
Fetal Programming of the Endocrine Pancreas: Impact of a Maternal Low-Protein Diet on Gene Expression in the Perinatal Rat Pancreas

 , and
, and 
Abstract
1. Introduction
2. Results
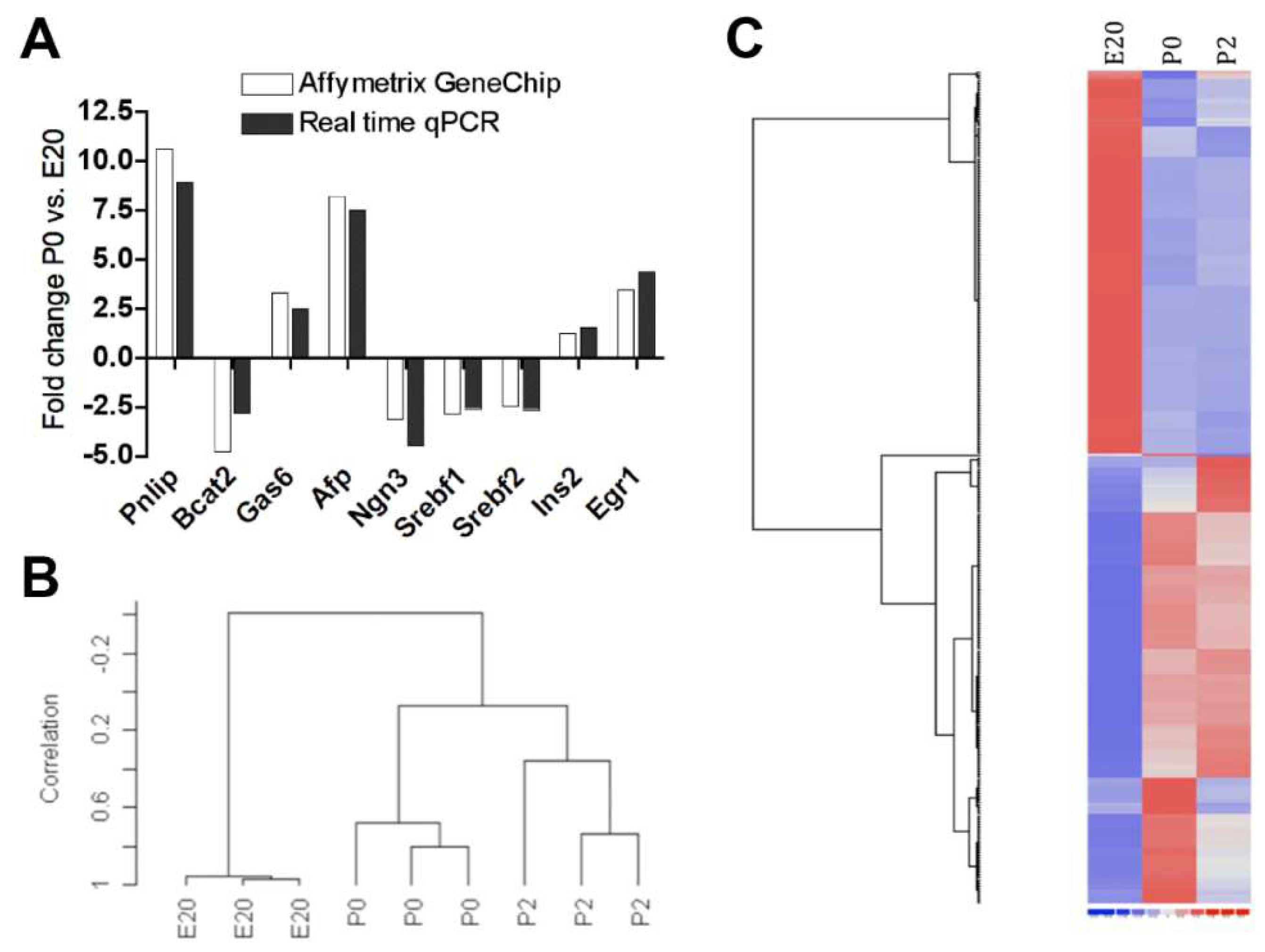
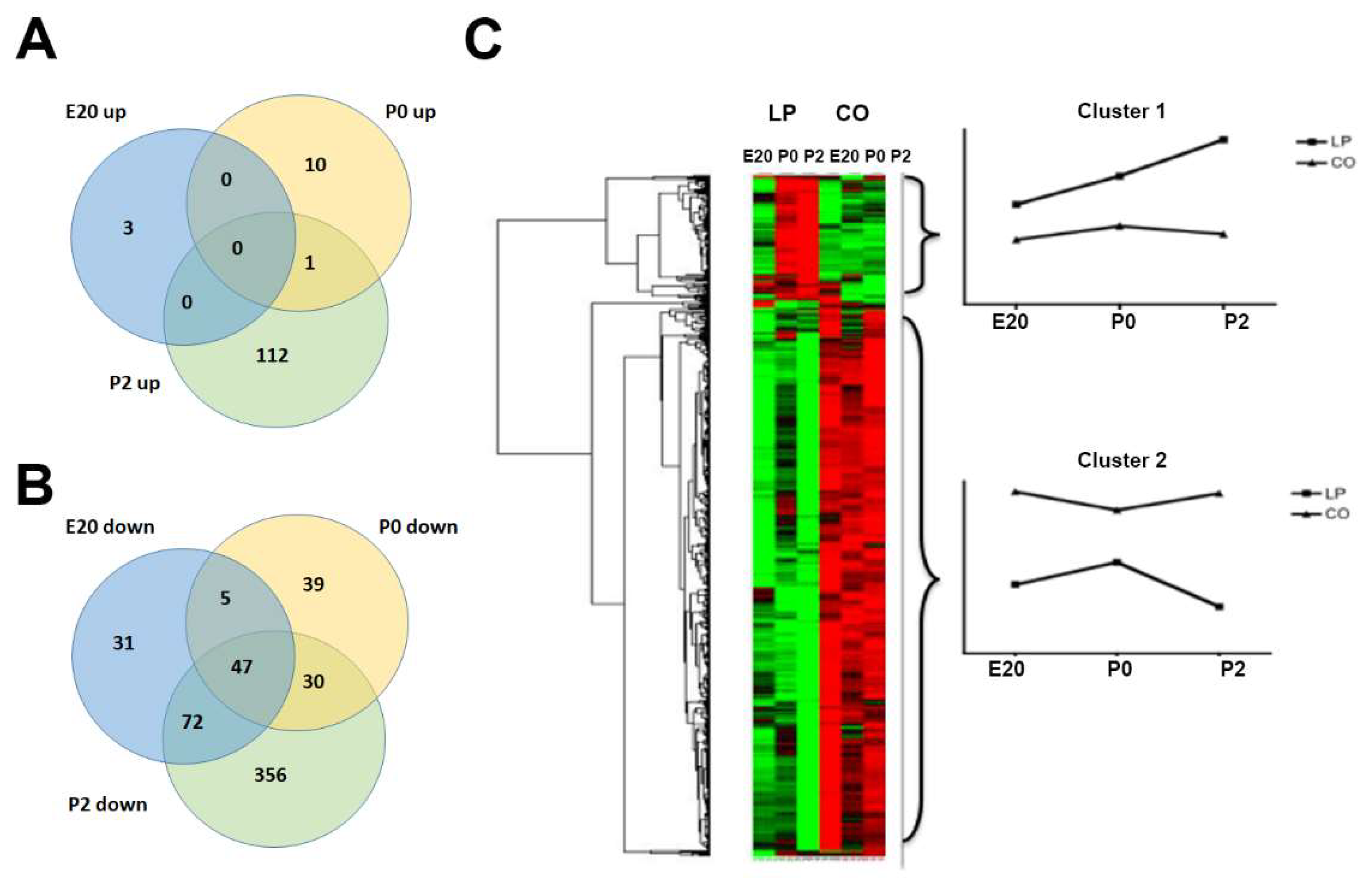
2.1. Gene Regulation in Normal Perinatal Rat Pancreas from E20 to P2
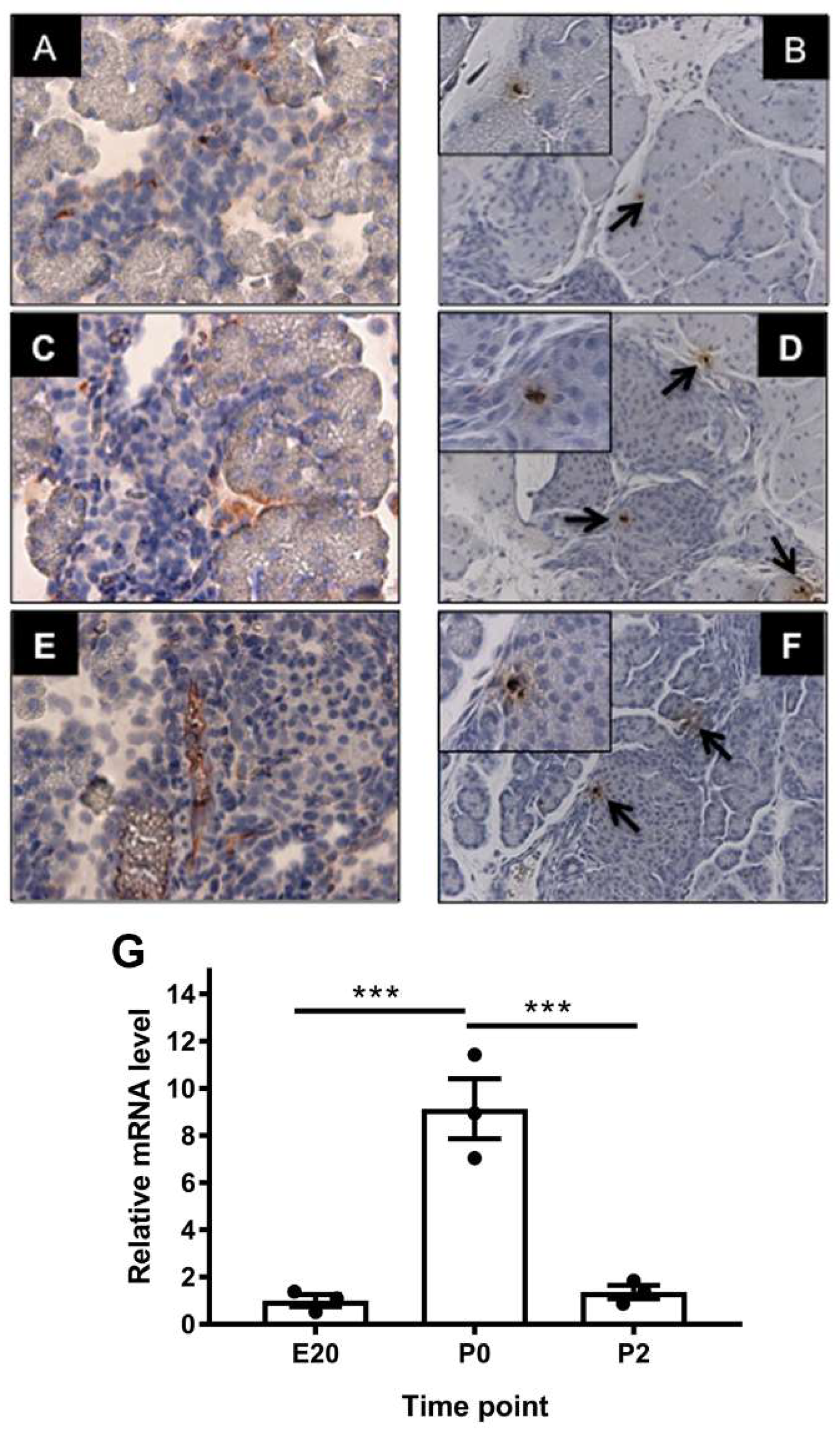
2.2. Localization of Alpha-Feto Protein (Afp) in Perinatal Rat Pancreas
2.3. Clustering and Functional Annotation of Profiles
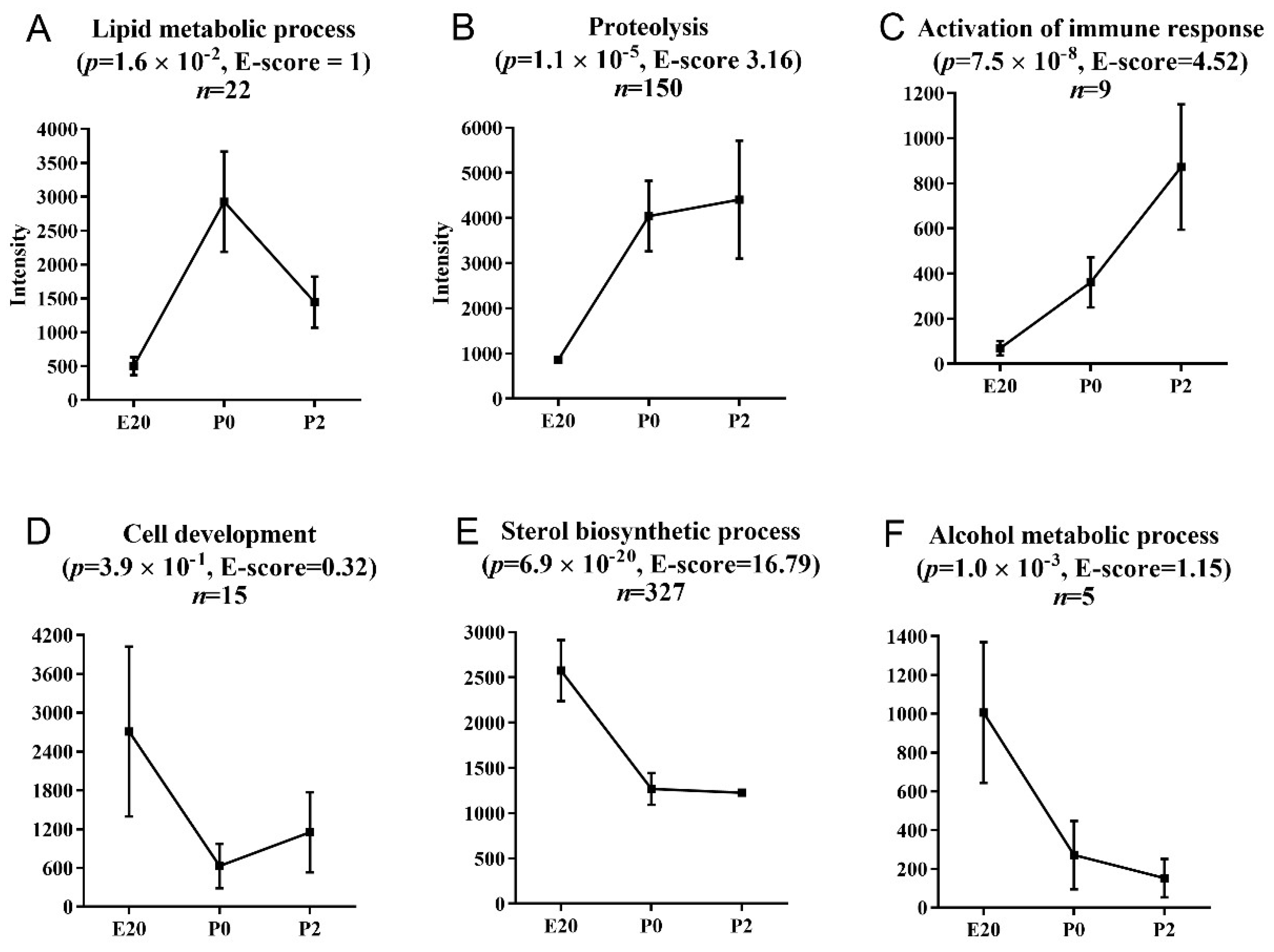
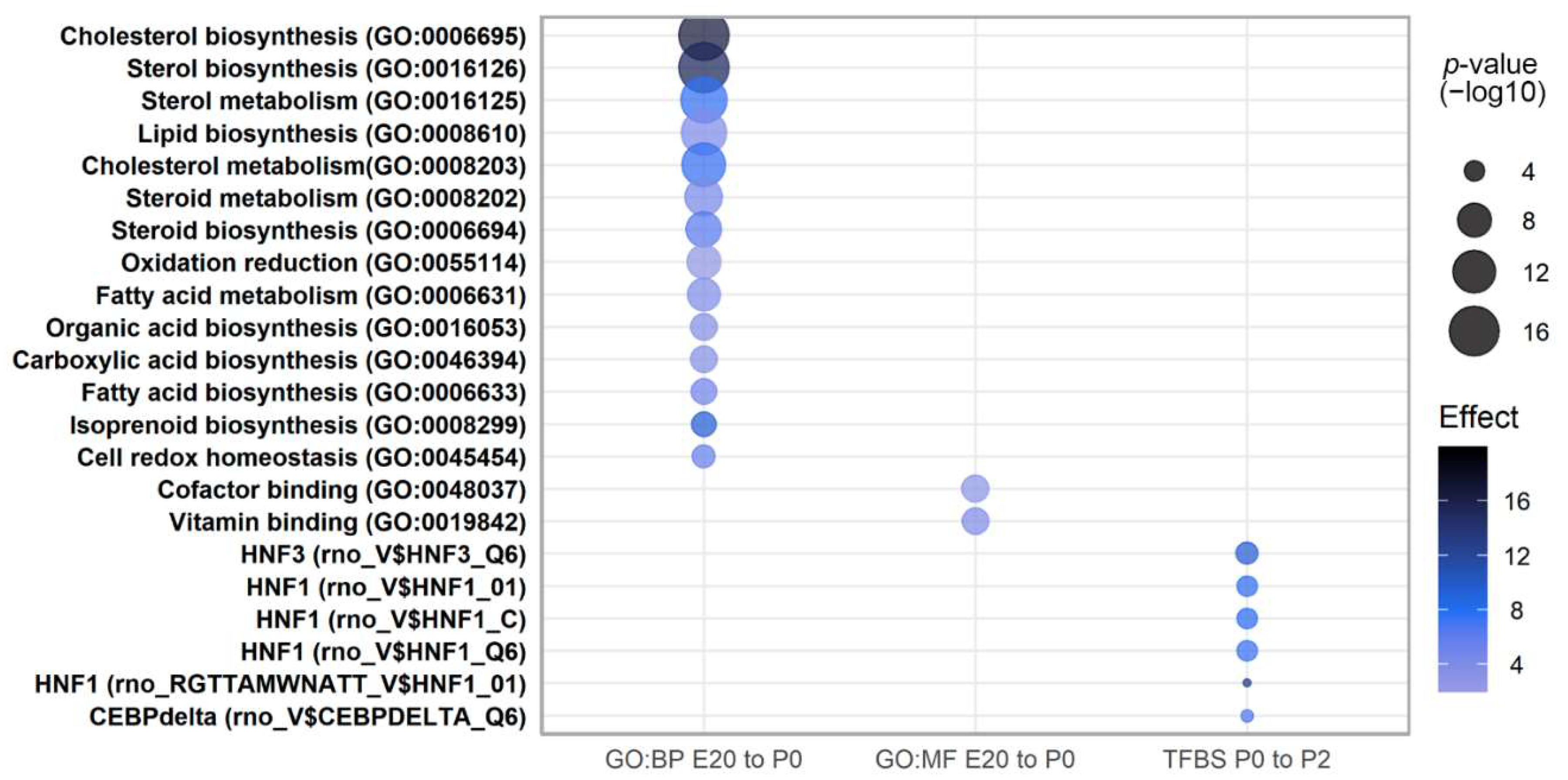
2.4. Pathway Analysis of Perinatally Regulated Transcripts
2.5. Characterization of the Low-Protein Malnutrition Model
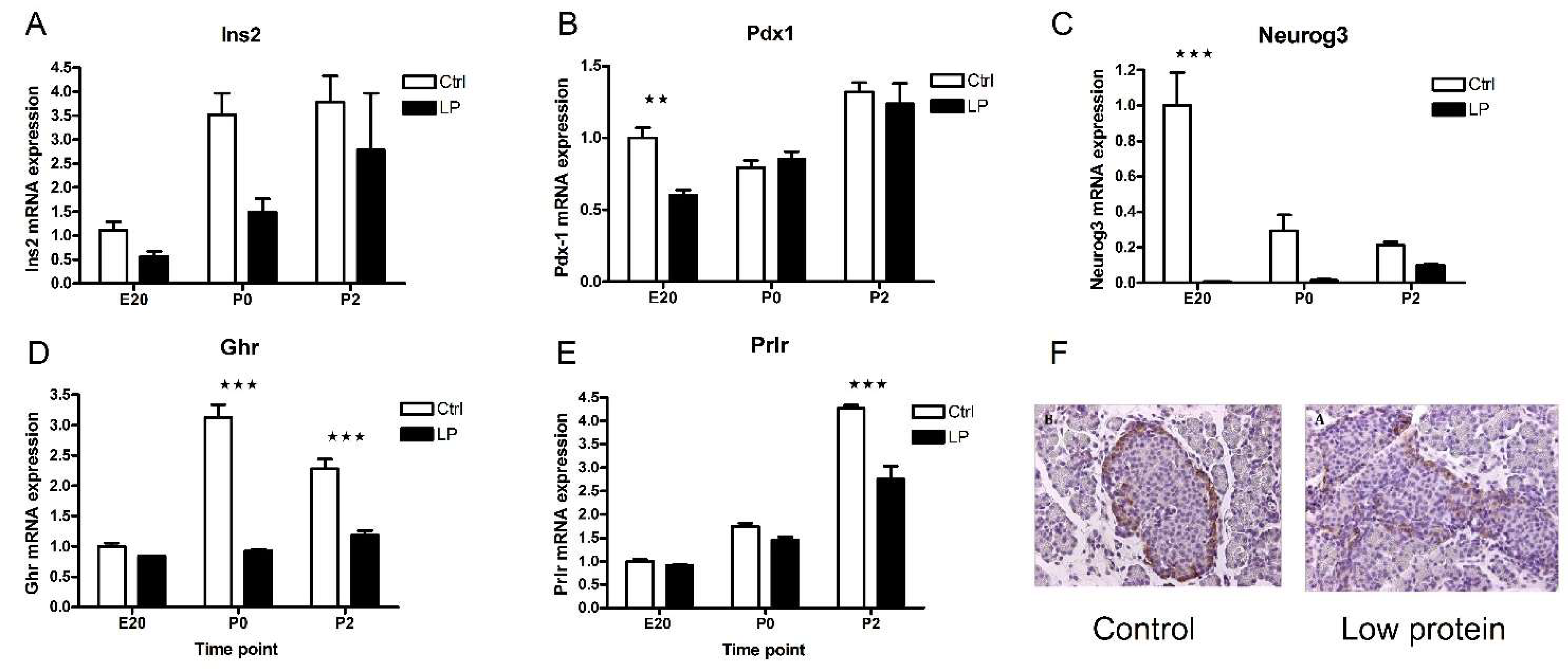
2.6. Transcripts Regulated by Maternal Low-Protein Diet
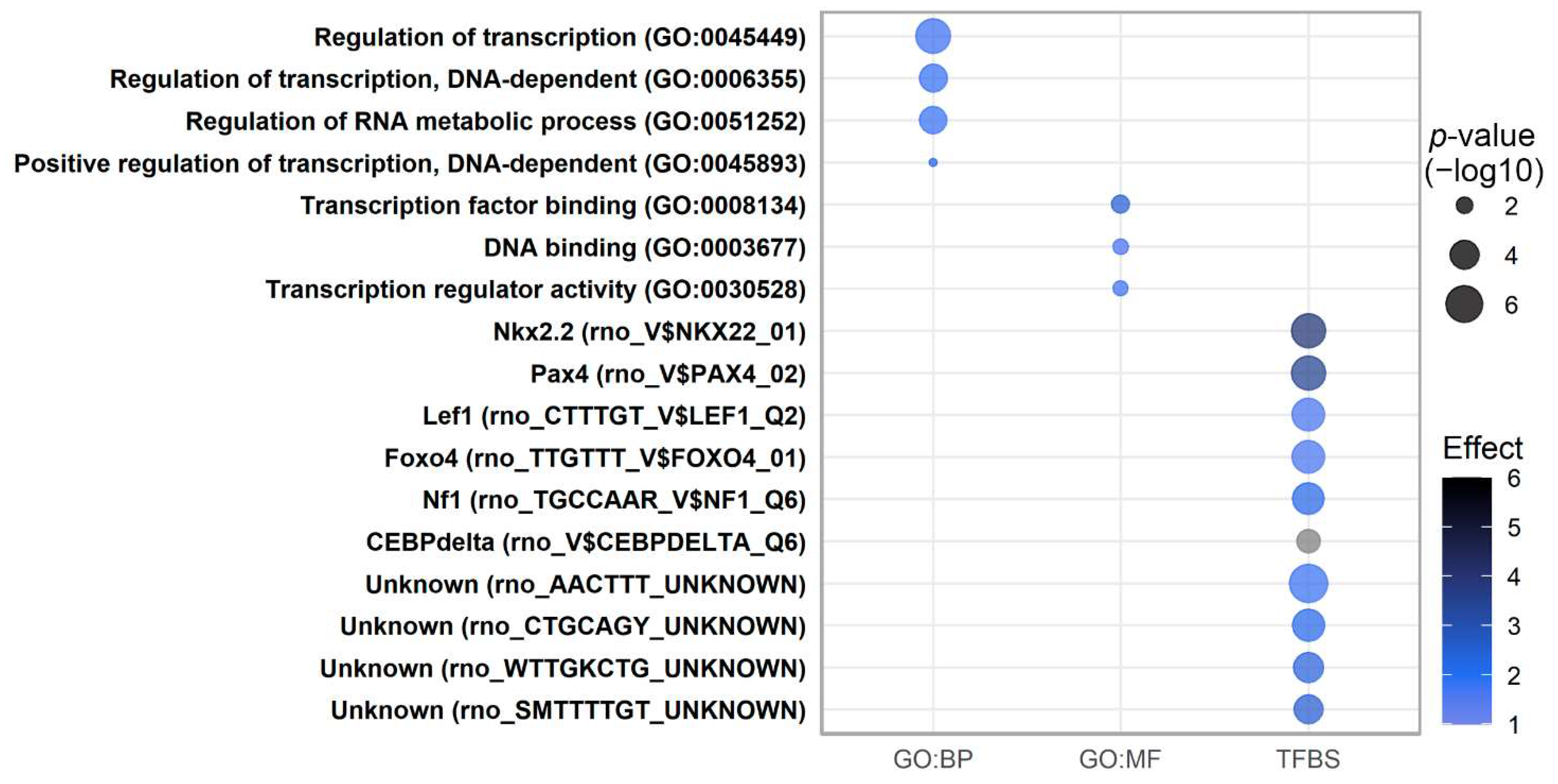
2.7. Pathway Analysis of Transcripts Regulated by Low-Protein Diet
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bouret, S.; Levin, B.E.; Ozanne, S.E. Gene-Environment Interactions Controlling Energy and Glucose Homeostasis and the Developmental Origins of Obesity. Physiol. Rev. 2015, 95, 47–82. [Google Scholar] [CrossRef] [PubMed]
- Weir, G.C.; Gaglia, J.; Bonner-Weir, S. Inadequate β-cell mass is essential for the pathogenesis of type 2 diabetes. Lancet Diabetes Endocrinol. 2020, 8, 249–256. [Google Scholar] [CrossRef]
- Nielsen, J.H.; Haase, T.N.; Jaksch, C.; Nalla, A.; Søstrup, B.; Nalla, A.A.; Larsen, L.; Rasmussen, M.; Dalgaard, L.; Gaarn, L.W.; et al. Impact of fetal and neonatal environment on beta cell function and development of diabetes. Acta Obstet. Gynecol. Scand. 2014, 93, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Gradwohl, G.; Dierich, A.; LeMeur, M.; Guillemot, F. neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas. Proc. Natl. Acad. Sci. USA 2000, 97, 1607–1611. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, M.C.; Ahnfelt-Ronne, J.; Hald, J.; Madsen, O.D.; Serup, P.; Hecksher-Sorensen, J. An Illustrated Review of Early Pancreas Development in the Mouse. Endocr. Rev. 2007, 28, 685–705. [Google Scholar] [CrossRef]
- Horn, S.; Kobberup, S.; Jørgensen, M.C.; Kalisz, M.; Klein, T.; Kageyama, R.; Gegg, M.; Lickert, H.; Lindner, J.; Magnuson, M.A.; et al. Mind bomb 1 is required for pancreatic β-cell formation. Proc. Natl. Acad. Sci. USA 2012, 109, 7356–7361. [Google Scholar] [CrossRef]
- Bouwens, L.; Rooman, I. Regulation of pancreatic beta-cell mass. Physiol. Rev. 2005, 85, 1255–1270. [Google Scholar] [CrossRef]
- Dhawan, S.; Georgia, S.; Bhushan, A. Formation and regeneration of the endocrine pancreas. Curr. Opin. Cell Biol. 2007, 19, 634–645. [Google Scholar] [CrossRef]
- Dor, Y.; Brown, J.; Martinez, O.I.; Melton, D.A. Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation. Nature 2004, 429, 41–46. [Google Scholar] [CrossRef]
- Foa, P.P.; Blázquez, E.; Sodoyez, J.C.; Sodoyez-Goffaux, F. The Ontogeny of Mammalian Insular Function. In The Evolution of Pancreatic Islets; Pergamon Press: London, UK, 1976. [Google Scholar]
- Aye, T.; Toschi, E.; Sharma, A.; Sgroi, D.; Bonner-Weir, S. Identification of markers for newly formed beta-cells in the perinatal period: A time of recognized beta-cell immaturity. J. Histochem. Cytochem. 2010, 58, 369–376. [Google Scholar] [CrossRef]
