
Deciphering the Effectiveness of Humic Substances and Biochar Modified Digestates on Soil Quality and Plant Biomass Accumulation

 , ,
, ,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
- i.
- Digestate enriched with Humac would improve microbial activities leading to enhanced mineralization of nutrients (C, N, P).
- ii.
- Digestate enriched with biochar (or biochar + Humac) would increase total carbon, C:N ratio, and microbial respiration.
- iii.
- Positive effects of digestate + biochar (eventually + Humac) on catabolism would be only short-term (until utilization of labile carbon in DG + amendment blend), with priming negative effects on microbial abundance.
- iv.
- Long-term effects of digestate + biochar (+Humac) on nutrient mineralization would be negative, due to biochar-mediated immobilization and sequestration of nutrients.
2. Materials and Methods
2.1. Procurement and Preparation of Soil Amendments
2.2. Experimental Design and Treatments
2.3. Soil Analysis
2.4. Statistical Analyses
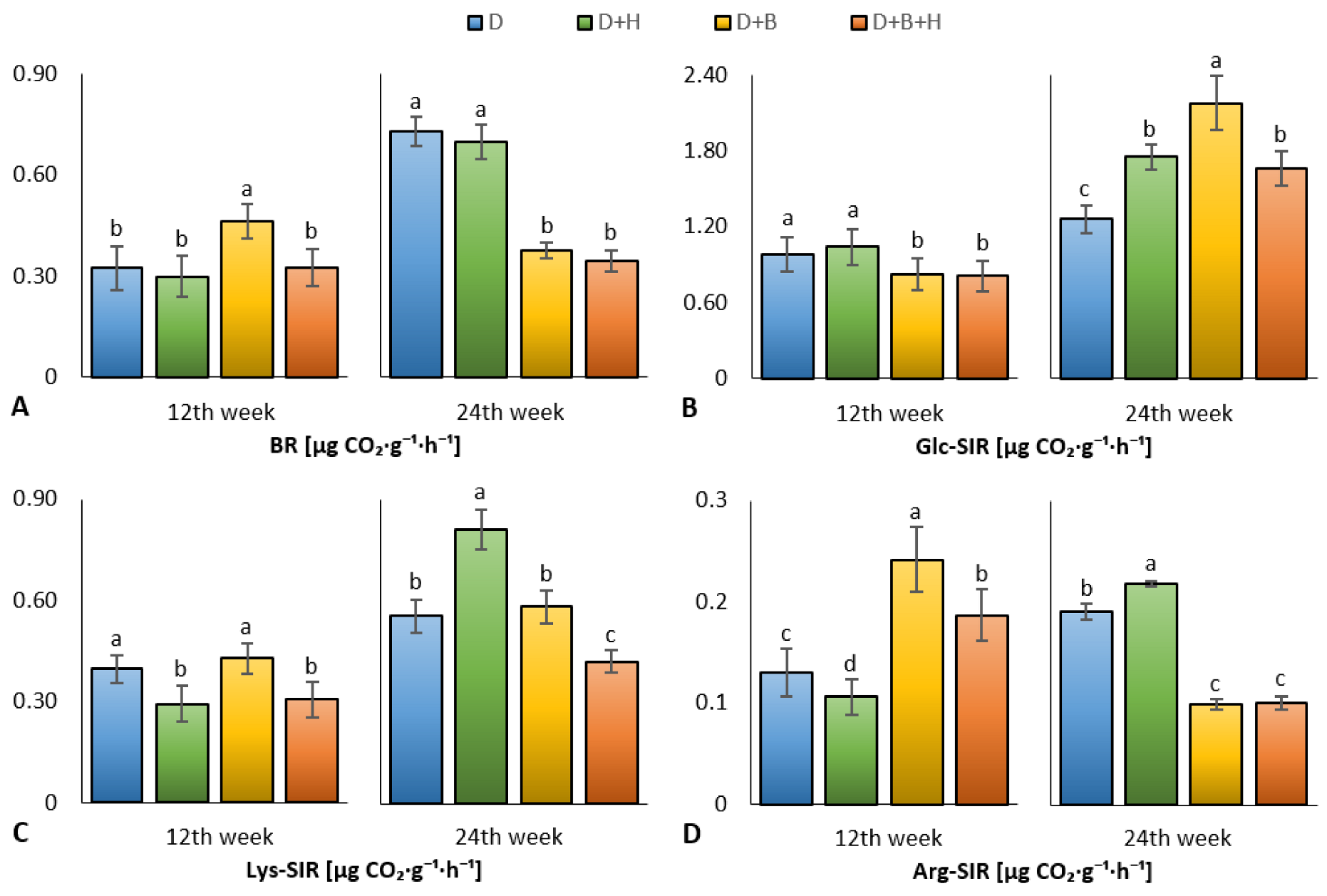
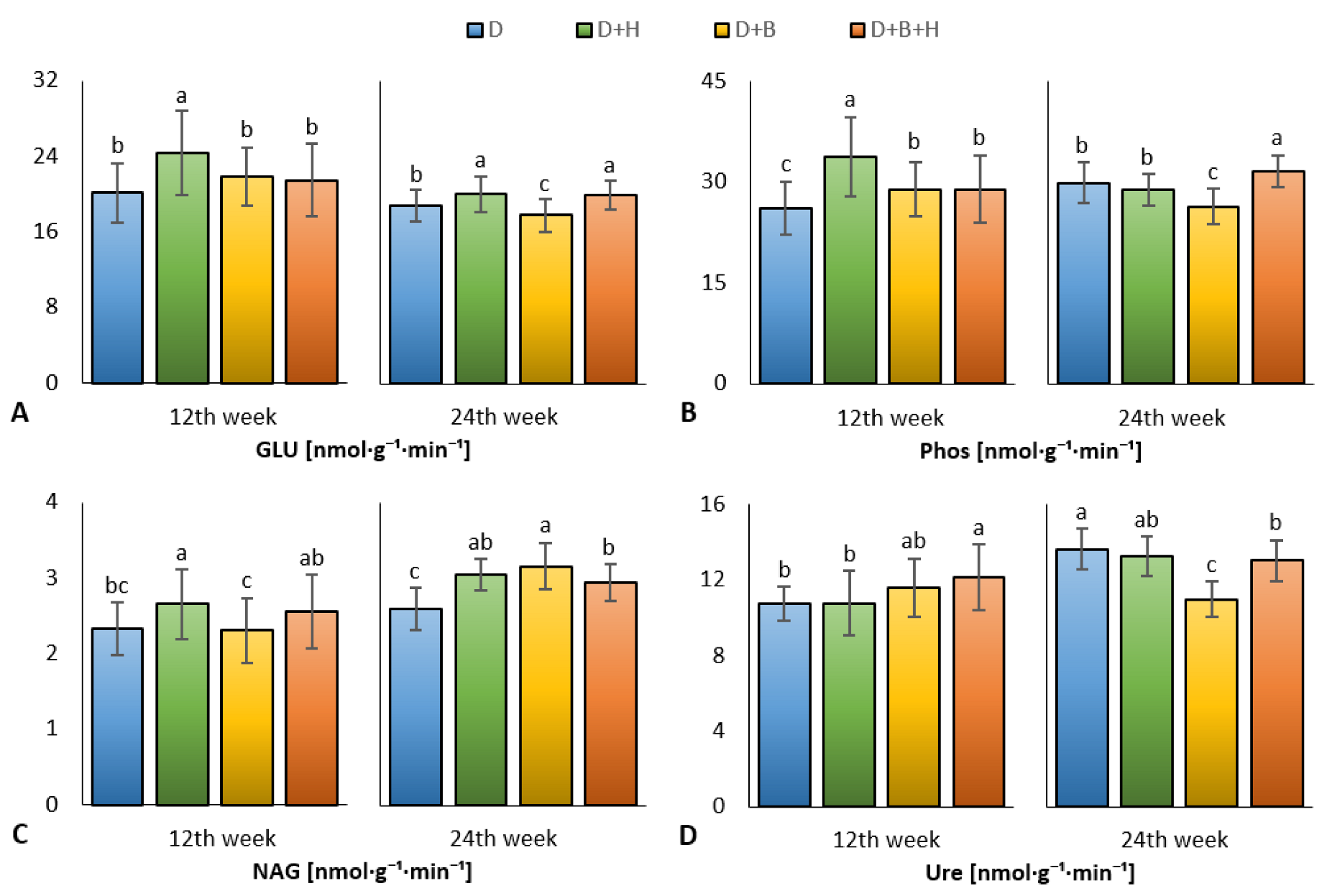
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Glæsner, N.; Helming, K.; De Vries, W. Do current european policies prevent soil threats and support soil functions? Sustainability 2014, 6, 9538–9563. [Google Scholar] [CrossRef] [Green Version]
- COWI. Technical Guidance Handbook—Setting up and Implementing Result-Based Carbon Farming Mechanisms in the EU Report to the European Commission; Contract No. CLIMA/C.3/ETU/2018/007; Ecologic Institute: Berlin, Germany; IEEP: Bruxelles, Belgium; DG Climate Action: Kongens Lyngby, Denmark, 2021. [Google Scholar]
- Kilpeläinen, S.; Aalto, P.; Toivanen, P.; Lehtonen, P.; Holttinen, H. How to achieve a more resource-efficient and climate-neutral energy system by 2030? Views of nordic stakeholders. Rev. Policy Res. 2019, 36, 448–472. [Google Scholar] [CrossRef]
- Luo, L.; Qin, L.; Wang, Y.; Wang, Q. Environmentally-friendly agricultural practices and their acceptance by smallholder farmers in china-a case study in xinxiang county, henan province. Sci. Total Environ. 2016, 571, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.C.D.; Farias, W.M.; Melo, B.A.D.; Chagas, J.K.M.; Vale, A.T.; Coser, T.R. Labile and stable pools of organic matter in soil amended with sewage sludge biochar. Arch. Agron. Soil Sci. 2018, 65, 770–781. [Google Scholar] [CrossRef]
