
Chitosan from Crabs (Scylla serrata) Represses Hyperlipidemia-Induced Hepato-Renal Dysfunctions in Rats: Modulation of CD43 and p53 Expression

 ,
,  ,
,  ,
,
Abstract
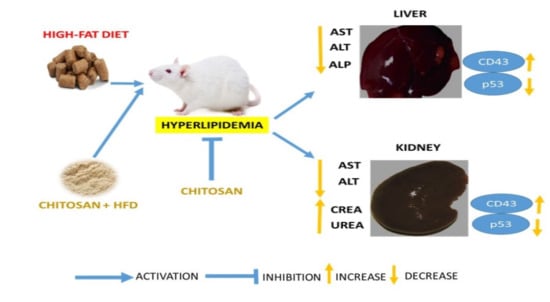
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Crab Shell Collection and Preparation of Raw Materials
2.1.2. Extraction of Chitosan
Purification of Chitosan
2.1.3. Experimental Animals
2.1.4. Experimental Feed Composition
2.2. Methods
2.2.1. Experimental Design
2.2.2. Sacrifice and Collection of Samples
2.2.3. Sample Preparation and Biochemical Analyses
2.2.4. Immuno-Histochemical Analyses for CD43 and p53
2.3. Statistical Analysis
3. Results
3.1. Cumulative Weight Gain of the Experimental Animals
3.2. Effects of Chitosan Supplementation on Renal Biomarkers
3.3. Effects of Chitosan Supplementation on Hepatic Biomarkers
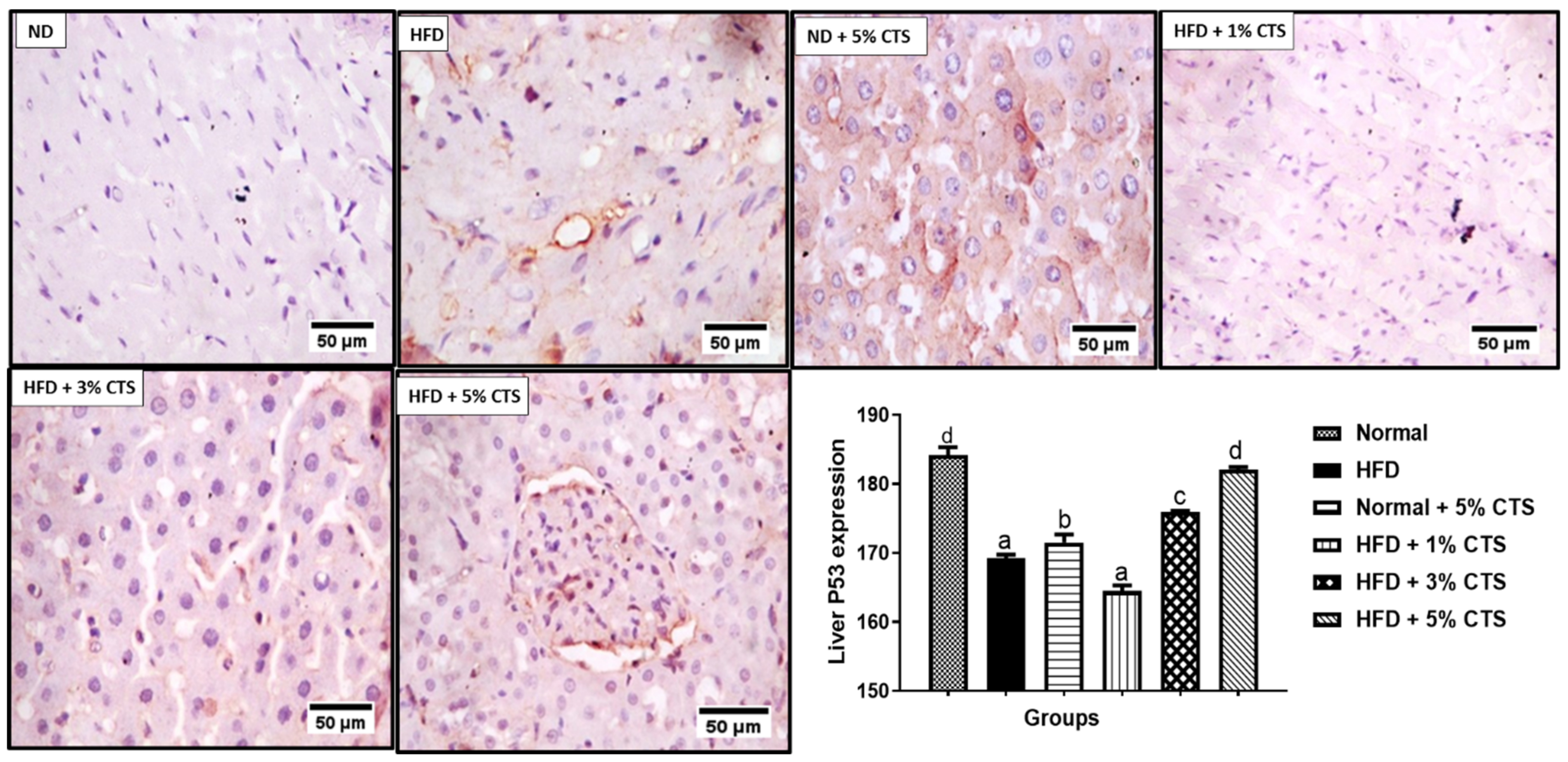
3.4. Effects of Chitosan Supplementation on Hepatic CD43 and p53 Expressions
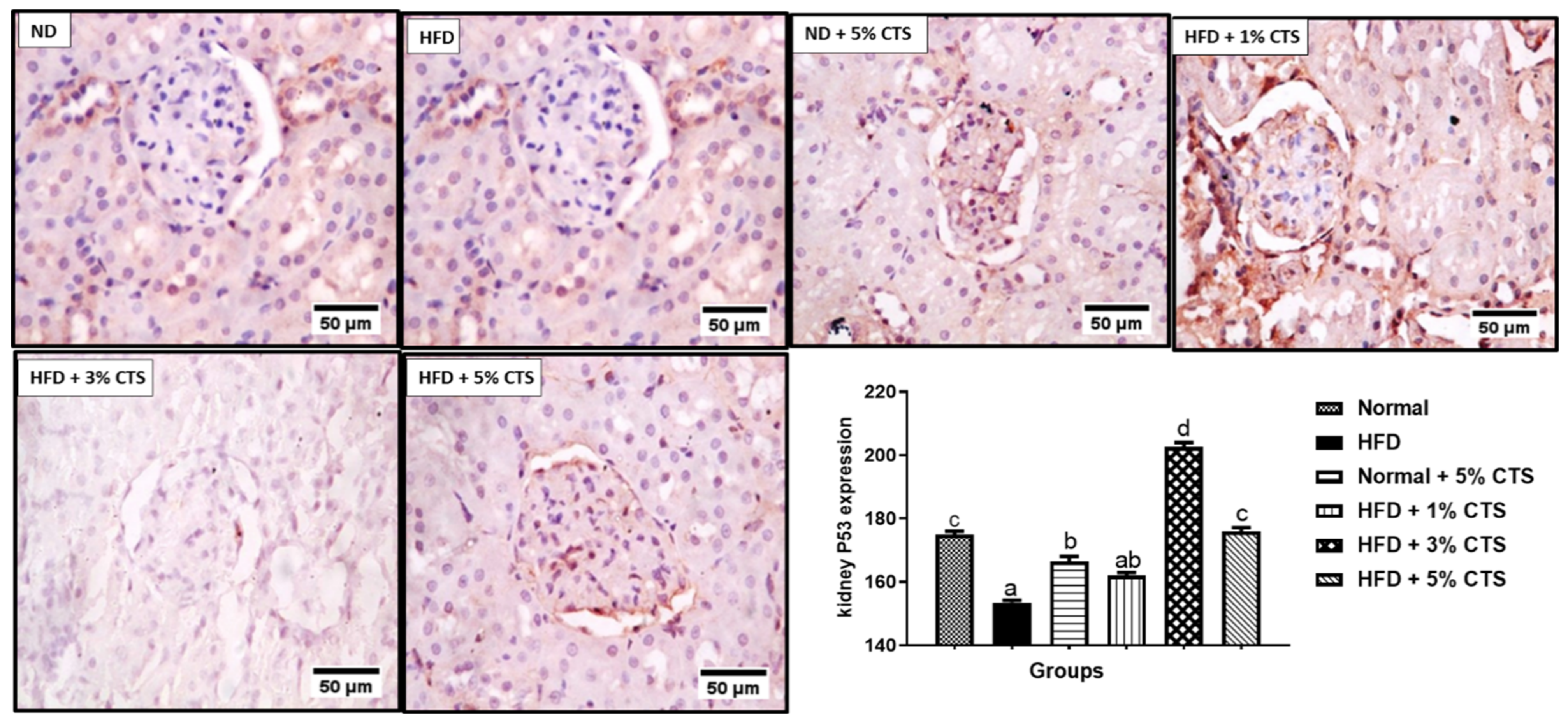
3.5. Effects of Chitosan Supplementation on Renal CD43 and p53 Expressions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ahmad, S.; Beg, Z.H. Hypolipidemic and antioxidant activities of thymoquinone and limonene in atherogenic suspension fed rats. Food Chem. 2013, 138, 1116–1124. [Google Scholar] [CrossRef] [PubMed]
- Navar-Boggan, A.M.; Peterson, E.D.; D’Agostino, S.R.B.; Neely, B.; Sniderman, A.D.; Pencina, M.J. Hyperlipidemia in Early Adulthood Increases Long-Term Risk of Coronary Heart Disease. Circulation 2015, 131, 451–458. [Google Scholar] [CrossRef]
- Ugbaja, R.N.; Akinloye, D.I.; James, A.S.; Ugwor, E.I.; Kareem, S.E.; David, G.; Abolade, S.O.; Oyebade, O.E. Crab derived dietary chitosan mollifies hyperlipidemia- induced oxidative stress and histopathological Derangements in male albino rats. Obes. Med. 2020, 20, 100300. [Google Scholar] [CrossRef]
- Vaziri, N.D. Dyslipidemia of chronic renal failure: The nature, mechanisms, and potential consequences. Am. J. Physiol. Physiol. 2006, 290, F262–F272. [Google Scholar] [CrossRef] [PubMed]
- Kaysen, G.A. Hyperlipidemia of Chronic Renal Failure. Blood Purif. 1994, 12, 60–67. [Google Scholar] [CrossRef]
- Al-Jameil, N.; A Khan, F.; Arjumand, S.; Khan, M.F.; Tabassum, H. Associated liver enzymes with hyperlipidemic profile in type 2 diabetes patients. Int. J. Clin. Exp. Pathol. 2014, 7, 4345–4349. [Google Scholar]
