


Adaptive Responses of the Sea Anemone Heteractis crispa to the Interaction of Acidification and Global Warming


{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions, Acclimation, and Treatments
2.2. Tissue Collection and Physiological Measurements
2.3. Sterol Extraction and Analysis
2.4. Statistical Analysis
3. Results
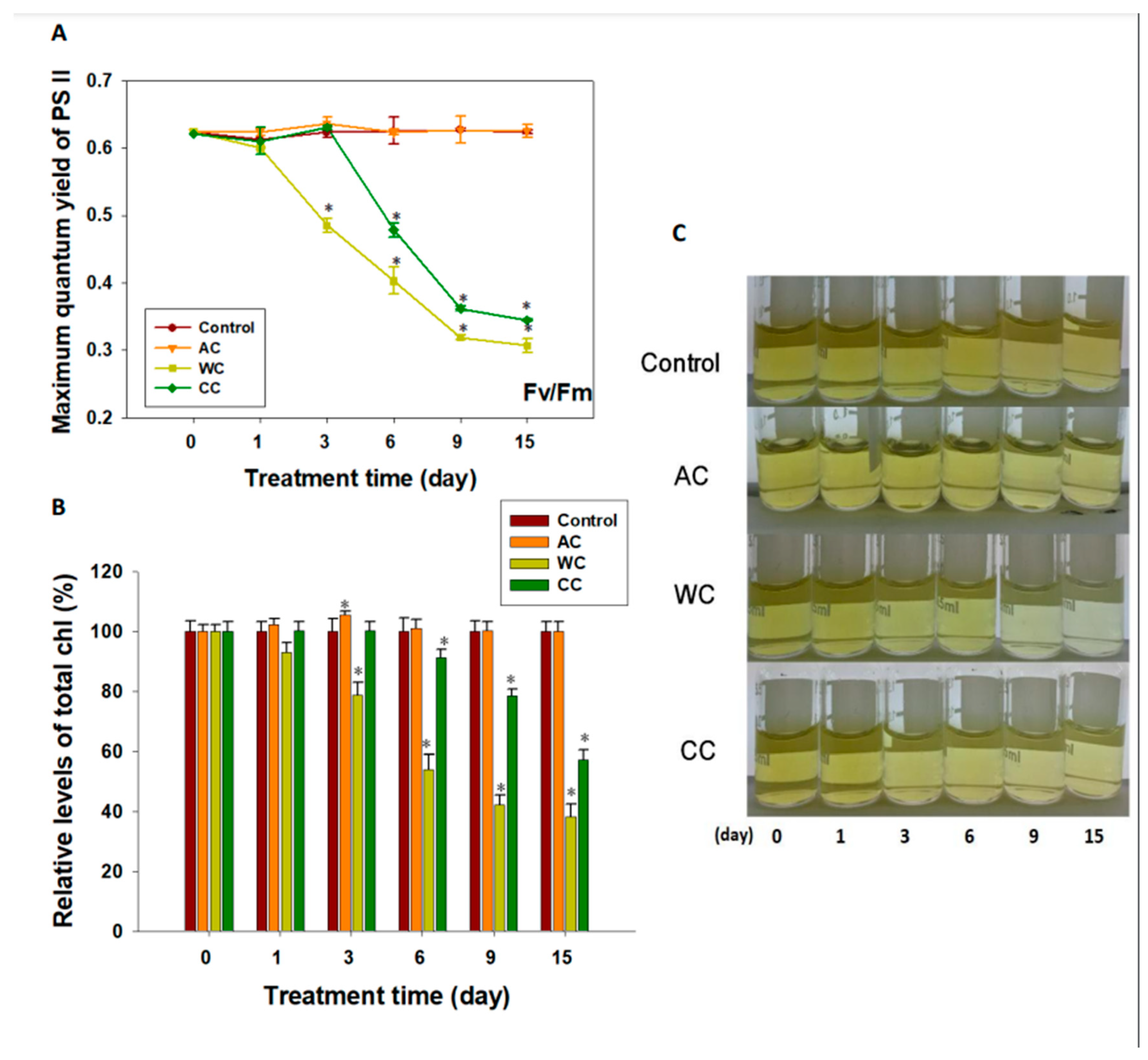
3.1. Maximum Quantum Yield of Photosystem II and Chlorophyll
3.2. Symbiodiniaceae Density
3.3. Cholesterol Contents and the Phenotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Paraschiv, S.; Paraschiv, L.S. Trends of carbon dioxide (CO2) emissions from fossil fuels combustion (coal, gas and oil) in the EU member states from 1960 to 2018. Energy Rep. 2020, 6, 237–242. [Google Scholar] [CrossRef]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean acidification: The other CO2 problem. Annu. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Bindoff, N.L.; Cheung, W.W.L.; Kairo, J.G.; Aristegui, J.; Guinder, V.A.; Hallberg, R.; Hilmi, N.; Jiao, N.; Karim, M.S.; Levin, L.S.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Portner, H.O., Roberts, D.C., Masson Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegria, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- Jackson, R.B.; Le Quere, C.; Andrew, R.M.; Canadell, J.G.; Peters, G.P.; Roy, J.; Wu, L. Warning signs for stabilizing global CO2 emissions. Environ. Res. Lett. 2017, 12, 110202. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, N., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Hoegh-Guldberg, O.; Bruno, J.F. The impact of climate change on the world’s marine ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Assan, D.; Kuebutornye, F.K.A.; Mustapha, U.F.; Chen, H.P.; Li, G.L. Effects of climate change on marine organisms. Am. J. Clim. Chang. 2020, 9, 204–216. [Google Scholar] [CrossRef]
- Martino, C.; Byrne, M.; Roccheri, M.C.; Chiarelli, R. Interactive effects of increased temperature and gadolinium pollution in Paracentrotus lividus sea urchin embryos: A climate change perspective. Aquat. Toxicol. 2021, 232, 105750. [Google Scholar] [CrossRef]
- Flores, F.; Marques, J.A.; Uthicke, S.; Fisher, R.; Patel, F.; Kaserzon, S.; Negri, A.P. Combined effects of climate change and the herbicide diuron on the coral acropora millepora. Mar. Pollut. Bull. 2021, 169, 112582. [Google Scholar] [CrossRef]
- Nordborg, F.M.; Jones, R.J.; Oelgemoller, M.; Negri, A.P. The effects of ultraviolet radiation and climate on oil toxicity to coral reef organisms—A review. Sci. Total Environ. 2020, 720, 137486. [Google Scholar] [CrossRef]
- Kibria, G.; Nugegoda, D.; Rose, G.; Haroon, A.K.Y. Climate change impacts on pollutants mobilization and interactive effects of climate change and pollutants on toxicity and bioaccumulation of pollutants in estuarine and marine biota and linkage to seafood security. Mar. Pollut. Bull. 2021, 167, 112364. [Google Scholar] [CrossRef]
- Suggett, D.J.; Smith, D.J. Coral bleaching patterns are the outcome of complex biological and environmental networking. Glob. Chang. Biol. 2020, 26, 68–79. [Google Scholar] [CrossRef]
