


Induction, Proliferation, Regeneration and Kinsenoside and Flavonoid Content Analysis of the Anoectochilus roxburghii (Wall.) Lindl Protocorm-like Body

Abstract
1. Introduction
2. Results
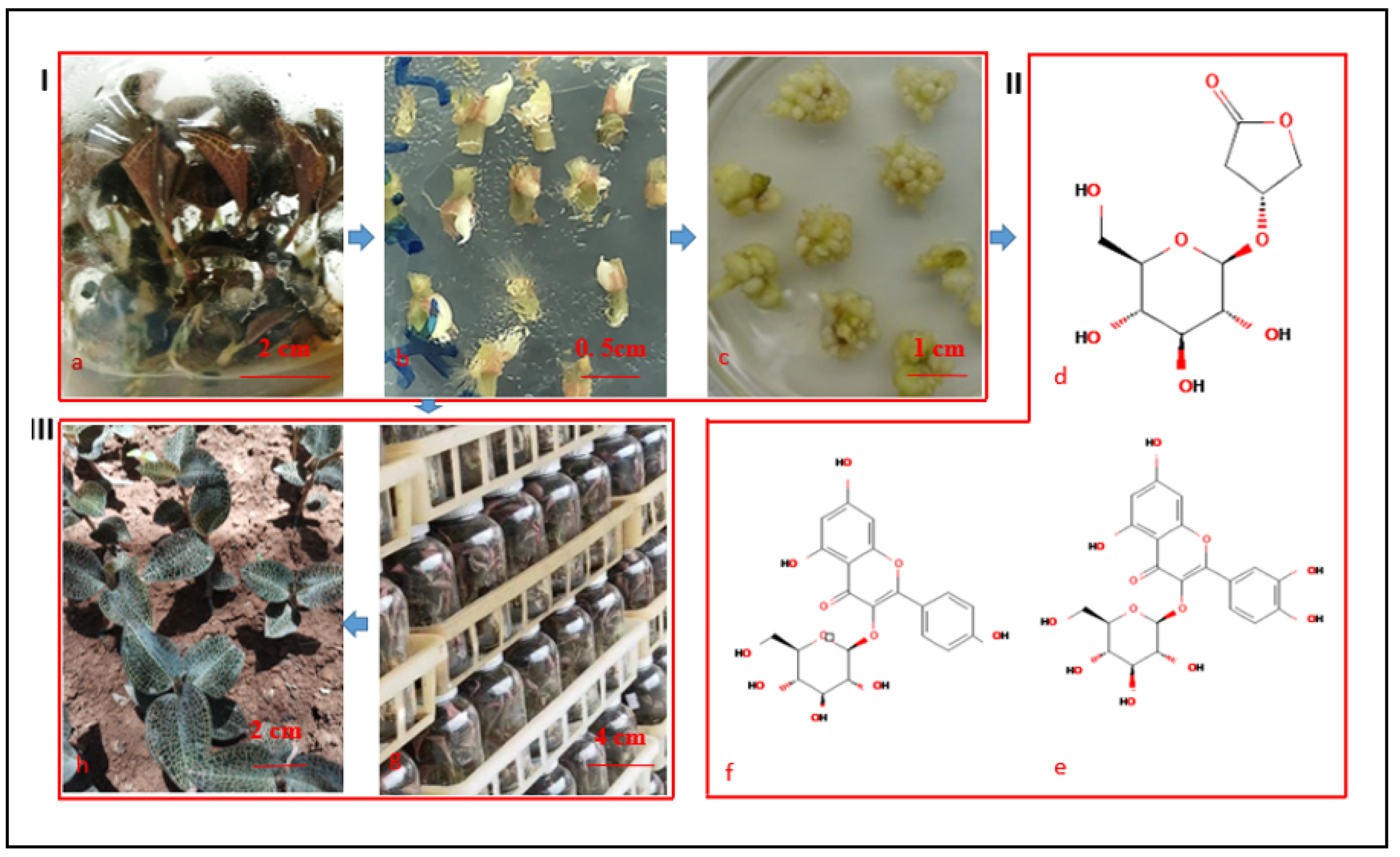
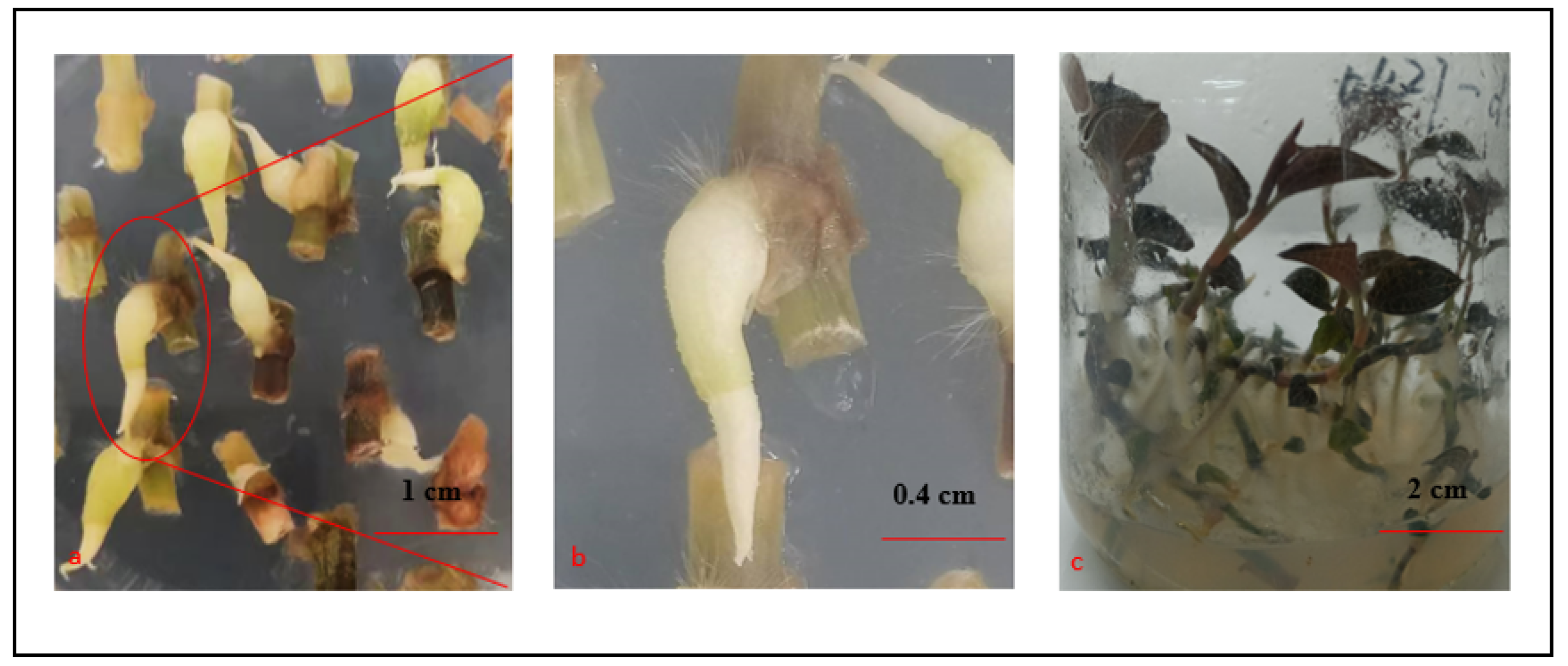
2.1. Induction of PLBs
2.2. Proliferation of PLBs (Induction and Proliferation of Secondary PLBs)
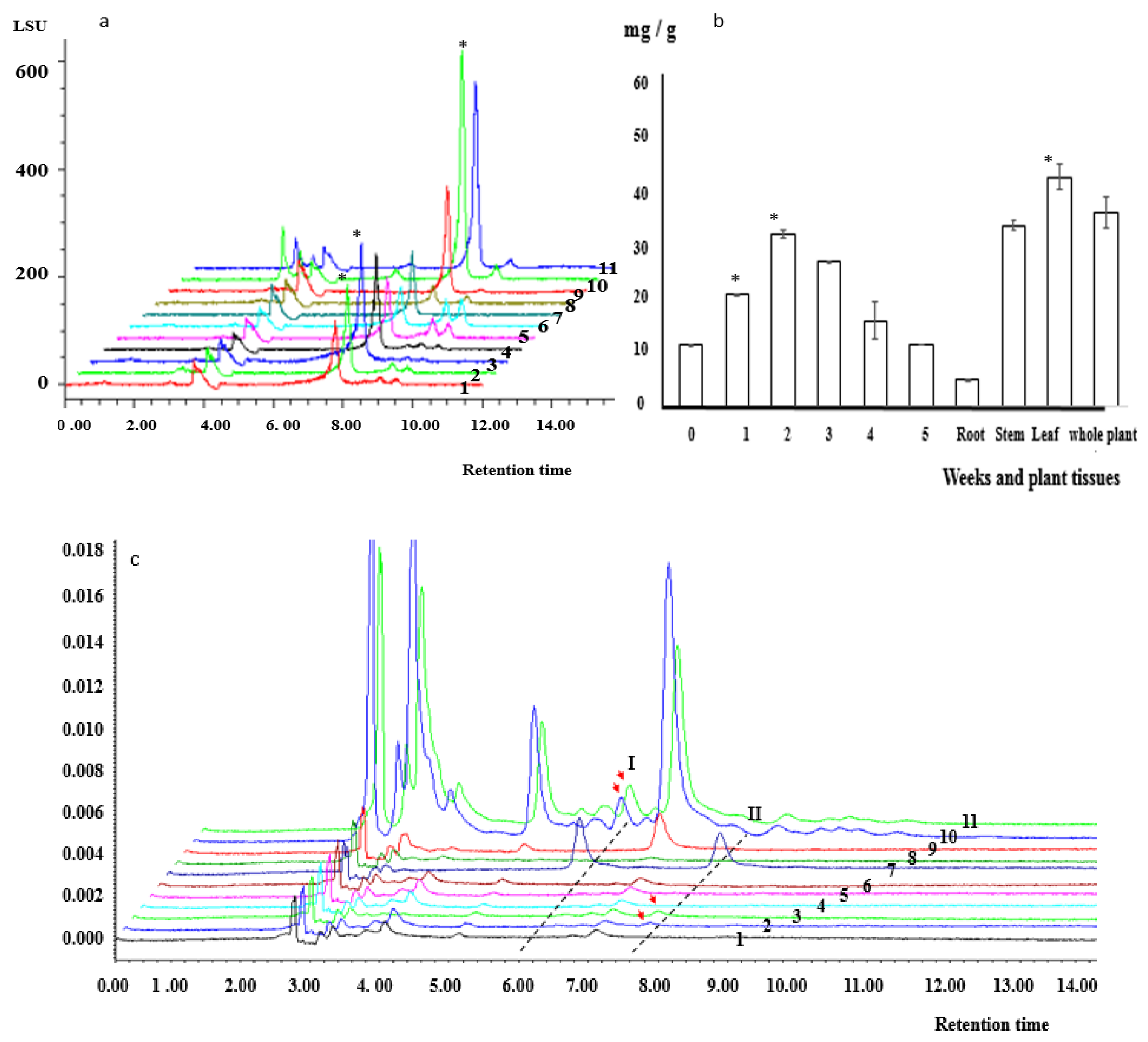
2.3. Kinsenoside and Flavonoid Contents of PLBs
2.4. Regeneration of PLBs and the Formation of Shoots and Roots
2.5. Acclimation of In Vitro Grown Plantlets
3. Discussion
3.1. Induction of A. roxburghii PLBs
3.2. Proliferation of A. roxburghii PLBs
3.3. Kinsenoside and Flavonoid Contents in PLBs
3.4. Regeneration of PLBs: Shoot and Root Formation
4. Materials and Methods
4.1. Plant Materials
4.2. PLB Induction
4.3. PLB Proliferation (Induction and Proliferation of Secondary PLBs)
4.4. Determination of Kinsenoside, Isoquercitrin, and Astragalin Contents
4.5. Regeneration of PLBs: Shoot and Root Formation
4.6. Acclimation of Plantlets
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nanjing University of Chinese Medicine. The Secondary Version of Chinese Materia Medica Dictionary; Shanghai Science and Technology Press: Shanghai, China, 2005; pp. 1956–1957. [Google Scholar]
- He, C.N.; Wang, C.L. Advances on chemical constituents and pharmacological activities of Anoectochilus plant. Chin. Pharm. J. 2004, 39, 81–84. [Google Scholar]
- Ye, S.Y.; Shao, Q.S. Anoectochilus roxburghii: A review of its photochemistry, pharmacology, and clinical applications. J. Ethnopharmacol. 2017, 209, 184–202. [Google Scholar] [CrossRef]
- Fujian Institute of Traditional Chinese Medicine. Record of Fujian Materia Medica, 2nd ed.; Fujian Science and Technology Press: Fuzhou, China, 1982. [Google Scholar]