- Klinglmair, M.; Thomsen, M. Using food waste in organic fertilizer: Modelling biogenic carbon sequestration with associated nutrient and micropollutant loads. Sustainability 2020, 12, 7399. [Google Scholar] [CrossRef]
- Shaji, H.; Chandran, V.; Mathew, L. Chapter 13—Organic Fertilizers as a Route to Controlled Release of Nutrients. In Controlled Release Fertilizers for Sustainable Agriculture; Lewu, F.B., Volova, T., Thomas, S., Rakhimol, K.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 231–245. [Google Scholar] [CrossRef]
- Barłóg, P.; Hlisnikovský, L.; Kunzová, E. Effect of digestate on soil organic carbon and plant-available nutrient content compared to cattle slurry and mineral fertilization. Agronomy 2020, 10, 379. [Google Scholar] [CrossRef] [Green Version]
- Slepetiene, A.; Kochiieru, M.; Jurgutis, L.; Mankeviciene, A.; Skersiene, A.; Belova, O. The effect of anaerobic digestate on the soil organic carbon and humified carbon fractions in different land-use systems in lithuania. Land 2022, 11, 133. [Google Scholar] [CrossRef]
- Chen, R.; Blagodatskaya, E.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Kuzyakov, Y. Decomposition of biogas residues in soil and their effects on microbial growth kinetics and enzyme activities. Biomass Bioenergy 2012, 45, 221–229. [Google Scholar] [CrossRef]
- Stumpe, B.; Werner, S.; Jung, R.; Heinze, S.; Jüschke, E.; Strippel, C.; Marschner, B. Organic carbon dynamics and enzyme activities in agricultural soils amended with biogas slurry, liquid manure and sewage sludge. Agric. Sci. 2012, 3, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, I.K.; Olesen, J.E.; Møller, H.B.; Sørensen, P.; Christensen, B.T. Carbon dynamics and retention in soil after anaerobic digestion of dairy cattle feed and faeces. Soil Biol. Biochem. 2013, 58, 82–87. [Google Scholar] [CrossRef]
- Béghin-Tanneau, R.; Guérin, F.; Guiresse, M.; Kleiber, D.; Scheiner, J.D. Carbon sequestration in soil amended with anaerobic digested matter. Soil Tillage Res. 2019, 192, 87–94. [Google Scholar] [CrossRef]
- Slepetiene, A.; Kadziuliene, Z.; Feiziene, D.; Liaudanskiene, I.; Amaleviciute-Volunge, K.; Slepetys, J.; Skersiene, A.; Velykis, A.; Armolaitis, K. The distribution of organic carbon, its forms and macroelements in agricultural soils. Zemdirb. Agric. 2020, 107, 291–300. [Google Scholar] [CrossRef]