- Roth, M.S. The engine of the reef: Photobiology of the coral-algal symbiosis. Front. Microbiol. 2014, 5, 422. [Google Scholar] [PubMed]
- Elias, S.A. Loss of Coral Reefs. In Encyclopedia of the Anthropocene; Dellasala, D.A., Goldstein, M.I., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 245–258. [Google Scholar]
- Tambutte, E.; Venn, A.A.; Holcomb, M.; Segonds, N.; Techer, N.; Zoccola, D.; Tambutte, S. Morphological plasticity of the coral skeleton under CO2-driven seawater acidification. Nat. Commun. 2015, 6, 7368. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.I.; Teneva, L.; Okamoto, D.K.; Schneider, K.; Caldeira, K.; Miard, T.; Chai, A.; Marker, M.; Dunbar, R.B.; Mitchell, B.G.; et al. Living coral tissue slows skeletal dissolution related to ocean acidification. Nat. Ecol. Evol. 2019, 3, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, H.; Wouters, J.; Yasuda, N. Seasonal calcification of the coral Acropora digitifera from a subtropical marginal Okinawa reef under ocean acidification. Coral Reefs 2019, 38, 443–454. [Google Scholar] [CrossRef]
- Van der Zande, R.M.; Achlatis, M.; Bender-Champ, D.; Kubicek, A.; Dove, S.; Hoegh-Guldberg, O. Paradise lost: End-of-century warming and acidification under business-as-usual emissions have severe consequences for symbiotic corals. Glob. Chang. Biol. 2020, 26, 2203–2219. [Google Scholar] [CrossRef] [PubMed]
- Tansik, A.L.; Hopkinson, B.M.; Meile, C. Inorganic carbon fluxes and perturbations by ocean acidification estimated using a data-constrained, process-based model of coral physiology. Mar. Biol. 2021, 168, 116. [Google Scholar] [CrossRef]
- Guo, W.F.; Bokade, R.; Cohen, A.L.; Mollica, N.R.; Leung, M.; Barainard, R.E. Ocean acidification has impacted coral growth on the Great Barrier Reef. Geophys. Res. Lett. 2020, 47, e2019GL086761. [Google Scholar] [CrossRef]
- Albright, R.; Takeshita, Y.; Koweek, D.A.; Ninokawa, A.; Wolfe, K.; Rivlin, T.; Nebuchina, Y.; Young, J.; Caldeira, K. Carbon dioxide addition to coral reef waters suppresses net community calcification. Nature 2018, 555, 516. [Google Scholar] [CrossRef]
- Eyre, B.D.; Cyronak, T.; Drupp, P.; De Carlo, E.H.; Sachs, J.P.; Andersson, A.J. Coral reefs will transition to net dissolving before end of century. Science 2018, 359, 908–911. [Google Scholar] [CrossRef]
- Hennige, S.J.; Wicks, L.C.; Kamenos, N.A.; Perna, G.; Findlay, H.S.; Roberts, J.M. Hidden impacts of ocean acidification to live and dead coral framework. Proc. R. Soc. B 2015, 282, 20150990. [Google Scholar] [CrossRef]
- Rivest, E.B.; Chen, C.S.; Fan, T.Y.; Li, H.H.; Hofmann, G.E. Lipid consumption in coral larvae differs among sites: A consideration of environmental history in a global ocean change scenario. Proc. R. Soc. B-Biol. Sci. 2017, 284, 20162825. [Google Scholar] [CrossRef] [PubMed]
- Biscere, T.; Zampighi, M.; Lorrain, A.; Jurriaans, S.; Foggo, A.; Houlbreque, F.; Rodolfo-Metalpa, R. High pCO2 promotes coral primary production. Biol. Lett. 2019, 15, 20180777. [Google Scholar] [CrossRef] [PubMed]