- Hellerstrom, C. The life story of the pancreatic B cell. Diabetologia 1984, 26, 393–400. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, R.C.; Madson, K.L. Pancreatic Insulin-, Glucagon-, and Somatostatin-Positive Islet Cell Populations during the Perinatal Development of the Rat. II. Changes in hormone content and concentration. Biol. Neonatol. 1980, 38, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Hellerstrom, C.; Swenne, I. Functional maturation and proliferation of fetal pancreatic beta-cells. Diabetes 1991, 40 (Suppl. 2), 89–93. [Google Scholar] [CrossRef] [PubMed]
- Martens, G.; Motte, E.; Kramer, G.; Stange, G.; Gaarn, L.W.; Hellemans, K.; Nielsen, J.H.; Aerts, J.; Ling, Z.; Pipeleers, D. Functional characteristics of neonatal rat β cells with distinct markers. J. Mol. Endocrinol. 2013, 52, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, O.; Pages, G.; Gimond, C. The dual-specificity MAP kinase phosphatases: Critical roles in development and cancer. Am. J. Physiol. Cell Physiol. 2010, 299, C189–C202. [Google Scholar] [CrossRef]
- Yamada, T.; Ozaki, N.; Kato, Y.; Miura, Y.; Oiso, Y. Insulin downregulates angiopoietin-like protein 4 mRNA in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2006, 347, 1138–1144. [Google Scholar] [CrossRef]
- Yi, P.; Park, J.-S.; Melton, D.A. RETRACTED: Betatrophin: A Hormone that Controls Pancreatic β Cell Proliferation. Cell 2013, 153, 747–758. [Google Scholar] [CrossRef]
- Møldrup, A.; Petersen, E.D.; Nielsen, J.H. Effects of sex and pregnancy hormones on growth hormone and prolactin receptor gene expression in insulin-producing cells. Endocrinology 1993, 133, 1165–1172. [Google Scholar] [CrossRef]
- Dolva, L.; Nielsen, J.H.; Welinder, B.S.; Hanssen, K.F. Biosynthesis and release of thyrotropin-releasing hormone immunoreactivity in rat pancreatic islets in organ culture. Effects of age, glucose, and streptozotocin. J. Clin. Investig. 1983, 72, 1867–1873. [Google Scholar] [CrossRef]
- Štrbák, V. Pancreatic Thyrotropin Releasing Hormone and Mechanism of Insulin Secretion. Cell. Physiol. Biochem. 2018, 50, 378–384. [Google Scholar] [CrossRef]
- Demarchi, F.; Verardo, R.; Varnum, B.; Brancolini, C.; Schneider, C. Gas6 anti-apoptotic signaling requires NF-kappa B activation. J. Biol. Chem. 2001, 276, 31738–31744. [Google Scholar] [CrossRef] [PubMed]
- Couchie, D.; Lafdil, F.; Martin-Garcia, N.; Laperche, Y.; Zafrani, E.S.; Mavier, P. Expression and role of Gas6 protein and of its receptor Axl in hepatic regeneration from oval cells in the rat. Gastroenterology 2005, 129, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Araki, H.; Sugimoto, T.; Takeuchi, K.; Yamane, T.; Maeda, T.; Yamamoto, Y.; Nishi, K.; Asano, M.; Shirahama-Noda, K.; et al. Legumain/asparaginyl endopeptidase controls extracellular matrix remodeling through the degradation of fibronectin in mouse renal proximal tubular cells. FEBS Lett. 2007, 581, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, C.; Huang, H.; Janda, K.; Edgington, T. Overexpression of legumain in tumors is significant for invasion/metastasis and a candidate enzymatic target for prodrug therapy. Cancer Res. 2003, 63, 2957–2964. [Google Scholar] [PubMed]
