
Saponins in Cancer Treatment: Current Progress and Future Prospects

 , ,
, ,
Abstract
1. Introduction
2. Classification of Saponins
2.1. Sources of Saponins
2.1.1. Synthetic Saponins
2.1.2. Natural Sources of Saponins
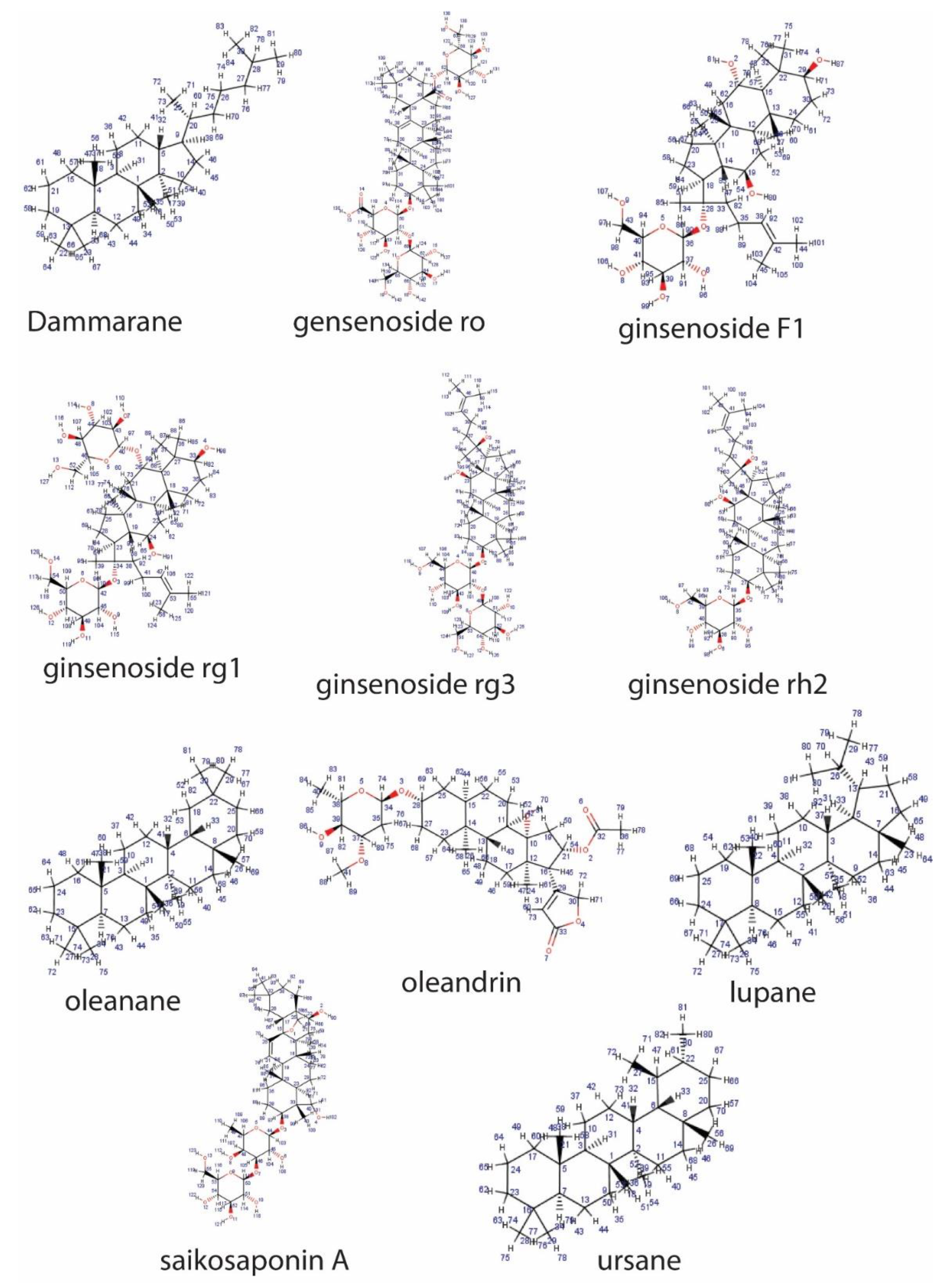
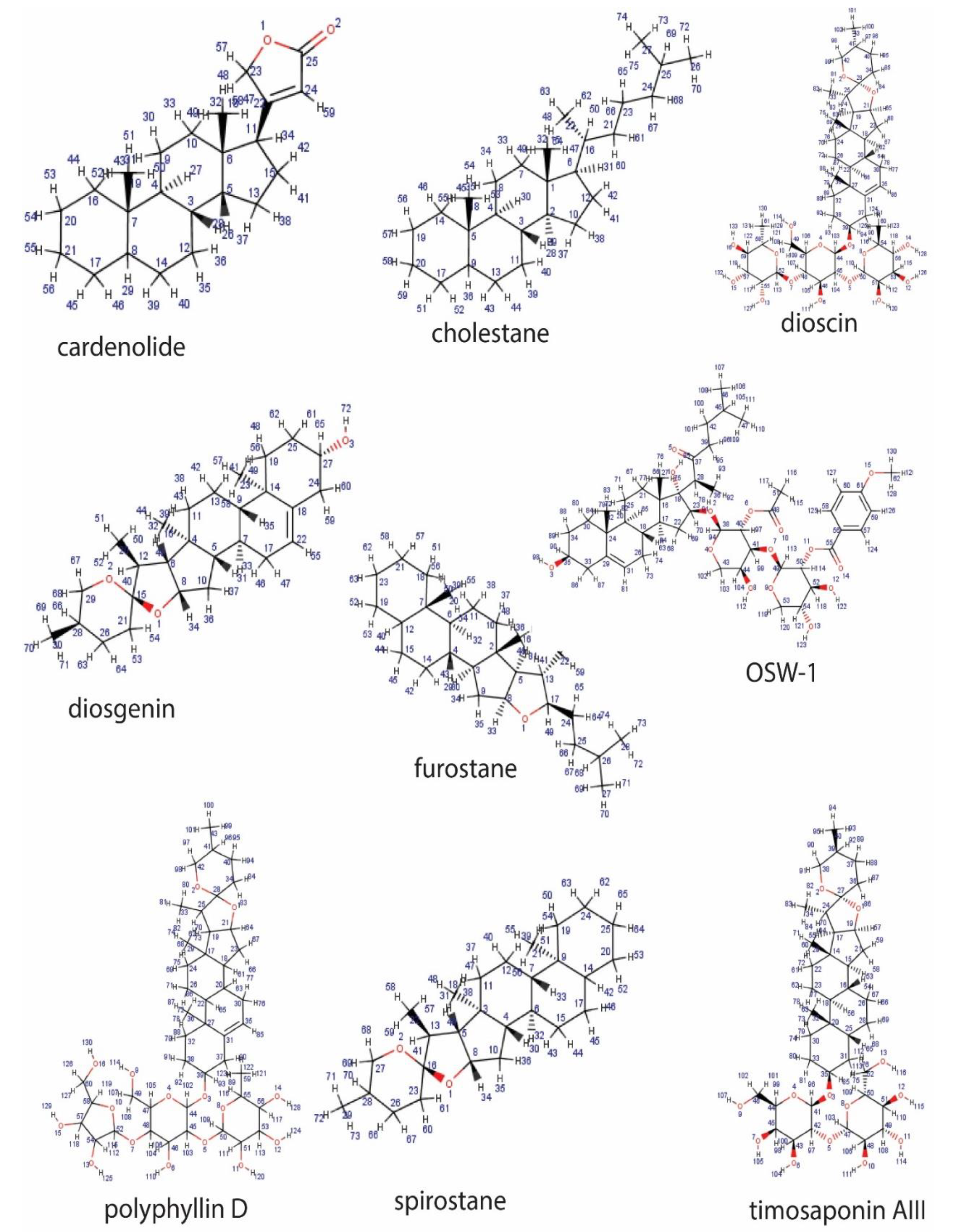
2.2. Classification Based on the Structure
3. Anticancer Mechanisms of Saponin
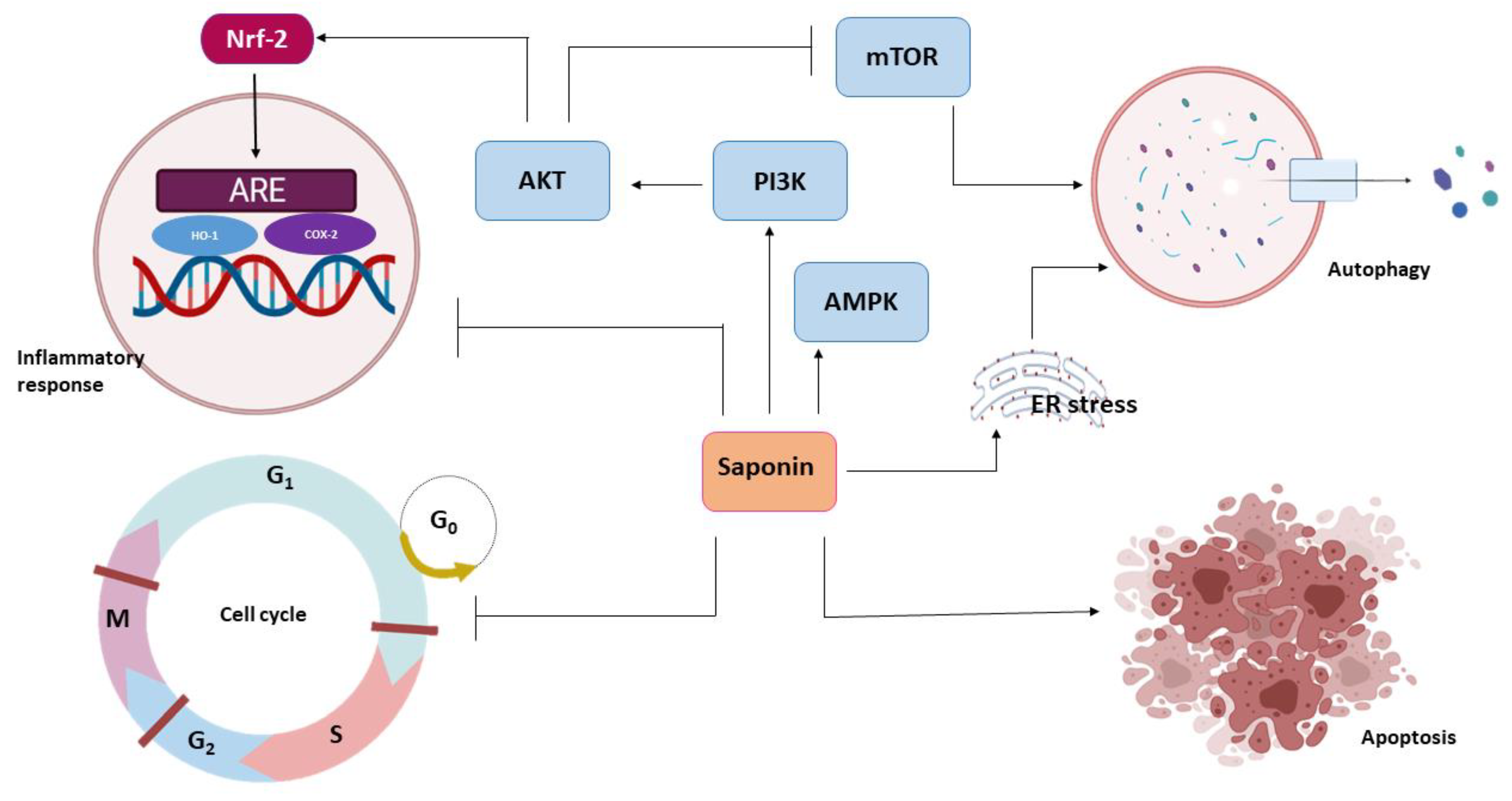
3.1. Chemoprevention and Saponin
3.1.1. Anti-Inflammatory Activity
3.1.2. Modulation of Redox Potential
3.1.3. Cell-Cycle Arrest
3.2. Cytotoxicity Effects of Saponins
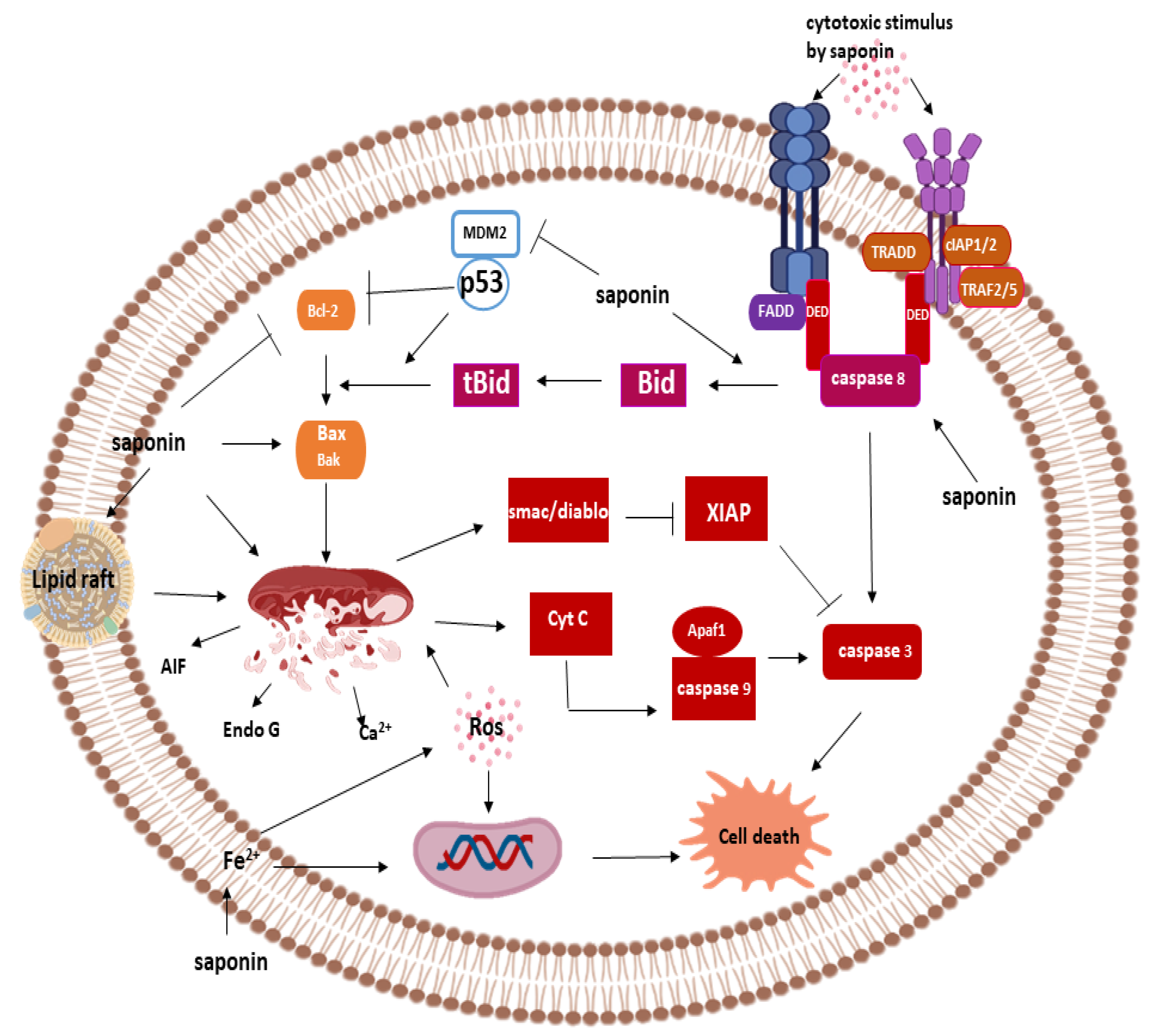
3.2.1. Apoptosis
Saponins and Caspase-Dependent Apoptosis
Extrinsic Pathway and Saponins
Intrinsic Pathway and Saponins
Saponin and Caspase Independent Apoptosis
3.2.2. Ferroptosis, Oncotic Necrosis and Necroptosis
3.2.3. Autophagy and Saponin
3.3. Metastasis and Saponins
3.4. Saponin in Multidrug Resistant Cancer
4. Limitations and Prospects
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Preventing Chronic Diseases: A Vital Investment; World Global Report; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef]
- Kassem, M.E.S.; Shoela, S.; Marzouk, M.M.; Sleem, A.A. A sulphated flavone glycoside from Livistona australis and its antioxidant and cytotoxic activity. Nat. Prod. Res. 2012, 26, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.; Kaur, P.; Shukla, S.; Abbas, A.; Fu, P.; Gupta, S. Plant flavone apigenin inhibits HDAC and remodels chromatin to induce growth arrest and apoptosis in human prostate cancer cells: In vitro and in vivo study. Mol. Carcinog. 2012, 51, 952–962. [Google Scholar] [CrossRef]
- Ameh, S.J.; Tarfa, F.D.; Ebeshi, B.U. Traditional herbal management of sickle cell anemia: Lessons from Nigeria. Anemia 2012, 2012, 1–9. [Google Scholar] [CrossRef]
- Cheng, G.; Gao, F.; Sun, X.; Bi, H.; Zhu, Y. Paris saponin VII suppresses osteosarcoma cell migration and invasion by inhibiting MMP-2/9 production via the p38 MAPK signaling pathway. Mol. Med. Rep. 2016, 14, 3199–3205. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Shen, X.; Li, S.; Li, Y.; Si, H.; Fan, J.; Li, J.; Gao, E.; Liu, S. Structure-activity relationships of 3-O-β-chacotriosyl oleanane-type triterpenoids as potential H5N1 entry inhibitors. Eur. J. Med. Chem. 2016, 119, 109–121. [Google Scholar] [CrossRef]
- Juang, Y.-P.; Liang, P.-H. Biological and pharmacological effects of synthetic saponins. Molecules 2020, 25, 4974. [Google Scholar] [CrossRef]
- Gauthier, C.; Legault, J.; Pichette, A. Recent progress in the synthesis of naturally occurring triterpenoid saponins. Mini Rev. Org. Chem. 2009, 6, 321–344. [Google Scholar] [CrossRef]
- Yang, Y.; Laval, S.; Yu, B. Chemical synthesis of saponins. In Horton DBT-A in CC and B; Academic Press: Cambridge, MA, USA, 2014; pp. 137–226. [Google Scholar]
- Liu, H.; Chou, G.-X.; Wu, T.; Guo, Y.-L.; Wang, S.-C.; Wang, C.-H.; Wang, Z.-T. Steroidal sapogenins and glycosides from the rhizomes of dioscorea bulbifera. J. Nat. Prod. 2009, 72, 1964–1968. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, A.E. Saponins in cereals. Phytochemistry 2003, 62, 1–4. [Google Scholar] [CrossRef]
- Zhang, X.-M.; Li, X.-B.; Zhang, S.-S.; He, Q.-X.; Hou, H.-R.; Dang, L.; Guo, J.-L.; Chen, Y.-F.; Yu, T.; Peng, D.-J.; et al. LC-MS/MS identification of novel saponins from the viscera of sea cucumber apostichopus japonicus. Chem. Nat. Compd. 2018, 54, 721–725. [Google Scholar] [CrossRef]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef]
- Xu, X.-H.; Li, T.; Fong, C.M.V.; Chen, X.; Chen, X.-J.; Wang, Y.-T.; Huang, M.-Q.; Lu, J.-J. Saponins from Chinese medicines as anticancer agents. Molecules 2016, 21, 1326. [Google Scholar] [CrossRef]
- Gevrenova, R.; Weng, A.; Voutguenne-Nazabadioko, L.; Thakur, M.; Doytchinova, I. Quantitative structure–activity relationship study on saponins as cytotoxicity enhancers. Lett. Drug Des. Discov. 2015, 12, 166–171. [Google Scholar] [CrossRef]
- Xu, K.; Shu, Z.; Xu, Q.-M.; Liu, Y.-L.; Li, X.-R.; Wang, Y.-L.; Yang, S.-L. Cytotoxic activity of Pulsatilla chinensis saponins and their structure–activity relationship. J. Asian Nat. Prod. Res. 2013, 15, 680–686. [Google Scholar] [CrossRef]
- Nag, S.A.; Qin, J.; Wang, W.; Wang, M.-H.; Wang, H.; Zhang, R. Ginsenosides as anticancer agents: In vitro and in vivo activities, structure-activity relationships, and molecular mechanisms of action. Front. Pharmacol. 2012, 3, 25. [Google Scholar] [CrossRef]
- Liu, M.-J.; Wang, Z.; Ju, Y.; Wong, R.N.-S.; Wu, Q.-Y. Diosgenin induces cell cycle arrest and apoptosis in human leukemia K562 cells with the disruption of Ca2+ homeostasis. Cancer Chemother. Pharmacol. 2005, 55, 79–90. [Google Scholar] [CrossRef]
- Bhuvanalakshmi, G.; Rangappa, K.S.; Dharmarajan, A.; Sethi, G.; Kumar, A.P.; Warrier, S. Breast cancer stem-like cells are inhibited by diosgenin, a steroidal saponin, by the attenuation of the Wnt β-catenin signaling via the Wnt antagonist secreted frizzled related protein-4. Front. Pharmacol. 2017, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Zheng, L.; Xu, L.; Yin, L.; Han, X.; Qi, Y.; Xu, Y.; Wang, C.; Peng, J. Dioscin suppresses human laryngeal cancer cells growth via induction of cell-cycle arrest and MAPK-mediated mitochondrial-derived apoptosis and inhibition of tumor invasion. Eur. J. Pharmacol. 2016, 774, 105–117. [Google Scholar] [CrossRef]