- Leno, N.; Sudharmaidevi, C.R.; Byju, G.; Thampatti, K.C.M.; Krishnaprasad, P.U.; Jacob, G.; Gopinath, P.P. Thermochemical digestate fertilizer from solid waste: Characterization, labile carbon dynamics, dehydrogenase activity, water holding capacity and biomass allocation in banana. Waste Manag. 2021, 123, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Voltr, V.; Menšík, L.; Hlisnikovský, L.; Hruška, M.; Pokorný, E.; Pospíšilová, L. The soil organic matter in connection with soil properties and soil inputs. Agronomy 2021, 11, 779. [Google Scholar] [CrossRef]
- Tang, Y.; Luo, L.; Carswell, A.; Misselbrook, T.; Shen, J.; Han, J. Changes in soil organic carbon status and microbial community structure following biogas slurry application in a wheat-rice rotation. Sci. Total Environ. 2021, 757, 143786. [Google Scholar] [CrossRef]
- Kolář, L.; Kužel, S.; Peterka, J.; Štindl, P.; Plát, V. Agrochemical value of organic matter of fermenter wastes in biogas production. Plant Soil Environ. 2008, 54, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Sarma, B.; Farooq, M.; Gogoi, N.; Borkotoki, B.; Kataki, R.; Garg, A. Soil organic carbon dynamics in wheat—Green gram crop rotation amended with vermicompost and biochar in combination with inorganic fertilizers: A comparative study. J. Clean. Prod. 2018, 201, 471–480. [Google Scholar] [CrossRef]
- Ruhlmann, J. Calculation of net nitrogen mineralization from the decomposable soil organic matter pool. In Proceedings of the International Workshop on Ecological Aspects of Vegetable Fertilisation in Integrated Crop Production in the Field, Wellesbourne Hastings, Warwick, UK, 27–31 July 1998; International Society Horticultural Science: Leuven, Belgium, 1998. [Google Scholar]
- Ludwig, B.; Geisseler, D.; Michel, K.; Joergensen, R.G.; Schulz, E.; Merbach, I.; Raupp, J.; Rauber, R.; Hu, K.; Niu, L.; et al. Effects of fertilization and soil management on crop yields and carbon stabilization in soils. A review. Agron. Sustain. Dev. 2010, 31, 361–372. [Google Scholar] [CrossRef] [Green Version]
- Piccoli, I.; Francioso, O.; Camarotto, C.; Delle Vedove, G.; Lazzaro, B.; Giandon, P.; Morari, F. Assessment of the short-term impact of anaerobic digestate on soil C stock and CO2 emissions in shallow water table conditions. Agronomy 2022, 12, 504. [Google Scholar] [CrossRef]
- Molinuevo-Salces, B.; Gomez, X.; Moran, A.; Garcia-Gonzalez, M.C. Anaerobic co-digestion of livestock and vegetable processing wastes: Fibre degradation and digestate stability. Waste Manag. 2013, 33, 1332–1338. [Google Scholar] [CrossRef]