- Choi, J.-H.; Lee, M.-Y.; Kim, Y.; Shim, J.-Y.; Han, S.-M.; Lee, K.-A.; Choi, Y.-K.; Jeon, H.-M.; Baek, K.-H. Isolation of genes involved in pancreas regeneration by subtractive hybridization. Biol. Chem. 2010, 391, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Ekman, G.C.; Garcia, T.; Carnes, K.; Zhang, Z.; Murphy, T.; Murphy, K.M.; Hess, R.A.; Cooke, P.S.; Hofmann, M.-C. ETV5 Regulates Sertoli Cell Chemokines Involved in Mouse Stem/Progenitor Spermatogonia Maintenance. Stem Cells 2010, 28, 1882–1892. [Google Scholar] [CrossRef]
- Kobberup, S.; Nyeng, P.; Juhl, K.; Hutton, J.; Jensen, J. ETS-family genes in pancreatic development. Dev. Dyn. 2007, 236, 3100–3110. [Google Scholar] [CrossRef]
- Haase, T.N.; Rasmussen, M.; Jaksch, C.A.M.; Gaarn, L.W.; Petersen, C.K.; Billestrup, N.; Nielsen, J.H. Growth arrest specific protein (GAS) 6: A role in the regulation of proliferation and functional capacity of the perinatal rat beta cell. Diabetologia 2013, 56, 763–773. [Google Scholar] [CrossRef]
- Winkel, L.; Bagge, A.; Larsen, L.; Haase, T.N.; Rasmussen, M.; Lykke, J.; Holmgaard, D.B.; Thim, L.; Nielsen, J.H.; Dalgaard, L.T. Trefoil factor 3 in perinatal pancreas is increased by gestational low protein diet and associated with accelerated β-cell maturation. Islets 2018, 10, e1472186-25. [Google Scholar] [CrossRef]
- Jackerott, M.; Lee, Y.C.; Møllgård, K.; Kofod, H.; Jensen, J.; Rohleder, S.; Neubauer, N.; Gaarn, L.W.; Lykke, J.; Dodge, R.; et al. Trefoil Factors Are Expressed in Human and Rat Endocrine Pancreas: Differential Regulation by Growth Hormone. Endocrinology 2006, 147, 5752–5759. [Google Scholar] [CrossRef][Green Version]
- Yang, Y.; Lin, Z.; Lin, Q.; Bei, W.; Guo, J. Pathological and therapeutic roles of bioactive peptide trefoil factor 3 in diverse diseases: Recent progress and perspective. Cell Death Dis. 2022, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Fang, Z.; Yu, X.; Zhang, M. Transcription factors involved in glucose-stimulated insulin secretion of pancreatic beta cells. Biochem. Biophys. Res. Commun. 2009, 384, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Iwasaki, Y.; Yatoh, S.; Toyonori, K.; Shin, K.; Noriyuki, I.; Takashi, Y.; Takashi, M.; Yoshimi, N.; Naoya, Y.; et al. Cholesterol accumulation and diabetes in pancreatic beta-cell-specific SREBP-2 transgenic mice: A new model for lipotoxicity. J. Lipid Res. 2008, 49, 2524–2534. [Google Scholar] [CrossRef]
- Reusens, B.; Theys, N.; Dumortier, O.; Goosse, K.; Remacle, C. Maternal malnutrition programs the endocrine pancreas in progeny. Am. J. Clin. Nutr. 2011, 94 (Suppl. 6), 1824S–1829S. [Google Scholar] [CrossRef]
- Søstrup, B.; Gaarn, L.W.; Nalla, A.; Billestrup, N.; Nielsen, J.H. Co-ordinated regulation of neurogenin-3 expression in the maternal and fetal pancreas during pregnancy. Acta Obstet. Gynecol. Scand. 2014, 93, 1190–1197. [Google Scholar] [CrossRef]
- Golden, T.N.; Simmons, R.A. Immune dysfunction in developmental programming of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 235–245. [Google Scholar] [CrossRef]
- Bruun, C.; Christensen, G.L.; Jacobsen, M.L.B.; Kanstrup, M.B.; Jensen, P.R.; Fjordvang, H.; Mandrup-Poulsen, T.; Billestrup, N. Inhibition of beta cell growth and function by bone morphogenetic proteins. Diabetologia 2014, 57, 2546–2554. [Google Scholar] [CrossRef]