- Editorial Committee of Flora of China and Chinese Academy of Sciences. Flora of China; Science Press: Beijing, China, 1999; Volume 17, p. 225. [Google Scholar]
- He, B.Z.; He, G.R. Micropropagation Technology of Anoectochilus roxburghii in Fujian Province. Agric. Eng. 2013, 3, 72–76. [Google Scholar]
- Yang, C.H.; Li, X. Research progress on tissue culture of Anoectochilus roxburghii. Pratcult. Sci. 2018, 35, 1047–1056. [Google Scholar]
- Shao, Q.S.; Ye, S.Y. Current researches and prospects of seedling propagation and cultivation modes of Jinxianlian. China J. Chin. Mater. Med. 2016, 40, 160–166. [Google Scholar]
- Lee, Y.I.; Hsu, S.T. Orchid protocorm-like bodies are somatic embryos. Am. J. Bot. 2013, 100, 2121–2131. [Google Scholar] [CrossRef]
- Cardoso, J.C.; Zanello, C.A.; Chen, J.T. An Overview of Orchid Protocorm-Like Bodies: Mass Propagation, Biotechnology, Molecular Aspects, and Breeding. Int. J. Mol. Sci. 2020, 21, 985. [Google Scholar] [CrossRef] [PubMed]
- Gantait, S.; Sinniah, U.R. Direct induction of protocorm-like bodies from shoot tips, plantlet formation, and clonal fidelity analysis in Anthurium andreanum cv. CanCan. Plant Growth Regul. 2012, 67, 257–270. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Li, A. Induction of Protocorm-like Bodies and Plant Regeneration of Pinellia ternate (Thunb.) Breit. Chin. Agric. Sci. Bull. 2013, 29, 153–158. [Google Scholar]
- Li, X.L.; Zhao, Z.C. Optimization of protocol of tissue culture and rapid propagation of Pinellia ternate of protocorm-like body. North. Hortic. 2018, 1, 14–18. [Google Scholar]
- Da Silva, J.A.T.; Cardoso, C.J. Dendrobium micropropagation: A review. Plant Cell Rep. 2015, 34, 671–704. [Google Scholar] [CrossRef] [PubMed]
