Palbociclib Induces the Apoptosis of Lung Squamous Cell Carcinoma Cells via RB-Independent STAT3 Phosphorylation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Lines and Culture
2.3. Cell Viability Assay
2.4. Flow Cytometry Analysis
2.5. Western Blotting
2.6. Real-Time PCR
2.7. ELISA
2.8. SiRNA Transfection
2.9. Myc Plasmid Transfection
2.10. Statistical Analysis
3. Results
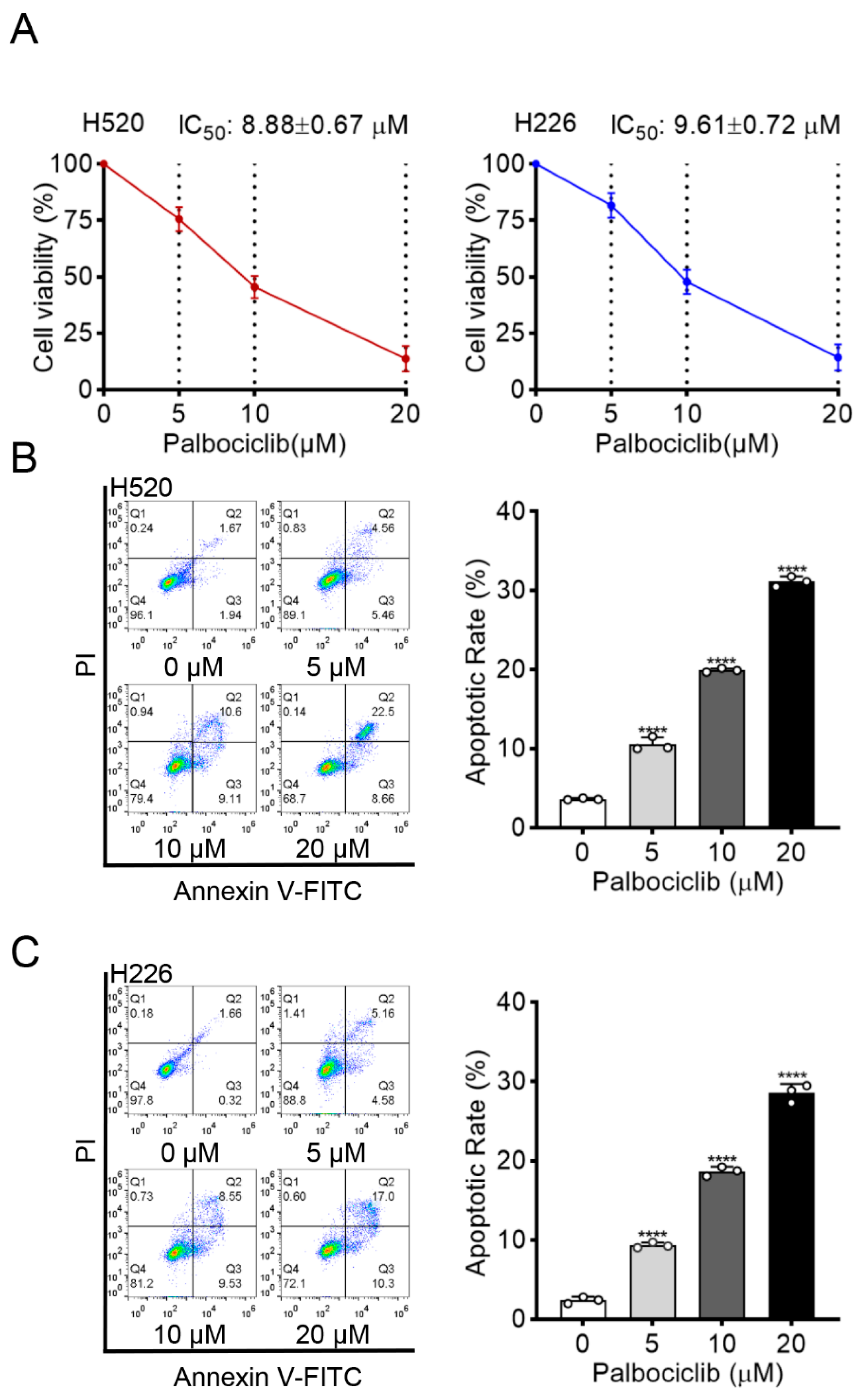
3.1. Palbociclib Exhibits a Pro-Apoptotic Activity for LUSC Cells
3.2. Inhibition of STAT3 Signaling Decreases Palbociclib-Induced LUSC Apoptosis
3.3. Palbociclib Inhibits STAT3 Phosphorylation via Src Inhibition in RB-Independent Manner
3.4. Palbociclib Inhibits Src/STAT3 Signaling via Suppressing IL-1β and IL-6 Expression in LUSC Cells
3.5. Myc Mediates the Sensitivity of LUSC Cells to Palbociclib
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| Cyclin D | D-type cyclins |
| CDK4/6 | Cyclin-dependent kinase 4/6 |
| DMSO | Dimethyl Sulfoxide |
| ER | estrogen receptor |
| ELISA | Enzyme-linked immunosorbent assay |
| FBS | Fetal bovine serum |
| IL | interleukin |
| JAK2 | Janus kinase 2 |
| LUSC | Lung squamous cell carcinoma |
| MTT | 3-(4:5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide, Thiazolyl Blue Tetrazolium Bromide |
| NSCLC | Non-small cell lung cancer |
| PBS | Phosphate buffered solution |
| PI | Propidium iodide |
| (RPMI)-1640 | Roswellpark memorial institute |
| STAT3 | Signal transducers and activators of transcription 3 |
References
- Bade, B.C.; Dela Cruz, C.S. Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clin. Chest Med. 2020, 41, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Perez-Moreno, P.; Brambilla, E.; Thomas, R.; Soria, J.C. Squamous cell carcinoma of the lung: Molecular subtypes and therapeutic opportunities. Clin. Cancer Res. 2012, 18, 2443–2451. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, I.; Dynlacht, B.D. New insights into cyclins, CDKs, and cell cycle control. Semin. Cell Dev. Biol. 2005, 16, 311–321. [Google Scholar] [CrossRef] [PubMed]
- VanArsdale, T.; Boshoff, C.; Arndt, K.T.; Abraham, R.T. Molecular Pathways: Targeting the Cyclin D-CDK4/6 Axis for Cancer Treatment. Clin. Cancer Res. 2015, 21, 2905–2910. [Google Scholar] [CrossRef]
- Blain, S.W. Switching cyclin D-Cdk4 kinase activity on and off. Cell Cycle 2008, 7, 892–898. [Google Scholar] [CrossRef]
- Inoue, Y.; Kitagawa, M.; Taya, Y. Phosphorylation of pRB at Ser612 by Chk1/2 leads to a complex between pRB and E2F-1 after DNA damage. EMBO J. 2007, 26, 2083–2093. [Google Scholar] [CrossRef]