- Dumartin, L.; Whiteman, H.J.; Weeks, M.E.; Hariharan, D.; Dmitrovic, B.; Iacobuzio-Donahue, C.A.; Brentnall, T.A.; Bronner, M.P.; Feakins, R.M.; Timms, J.F.; et al. AGR2 Is a Novel Surface Antigen That Promotes the Dissemination of Pancreatic Cancer Cells through Regulation of Cathepsins B and D. Cancer Res. 2011, 71, 7091–7102. [Google Scholar] [CrossRef]
- Park, S.-W.; Zhen, G.; Verhaeghe, C.; Nakagami, Y.; Nguyenvu, L.T.; Barczak, A.J.; Killeen, N.; Erle, D.J. The protein disulfide isomerase AGR2 is essential for production of intestinal mucus. Proc. Natl. Acad. Sci. USA 2009, 106, 6950–6955. [Google Scholar] [CrossRef]
- Kinsey, C.; Balakrishnan, V.; O’Dell, M.R.; Huang, J.L.; Newman, L.; Whitney-Miller, C.L.; Hezel, A.F.; Land, H. Plac8 Links Oncogenic Mutations to Regulation of Autophagy and Is Critical to Pancreatic Cancer Progression. Cell Rep. 2014, 7, 1143–1155. [Google Scholar] [CrossRef]
- Conrad, E.; Stein, R.; Hunter, C.S. Revealing transcription factors during human pancreatic β cell development. Trends Endocrinol. Metab. 2014, 25, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Swenne, I.; Eriksson, U. Diabetes in pregnancy: Islet cell proliferation in the fetal rat pancreas. Diabetologia 1982, 23, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Diraison, F.; Ravier, M.A.; Richards, S.K.; Smith, R.M.; Shimano, H.; Rutter, G.A. SREBP1 is required for the induction by glucose of pancreatic beta-cell genes involved in glucose sensing. J. Lipid Res. 2008, 49, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.; Rosenstierne, M.W.; Gaarn, L.W.; Bagge, A.; Pedersen, L.; Dahmcke, C.M.; Nielsen, J.H.; Dalgaard, L.T. Expression and Localization of microRNAs in Perinatal Rat Pancreas: Role of miR-21 in Regulation of Cholesterol Metabolism. PLoS ONE 2011, 6, e25997. [Google Scholar] [CrossRef] [PubMed]
- Freie, H.M.; Pasma, A.; Bouman, P.R. Quantitative analysis of pancreatic islet development and insulin storage in the foetal and newborn rat. Acta Endocrinol. 1975, 80, 657–666. [Google Scholar] [CrossRef]
- Chen, Z.; Downing, S.; Tzanakakis, E.S. Four Decades After the Discovery of Regenerating Islet-Derived (Reg) Proteins: Current Understanding and Challenges. Front. Cell Dev. Biol. 2019, 7, 235. [Google Scholar] [CrossRef]
- Okamoto, H. The Reg gene family and Reg proteins: With special attention to the regeneration of pancreatic beta-cells. J. Hepatobiliary-Pancreat. Surg. 1999, 6, 254–262. [Google Scholar] [CrossRef]
- Sala, P.; Torrinhas, R.S.; Fonseca, D.C.; Heymsfield, S.; Giannella-Neto, D.; Waitzberg, D.L. Type 2 Diabetes Remission After Roux-en-Y Gastric Bypass: Evidence for Increased Expression of Jejunal Genes Encoding Regenerating Pancreatic Islet-Derived Proteins as a Potential Mechanism. Obes. Surg. 2017, 27, 1123–1127. [Google Scholar] [CrossRef]
- Rezania, A.; Bruin, J.E.; Arora, P.; Rubin, A.; Batushansky, I.; Asadi, A.; O’Dwyer, S.; Quiskamp, N.; Mojibian, M.; Albrecht, T.; et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 2014, 32, 1121–1133. [Google Scholar] [CrossRef]
- Zhang, L.; Theise, N.; Chua, M.; Reid, L.M. The stem cell niche of human livers: Symmetry between development and regeneration. Hepatology 2008, 48, 1598–1607. [Google Scholar] [CrossRef]
- Jelnes, P.; Santoni-Rugiu, E.; Rasmussen, M.; Friis, S.L.; Nielsen, J.H.; Tygstrup, N.; Bisgaard, H.C. Remarkable heterogeneity displayed by oval cells in rat and mouse models of stem cell–mediated liver regeneration. Hepatology 2007, 45, 1462–1470. [Google Scholar] [CrossRef]