- Han, X.H.; Wang, C.L. Effects of different hormone levels on tissue culture of Anoectochilus roxburghii. Guangdong Agric. Sci. 2012, 39, 94–97. [Google Scholar]
- Han, X.H.; Luo, M. Study on the proliferation of protocorm-like bodies in Anoetochochilus roburghii. North. Hortic. 2012, 36, 144–146. [Google Scholar]
- Jiao, Z. Optimization of the extraction condition of flavone form protocorm-like bodies of Anoectochilus roxburghii. Heilongjiang Agric. Sci. 2018, 10, 144–148. [Google Scholar]
- Jiao, Z.; An, S.J. Establishment of new protocorm-like bodies system of Anoectochilus roxburghii by ‘double two-steps’ Method. Chin. J. Trop. Crops 2019, 40, 314–322. [Google Scholar]
- Picolotto, D.R.N.; Paiva Neto, V.B. Micropropagation of Cyrtopodium paludicolum (Orchidaceae) from root tip explants. Crop Breed. App. Biotech. 2017, 17, 191–197. [Google Scholar] [CrossRef][Green Version]
- Samala, S.; Te-chato, S. Protocorm-like body of Grammatophyllum speciosum through a symbiotic seed germination. Sci. Asia 2014, 40, 379–383. [Google Scholar] [CrossRef][Green Version]
- Naing, A.H.; Chung, J.D. Efficient plant regeneration of the endangered medicinal orchid, Coelogyne cristata using protocorm-like bodies. Acta Physiol. Plant 2011, 33, 659–666. [Google Scholar] [CrossRef]
- Kalyan, K.; Sil, S. Protocorm-like bodies and plant regeneration from foliar explants of Coelogyne flaccida, a horticulturally and medicinally important endangered orchid of eastern himalaya. Lanke 2015, 15, 151–158. [Google Scholar]
- Chin, C.K.; Stanly, C. Protocorm-like bodies (PLBs) of Dendrobium Sabin Blue: A novel source for in vitro production of dendrobine and anthocyanin. In Vitro Cell. Dev. Biol. Plant 2021, 57, 874–882. [Google Scholar] [CrossRef]