- Balikova, A.; Jääger, K.; Viil, J.; Maimets, T.; Kadaja-Saarepuu, L. Leukocyte marker CD43 promotes cell growth in co-operation with β-catenin in non-hematopoietic cancer cells. Int. J. Oncol. 2012, 41, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Vousden, K.H.; Prives, C. Blinded by the Light: The Growing Complexity of p53. Cell 2009, 137, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Napoli, M.; Flores, E.R. The p53 family orchestrates the regulation of metabolism: Physiological regulation and implications for cancer therapy. Br. J. Cancer 2017, 116, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Orimo, M.; Shimizu, I.; Kunieda, T.; Yokoyama, M.; Ito, T.; Nojima, A.; Nabetani, A.; Oike, Y.; Matsubara, H.; et al. A crucial role for adipose tissue p53 in the regulation of insulin resistance. Nat. Med. 2009, 15, 1082–1087. [Google Scholar] [CrossRef]
- Zhang, H.-L.; Zhong, X.-B.; Tao, Y.; Wu, S.-H.; Su, Z.-Q. Effects of chitosan and water-soluble chitosan micro- and nanoparticles in obese rats fed a high-fat diet. Int. J. Nanomed. 2012, 7, 4069–4076. [Google Scholar] [CrossRef]
- Selvasudha, N.; Koumaravelou, K. The multifunctional synergistic effect of chitosan on simvastatin loaded nanoparticulate drug delivery system. Carbohydr. Polym. 2017, 163, 70–80. [Google Scholar] [CrossRef]
- Zhang, H.-L.; Tao, Y.; Guo, J.; Hu, Y.-M.; Su, Z.-Q. Hypolipidemic effects of chitosan nanoparticles in hyperlipidemia rats induced by high fat diet. Int. Immunopharmacol. 2011, 11, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, J.; Jiang, Q.; Xia, W. The hypolipidemic activity of chitosan nanopowder prepared by ultrafine milling. Carbohydr. Polym. 2013, 95, 487–491. [Google Scholar] [CrossRef]
- Kumar, M.N.V.R.; Muzzarelli, R.A.A.; Muzzarelli, C.; Sashiwa, H.; Domb, A.J. Chitosan Chemistry and Pharmaceutical Perspectives. Chem. Rev. 2004, 104, 6017–6084. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Park, P.J.; Kim, S.K. Antimicrobial effect of chitooligosaccharides produced by bioreactor. Carbohydr. Polym. 2011, 44, 71–76. [Google Scholar] [CrossRef]
- Vo, T.-S.; Kim, S.-K. Marine-derived polysaccharides for regulation of allergic responses. In Advances in Food and Nutrition Research; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 73, pp. 1–13. [Google Scholar]
- Chung, M.J.; Park, J.K.; Park, Y.I. Anti-inflammatory effects of low-molecular weight chitosan oligosaccharides in IgE–antigen complex-stimulated RBL-2H3 cells and asthma model mice. Int. Immunopharmacol. 2012, 12, 453–459. [Google Scholar] [CrossRef]