- Mizejewski, G.J. Biological roles of alpha-fetoprotein during pregnancy and perinatal development. Exp. Biol. Med. 2004, 229, 439–463. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guo, J.; Yuan, L.; Cheng, M.; Cao, L.; Shi, H.; Tong, H.; Wang, N.; De, W. Alpha-fetoprotein is dynamically expressed in rat pancreas during development. Dev. Growth Differ. 2007, 49, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, G.; Jabs, N.; Konstantinova, I.; Domogatskaya, A.; Tryggvason, K.; Sorokin, L.; Fässler, R.; Gu, G.; Gerber, H.-P.; Ferrara, N.; et al. The vascular basement membrane: A niche for insulin gene expression and Beta cell proliferation. Dev. Cell 2006, 10, 397–405. [Google Scholar] [CrossRef]
- Eberhard, D.; Kragl, M.; Lammert, E. ‘Giving and taking’: Endothelial and beta-cells in the islets of Langerhans. Trends Endocrinol. Metab. 2010, 21, 457–463. [Google Scholar] [CrossRef]
- Schumacher, A.; Costa, S.-D.; Zenclussen, A.C. Endocrine Factors Modulating Immune Responses in Pregnancy. Front. Immunol. 2014, 5, 196. [Google Scholar] [CrossRef]
- Tian, L.; Zhou, J.; Casimiro, M.C.; Pestell, R.G.; Wang, C. Activating peroxisome proliferator-activated receptor gamma mutant promotes tumor growth in vivo by enhancing angiogenesis. Cancer Res. 2009, 69, 9236–9244. [Google Scholar] [CrossRef]
- Chen, J.M.; Fortunato, M.; Stevens, R.A.; Barrett, A.J. Activation of progelatinase A by mammalian legumain, a recently discovered cysteine proteinase. Biol. Chem. 2001, 382, 777–783. [Google Scholar] [CrossRef]
- Gutierrez-Aguilar, R.; Kim, D.-H.; Casimir, M.; Dai, X.-Q.; Pfluger, P.; Park, J.; Haller, A.; Donelan, E.; Park, J.; D’Alessio, D.; et al. The role of the transcription factor ETV5 in insulin exocytosis. Diabetologia 2013, 57, 383–391. [Google Scholar] [CrossRef]
- Planaguma, J.; Liljestrom, M.; Alameda, F.; Bützow, R.; Virtanen, I.; Reventós, J.; Hukkanen, M. Matrix metalloproteinase-2 and matrix metalloproteinase-9 codistribute with transcription factors RUNX1/AML1 and ETV5/ERM at the invasive front of endometrial and ovarian carcinoma. Hum. Pathol. 2011, 42, 57–67. [Google Scholar] [CrossRef]
- Miralles, F.; Battelino, T.; Czernichow, P.; Scharfmann, R. TGF-beta plays a key role in morphogenesis of the pancreatic islets of Langerhans by controling the activity of the matrix metalloproteinase MMP-2. J. Cell Biol. 1998, 143, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, P.; Huotari, M.; Koivisto, T.; Ustinov, J.; Palgi, J.; Rasilainen, S.; Lehtonen, E.; Keski-Oja, J.; Otonkoski, T. Impaired migration and delayed differentiation of pancreatic islet cells in mice lacking EGF-receptors. Development 2000, 127, 2617–2627. [Google Scholar] [CrossRef] [PubMed]
- Stetler-Stevenson, W.G.; Aznavoorian, S.; Liotta, L.A. Tumor cell interactions with the extracellular matrix during invasion and metastasis. Annu. Rev. Cell Biol. 1993, 9, 541–573. [Google Scholar] [CrossRef] [PubMed]
- Liotta, L.A.; Tryggvason, K.; Garbisa, S.; Hart, I.; Foltz, C.M.; Shafie, S. Metastatic potential correlates with enzymatic degradation of basement membrane collagen. Nature 1980, 284, 67–68. [Google Scholar] [CrossRef]