- Qi, C.X.; Zho, Q. Kinsenoside: A Promising Bioactive Compound from Anoectochilus Species. Curr. Med. Sci. 2018, 38, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.D.; Wang, H.Z. Recent advances in kinsenoside. Chin. Hosp. Pharm. 2015, 35, 1795–1798. [Google Scholar]
- Hong, L.; Shao, Q.S. Current status and sustainable development countermeasures of Anoectochilus roxburghii. China J. Chin. Mater. Med. 2015, 40, 553–558. [Google Scholar]
- Feng, Y.L.; Yu, Y.T. Effect of Plant Growth Substance on Regeneration of Anoectochilus formosanus. North. Horticult. 2010, 34, 136–138. [Google Scholar]
- Wei, C.H. Optimization of PLB induction conditions for Oncidium. Fujian J. Agric. Sci. 2007, 22, 332–335. [Google Scholar]
- Li, W.L.; Zhai, L.S. Study on induction and culture of Oncidium protocorm-like body (PLB).Henan. Science 2004, 22, 360–362. [Google Scholar]
- Gow, W.; Chen, J. Enhancement of direct somatic embryogenesis and plantlet growth from leaf explants of Phalaenopsis by adjusting culture period and explant length. Acta Physiol. Plant. 2010, 32, 621–627. [Google Scholar] [CrossRef]
- Zanello, C.A.; Cardoso, J.C. PLBs induction and clonal plantlet regeneration from leaf segment of commercial hybrids of Phalaenopsis. J. Hortic. Sci. Biotechnol. 2019, 94, 627–631. [Google Scholar] [CrossRef]
- Wang, J.Q.; Chen, G. Tissue culture induction of Anoectochilus rocburghii protocorm. J. Chin. Med. Mater. 1995, 1, 3–5. [Google Scholar]
- Wang, J.Q.; Lin, L. Protocorm initiation and plantlet regeneration of Anoectochilus roxbburhii. Chin. Bull. Bot. 1996, 13, 54–55. [Google Scholar]