- Burrows, F.; Louime, C.; Abazinge, M.; Onokpise, O. Extraction and evaluation of chitosan from crab exoskeleton as a seed fungicide and plant growth enhancer. Am. Eurasian J. Agric. Environ. Sci. 2007, 2, 103–111. [Google Scholar]
- Ogungbemi, K.; Ugbaja, R.N.; Ilesanmi, F.F.; Ilori, A.O.; Odeniyi, T.A.; Adeniyi, B.M.; Balogun, D.A.; Ajisafe, S.S. Effect of dietary chitosan on the feed efficiency and weight performance of high fat diet induced hyperlipidemia in male wistar rat. Int. J. Sci. Rep. 2020, 6, 90–94. [Google Scholar] [CrossRef]
- Hussain, M.R.; Iman, M.; Maji, T.K. Determination of degree of deacetylation of chitosan and their effect on the release behavior of essential oil from chitosan and chitosan-gelatin complex microcapsules. Int. J. Adv. Eng. Appl. 2013, 6, 4–12. [Google Scholar]
- Czechowska-Biskup, R.; Jarosińska, D.; Rokita, B.; Ulański, P.; Rosiak, J.M. Determination of degree of deacetylation of chitosan-comparision of methods. Progress Chem. Appl. Chitin Deriv. 2012, 17, 5–20. [Google Scholar]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Oguntoye, O.C.; Oke, O.B. A Comparison of xylazine/ketamine, diazepam/ketamine and acepromazine/ketamine anaesthesia in Rabbit. Sokoto J. Veter. Sci. 2015, 12, 21. [Google Scholar] [CrossRef]
- Krishna, M.B.; Min, X.; Justin, M.A.; Guangbi, L.; Ashley, L.P.; Todd, B.G.; Yang, Z.; Pin-Lan, L. Activation of inflammasomes in podocyte injury of mice on the high fat diet: Effects of ASC gene deletion and silencing. Biochim. et Biophys. Acta 2014, 1843, 836–845. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Gyurászová, M.; Gurecká, R.; Bábíčková, J.; Tóthová, Ľ. Oxidative Stress in the Pathophysiology of Kidney Disease: Implications for Noninvasive Monitoring and Identification of Biomarkers. Oxidative Med. Cell. Longev. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Timofeev, O.; Schlereth, K.; Wanzel, M.; Braun, A.; Nieswandt, B.; Pagenstecher, A.; Rosenwald, A.; Elsässer, H.-P.; Stiewe, T. p53 DNA Binding Cooperativity Is Essential for Apoptosis and Tumor Suppression In Vivo. Cell Rep. 2013, 3, 1512–1525. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Riscal, R.; Arena, G.; Linares, L.K.; Le Cam, L. Metabolic functions of the tumor suppressor p53: Implications in normal physiology, metabolic disorders, and cancer. Mol. Metab. 2020, 33, 2–22. [Google Scholar] [CrossRef]