- Zhao, X.; Tao, X.; Xu, L.; Yin, L.; Qi, Y.; Xu, Y.; Han, X.; Peng, J. Dioscin induces apoptosis in human cervical carcinoma hela and siha cells through ROS-mediated DNA damage and the mitochondrial signaling pathway. Molecules 2016, 21, 730. [Google Scholar] [CrossRef]
- Kim, E.-A.; Jang, J.-H.; Lee, Y.-H.; Sung, E.-G.; Song, I.-H.; Kim, J.-Y.; Kim, S.; Sohn, H.-Y.; Lee, T.-J. Dioscin induces caspase-independent apoptosis through activation of apoptosis-inducing factor in breast cancer cells. Apoptosis 2014, 19, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Suzuki, T.; Hara, F.; Yasui, T.; Uga, N.; Naoe, A. Polyphyllin D, a steroidal saponin in Paris polyphylla, induces apoptosis and necroptosis cell death of neuroblastoma cells. Pediatric Surg. Int. 2017, 33, 713–719. [Google Scholar] [CrossRef]
- Yang, C.; Cai, H.; Meng, X. Polyphyllin D induces apoptosis and differentiation in K562 human leukemia cells. Int. Immunopharmacol. 2016, 36, 17–22. [Google Scholar] [CrossRef]
- Gao, M.; Cheung, K.L.; Lau, I.P.; Yu, W.S.; Fung, K.P.; Yu, B.; Loo, J.F.; Kong, S.K. Polyphyllin D induces apoptosis in human erythrocytes through Ca 2+ rise and membrane permeabilisation. Arch. Toxicol. 2012, 86, 741–752. [Google Scholar] [CrossRef]
- Lee, M.-S.; Chan, J.Y.-W.; Kong, S.-K.; Yu, B.; Eng-Choon, V.O.; Nai-Ching, H.W.; Chung-Wai, T.M.; Fung, K.-P.; Yuet-Wa, J.C. Effects of polyphyllin D, a steroidal saponin inParis Polyphylla, in growth inhibition of human breast cancer cells and in xenograft. Cancer Biol. Ther. 2005, 4, 1248–1254. [Google Scholar] [CrossRef]
- Kanwal, N.; Rasul, A.; Hussain, G.; Anwar, H.; Shah, M.A.; Sarfraz, I.; Riaz, A.; Batool, R.; Shahbaz, M.; Hussain, A.; et al. Oleandrin: A bioactive phytochemical and potential cancer killer via multiple cellular signaling pathways. Food Chem. Toxicol. 2020, 143, 111570. [Google Scholar] [CrossRef]
- Yuan, B.; Yang, R.; Ma, Y.; Zhou, S.; Zhang, X.; Liu, Y. A systematic review of the active saikosaponins and extracts isolated from Radix Bupleuri and their applications. Pharm. Biol. 2017, 55, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.-F.; Wang, X.; Kang, H.-F.; Bai, M.-H.; Guan, H.-T.; Wang, Z.-W.; Zan, Y.; Song, L.-Q.; Min, W.-L.; Lin, S. Saikosaponin-D enhances radiosensitivity of hepatoma cells under hypoxic conditions by inhibiting hypoxia-inducible factor-1α. Cell Physiol Biochem. 2014, 33, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Siu, F.-M.; Ma, D.-L.; Cheung, Y.-W.; Lok, C.-N.; Yan, K.; Yang, Z.; Yang, M.; Xu, S.; Ko, B.C.-B.; He, Q.-Y.; et al. Proteomic and transcriptomic study on the action of a cytotoxic saponin (Polyphyllin D): Induction of endoplasmic reticulum stress and mitochondria-mediated apoptotic pathways. Proteomics 2008, 8, 3105–3117. [Google Scholar] [CrossRef]
- Pan, H.-J.; Nie, X.-Q.; Liu, D.; Bian, K. Effects of four kinds of Chinese medicine monomer on growth of PANC-1 xenograft tumor and studying of molecular mechanism. China J. Chin. Mater. Med. 2013, 38, 245–248. [Google Scholar]
- Nho, K.J.; Chun, J.M.; Kim, H.K. Induction of mitochondria-dependent apoptosis in HepG2 human hepatocellular carcinoma cells by timosaponin A-III. Environ. Toxicol. Pharmacol. 2016, 45, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-H.; Yang, C.-W.; Wang, J.-Y.; Tsai, Y.-F.; Tseng, L.-M.; King, K.-L.; Chen, W.-S.; Chiu, J.-H.; Shyr, Y.-M. Timosaponin AIII suppresses hepatocyte growth factor-induced invasive activity through sustained ERK activation in breast cancer MDA-MB-231 Cells. Evid. Based Complement. Altern. Med. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, F.; Fan, K.; Zhang, Y.; Zhang, J.; Guo, H.; Yu, P.; Ma, J. Effective cytotoxic activity of OSW-1 on colon cancer by inducing apoptosis in vitro and in vivo. Oncol. Rep. 2017, 37, 3509–3519. [Google Scholar] [CrossRef]
- Malabed, R.; Hanashima, S.; Murata, M.; Sakurai, K. Interactions of OSW-1 with lipid bilayers in comparison with digitonin and soyasaponin. Langmuir 2020, 36, 3600–3610. [Google Scholar] [CrossRef]
- Kimura, M.; Sasaki, K.; Fukutani, Y.; Yoshida, H.; Ohsawa, I.; Yohda, M.; Sakurai, K. Anticancer saponin OSW-1 is a novel class of selective Golgi stress inducer. Bioorg. Med. Chem. Lett. 2019, 29, 1732–1736. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, C.; Zhou, C.; Zhuang, J.; Tang, S.; Yu, J.; Tian, J.; Feng, F.; Liu, L.; Zhang, T.; et al. Meta-analysis of the association between the dietary inflammatory index (DII) and breast cancer risk. Eur. J. Clin. Nutr. 2018, 73, 509–517. [Google Scholar] [CrossRef]
- Chen, M.; Ye, K.; Zhang, B.; Xin, Q.; Li, P.; Kong, A.-N.; Wen, X.; Yang, J. Paris Saponin II inhibits colorectal carcinogenesis by regulating mitochondrial fission and NF-κB pathway. Pharmacol. Res. 2019, 139, 273–285. [Google Scholar] [CrossRef]
- Xia, L.; Zhang, B.; Yan, Q.; Ruan, S. Effects of saponins of patrinia villosa against invasion and metastasis in colorectal cancer cell through NF-κB signaling pathway and EMT. Biochem. Biophys. Res. Commun. 2018, 503, 2152–2159. [Google Scholar] [CrossRef]
- Baharara, J.; Amini, E.; Salek-Abdollahi, F. Anti-inflammatory properties of saponin fraction from Ophiocoma erinaceus. Iran. J. Fish. Sci. 2020, 19, 638–652. [Google Scholar]