- Gow, W.; Chen, J. Effects of genotype, light regime, explant position, and orientation on direct somatic embryogenesis from leaf explants of Phalaenopsis orchid. Acta Physiol. Plant 2009, 31, 363–369. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, W. Changes in Kinsenoside Content of Different Strains of Anoectochilus roxburghii at Different Growth Periods Mod. Chin. Med. 2021, 23, 1423–1429. [Google Scholar]
- Neugart, S.; Fiol, M. Interaction of Moderate UV-B Exposure and Temperature on the Formation of Structurally Different Flavonol Glycosides and Hydroxycinnamic Acid Derivatives in Kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2014, 62, 4054–4062. [Google Scholar] [CrossRef]
- Neugart, S.; Klaring, H.P. The effect of temperature and radiation on flavonol aglycones and flavonol glycosides of kale (Brassica oleracea var. sabellica). Food Chem. 2012, 133, 1456–1465. [Google Scholar] [CrossRef]
- Neugart, S.; Zietz, M. Structurally different flavonol glycosides and hydroxycinnamic acid derivatives respond differently to moderate UV-B radiation exposure. Physiol. Plant. 2012, 145, 582–593. [Google Scholar] [CrossRef]
- Yu, X.F.; Zhu, Y.L. Accumulation of Flavonoid Glycosides and UFGT Gene Expression in Mulberry Leaves (Morus alba L.) before and after Frost. Chem. Biodivers. 2017, 14, e1600496. [Google Scholar] [CrossRef]
- Cavallaro, V.; Pellegrino, A. Light and Plant Growth Regulators on In Vitro Proliferation. Plants 2022, 11, 844. [Google Scholar] [CrossRef]
- Suzuki, R.M.; Kerbauy, G.B. Effects of light and ethylene on endogenous hormones and development of Catasetum fimbriatum (Orchidaceae). Braz. J. Plant Physiol. 2006, 18, 359–365. [Google Scholar] [CrossRef]