- Bergman, J.L. Components of a Flexible Phenotype in Two Species of Scleractinian Coral under Ocean Acidification. Ph.D. Thesis, California State University, Northridge, CA, USA, 2017. [Google Scholar]
- Liew, Y.J.; Zoccola, D.; Li, Y.; Tambutte, E.; Venn, A.A.; Michell, C.T.; Gui, G.; Deutekom, E.S.; Kaandorp, J.A.; Voolstra, C.R.; et al. Epigenome-associated phenotypic acclimatization to ocean acidification in a reef-building coral. Sci. Adv. 2018, 24, eaar8028. [Google Scholar] [CrossRef] [PubMed]
- Anthony, K.R.N.; Kline, D.I.; Diaz-Pulido, G.; Dove, S.; HoeghGuldberg, O. Ocean acidification causes bleaching and productivity loss in coral reef builders. Proc. Natl. Acad. Sci. USA 2008, 105, 17442–17446. [Google Scholar] [CrossRef] [PubMed]
- Kavousi, J.; Tanaka, Y.; Nishida, K.; Suzuki, A.; Nojiri, Y.; Nakamura, T. Colony-specific calcification and mortality under ocean acidification in the branching coral Montipora digitata. Mar. Environ. Res. 2016, 119, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Harii, S.; Hongo, C.; Ishihara, M.; Ide, Y.; Kayanne, H. Impacts of multiple disturbances on coral communities at Ishigaki Island, Okinawa, Japan, during a 15 year survey. Mar. Ecol. Prog. Ser. 2014, 509, 171–180. [Google Scholar] [CrossRef]
- Jaroensutasinee, K.; Somchuea, S.; Jaroensutasinee, M. Coral and reef fish community recovery following the 2010 extreme ocean warming event (mass bleaching event) at Thailand. J. Anim. Behav. Biometeorol. 2021, 9, 21004. [Google Scholar] [CrossRef]
- Goyen, S.; Camp, E.F.; Fujise, L.; Lloyd, A.; Nitschke, M.R.; Lajeunensse, T.; Kahlke, T.; Ralph, P.J.; Suggett, D. Mass coral bleaching of P. versipora in Sydney Harbour driven by the 2015–2016 heatwave. Coral Reefs 2019, 38, 815–830. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, J.; Zhao, H.; Han, X.; Xiang, Y.; Zhou, W. Clade-specific sterol metabolites in dinoflagellate endosymbionts are associated with coral bleaching in response to environmental cues. mSystems 2020, 5, e00765-20. [Google Scholar] [CrossRef]
- Pryor, S.H.; Andrews, L.; Kelaher, B.P.; Tagliafico, A.; Scott, A. Ocean temperature, but not acidification, causes sea anemone bleaching under a near-future climate scenario. Coral Reefs 2021, 40, 355–364. [Google Scholar]
- Hillyer, K.E.; Dias, D.A.; Lutz, A.; Wilkinson, S.P.; Roessner, U.; Davy, S.K. Metabolite profiling of symbiont and host during thermal stress and bleaching in the coral Acropora aspera. Coral Reefs 2016, 36, 105–118. [Google Scholar] [CrossRef]
- Roach, T.; Dilworth, J.; Christian, M.H.; Jones, D.; Quinn, R.A.; Drury, C. Metabolomic signatures of coral bleaching history. Nat. Ecol. Evol. 2021, 5, 495–503. [Google Scholar] [CrossRef]
- Williams, A.; Chiles, E.N.; Conetta, D.; Pathmanathan, J.S.; Cleves, P.A.; Putnam, H.M.; Su, X.Y.; Bhattacharya, D. Metabolomic shifts associated with heat stress in coral holobionts. Sci. Adv. 2021, 7, eabd4210. [Google Scholar] [CrossRef] [PubMed]
- Bove, C.B.; Umbanhowar, J.; Castillo, K.D. Meta-analysis reveals reduced coral calcification under projected ocean warming but not under acidification across the Caribbean sea. Front. Mar. Sci. 2020, 7, 127. [Google Scholar] [CrossRef]