- Tambone, F.; Adani, F.; Gigliotti, G.; Volpe, D.; Fabbri, C.; Provenzano, M.R. Organic matter characterization during the anaerobic digestion of different biomasses by means of cpmas 13c nmr spectroscopy. Biomass Bioenergy 2013, 48, 111–120. [Google Scholar] [CrossRef]
- Shakeri Yekta, S.; Hedenstrom, M.; Svensson, B.H.; Sundgren, I.; Dario, M.; Enrich-Prast, A.; Hertkorn, N.; Bjorn, A. Molecular characterization of particulate organic matter in full scale anaerobic digesters: An nmr spectroscopy study. Sci. Total Environ. 2019, 685, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Tambone, F.; Orzi, V.; Zilio, M.; Adani, F. Measuring the organic amendment properties of the liquid fraction of digestate. Waste Manag. 2019, 88, 21–27. [Google Scholar] [CrossRef]
- Greenberg, I.; Kaiser, M.; Gunina, A.; Ledesma, P.; Polifka, S.; Wiedner, K.; Mueller, C.W.; Glaser, B.; Ludwig, B. Substitution of mineral fertilizers with biogas digestate plus biochar increases physically stabilized soil carbon but not crop biomass in a field trial. Sci. Total Environ. 2019, 680, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Akimbekov, N.; Qiao, X.; Digel, I.; Abdieva, G.; Ualieva, P.; Zhubanova, A. The effect of leonardite-derived amendments on soil microbiome structure and potato yield. Agriculture 2020, 10, 147. [Google Scholar] [CrossRef]
- Al-Maliki, S.; Al-Mammory, H.; Scullion, J. Interactions between humic substances and organic amendments affecting soil biological properties and growth ofzea maysl. In the arid land region. Arid. Land Res. Manag. 2018, 32, 455–470. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fang, F.; Wei, J.; Wu, X.; Cui, R.; Li, G.; Zheng, F.; Tan, D. Humic acid fertilizer improved soil properties and soil microbial diversity of continuous cropping peanut: A three-year experiment. Sci. Rep. 2019, 9, 12014. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Wang, J.; Xiong, Z.; Kuzyakov, Y. Biochar stability in soil: Meta-analysis of decomposition and priming effects. GCB Bioenergy 2015, 8, 512–523. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.M.; Srivastava, A.K.; Cheraghi, M. Soil health and crop response of biochar: An updated analysis. Arch. Agron. Soil Sci. 2022, 62, 1–26. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management; Routledge: London, UK, 2015. [Google Scholar] [CrossRef]
- Liao, N.; Li, Q.; Zhang, W.; Zhou, G.; Ma, L.; Min, W.; Ye, J.; Hou, Z. Effects of biochar on soil microbial community composition and activity in drip-irrigated desert soil. Eur. J. Soil Biol. 2016, 72, 27–34. [Google Scholar] [CrossRef]