- Bravo-Adame, M.E.; Vera-Estrella, R.; Barkla, B.J.; Martínez-Campos, C.; Flores-Alcantar, A.; Ocelotl-Oviedo, J.P.; Pedraza-Alva, G.; Rosenstein, Y. An alternative mode of CD43 signal transduction activates pro-survival pathways of T lymphocytes. Immunology 2016, 150, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Fierro, N.A.; Pedraza-Alva, G.; Rosenstein, Y. TCR-Dependent Cell Response Is Modulated by the Timing of CD43 Engagement. J. Immunol. 2006, 176, 7346–7353. [Google Scholar] [CrossRef]
- Xu, R.-Y.; Wan, Y.-P.; Tang, Q.-Y.; Wu, J.; Cai, W. The effects of high fat on central appetite genes in Wistar rats: A microarray analysis. Clin. Chim. Acta 2008, 397, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Dreher, M.L. Role of Fiber and Healthy Dietary Patterns in Body Weight Regulation and Weight Loss. Adv. Obes. Weight Manag. Control. 2015, 3, 00068. [Google Scholar] [CrossRef]
- Ochiai, H.; Shirasawa, T.; Yoshimoto, T.; Nagahama, S.; Watanabe, A.; Sakamoto, K.; Kokaze, A. Elevated alanine aminotransferase and low aspartate aminotransferase/alanine aminotransferase ratio are associated with chronic kidney disease among middle-aged women: A cross-sectional study. BMC Nephrol. 2020, 21, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mansourian, M.; Mirzaei, A.; Azarmehr, N.; Vakilpour, H.; Kokhdan, E.P.; Doustimotlagh, A.H. Hepatoprotective and antioxidant activity of hydroalcoholic extract of Stachys pilifera. Benth on acetaminophen-induced liver toxicity in male rats. Heliyon 2019, 5, e03029. [Google Scholar] [CrossRef]
- Deji, N.; Kume, S.; Araki, S.-I.; Soumura, M.; Sugimoto, T.; Isshiki, K.; Chin-Kanasaki, M.; Sakaguchi, M.; Koya, D.; Haneda, M.; et al. Structural and functional changes in the kidneys of high-fat diet-induced obese mice. Am. J. Physiol. Physiol. 2009, 296, F118–F126. [Google Scholar] [CrossRef]
- Zaid, O.A.; Elsonbaty, S.; Moawad, F.; Abdelghaffar, M. Antioxidants and hepatoprotective effects of chitosan nanoparticles against hepatotoxicity induced in rats. Benha Veter. Med. J. 2019, 36, 252–261. [Google Scholar] [CrossRef]
- Anna, B.; Solaiman, D.; Alexey, S.; Sali, D. Pharmacological and biological effects of chitosan. Res. J. Pharm. Technol. 2020, 13, 1043. [Google Scholar] [CrossRef]
- Matica, A.; Menghiu, G.; Ostafe, V. Toxicity of Chitosan Based Products. New Front. Chem. 2017, 26, 65–74. [Google Scholar]