- Schoepf, V.; D’Olivo, J.P.; Rigal, C.; Jung, E.M.; McCulloch, M.T. Heat stress differentially impacts key calcification mechanisms in reef-building corals. Coral Reefs 2021, 40, 459–471. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Langdon, C.; Uthicke, S.; Humphrey, C.; Noonan, S.; Death, G.; Okazaki, R.; Muehllehner, N.; Glas, M.S.; Lough, J.M. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nat. Clim. Chang. 2011, 1, 165–169. [Google Scholar] [CrossRef]
- Guillermic, M.; Cameron, L.P.; Corte, I.D.; Misra, S.; Bijma, J.; de Beer, D.; Reymond, C.E.; Westphal, H.; Ries, J.B.; Eagle, R.A. Thermal stress reduces pocilloporid coral resilience to ocean acidification by impairing control over calcifying fluid chemistry. Sci. Adv. 2021, 7, eaba9958. [Google Scholar] [CrossRef] [PubMed]
- Araujo-Silva, C.L.; Sarmento, V.C.; Santos, P.J.P. Climate change scenarios of increased CO2 and temperature affect a coral reef peracarid (Crustacea) community. Mar. Environ. Res. 2021, 173, 105518. [Google Scholar] [CrossRef]
- Kayal, E.; Roure, B.; Philippe, H.; Collins, A.G.; Lavrov, D.V. Cnidarian phylogenetic relationships as revealed by mitogenomics. BMC Evol. Biol. 2013, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Harrison, P.L. Embryonic and larval development of the host sea anemones Entacmaea quadricolor and Heteractis crispa. Biol. Bull. 2007, 213, 110–121. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ollerton, J.; McCollin, D.; Fautin, D.G.; Allen, G.R. Finding NEMO: Nestedness engendered by mutualistic organization in anemonefish and their hosts. Proc. R. Soc. B 2007, 274, 591–598. [Google Scholar] [CrossRef]
- Scott, A. Effects of feeding on the growth rates of captive-bred Heteractis crispa: A popular marine ornamental for aquariums. Bull. Mar. Sci. 2012, 88, 81–87. [Google Scholar] [CrossRef]
- Scott, A.; Dixson, D.L. Reef fishes can recognize bleached habitat during settlement: Sea anemone bleaching alters anemonefish host selection. Proc. R. Soc. B-Biol. Sci. 2016, 283, 20152694. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.M.; Ndobe, S.; Yasir, I.; Ambo-Rappe, R.; Jompa, J. Banggai cardinalfish and its microhabitats in a warming world: A preliminary study. IOP Conf. Ser. Earth Environ. Sci. 2019, 253, 012021. [Google Scholar] [CrossRef]
- Hill, L.J.; Paradas, W.C.; Willemes, M.J.; Pereira, M.G.; Salomon, P.S.; Mariath, R.; Moura, R.L.; Atella, G.C.; Farina, M.; Amado-Filho, G.M.; et al. Acidification-induced cellular changes in Symbiodinium isolated from Mussismilia braziliensis. PLoS ONE 2019, 14, e0220130. [Google Scholar] [CrossRef] [PubMed]
- Kelaher, B.P.; Coleman, M.A.; Bishop, M.J. Ocean warming, but not acidification, accelerates seagrass decomposition under near future climate scenarios. Mar. Ecol. Prog. Ser. 2018, 605, 103–110. [Google Scholar] [CrossRef]
- Lewis, E.; Wallace, D.W.R. Program Developed for CO2 System Calculation; ORNL/CDIAC-105; Carbon Dioxide Information Analysis Center: Oak Ridge, TN, USA, 1998.