- Miller, K.; Kim, A.; Kilimnik, G.; Jo, J.; Moka, U.; Periwal, V.; Hara, M. Islet Formation during the Neonatal Development in Mice. PLoS ONE 2009, 4, e7739. [Google Scholar] [CrossRef]
- Friedrichsen, B.N.; Carlsson, C.; Moldrup, A.; Michelsen, B.; Jensen, C.H.; Teisner, B.; Nielsen, J.H. Expression, biosynthesis and release of preadipocyte factor-1/ delta-like protein/fetal antigen-1 in pancreatic beta-cells: Possible physiological implications. J. Endocrinol. 2003, 176, 257–266. [Google Scholar] [CrossRef]
- Pagliuca, F.W.; Millman, J.R.; Gürtler, M.; Segel, M.; Van Dervort, A.; Ryu, J.H.; Peterson, Q.P.; Greiner, D.; Melton, D.A. Generation of Functional Human Pancreatic β Cells In Vitro. Cell 2014, 159, 428–439. [Google Scholar] [CrossRef]
- Simon, R.; Lam, A.; Li, M.C.; Ngan, M.; Menenzes, S.; Zhao, Y. Analysis of gene expression data using BRB-ArrayTools. Cancer Inform. 2007, 3, 11–17. [Google Scholar] [CrossRef]
- Korn, E.L.; Troendle, J.F.; McShane, L.M.; Simon, R. Controling the number of false discoveries: Application to high-dimensional genomic data. J. Stat. Plan. Inference 2004, 124, 379–398. [Google Scholar] [CrossRef]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef]
- Li, C.; Wong, W.H. Model-based analysis of oligonucleotide arrays: Expression index computation and outlier detection. Proc. Natl. Acad. Sci. USA 2001, 98, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, 3. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Gerds, T.A.; Nielsen, O.H.; Seidelin, J.B.; Troelsen, J.T.; Olsen, J. pcaGoPromoter—An R Package for Biological and Regulatory Interpretation of Principal Components in Genome-Wide Gene Expression Data. PLoS ONE 2012, 7, e32394. [Google Scholar] [CrossRef]
- Sandelin, A.; Alkema, W.; Engstrom, P.; Wasserman, W.W.; Lenhard, B. JASPAR: An open-access database for eukaryotic transcription factor binding profiles. Nucl. Acids Res. 2004, 32, D91–D94. [Google Scholar] [CrossRef]
- Matys, V. TRANSFAC(R) and its module TRANSCompel(R): Transcriptional gene regulation in eukaryotes. Nucl. Acids Res. 2006, 34, D108–D110. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Increased at P0 vs. E20 | Increased at P2 vs. P0 | ||||||
|---|---|---|---|---|---|---|---|
| Gene Symbol | Gene Name | FC CO | FC LP | Gene Symbol | Gene Name | FC CO | FC LP |
| Reg3a/3b | Regenerating family member 3 alpha/beta | 461/335 | 617/617 | LOC688750 | CD209 antigen | 5.1 | 1.8 |
| Hmgcs2 | 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 2 | 270 | 55 | Prss35 | Protease, serine, 35 | 4.9 | 1.2 |
| LOC286960 | Preprotrypsinogen IV | 155 | 103 | C5 | Complement C5 | 4.8 | −1.3 |
| LOC312273 | Trypsin V-A | 145 | 151 | LOC500183 | NGF-binding Ig light chain | 4.7 | 1.3 |
| Gif | Gastric intrinsic factor | 61 | 61 | C4a | Complement component 4a | 3.6 | 1.6 |
| Alb | Albumin | 58 | 12 | LOC365985 | Adenylate kinase 5 isoform 1 | 3.4 | 1.6 |
| Angptl4 | Angiopoietin-like 4 | 54 | 18 | Tinag | Tubulointerstitial nephritis antigen | 3.1 | −1.5 |
| Apoa1 | Apolipoprotein A1 | 39 | 117 | Spink3 | Serine protease inhibitor, Kazal type 3 | 2.9 | 1.7 |
| Try10 | Pancreatic trypsin 1 | 30 | 15 | LOC686268 | SUMO/sentrin specific protease 5 | 2.9 | 1.4 |
| Ahsg | Alpha-2-HS-glycoprotein | 29 | 19 | Cuzd1 | CUB and zona pellucida-like domains 1 | 2.9 | 1.1 |