- Zhang, A.L.; Wang, H.Z. Large scale in vitro propagation of Anoectochilus roxburghii for commercial application: Pharmaceutically important and ornamental plant. Ind. Crops Prod. 2015, 70, 158–162. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Neto, V.B. In vitro propagation of Cyrtopodium saintlegerianum rchb. f. (orchidaceae), a native orchid of the Brazilian savannah. Crop Breed. Appl. Biotechnol. 2015, 15, 10–17. [Google Scholar] [CrossRef]
- Lin, X.L.; Yang, Z.X. Micropropagation and transplanting techniques in Anoectochilus roxburghii (Wall.) Lindl. Hortic. Seed 2016, 5, 6–9. [Google Scholar]
- Parthibhan, S.; Kumar, T.S.; Rao, M.V. Phenology and reintroduction strategies for Dendrobium aqueum Lindley—An endemic, near threatened orchid. J. Nat. Conserv. 2015, 24, 68–71. [Google Scholar] [CrossRef]
- Mohanty, P.; Paul, S. A simple and efficient protocol for the mass propagation of Cymbidium mastersii: An ornamental orchid of Northeast India. AoB Plants 2012, 2012, pls023. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGRs (mg/L) | PLB Appearance Time | PLB Induction Rate (%) | |||
|---|---|---|---|---|---|
| 6-BA | NAA | ZT | 2, 4-D | ||
| 0 | 0 | 0 | 0 | 25 ± 1.00 | 60 ± 1.50 |
| 3 | 0.5 | 0 | 0 | 20.3 ± 1.53 | 65 ± 1.44 |
| 3 | 0.5 | 0.2 | 0 | 22.6 ± 2.08 * | 80 ± 1.52 |
| 3 | 0.5 | 0.5 | 0 | 23.6 ± 1.52 | 83.5 ± 1.32 * |
| 3 | 0.5 | 0.8 | 0 | 21.0 ± 1.00 | 83 ± 1.22 |
| 3 | 0.5 | 0.8 | 0.2 | 19.3 ± 1.53 * | 89 ± 2.08 * |
| Hormone (mg/L) | Induction Rate (Shoots/PLB Mass) (Mean ± SD) | Shoot Length (Mean ± SD) (cm) | Shoot Diameter (Mean ± SD) (mm) | |
|---|---|---|---|---|
| 6-BA | NAA | |||
| 0 | 0 | 5.0 ± 0.23 | 3.01 ± 0.45 | 2.02 ± 0.44 |
| 1.0 | 0.5 | 6.5 ± 0.51 * | 2.89 ± 0.56 | 2.61 ± 0.11 |
| 2.0 | 0.5 | 8.2 ± 0.05 | 2.78. ± 0.11 * | 2.97 ± 0.45 |
| 3.0 | 0.5 | 10.5 ± 0.64 * | 2.79 ± 0.75 * | 3.19 ± 0.33 * |
| 4.0 | 0.5 | 9.1 ± 0.67 | 2.56 ± 0.33 | 2.60 ± 0.22 |