- Ma, L.; Liu, H.; Qin, P.; Hu, C.; Man, S.; Li, Y.; Liu, Z.; Liu, Z.; Diao, A. Saponin fraction isolated from Conyza blinii H. Lév. demonstrates strong anticancer activity that is due to its NF-κB inhibition. Biochem. Biophys. Res. Commun. 2017, 483, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Deng, S.; Zhou, D.; Huang, Y.; Li, C.; Hao, L.; Zhang, G.; Su, S.; Xu, X.; Yang, R. 3, 4-Seco-dammarane triterpenoid saponins with anti-inflammatory activity isolated from the leaves of Cyclocarya paliurus. J. Agric. Food Chem. 2020, 68, 2041–2053. [Google Scholar] [CrossRef] [PubMed]
- Morry, J.; Ngamcherdtrakul, W.; Yantasee, W. Oxidative stress in cancer and fibrosis: Opportunity for therapeutic intervention with antioxidant compounds, enzymes, and nanoparticles. Redox. Biol. 2017, 11, 240–253. [Google Scholar] [CrossRef]
- Alwash, B.M.J. Triterpenoid saponins investigation and pharmacological (cytotoxic and antioxidant) properties of Bacopa monnieri L. cultivated in Iraq. Baghdad Sci. J. 2018, 15, 123–129. [Google Scholar]
- Choudhry, Q.N.; Kim, M.J.; Kim, T.G.; Pan, J.H.; Kim, J.H.; Park, S.J.; Lee, J.H.; Kim, Y.J. Saponin-based nanoemulsification improves the antioxidant properties of vitamin A and E in AML-12 Cells. Int. J. Mol. Sci. 2016, 17, 1406. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wu, Y.; Zhao, B.; Hu, H.; Zhu, B.; Sun, Z.; Li, P.; Du, S. Panax notoginseng saponins protect cerebral microvascular endothelial cells against oxygen-glucose deprivation/reperfusion-induced barrier dysfunction via activation of PI3K/Akt/Nrf2 antioxidant signaling pathway. Molecules 2018, 23, 2781. [Google Scholar] [CrossRef]
- Sznarkowska, A.; Kostecka, A.; Meller, K.; Bielawski, K.P. Inhibition of cancer antioxidant defense by natural compounds. Oncotarget 2017, 8, 15996. [Google Scholar] [CrossRef]
- Mu, L.-H.; Wang, L.-H.; Yu, T.-F.; Wang, Y.-N.; Yan, H.; Liu, P.; Yan, C. Triterpenoid Saponin AG8 from Ardisia gigantifolia stapf. induces triple negative breast cancer cells apoptosis through oxidative stress pathway. Oxid. Med. Cell. Longev. 2020, 2020, 7963212. [Google Scholar] [CrossRef]
- Kim, E.H.; Baek, S.; Shin, D.; Lee, J.; Roh, J.-L. Hederagenin induces apoptosis in cisplatin-resistant head and neck cancer cells by inhibiting the Nrf2-ARE antioxidant pathway. Oxidative Med. Cell. Longev. 2017, 2017, 5498908. [Google Scholar] [CrossRef]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, G. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348. [Google Scholar] [PubMed]
- Jaramillo, S.; Muriana, F.J.; Guillen, R.; Jimenez-Araujo, A.; Arcos, R.R.; Lopez, S. Saponins from edible spears of wild asparagus inhibit AKT, p70S6K, and ERK signalling, and induce apoptosis through G0/G1 cell cycle arrest in human colon cancer HCT-116 cells. J. Funct. Foods 2016, 26, 1–10. [Google Scholar] [CrossRef]
- Yan, T.; Hu, G.; Wang, A.; Sun, X.; Yu, X.; Jia, J. Paris saponin VII induces cell cycle arrest and apoptosis by regulating Akt/MAPK pathway and inhibition of P-glycoprotein in K562/ADR cells. Phytother. Res. 2018, 32, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, Q.-S.; Meng, F.-C.; Tang, Z.-H.; Chen, X.; Lin, L.-G.; Chen, P.; Qiang, W.-A.; Wang, Y.-T.; Zhang, Q.-W. Chikusetsusaponin IVa methyl ester induces G1 cell cycle arrest, triggers apoptosis and inhibits migration and invasion in ovarian cancer cells. Phytomedicine 2016, 23, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, N.; Rankin, G.O.; Li, B.; Rojanasakul, Y.; Tu, Y.; Chen, Y.C. Anti-proliferative effect and cell cycle arrest induced by saponins extracted from tea (Camellia sinensis) flower in human ovarian cancer cells. J. Funct. Foods 2017, 37, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Du, L.; Jiang, H.; Zhu, X.; Li, J.; Xu, J. Paris saponin I sensitises gastric cancer cell lines to cisplatin via cell cycle arrest and apoptosis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 3798. [Google Scholar]
- Yao, M.; Li, R.; Yang, Z.; Ding, Y.; Zhang, W.; Li, W.; Liu, M.; Zhao, C.; Wang, Y.; Tang, H. PP9, a steroidal saponin, induces G2/M arrest and apoptosis in human colorectal cancer cells by inhibiting the PI3K/Akt/GSK3β pathway. Chem. Biol. Interact. 2020, 331, 109246. [Google Scholar] [CrossRef]
- Deng, L.-J.; Hu, L.-P.; Peng, Q.-L.; Yang, X.-L.; Bai, L.-L.; Yiu, A.; Li, Y.; Tian, H.-Y.; Ye, W.-C.; Zhang, D.-M. Hellebrigenin induces cell cycle arrest and apoptosis in human hepatocellular carcinoma HepG2 cells through inhibition of Akt. Chem. Biol. Interact. 2014, 219, 184–194. [Google Scholar] [CrossRef]
- Liao, W.-L.; Lin, J.-Y.; Shieh, J.-C.; Yeh, H.-F.; Hsieh, Y.-H.; Cheng, Y.-C.; Lee, H.-J.; Shen, C.-Y.; Cheng, C.-W. Induction of G2/M phase arrest by diosgenin via activation of chk1 kinase and cdc25c regulatory pathways to promote apoptosis in human breast cancer cells. Int. J. Mol. Sci. 2019, 21, 172. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz-Hajduk, J.; Bartoszewski, R.; Bartoszewska, S.; Kochan, K.; Adamska, A.; Kosiński, I.; Ochocka, J.R. Pennogenyl Saponins from Paris quadrifolia L. Induce Extrinsic and Intrinsic Pathway of Apoptosis in Human Cervical Cancer HeLa Cells. PLoS ONE 2015, 10, e0135993. [Google Scholar] [CrossRef]
- Qin, H.; Du, X.; Zhang, Y.; Wang, R. Platycodin D, a triterpenoid saponin from Platycodon grandiflorum, induces G2/M arrest and apoptosis in human hepatoma HepG2 cells by modulating the PI3K/Akt pathway. Tumor. Biol. 2014, 35, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.K.-S.; Auyeung, K.K.-W.; Mok, N.-L.; Wong, C.-M.; Cho, C.-H. Astragalus saponins modulate mTOR and ERK signaling to promote apoptosis through the extrinsic pathway in HT-29 colon cancer cells. Int. J. Mol. Med. 2010, 26, 341–349. [Google Scholar] [CrossRef]