| Spink1 | Serine protease inhibitor, Kazal type 1 | 29 | 11 | Egfl6 | EGF-like-domain, multiple 6 | 2.9 | 2.0 |
| Kng1 | Kininogen 1 | 28 | 23 | Ak7 | Adenylate kinase 7 | 2.8 | −1.1 |
| Gas6 | Growth arrest specific 6 | 28 | 18 | Nradd | Neurotrophin receptor associated death domain | 2.8 | 1.3 |
| Apob | Apolipoprotein B | 25 | 84 | Ret | Ret proto-oncogene | 2.7 | −1.0 |
| Slc18a2 | Solute carrier family 18 (vesicular monoamine), member 2 | 23 | 5.3 | Zcchc12 | Zinc finger, CCHC domain containing 12 | 2.6 | −1.1 |
| Decreased at P0 vs. E20 | Decreased at P2 vs. P0 | ||||||
| Gene Symbol | Gene Name | FC CO | FC LP | Gene Symbol | Gene Name | FC CO | FC LP |
| Serpina6 | Serpin family A member 6 | −47 | −74 | Ahsg | Alpha-2-HS-glycoprotein | −31 | −1.4 |
| Hbe1 | Hemoglobin subunit epsilon 1 | −44 | −30 | Fga | Fibrinogen, alpha polypeptide | −23 | −1.2 |
| Tinag | Tubulointerstitial nephritis antigen | −30 | −7.6 | Fabp1 | Fatty acid binding protein 1 | −23 | −1.3 |
| Tnni | Troponin I, skeletal, slow | −28 | −23 | Kng1 | Kininogen 1 | −21 | −1.6 |
| Hbg1 | Hemoglobin, gamma A | −27 | −19 | Apoc2 | Apolipoprotein C-II | −19 | −4.6 |
| Hdc | Histidine decarboxylase | −24 | −17 | Fgg | Fibrinogen, gamma polypeptide | −17 | 2.0 |
| Ptges | Prostaglandin E synthase | −22 | −14 | Apoc1 | Apolipoprotein C-I | −16 | −1.2 |
| Tm7sf2 | Transmembrane 7 superfamily member 2 | −17 | −4.1 | Apoh | Apolipoprotein H | −15 | −1.0 |
| Pln | Phospholamban | −17 | −4.2 | Fgb | Fibrinogen, beta polypeptide | −13 | −1.1 |
| Nags | N-acetylglutamate synthase | −17 | −3.2 | Apoa1 | Apolipoprotein A-I | −13 | −5.4 |
| Clic3 | Chloride intracellular channel 3 | −13 | −7.1 | Hpx | Hemopexin | −13 | −1.2 |
| Adam2 | A disintegrin and metalloprotease domain 2 | −12 | −9.8 | LOC299282 | Serine protease inhibitor | −12 | −1.3 |
| Fdft1 | Farnesyl diphosphate farnesyl transferase 1 | −12 | −4.4 | Itih3 | Inter-alpha trypsin inhibitor, heavy chain 3 | −10 | −1.2 |
| LOC682690 | Chromodomain helicase DNA binding protein 9 | −11 | 1.2 | Pck1 | Phosphoenolpyruvate carboxykinase 1 | −9.9 | −5.3 |
| Camkk2 | Calcium/calmodulin-dependent protein kinase kinase 2, beta | −11 | −2.4 | Serpina3k | Serine peptidase inhibitor, clade A, member 3K | −9.7 | −1.2 |
| Gene Symbol | Gene Name | E20 | Gene Symbol | Gene Name | P0 | Gene Symbol | Gene Name | P2 |
|---|---|---|---|---|---|---|---|---|
| Increased | LP vs. Control (Fold Regulation) | Increased | LP vs. Control (Fold Regulation) | Increased | LP vs. Control (Fold Regulation) | |||
| Agtr2 | Angiotensin II receptor, type 2 | 11 | Apoa4 | Apolipoprotein A4 | 142 | Lgals4 | Lectin, galactose binding, soluble 4 | 236 |
| LOC686892 | Muscleblind-like 1 isoform d | 8.7 | Pga5 | Pepsinogen 5 | 112 | Pga5 | Pepsinogen 5 | 215 |
| Cav | Caveolin | 8.6 | Rbp2 | Retinol binding protein 2 | 86 | Agr2 | Anterior gradient 2 | 110 |
| Ppp3r1 | Calcineurin B, type I | 8.5 | Lgals4 | Lectin, galactose binding, soluble 4 | 79 | Retnla | Resistin like alpha | 93 |
| Zfp260 | Zinc finger protein 260 | 7.7 | Tff1 | Trefoil factor 1 | 73 | Fabp1 | Fatty acid binding protein 1 | 92 |
| Sept2 | Septin 2 | 7.6 | Clca3 | Chloride channel calcium activated 3 | 55 | Tff1 | Trefoil factor 1 | 87 |
| Ogn | Osteoglycin | 7.4 | Agr2 | Anterior gradient 2 | 48 | Gkn1 | Gastrokine 1 | 82 |