- Cleves, P.A.; Krediet, C.J.; Lehnert, E.M.; Onishi, M.; Pringle, J.R. Insights into coral bleaching under heat stress from analysis of gene expression in a sea anemone model system. Proc. Natl. Acad. Sci. USA 2020, 117, 28906–28917. [Google Scholar] [CrossRef]
- Molina, V.H.; Castillo-Medina, R.E.; Thome, P.E. Experimentally induced bleaching in the Sea Anemone Exaiptasia supports glucose as a main metabolite associated with its symbiosis. J. Mar. Biol. 2017, 2017, 3130723. [Google Scholar] [CrossRef]
- Jiang, J.; Lu, Y. Metabolite profiling of Breviolum minutum in response to acidification. Aquat. Toxicol. 2019, 213, 105215. [Google Scholar] [CrossRef]
- Ventura, P.; Jarrold, M.D.; Merle, P.L.; Barnay-Verdier, S.; Zamoum, T.; Rodolfo-Metalpa, R.; Calosi, P.; Furla, P. Resilience to ocean acidification: Decreased carbonic anhydrase activity in sea anemones under high pCO2 conditions. Mar. Ecol. Prog. Ser. 2016, 559, 257–263. [Google Scholar] [CrossRef][Green Version]
- Hoadley, K.D.; Pettay, D.T.; Dodge, D.; Warner, M.E. Contrasting physiological plasticity in response to environmental stress within different cnidarians and their respective symbionts. Coral Reefs 2016, 35, 529–542. [Google Scholar] [CrossRef]
- Noonan, S.H.C.; Fabricius, K.E. Ocean acidification affects productivity but not the severity of thermal bleaching in some tropical corals. ICES J. Mar. Sci. 2016, 73, 715–726. [Google Scholar] [CrossRef]
- Davies, S.W.; Ries, J.B.; Marchetti, A.; Castillo, K.D. Symbiodinium functional diversity in the coral Siderastrea siderea is influenced by thermal stress and reef environment, but not ocean acidification. Front. Mar. Sci. 2018, 5, 150. [Google Scholar] [CrossRef]
- Rivest, E.B.; Kelly, M.W.; DeBiasse, M.B.; Hofmann, G.E. Host and symbionts in Pocillopora damicornis larvae display different transcriptomic responses to ocean acidification and warming. Front. Mar. Sci. 2018, 5, 186. [Google Scholar] [CrossRef]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, A.; Deng, X.; Zhou, W.; Gan, Q.; Lu, Y. How Symbiodiniaceae meets the challenges of life during coral bleaching. Coral Reefs 2021, 40, 1339–1353. [Google Scholar] [CrossRef]
- Sogin, E.M.; Putnam, H.M.; Anderson, P.E.; Gates, R.D. Metabolomic signatures of increases in temperature and ocean acidification from the reef-building coral, Pocillopora damicornis. Metabolomics 2016, 12, 71. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Tian, W.; Chen, C.; Ye, Q.; Yang, L.; Jiang, J. Adaptive Responses of the Sea Anemone Heteractis crispa to the Interaction of Acidification and Global Warming. Animals 2022, 12, 2259. https://doi.org/10.3390/ani12172259
Wu Y, Tian W, Chen C, Ye Q, Yang L, Jiang J. Adaptive Responses of the Sea Anemone Heteractis crispa to the Interaction of Acidification and Global Warming. Animals. 2022; 12(17):2259. https://doi.org/10.3390/ani12172259
Chicago/Turabian StyleWu, Yangyang, Wenfei Tian, Chunxing Chen, Quanqing Ye, Liu Yang, and Jiaoyun Jiang. 2022. "Adaptive Responses of the Sea Anemone Heteractis crispa to the Interaction of Acidification and Global Warming" Animals 12, no. 17: 2259. https://doi.org/10.3390/ani12172259
APA StyleWu, Y., Tian, W., Chen, C., Ye, Q., Yang, L., & Jiang, J. (2022). Adaptive Responses of the Sea Anemone Heteractis crispa to the Interaction of Acidification and Global Warming. Animals, 12(17), 2259. https://doi.org/10.3390/ani12172259