- Cho, S.-H.; Chung, K.-S.; Choi, J.-H.; Kim, N.-H.; Lee, K.-T. Compound K, a metabolite of ginseng saponin, induces apoptosis via caspase-8-dependent pathway in HL-60 human leukemia cells. BMC Cancer 2009, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Sayers, T.J. Targeting the extrinsic apoptosis signaling pathway for cancer therapy. Cancer Immunol. Immunother. 2011, 60, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A. Targeting the extrinsic apoptosis pathway in cancer. Cytokine Growth Factor Rev. 2008, 19, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Kang, K.A.; Lim, C.M.; Kim, K.C.; Kim, H.S.; Kim, D.H.; Kim, B.J.; Chang, W.Y.; Choi, J.H.; Hyun, J.W. Compound K, a metabolite of ginseng saponin, induces mitochondria-dependent and caspase-dependent apoptosis via the generation of reactive oxygen species in human colon cancer cells. Int. J. Mol. Sci. 2010, 11, 4916–4931. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Bai, P.; Nguyen, T.M.B.; Xiao, J.; Liu, S.; Yang, G.; Hu, L.; Chen, X.; Zhang, X.; Liu, J.; et al. The antitumoral effect of Paris Saponin I associated with the induction of apoptosis through the mitochondrial pathway. Mol. Cancer Ther. 2009, 8, 1179–1188. [Google Scholar] [CrossRef]
- Lv, L.; Zheng, L.; Dong, D.; Xu, L.; Yin, L.; Xu, Y.; Qi, Y.; Han, X.; Peng, J. Dioscin, a natural steroid saponin, induces apoptosis and DNA damage through reactive oxygen species: A potential new drug for treatment of glioblastoma multiforme. Food Chem. Toxicol. 2013, 59, 657–669. [Google Scholar] [CrossRef]
- Wei, G.; Sun, J.; Hou, Z.; Luan, W.; Wang, S.; Cui, S.; Cheng, M.; Liu, Y. Novel antitumor compound optimised from natural saponin Albiziabioside A induced caspase-dependent apoptosis and ferroptosis as a p53 activator through the mitochondrial pathway. Eur. J. Med. Chem. 2018, 157, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Lawen, A. Apoptosis—An introduction. Bioessays 2003, 25, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Martin, S. Caspase-independent cell death. Nat. Med. 2005, 11, 725–730. [Google Scholar] [CrossRef]
- Lorent, J.H.; Léonard, C.; Abouzi, M.; Akabi, F.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.-P. α-Hederin induces apoptosis, membrane permeabilisation and morphologic changes in two cancer cell lines through a cholesterol-dependent mechanism. Planta. Med. 2016, 82, 1532–1539. [Google Scholar]
- Verstraeten, S.L.; Albert, M.; Paquot, A.; Muccioli, G.G.; Tyteca, D.; Mingeot-Leclercq, M.-P. Membrane cholesterol delays cellular apoptosis induced by ginsenoside Rh2, a steroid saponin. Toxicol. Appl. Pharmacol. 2018, 352, 59–67. [Google Scholar] [CrossRef]
- Yun, U.-J.; Lee, J.-H.; Koo, K.H.; Ye, S.-K.; Kim, S.-Y.; Lee, C.-H.; Kim, Y.-N. Lipid raft modulation by Rp1 reverses multidrug re-sistance via inactivating MDR-1 and Src inhibition. Biochem. Pharmacol. 2013, 85, 1441–1453. [Google Scholar] [CrossRef] [PubMed]
- Holze, C.; Michaudel, C.; Mackowiak, C.; Haas, D.A.; Benda, C.; Hubel, P.; Pennemann, F.L.; Schnepf, D.; Wettmarshausen, J.; Braun, M.; et al. Oxeiptosis, a ROS-induced caspase-independent apoptosis-like cell-death pathway. Nat. Immunol. 2018, 19, 130–140. [Google Scholar] [CrossRef]
- Hong, J.-M.; Kim, J.-H.; Kim, H.; Lee, W.J.; Hwang, Y.-I. SB365, Pulsatilla saponin d induces caspase-independent cell death and augments the anticancer effect of temozolomide in glioblastoma multiforme cells. Molecules 2019, 24, 3230. [Google Scholar] [CrossRef]
- Saelens, X.; Festjens, N.; Walle LVande Van Gurp, M.; Van Loo, G.; Vandenabeele, P. Toxic proteins released from mito-chondria in cell death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Yang, J.-S.; Lu, K.-W.; Yu, C.-S.; Chou, S.-T.; Lin, J.-J.; Chen, Y.-Y.; Lin, M.-L.; Chueh, F.-S.; Chen, S.-S.; et al. An experimental study on the antileukemia effects of gypenosides in vitro and in vivo. Integr. Cancer Ther. 2010, 10, 101–112. [Google Scholar] [CrossRef]
- Li, L.Y.; Luo, X.; Wang, X. Endonuclease G is an apoptotic DNase when released from mitochondria. Nat. Cell Biol. 2001, 412, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Liu, Y.; Dai, R.; Ismail, N.; Su, W.; Li, B. Ferroptosis and its potential role in human diseases. Front. Pharmacol. 2020, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Mbaveng, A.T.; Ndontsa, B.L.; Kuete, V.; Nguekeu, Y.M.; Çelik, I.; Mbouangouere, R.; Tane, P.; Efferth, T. A naturally occuring triterpene saponin ardisiacrispin B displayed cytotoxic effects in multi-factorial drug resistant cancer cells via ferroptotic and apoptotic cell death. Phytomedicine 2018, 43, 78–85. [Google Scholar] [CrossRef]
- Su, C.-L.; Lin, P.-L. Natural Saponin Formosanin C-induced Ferroptosis in Human Hepatocellular Carcinoma Cells In-volved Ferritinophagy. FASEB J. 2020, 34, 1. [Google Scholar]
- Song, Z.; Xiang, X.; Li, J.; Deng, J.; Fang, Z.; Zhang, L.; Xiong, J. Ruscogenin induces ferroptosis in pancreatic cancer cells. Oncol. Rep. 2019, 43, 516–524. [Google Scholar] [CrossRef]