- Gandara, D.R.; Hammerman, P.S.; Sos, M.L.; Lara, P.N., Jr.; Hirsch, F.R. Squamous cell lung cancer: From tumor genomics to cancer therapeutics. Clin. Cancer Res. 2015, 21, 2236–2243. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Comprehensive genomic characterization of squamous cell lung cancers. Nature 2012, 489, 519–525. [Google Scholar] [CrossRef]
- O’Leary, B.; Finn, R.S.; Turner, N.C. Treating cancer with selective CDK4/6 inhibitors. Nat. Rev. Clin. Oncol. 2016, 13, 417–430. [Google Scholar] [CrossRef]
- Rocca, A.; Schirone, A.; Maltoni, R.; Bravaccini, S.; Cecconetto, L.; Farolfi, A.; Bronte, G.; Andreis, D. Progress with palbociclib in breast cancer: Latest evidence and clinical considerations. Ther. Adv. Med. Oncol. 2017, 9, 83–105. [Google Scholar] [CrossRef]
- Ahn, E.R.; Mangat, P.K.; Garrett-Mayer, E.; Halabi, S.; Dib, E.G.; Haggstrom, D.E.; Alguire, K.B.; Calfa, C.J.; Cannon, T.L.; Crilley, P.A.; et al. Palbociclib in Patients with Non-Small-Cell Lung Cancer with Alterations: Results from the Targeted Agent and Profiling Utilization Registry Study. JCO Precis. Oncol. 2020, 4, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Jin, Z.; Liu, Z.; Sun, L.; Wang, L.; Li, K. Correlation of activated STAT3 expression with clinicopathologic features in lung adenocarcinoma and squamous cell carcinoma. Mol. Digan. Ther. 2011, 15, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Fathi, N.; Rashidi, G.; Khodadadi, A.; Shahi, S.; Sharifi, S. STAT3 and apoptosis challenges in cancer. Int. J. Biol. Macromol. 2018, 117, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Lee, J.; Kang, D.; Lee, D.H.; Kim, Y.J.; Hwang, S.G.; Kim, D.I.; Lee, C.W.; Lee, K.H. Multipotent neurogenic fate of mesenchymal stem cell is determined by Cdk4-mediated hypophosphorylation of Smad-STAT3. Cell Cycle 2016, 15, 1787–1795. [Google Scholar]
- Chai, E.Z.; Shanmugam, M.K.; Arfuso, F.; Dharmarajan, A.; Wang, C.; Kumar, A.P.; Samy, R.P.; Lim, L.H.; Wang, L.; Goh, B.C.; et al. Targeting transcription factor STAT3 for cancer prevention and therapy. Pharmacol. Ther. 2016, 162, 86–97. [Google Scholar] [CrossRef]
- Garcia, R.; Bowman, T.L.; Niu, G.; Yu, H.; Minton, S.; Muro-Cacho, C.A.; Cox, C.E.; Falcone, R.; Fairclough, R.; Parsons, S.; et al. Constitutive activation of Stat3 by the Src and JAK tyrosine kinases participates in growth regulation of human breast carcinoma cells. Oncogene 2001, 20, 2499–2513. [Google Scholar] [CrossRef]
- Massó-Vallés, D.; Beaulieu, M.E.; Soucek, L. MYC, MYCL, and MYCN as therapeutic targets in lung cancer. Expert Opin. Ther. Targets 2020, 24, 101–114. [Google Scholar] [CrossRef]