- Barrea, L.; Altieri, B.; Polese, B.; De Conno, B.; Muscogiuri, G.; Colao, A.; Savastano, S. Nutritionist and obesity: Brief overview on efficacy, safety, and drug interactions of the main weight-loss dietary supplements. Int. J. Obes. Suppl. 2019, 9, 32–49. [Google Scholar] [CrossRef]
- Aylon, Y.; Oren, M. The Hippo pathway, p53 and cholesterol. Cell Cycle 2016, 15, 2248–2255. [Google Scholar] [CrossRef]
- Komarova, E.A.; Krivokrysenko, V.; Wang, K.; Neznanov, N.; Chernov, M.V.; Komarov, P.G.; Brennan, M.; Golovkina, T.V.; Rokhlin, O.W.; Kuprash, D.V.; et al. p53 is a suppressor of inflammatory response in mice. FASEB J. 2005, 19, 1030–1032. [Google Scholar] [CrossRef] [PubMed]
- Krstic, J.; Reinisch, I.; Schupp, M.; Schulz, T.J.; Prokesch, A. p53 Functions in Adipose Tissue Metabolism and Homeostasis. Int. J. Mol. Sci. 2018, 19, 2622. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kim, T.-H.; Franklin, D.A.; Zhang, Y. Protection against High-Fat-Diet-Induced Obesity in MDM2 C305F Mice Due to Reduced p53 Activity and Enhanced Energy Expenditure. Cell Rep. 2017, 18, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Thomas, J.L.; Chen, J.-Z.; Jan, J.-S.; Lin, H.-Y. Activation of tumor suppressor p53 gene expression by magnetic thymine-imprinted chitosan nanoparticles. Chem. Commun. 2015, 52, 2137–2140. [Google Scholar] [CrossRef] [PubMed]
- Meiler, S.; Baumer, Y.; McCurdy, S.; Lee, B.-H.; Kitamoto, S.; Boisvert, W.A. Cluster of Differentiation 43 Deficiency in Leukocytes Leads to Reduced Atherosclerosis—Brief Report. Arter. Thromb. Vasc. Biol. 2015, 35, 309–311. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gruber, S.; Hendrikx, T.; Tsiantoulas, D.; Ozsvar-Kozma, M.; Göderle, L.; Mallat, Z.; Witztum, J.L.; Shiri-Sverdlov, R.; Nitschke, L.; Binder, C.J. Sialic Acid-Binding Immunoglobulin-like Lectin G Promotes Atherosclerosis and Liver Inflammation by Suppressing the Protective Functions of B-1 Cells. Cell Rep. 2016, 14, 2348–2361. [Google Scholar] [CrossRef]
- Kadaja, L.; Laos, S.; Maimets, T. Overexpression of leukocyte marker CD43 causes activation of the tumor suppressor proteins p53 and ARF. Oncogene 2003, 23, 2523–2530. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ngo, D.-H.; Vo, T.-S.; Kang, K.-H.; Je, J.-Y.; Pham, H.N.-D.; Byun, H.-G.; Kim, S.-K. Biological effects of chitosan and its derivatives. Food Hydrocoll. 2015, 51, 200–216. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Composition | Ingredients | Normal Diet (g/100 g Diet) | High-Fat Diet (g/100 g Diet) |