- Gao, J.; Li, X.; Gu, G.; Sun, B.; Cui, M.; Ji, M.; Lou, H.-X. Efficient synthesis of trisaccharide saponins and their tumor cell killing effects through oncotic necrosis. Bioorg. Med. Chem. Lett. 2011, 21, 622–627. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Chi, G.F.; Nguenang, G.S.; Abdelfatah, S.; Sop, R.V.T.; Ngadjui, B.T.; Kuete, V.; Efferth, T. Cytotoxicity of a naturally occuring spirostanol saponin, progenin III, towards a broad range of cancer cell lines by induction of apoptosis, autophagy and necroptosis. Chem. Interact. 2020, 326, 109141. [Google Scholar] [CrossRef] [PubMed]
- Linkermann, A.; Green, D.R. Necroptosis. N. Engl. J. Med. 2014, 370, 455–465. [Google Scholar] [CrossRef]
- White, E. The role for autophagy in cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting autophagy in cancer. Nat. Rev. Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef]
- Mathew, R.; Karantza-Wadsworth, V.; White, E. Role of autophagy in cancer. Nat. Rev. Cancer 2007, 7, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bao, J.; Wang, K.; Jia, X.; Zhang, C.; Huang, B.; Chen, M.; Wan, J.-B.; Su, H.; Wang, Y. Pulsatilla saponin D inhibits au-tophagic flux and synergistically enhances the anticancer activity of chemotherapeutic agents against HeLa cells. Am. J. Chin. Med. 2015, 43, 1657–1670. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. An overview of the molecular mechanism of autophagy. Curr. Top. Microbiol. Immunol. 2009, 335, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J. Autophagy: From phenomenology to molecular understanding in less than a decade. Nat. Rev. Mol. Cell Biol. 2007, 8, 931–937. [Google Scholar] [CrossRef]
- Xie, Z.-Z.; Li, M.-M.; Deng, P.-F.; Wang, S.; Wang, L.; Lu, X.-P.; Hu, L.-B.; Chen, Z.; Jie, H.-Y.; Wang, Y.-F. Paris saponin-induced au-tophagy promotes breast cancer cell apoptosis via the Akt/mTOR signaling pathway. Chem. Biol. Interact. 2017, 264, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Zhou, Y.; Zhang, J.; Xiang, Z.; Liu, Y.; Miao, T.; Liu, G.; Liu, B.; Liu, X.; Shen, L.; et al. Anemoside B4 exerts anti-cancer effect by inducing apoptosis and autophagy through inhibiton of PI3K/Akt/mTOR pathway in hepatocellular carcinoma. Am. J. Transl. Res. 2019, 11, 2580–2589. [Google Scholar]
- Steeves, M.A.; Dorsey, F.C.; Cleveland, J.L. Targeting the autophagy pathway for cancer chemoprevention. Curr. Opin. Cell Biol. 2010, 22, 218–225. [Google Scholar] [CrossRef]
- Shinojima, N.; Yokoyama, T.; Kondo, Y.; Kondo, S. Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in curcumin-induced autophagy. Autophagy 2007, 3, 635–637. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, C.; Chen, L.; Chen, Y.C.; Tu, Y. Saponins extracted from tea (Camellia Sinensis) flowers induces autophagy in ovarian cancer cells. Molecules 2020, 25, 5254. [Google Scholar] [CrossRef]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Tong, S.; Wu, J.; Tian, L.; Qi, Z.; Chen, B.; Zhu, D.; Zhang, Y. Paris saponin VII extracted from Trillium tschonoskii induces autophagy and apoptosis in NSCLC cells. J. Ethnopharmacol. 2020, 248, 112304. [Google Scholar] [CrossRef]
- Zhang, C.; Jia, X.; Wang, K.; Bao, J.; Li, P.; Chen, M.; Wan, J.-B.; Su, H.; Mei, Z.; He, C. Polyphyllin VII induces an autophagic cell death by activation of the JNK pathway and inhibition of PI3K/AKT/mTOR pathway in HepG2 cells. PLoS ONE 2016, 11, e0147405. [Google Scholar] [CrossRef]
- Wu, Q.; Deng, J.; Fan, D.; Duan, Z.; Zhu, C.; Fu, R.; Wang, S. Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem. Pharmacol. 2018, 148, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Amaravadi, R.; Kimmelman, A.C.; White, E. Recent insights into the function of autophagy in cancer. Genes Dev. 2016, 30, 1913–1930. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hu, C.; Sun, N.; Li, Y.; Man, S.; Liu, Z.; Diao, A.; Ma, L. A triterpenoidal saponin fraction of Conyza blinii H.Lév. is a dual-targeting autophagy inhibitor for HeLa cells. RSC Adv. 2017, 7, 24291–24297. [Google Scholar] [CrossRef]
- Souza ASC de Gonçalves, L.B.; Lepique, A.P.; Araujo-Souza, D.; Savio, P. The role of autophagy in tumor immunolo-gy-complex mechanisms that may be explored therapeutically. Front. Oncol. 2020, 10, 2691. [Google Scholar]
- Koczurkiewicz, P.; Czyż, J.; Podolak, I.; Wójcik, K.; Galanty, A.; Janeczko, Z.; Michalik, M. Multidirectional effects of triterpene saponins on cancer cells—Mini-review of in vitro studies. Acta Biochim. Pol. 2015, 62, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Woodhouse, E.C.; Chuaqui, R.F.; Liotta, L.A. General mechanisms of metastasis. Cancer Interdiscip. Int. J. Am. Cancer Soc. 1997, 80, 1529–1537. [Google Scholar] [CrossRef]
- Mochizuki, M.; YOO, Y.; Matsuzawa, K.; Sato, K.; Saiki, I.; Tonooka, S.; Samukawa, K.; Azuma, I. Inhibitory effect of tumor metastasis in mice by saponins, ginsenoside-Rb2, 20(R)- and 20(S)-ginsenoside-Rg3, of red ginseng. Biol. Pharm. Bull. 1995, 18, 1197–1202. [Google Scholar] [CrossRef]
- Sato, K.; Mochizuki, M.; Saiki, I.; YOO, Y.; Samukawa, K.; Azuma, I. Inhibition of tumor angiogenesis and metastasis by a saponin of panax ginseng, ginsenoside-Rb. Biol. Pharm. Bull. 1994, 17, 635–639. [Google Scholar] [CrossRef]
- Chan, J.Y.-W.; Koon, J.C.-M.; Liu, X.; Detmar, M.; Yu, B.; Kong, S.-K.; Fung, K.P. Polyphyllin D, a steroidal saponin from Paris polyphylla, inhibits endothelial cell functions in vitro and angiogenesis in zebrafish embryos in vivo. J. Ethnopharmacol. 2011, 137, 64–69. [Google Scholar] [CrossRef]