- Goga, A.; Yang, D.; Tward, A.D.; Morgan, D.O.; Bishop, J.M. Inhibition of CDK1 as a potential therapy for tumors over-expressing MYC. Nat. Med. 2007, 13, 820–827. [Google Scholar] [CrossRef]
- Thangavel, C.; Boopathi, E.; Liu, Y.; McNair, C.; Haber, A.; Perepelyuk, M.; Bhardwaj, A.; Addya, S.; Ertel, A.; Shoyele, S.; et al. Therapeutic Challenge with a CDK 4/6 Inhibitor Induces an RB-Dependent SMAC-Mediated Apoptotic Response in Non-Small Cell Lung Cancer. Clin. Cancer Res. 2018, 24, 1402–1414. [Google Scholar] [CrossRef]
- Chen, L.; Pan, J. Dual cyclin-dependent kinase 4/6 inhibition by PD-0332991 induces apoptosis and senescence in oesophageal squamous cell carcinoma cells. Br. J. Pharmacol. 2017, 174, 2427–2443. [Google Scholar] [CrossRef]
- Germain, D.; Frank, D.A. Targeting the cytoplasmic and nuclear functions of signal transducers and activators of transcription 3 for cancer therapy. Clin. Cancer Res. 2007, 13, 5665–5669. [Google Scholar] [CrossRef] [PubMed]
- Diaz, N.; Minton, S.; Cox, C.; Bowman, T.; Gritsko, T.; Garcia, R.; Eweis, I.; Wloch, M.; Livingston, S.; Seijo, E.; et al. Activation of stat3 in primary tumors from high-risk breast cancer patients is associated with elevated levels of activated SRC and survivin expression. Clin. Cancer Res. 2006, 12, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.L.; Thangavel, C.; McClendon, A.K.; Reed, C.A.; Knudsen, E.S. Therapeutic CDK4/6 inhibition in breast cancer: Key mechanisms of response and failure. Oncogene 2010, 29, 4018–4032. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneira, D.B.; Mayhew, C.N.; Thangavel, C.; Sotillo, E.; Reed, C.A.; Graña, X.; Knudsen, E.S. Proliferative suppression by CDK4/6 inhibition: Complex function of the retinoblastoma pathway in liver tissue and hepatoma cells. Gastroenterology 2010, 138, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Dubé, N.; Kim, J.W.; Cheng, A.; Ibarra-Sanchez Mde, J.; Tremblay, M.L.; Boisclair, Y.R. Protein tyrosine phosphatase 1B attenuates growth hormone-mediated JAK2-STAT signaling. Mol. Cell Biol. 2003, 23, 3753–3762. [Google Scholar] [CrossRef]
- Kim, H.Y.; Park, E.J.; Joe, E.H.; Jou, I. Curcumin suppresses Janus kinase-STAT inflammatory signaling through activation of Src homology 2 domain-containing tyrosine phosphatase2 in brain microglia. J. Immunol. 2003, 171, 6072–6079. [Google Scholar] [CrossRef]
- Kuninaka, S.; Yano, T.; Yokoyama, H.; Fukuyama, Y.; Terazaki, Y.; Uehara, T.; Kanematsu, T.; Asoh, H.; Ichinose, Y. Direct influences of pro-inflammatory cytokines (IL-1beta, TNF-alpha, IL-6) on the proliferation and cell-surface antigen expression of cancer cells. Cytokine 2000, 12, 8–11. [Google Scholar] [CrossRef]