| 5.0 | 0.5 | 4.3 ± 1.2 * | 2.43 ± 0.24 | 2.43 ± 0.35 |
| PGRs (mg/L) | Rooting Rate (Mean ± SD)% | Root NUMBER (Mean ± SD) cm | Root Length (Mean ± SD) cm | ||
|---|---|---|---|---|---|
| NAA | 6-BA | Indole Butyric Acid (IBA) | |||
| 0.5 | 0 | 0 | 98.35 ± 0.05 | 3.22 ± 0.21 | 3.51 ± 0.12 |
| 0.5 | 0.1 | 0 | 93.45 ± 0.51 * | 2.89 ± 0.56 | 3.34 ± 0.17 * |
| 0.5 | 0.1 | 4 | 93.14 ± 0.23 * | 3.01 ± 0.45 * | 2.89 ± 0.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Chen, X.; Yan, X.; Xu, Z.; Shao, Q.; Wu, X.; Tou, L.; Fang, L.; Wei, M.; Wang, H. Induction, Proliferation, Regeneration and Kinsenoside and Flavonoid Content Analysis of the Anoectochilus roxburghii (Wall.) Lindl Protocorm-like Body. Plants 2022, 11, 2465. https://doi.org/10.3390/plants11192465
Wang H, Chen X, Yan X, Xu Z, Shao Q, Wu X, Tou L, Fang L, Wei M, Wang H. Induction, Proliferation, Regeneration and Kinsenoside and Flavonoid Content Analysis of the Anoectochilus roxburghii (Wall.) Lindl Protocorm-like Body. Plants. 2022; 11(19):2465. https://doi.org/10.3390/plants11192465
Chicago/Turabian StyleWang, Hongzhen, Xinying Chen, Xiaoyun Yan, Zhixia Xu, Qingsong Shao, Xueqian Wu, Lingjuan Tou, Li Fang, Mingming Wei, and Huasen Wang. 2022. "Induction, Proliferation, Regeneration and Kinsenoside and Flavonoid Content Analysis of the Anoectochilus roxburghii (Wall.) Lindl Protocorm-like Body" Plants 11, no. 19: 2465. https://doi.org/10.3390/plants11192465
APA StyleWang, H., Chen, X., Yan, X., Xu, Z., Shao, Q., Wu, X., Tou, L., Fang, L., Wei, M., & Wang, H. (2022). Induction, Proliferation, Regeneration and Kinsenoside and Flavonoid Content Analysis of the Anoectochilus roxburghii (Wall.) Lindl Protocorm-like Body. Plants, 11(19), 2465. https://doi.org/10.3390/plants11192465