- Yang, B.-R.; Cheung, K.K.; Zhou, X.; Xie, R.-F.; Cheng, P.-P.; Wu, S.; Zhou, Z.-Y.; Tang, J.Y.; Hoi, P.M.; Wang, Y.-H.; et al. Amelioration of acute myocardial infarction by saponins from flower buds of Panax notoginseng via pro-angiogenesis and anti-apoptosis. J. Ethnopharmacol. 2016, 181, 50–58. [Google Scholar] [CrossRef]
- Yang, M.; Zou, J.; Zhu, H.; Liu, S.; Wang, H.; Bai, P.; Xiao, X. Paris saponin II inhibits human ovarian cancer cell-induced an-giogenesis by modulating NF-κB signaling. Oncol. Rep. 2015, 33, 2190–2198. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.-Y.; Liu, H.-J.; Luan, X.; Xu, J.-R.; Lu, Q.; Liu, Y.-R.; Gao, Y.-G.; Zhao, M.; Chen, H.-Z.; Fang, C. Raddeanin A, a triterpenoid saponin isolated from Anemone raddeana, suppresses the angiogenesis and growth of human colorectal tumor by inhibiting VEGFR2 signaling. Phytomedicine 2015, 22, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Zhang, X.; Tong, Y.; Yi, Y.; Zhang, S.; Liping, L.; Sun, P.; Lin, L.; Ding, J. PE, a new sulfated saponin from sea cucumber, Exhibits anti-angiogenic and anti-tumor activities in vitro and in vivo. Cancer Biol. Ther. 2005, 4, 874–882. [Google Scholar] [CrossRef]
- Li, Y.; Gu, J.-F.; Zou, X.; Wu, J.; Zhang, M.-H.; Jiang, J.; Qin, D.; Zhou, J.-Y.; Liu, B.-X.-Z.; Zhu, Y.-T.; et al. The anti-lung cancer activities of steroidal saponins of P. polyphylla Smith var. chinensis (Franch.) Hara through enhanced immunostimulation in experimental lewis tumor-bearing C57BL/6 mice and induction of apoptosis in the A549 cell line. Molecules 2013, 18, 12916–12936. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wan, J.-B.; Chan, S.-W.; Deng, Y.-H.; Yu, N.; Zhang, Q.-W.; Wang, Y.-T.; Lee, S.-M. Comparative study on saponin fractions from Panax notoginseng inhibiting inflammation-induced endothelial adhesion molecule expression and monocyte adhesion. Chin. Med. 2011, 6, 37. [Google Scholar] [CrossRef]
- Sun, L.; Lin, S.; Zhao, R.; Yu, B.; Yuan, S.; Zhang, L. The saponin monomer of dwarf lilyturf tuber, dt-13, reduces human breast cancer cell adhesion and migration during hypoxia via regulation of tissue factor. Biol. Pharm. Bull. 2010, 33, 1192–1198. [Google Scholar] [CrossRef][Green Version]
- Lou, L.; Ye, W.; Chen, Y.; Wu, S.; Jin, L.; He, J.; Tao, X.; Zhu, J.; Chen, X.; Deng, A.; et al. Ardipusilloside inhibits survival, invasion and metastasis of human hepatocellular carcinoma cells. Phytomedicine 2012, 19, 603–608. [Google Scholar] [CrossRef]
- Shuli, M.; Wenyuan, G.; Yanjun, Z.; Chaoyi, M.; Liu, Y.; Yiwen, L. Paridis saponins inhibiting carcinoma growth and metastasis In vitro and in vivo. Arch. Pharmacal Res. 2011, 34, 43–50. [Google Scholar] [CrossRef]
- Wang, P.; Cui, J.; Du, X.; Yang, Q.; Jia, C.; Xiong, M.; Yu, X.; Li, L.; Wang, W.; Chen, Y.; et al. Panax notoginseng saponins (PNS) inhibits breast cancer metastasis. J. Ethnopharmacol. 2014, 154, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Choi, Y.-J.; Cha, S.-W.; Lee, S.-G. Anti-metastatic effects of ginsenoside Rd via inactivation of MAPK signaling and induction of focal adhesion formation. Phytomedicine 2012, 19, 284–292. [Google Scholar] [CrossRef]
- Bauvois, B. New facets of matrix metalloproteinases MMP-2 and MMP-9 as cell surface transducers: Outside-in signaling and relationship to tumor progression. Biochim. Biophys. Acta (BBA) Bioenerg. 2012, 1825, 29–36. [Google Scholar] [CrossRef]
- Man, S.; Gao, W.; Zhang, Y.; Yan, L.; Ma, C.; Liu, C.; Huang, L. Antitumor and antimetastatic activities of Rhizoma Paridis saponins. Steroids 2009, 74, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Hwang, Y.S.; Park, K.-K.; Park, H.-J.; Seo, J.Y.; Chung, W.-Y. Kalopanaxsaponin A inhibits PMA-induced invasion by reducing matrix metalloproteinase-9 via PI3K/Akt-and PKCδ-mediated signaling in MCF-7 human breast cancer cells. Car-cinogenesis 2009, 30, 1225–1233. [Google Scholar]
- Jung, S.-H.; Woo, M.-S.; Kim, S.-Y.; Kim, W.-K.; Hyun, J.-W.; Kim, E.-J.; Kim, D.-H.; Kim, H.-S. Ginseng saponin metabolite suppresses phorbol ester-induced matrix metalloproteinase-9 expression through inhibition of activator protein-1 and mitogen-activated protein kinase signaling pathways in human astroglioma cells. Int. J. Cancer 2006, 118, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Luan, X.; Zhao, J.; Li, W. Inhibitory effect of Trillium saponins on invasion and migration of HuH-7 cells by promoting human β defensin-2 expression. Chin. J. Cell Mol. Immunol. 2019, 35, 526–532. [Google Scholar]
- Kang, J.-H.; Han, I.-H.; Sung, M.-K.; Yoo, H.; Kim, Y.-G.; Kim, J.-S.; Kawada, T.; Yu, R. Soybean saponin inhibits tumor cell metastasis by modulating expressions of MMP-2, MMP-9 and TIMP-2. Cancer Lett. 2008, 261, 84–92. [Google Scholar] [CrossRef]
- Chen, P.-S.; Shih, Y.-W.; Huang, H.-C.; Cheng, H.-W. Diosgenin, a steroidal saponin, inhibits migration and invasion of human prostate cancer PC-3 cells by reducing matrix metalloproteinases expression. PLoS ONE 2011, 6, e20164. [Google Scholar] [CrossRef]
- Lu, K.V.; Jong, K.A.; Rajasekaran, A.K.; Cloughesy, T.F.; Mischel, P.S. Upregulation of tissue inhibitor of metalloproteinases (TIMP)-2 promotes matrix metalloproteinase (MMP)-2 activation and ce ll invasion in a human glioblastoma cell line. Lab. Investig. 2004, 84, 8–20. [Google Scholar] [CrossRef]