- Vahl, J.M.; Friedrich, J.; Mittler, S.; Trump, S.; Heim, L.; Kachler, K.; Balabko, L.; Fuhrich, N.; Geppert, C.I.; Trufa, D.I.; et al. Interleukin-10-regulated tumour tolerance in non-small cell lung cancer. Br. J. Cancer 2017, 117, 1644–1655. [Google Scholar] [CrossRef]
- Chang, C.H.; Hsiao, C.F.; Yeh, Y.M.; Chang, G.C.; Tsai, Y.H.; Chen, Y.M.; Huang, M.S.; Chen, H.L.; Li, Y.J.; Yang, P.C.; et al. Circulating interleukin-6 level is a prognostic marker for survival in advanced nonsmall cell lung cancer patients treated with chemotherapy. Int. J. Cancer 2013, 132, 1977–1985. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Wang, Y.; Liu, Q. Palbociclib improves cardiac dysfunction in diabetic cardiomyopathy by regulating RB phosphorylation. Am. J. Transl. Res. 2019, 11, 3481–3489. [Google Scholar]
- Iwakawa, R.; Kohno, T.; Kato, M.; Shiraishi, K.; Tsuta, K.; Noguchi, M.; Ogawa, S.; Yokota, J. MYC amplification as a prognostic marker of early-stage lung adenocarcinoma identified by whole genome copy number analysis. Clin. Cancer Res. 2011, 17, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Sergio, C.M.; Sutherland, R.L.; Musgrove, E.A. Targeting cyclin-dependent kinase 1 (CDK1) but not CDK4/6 or CDK2 is selectively lethal to MYC-dependent human breast cancer cells. BMC Cancer 2014, 14, 32. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, W.; Qi, W.; Li, H.; Sun, J.; Dong, C.; Ou, H.; Liu, B. Palbociclib Induces the Apoptosis of Lung Squamous Cell Carcinoma Cells via RB-Independent STAT3 Phosphorylation. Curr. Oncol. 2022, 29, 5855-5868. https://doi.org/10.3390/curroncol29080462
Xiang W, Qi W, Li H, Sun J, Dong C, Ou H, Liu B. Palbociclib Induces the Apoptosis of Lung Squamous Cell Carcinoma Cells via RB-Independent STAT3 Phosphorylation. Current Oncology. 2022; 29(8):5855-5868. https://doi.org/10.3390/curroncol29080462
Chicago/Turabian StyleXiang, Wenjing, Wanchen Qi, Huayu Li, Jia Sun, Chao Dong, Haojie Ou, and Bing Liu. 2022. "Palbociclib Induces the Apoptosis of Lung Squamous Cell Carcinoma Cells via RB-Independent STAT3 Phosphorylation" Current Oncology 29, no. 8: 5855-5868. https://doi.org/10.3390/curroncol29080462
APA StyleXiang, W., Qi, W., Li, H., Sun, J., Dong, C., Ou, H., & Liu, B. (2022). Palbociclib Induces the Apoptosis of Lung Squamous Cell Carcinoma Cells via RB-Independent STAT3 Phosphorylation. Current Oncology, 29(8), 5855-5868. https://doi.org/10.3390/curroncol29080462