|---|---|---|---|
| Carbohydrates | Maize | 40 | 10 |
| Sucrose | 10 | 10 | |
| Lipids | Lard | – | 35 |
| Soya oil | 5 | 5 | |
| Protein | Soya | 10 | 10 |
| Groundnut cake | 15 | 15 | |
| Fish meal | 10 | 10 | |
| Fiber | Wheat fiber | 5 | - |
| Mineral mix * | 2.5 | 2.5 | |
| Vitamin mix # | 1 | 1 | |
| Methionine | 0.5 | 0.5 | |
| Lysine | 0.5 | 0.5 | |
| Salt (NaCl) | 0.5 | 0.5 |
| Groups | Final Body Weight (g) | Average Weekly Weight Gain (g/week) |
|---|---|---|
| Normal control | 132.34 ± 1.62 b | 7.06 ± 0.28 b |
| High-fat diet (HFD) | 150.06 ± 4.45 c | 10.01 ± 0.57 c |
| Normal + 5% CTS | 117.52 ± 1.46 a | 4.59 ± 0.18 a |
| HFD + 1% CTS | 116.60 ± 2.07 a | 4.43 ± 0.17 a |
| HFD + 3% CTS | 136.42 ± 2.89 b | 7.74 ± 0.26 b |
| HFD + 5% CTS | 130.78 ± 1.35 b | 6.80 ± 0.21 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ugbaja, R.N.; Ogungbemi, K.; James, A.S.; Peter Folorunsho, A.; Abolade, S.O.; Ajamikoko, S.O.; Atayese, E.O.; Adedeji, O.V. Chitosan from Crabs (Scylla serrata) Represses Hyperlipidemia-Induced Hepato-Renal Dysfunctions in Rats: Modulation of CD43 and p53 Expression. Pathophysiology 2021, 28, 224-237. https://doi.org/10.3390/pathophysiology28020015
Ugbaja RN, Ogungbemi K, James AS, Peter Folorunsho A, Abolade SO, Ajamikoko SO, Atayese EO, Adedeji OV. Chitosan from Crabs (Scylla serrata) Represses Hyperlipidemia-Induced Hepato-Renal Dysfunctions in Rats: Modulation of CD43 and p53 Expression. Pathophysiology. 2021; 28(2):224-237. https://doi.org/10.3390/pathophysiology28020015
Chicago/Turabian StyleUgbaja, Regina Ngozi, Kunle Ogungbemi, Adewale Segun James, Ayodele Peter Folorunsho, Samuel Olanrewaju Abolade, Stella Onajite Ajamikoko, Eniola Olapeju Atayese, and Omowunmi Victoria Adedeji. 2021. "Chitosan from Crabs (Scylla serrata) Represses Hyperlipidemia-Induced Hepato-Renal Dysfunctions in Rats: Modulation of CD43 and p53 Expression" Pathophysiology 28, no. 2: 224-237. https://doi.org/10.3390/pathophysiology28020015
APA StyleUgbaja, R. N., Ogungbemi, K., James, A. S., Peter Folorunsho, A., Abolade, S. O., Ajamikoko, S. O., Atayese, E. O., & Adedeji, O. V. (2021). Chitosan from Crabs (Scylla serrata) Represses Hyperlipidemia-Induced Hepato-Renal Dysfunctions in Rats: Modulation of CD43 and p53 Expression. Pathophysiology, 28(2), 224-237. https://doi.org/10.3390/pathophysiology28020015