- Hu, C.-T.; Wu, J.-R.; Cheng, C.-C.; Wu, W.-S. The therapeutic targeting of HGF/c-met signaling in hepatocellular carcinoma: Alternative approaches. Cancers 2017, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.; Xin, G.; Wei, Z.; Li, S.; Xing, Z.; Ji, C.; Du, J.; Niu, H.; Huang, W. Dihydrodiosgenin inhibits endothelial cell-derived factor VIII and platelet-mediated hepatocellular carcinoma metastasis. Cancer Manag. Res. 2019, 11, 4871–4882. [Google Scholar] [CrossRef]
- Song, G.; Guo, S.; Wang, W.; Hu, C.; Mao, Y.; Zhang, B.; Zhang, H.; Hu, T. Intestinal Metabolite Compound K of Ginseng Saponin Potently Attenuates Metastatic Growth of Hepatocellular Carcinoma by Augmenting Apoptosis via a Bid-Mediated Mitochondrial Pathway. J. Agric. Food Chem. 2010, 58, 12753–12760. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Cheng, X.; Wink, M. Natural lignans from Arctium lappa modulate P-glycoprotein efflux function in multidrug resistant cancer cells. Phytomedicine 2015, 22, 301–307. [Google Scholar] [CrossRef]
- Wang, H.; Zhai, Z.; Li, N.; Jin, H.; Chen, J.; Yuan, S.; Wang, L.; Zhang, J.; Li, Y.; Yun, J.; et al. Steroidal saponin of Trillium tschonoskii. Reverses multidrug resistance of hepatocellular carcinoma. Phytomedicine 2013, 20, 985–991. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Fan, L.; Zhang, F.; Meng, J.; Han, J.; Guo, X.; Zhang, D.; Zhang, R.; Yue, Z.; et al. Paris saponin VII inhibits growth of colorectal cancer cells through Ras signaling pathway. Biochem. Pharmacol. 2014, 88, 150–157. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, Q.; Chen, W.; Wu, M.; Pan, G.; Yang, K.; Li, X.; Man, S.; Teng, Y.; Yu, P.; et al. Chemosensitizing effect of Paris Saponin I on Camptothecin and 10-hydroxycamptothecin in lung cancer cells via p38 MAPK, ERK, and Akt signaling pathways. Eur. J. Med. Chem. 2017, 125, 760–769. [Google Scholar] [CrossRef]
- Jiang, Y.-X.; Yang, S.-W.; Li, P.-A.; Luo, X.; Li, Z.-Y.; Hao, Y.-X.; Yu, P.-W. The promotion of the transformation of quiescent gastric cancer stem cells by IL-17 and the underlying mechanisms. Oncogene 2017, 36, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Chaudhuri, P.K. Structural characteristics, bioavailability and cardioprotective potential of saponins. Integr. Med. Res. 2018, 7, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Basu, S.; Yang, Z.; Deb, A.; Hu, M. Bioavailability challenges associated with development of saponins as therapeutic and chemopreventive agents. Curr. Drug Targets 2012, 13, 1885–1899. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Gao, S.; Wang, J.; Yin, T.; Teng, Y.; Wu, B.; You, M.; Jiang, Z.; Hu, M. Enhancement of oral bioavailability of 20(S)-ginsenoside Rh2 through improved understanding of its absorption and efflux mechanisms. Drug Metab. Dispos. 2011, 39, 1866–1872. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Han, L.; Duan, H.; Cui, Y.; Feng, Y.; Zhu, Y.; Chen, Z.; Yang, S. The derivatives of Pulsatilla saponin A, a bioactive compound from Pulsatilla chinensis: Their synthesis, cytotoxicity, haemolytic toxicity and mechanism of action. Eur. J. Med. Chem. 2017, 129, 325–336. [Google Scholar] [CrossRef]
- Chwalek, M.; Lalun, N.; Bobichon, H.; Plé, K.; Voutquenne-Nazabadioko, L. Structure–activity relationships of some hed-eragenin diglycosides: Haemolysis, cytotoxicity and apoptosis induction. Biochim. Biophys. Acta. (BBA) General Subj. 2006, 1760, 1418–1427. [Google Scholar] [CrossRef]
- Vo, N.N.Q.; Fukushima, E.O.; Muranaka, T. Structure and hemolytic activity relationships of triterpenoid saponins and sapogenins. J. Nat. Med. 2017, 71, 50–58. [Google Scholar] [CrossRef]
- Witthawaskul, P.; Panthong, A.; Kanjanapothi, D.; Taesothikul, T.; Lertprasertsuke, N. Acute and subacute toxicities of the saponin mixture isolated from Schefflera leucantha Viguier. J. Ethnopharmacol. 2003, 89, 115–121. [Google Scholar] [CrossRef]
- Liang, J.; Jin, W.; Li, H.; Liu, H.; Huang, Y.; Shan, X.; Li, C.; Shan, L.; Efferth, T. In vivo cardiotoxicity induced by sodium aescinate in Zebrafish larvae. Molecules 2016, 21, 190. [Google Scholar] [CrossRef]
- Oleszek, W.; Junkuszew, M.; Stochmal, A. Determination and toxicity of saponins from Amaranthus cruentus seeds. J. Agric. Food Chem. 1999, 47, 3685–3687. [Google Scholar] [CrossRef]
- Qin, Y.; Wu, X.; Huang, W.; Gong, G.; Li, D.; He, Y.; Zhao, Y. Acute toxicity and sub-chronic toxicity of steroidal saponins from Dioscorea zingiberensis CH Wright in rodents. J. Ethnopharmacol. 2009, 126, 543–550. [Google Scholar] [CrossRef]
- Hassan, S.; Haq, A.; Byrd, J.; Berhow, M.; Cartwright, A.; Bailey, C. Haemolytic and antimicrobial activities of saponin-rich extracts from guar meal. Food Chem. 2010, 119, 600–605. [Google Scholar] [CrossRef]
- Yang, Z.-G.; Sun, H.-X.; Fang, W.-H. Haemolytic activities and adjuvant effect of Astragalus membranaceus saponins (AMS) on the immune responses to ovalbumin in mice. Vaccine 2005, 23, 5196–5203. [Google Scholar] [CrossRef] [PubMed]
- Akbal, O.; Vural, T.; Malekghasemi, S.; Bozdoğan, B.; Denkbaş, E.B. Saponin loaded montmorillonite-human serum albumin nanocomposites as drug delivery system in colorectal cancer therapy. Appl. Clay Sci. 2018, 166, 214–222. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, J.; Yang, X.; Jin, Y.; Yang, Z.; Wang, R.; Zhang, F.; Linhardt, R.J. Mechanism of enhanced oral absorption of akebia saponin D by a self-nanoemulsifying drug delivery system loaded with phospholipid complex. Drug Dev. Ind. Pharm. 2018, 45, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.-W.; Wang, F.-J.; Liew, O.-W.; Lu, Y.-Z.; Zhao, J. Analysis of triterpenoid saponins reveals insights into structural features associated with potent protein drug enhancement effects. Mol. Pharm. 2020, 17, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Santana-Davila, R.; Devisetty, K.; Szabo, A.; Sparapani, R.; Arce-Lara, C.; Gore, E.M.; Moran, A.; Williams, C.D.; Kelley, M.; Whittle, J. Cisplatin and etoposide versus carboplatin and paclitaxel with concurrent radiotherapy for stage III non–small-cell lung cancer: An analysis of veterans health administration data. J. Clin. Oncol. 2015, 33, 567–574. [Google Scholar] [CrossRef]