| Mat2a | Methionine adenosyltransferase II, alpha | 6.3 | Fabp2 | Fatty acid binding protein 2 | 46 | LOC56825 | Prochymosin | 66 |
| Il13ra1 | Interleukin 13 receptor, alpha 1 | 5.9 | Clca6 | Chloride channel calcium activated 6 | 45 | Clca3 | Chloride channel calcium activated 3 | 42 |
| LOC498358 | Solute carrier family 30 (zinc transporte), member 9 | 5.6 | Gkn1 | Gastrokine 1 | 42 | Sult1b1 | Sulfotransferase family 1B | 39 |
| Decreased | LP vs. Control (Fold Regulation) | E20 | Decreased | LP vs. Control (Fold Regulation) | P0 | Decreased | LP vs. Control (Fold Regulation) | P2 |
| Fos | FBJ murine osteosarcoma viral oncogene | −14 | Myo5c | Myosin Vc | −5.1 | Atp8b1 | ATPase, Class I, type 8B, member 1 | −11 |
| LOC680231 | Chromodomain helicase DNA binding protein 9 | −12 | Phlda1 | Chromodomain helicase DNA binding protein 9 | −4.3 | Aff4 | AF4/FMR2 family, member 4 | −9.9 |
| Zfhx1b | Zinc finger homeobox 1b (ZEB2) | −6.8 | Gtl2 | GTL2, imprinted maternally expressed untranslated | −4.2 | Foxo1a | Forkhead box O1A | −9.6 |
| Atxn2 | Ataxin 2 | −6.7 | Smoc1 | SPARC-related modular calcium binding protein 1 | −4.1 | Eif2c2 | Eukaryotic translation initiation factor 2C, 2 | −9.4 |
| RGD1561386 | CBL E3 ubiquitin protein ligase | −6.4 | LOC682488 | Ras-related protein Rab-1B | −4.1 | Tns | Tensin | −9.0 |
| Adipor2 | Adiponectin receptor 2 | −6.1 | Lamc1 | Laminin, gamma 1 | −3.9 | Akap9 | A kinase (PRKA) anchor protein 9 | −8.9 |
| Eif2c2 | Eukaryotic translation initiation factor 2C, 2 | −5.8 | Adhfe1 | Alcohol dehydrogenase, iron containing, 1 | −3.9 | Ash1l | Absent, small, or homeotic)-like | −8.8 |
| Mt1a | Metallothionein 1a | −5.8 | Fgfr1 | Fibroblast growth factor receptor 1 | −3.8 | Rck | DEAD box protein rck/p54 | −7.4 |
| Tbl1x | Transducin (beta)-like 1 X-linked | −5.8 | P34 | P34 protein | −3.7 | Tbl1x | Transducin (beta)-like 1 X-linked | −7.4 |
| Atp8b1 | ATPase, Class I, type 8B, member 1 | −5.4 | Ccnl2 | Cyclin L2 | −3.5 | Ubn1 | Ubinuclein 1 | −6.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winkel, L.; Rasmussen, M.; Larsen, L.; Dalgaard, L.T.; Nielsen, J.H. Fetal Programming of the Endocrine Pancreas: Impact of a Maternal Low-Protein Diet on Gene Expression in the Perinatal Rat Pancreas. Int. J. Mol. Sci. 2022, 23, 11057. https://doi.org/10.3390/ijms231911057
Winkel L, Rasmussen M, Larsen L, Dalgaard LT, Nielsen JH. Fetal Programming of the Endocrine Pancreas: Impact of a Maternal Low-Protein Diet on Gene Expression in the Perinatal Rat Pancreas. International Journal of Molecular Sciences. 2022; 23(19):11057. https://doi.org/10.3390/ijms231911057
Chicago/Turabian StyleWinkel, Louise, Morten Rasmussen, Louise Larsen, Louise T. Dalgaard, and Jens H. Nielsen. 2022. "Fetal Programming of the Endocrine Pancreas: Impact of a Maternal Low-Protein Diet on Gene Expression in the Perinatal Rat Pancreas" International Journal of Molecular Sciences 23, no. 19: 11057. https://doi.org/10.3390/ijms231911057
APA StyleWinkel, L., Rasmussen, M., Larsen, L., Dalgaard, L. T., & Nielsen, J. H. (2022). Fetal Programming of the Endocrine Pancreas: Impact of a Maternal Low-Protein Diet on Gene Expression in the Perinatal Rat Pancreas. International Journal of Molecular Sciences, 23(19), 11057. https://doi.org/10.3390/ijms231911057