- Zhao, P.; Song, S.; Du, L.; Zhou, G.; Ma, S.; Li, J.; Feng, J.; Zhu, X.; Jiang, H. Paris Saponins enhance radiosensitivity in a ge-fitinib-resistant lung adenocarcinoma cell line by inducing apoptosis and G2/M cell cycle phase arrest. Mol. Med. Rep. 2016, 13, 2878–2884. [Google Scholar] [CrossRef]
- Chae, S.; Kang, K.A.; Chang, W.Y.; Kim, M.J.; Lee, S.J.; Lee, Y.S.; Kim, H.S.; Kim, N.H.; Hyun, J.W. Effect of compound K, a metabolite of ginseng saponin, combined with γ-Ray radiation in human lung cancer cells in vitro and in vivo. J. Agric. Food Chem. 2009, 57, 5777–5782. [Google Scholar] [CrossRef]
- Koczurkiewicz, P.; Klaś, K.; Grabowska, K.; Piska, K.; Rogowska, K.; Wójcik-Pszczoła, K.; Podolak, I.; Galanty, A.; Michalik, M.; Pękala, E.; et al. Saponins as chemosensitizing substances that improve effectiveness and selectivity of anticancer drug—Minireview of in vitro studies. Phyther. Res. 2019, 33, 2141–2151. [Google Scholar] [CrossRef]
- Baharara, J.; Amini, E.; Nikdel, N.; Salek-Abdollahi, F. The cytotoxicity of dacarbazine potentiated by sea cucumber saponin in resistant B16F10 melanoma cells through apoptosis induction. Avicenna J. Med. Biotechnol. 2016, 8, 112–119. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cells/Tissue Type | Molecular Target | References |
|---|---|---|---|
| Diosgenin | MCF-7, breast cancer | The activation of p53, disruption of intracellular Ca2+ homeostasis, generation of ROS and caspase activation | [21,22] |
| Dioscin | Leukemia, lung cancer, gastric carcinoma, hepatocellular carcinoma, cervical cancer, breast cancer | Upregulates FADD, p53, Bid and Bax. Downregulates CDK2,Bcl-2, Clap-1 and Mcl-1 | [23,24,25] |
| Polyphyllin D | Ovarian cancer, cervical cancer, breast cancer, glioblastoma, glioma | Upregulates p53, p21, PDI and JNX. Downregulates CDK1, Bcl-2, HIF- and VEGF | [26,27,28,29] |
| Oleandrin | Pancreatic cancer, prostate cancer, breast cancer, lymphoma, melanoma, osteosarcoma | Upregulates Akt, ERK and ROS. Downregulates NF-κB, MAPK, JNK, pS6, p4EPB1, PI3K/Akt and mTOR. | [30] |
| Ginsenoside Rg3 | Lung cancer, esophageal carcinoma, gastric cancer, colon cancer, hepatoma, renal cancer, bladder cancer, breast cancer, ovarian cancer, prostate cancer and melanoma | Upregulates p63,p21, Bax and Smac Downregulates VEGF, p38 and P13K, | [17] |
| Ginsenoside Rh2 | Leukemia, colon cancer, hepatocellular carcinoma, breast cancer, ovarian cancer, prostate cancer | Upregulates p53, p21, p27 and p16 Downregulates AKT, CDK4, CDK6 and AP-1. | [17] |
| Saikosaponin A | Hepatocellular carcinoma, breast cancer, colon cancer | Upregulates p15, p16, ERK and cleaved-PARP Downregulates Bcl-2, XIAP, Clap2 and Pgp | [31] |
| Saikosaponin D | Lung cancer, hepatocellular carcinoma, prostate cancer, thyroid cancer | Upregulates p53, p21, Fas and Bax, Downregulates Bcl-2, CDK2, COX-2 and STAT3 | [32] |
| Polyphyllin D | Human non-small-cell lung cancer NCI-H460 cell line. | ER stress-mediated apoptosis, induction of tumor suppressor p53, disruption of mitochondrial membrane and activation of caspase-9 and caspase-3 | [33] |
| Timosaponin AIII (TAIII) | Breast, prostate, HepG2, pancreatic and osteosarcoma cancer cells. PANC-1 cell xenograft nude mice model | ER stress induction, activation of caspase-3, downregulation of Bcl-2, X-linked inhibitor of apoptosis protein (XIAP), Mcl-1 and IAPs, induction of cytochrome c and stimulation of caspases 3, 7, 8 and 9 | [34,35,36] |
| OSW-1(3β,16β,17α-trihydroxycholest-5-en-22-one16- O -(2- O -4-methoxybenzoyl-β- D -xylopyranosyl)-(1→3)-(2- O -acetyl-α- L -arabinopyranoside) | Leukemia cancer and pancreatic cancer cells | Mitochondria membrane permeabilization. Intrinsic apoptosis. Calcium-dependent GRP78 (survival factor) cleavage. Binding to oxysterol binding protein to activate the Golgi stress response leading to apoptosis | [37,38,39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elekofehinti, O.O.; Iwaloye, O.; Olawale, F.; Ariyo, E.O. Saponins in Cancer Treatment: Current Progress and Future Prospects. Pathophysiology 2021, 28, 250-272. https://doi.org/10.3390/pathophysiology28020017
Elekofehinti OO, Iwaloye O, Olawale F, Ariyo EO. Saponins in Cancer Treatment: Current Progress and Future Prospects. Pathophysiology. 2021; 28(2):250-272. https://doi.org/10.3390/pathophysiology28020017
Chicago/Turabian StyleElekofehinti, Olusola Olalekan, Opeyemi Iwaloye, Femi Olawale, and Esther Opeyemi Ariyo. 2021. "Saponins in Cancer Treatment: Current Progress and Future Prospects" Pathophysiology 28, no. 2: 250-272. https://doi.org/10.3390/pathophysiology28020017
APA StyleElekofehinti, O. O., Iwaloye, O., Olawale, F., & Ariyo, E. O. (2021). Saponins in Cancer Treatment: Current Progress and Future Prospects. Pathophysiology, 28(2), 250-272. https://doi.org/10.3390/pathophysiology28020017