- Hoskins, B.; Wolf, A.; Wolf, N. Dry Matter Analysis. In Recommended Methods of Manure Analysis; Peters, J., Ed.; University of Wisconsin Cooperative Extension Publishing: Madison, WI, USA, 2003. [Google Scholar]
- ISO 11261; Soil Quality—Determination of Total Nitrogen—Modified Kjeldahl Method. International Organization for Standardization: Geneva, Switzerland, 1995.
- Egnér, H.A.; Riehm, H.; Domingo, W.R. Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nährstoffzustandes der Böden. II. Chemische Extraktionsmethoden zur Phosphor-und Kaliumbestimmung. K. Lantbr. Ann. 1960, 26, 199–215. [Google Scholar]
- EN_15749; Fertilizers—Determination of Sulfates Content Using Three Different Methods. European Standardization Organization CEN-CENELEC: Brussels, Belgium, 2009.
- ISO 9964-3; Water Quality—Determination of Sodium and Potassium—Part 3: Determination of Sodium and Potassium by Flame Emission Spectrometry. International Organization for Standardization: Geneva, Switzerland, 2001.
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeler, E.; Deiglmayr, K.; Tscherko, D.; Bru, D.; Philippot, L. Abundance of narG, nirS, nirK, and nosZ genes of denitrifying bacteria during primary successions of a glacier foreland. Appl. Environ. Microbiol. 2006, 72, 5957–5962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef] [PubMed]
- ISO 10390; Soil Quality—Determination of pH. International Organization for Standardization: Geneva, Switzerland, 2005.
- ISO 10694; Soil Quality—Determination of Organic and Total Carbon after Dry Combustion (Elementary Analysis). International Organization for Standardization: Geneva, Switzerland, 1995.
- ISO 13878; Soil Quality—Determination of Total Nitrogen Content by Dry Combustion (Elemental Analysis). International Organization for Standardization: Geneva, Switzerland, 1998.
- Campbell, C.D.; Chapman, S.J.; Cameron, C.M.; Davidson, M.S.; Potts, J.M. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl. Environ. Microbiol 2003, 69, 3593–3599. [Google Scholar] [CrossRef] [Green Version]
- ISO 20130; Soil Quality—Measurement of Enzyme Activity Patterns in Soil Samples Using Colorimetric Substrates in Micro-Well Plates. International Organization for Standardization: Geneva, Switzerland, 2018.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Hinkle, D.E.; Wiersma, W.; Jurs, S.G. Applied Statistics for the Behavioral Sciences; Houghton Mifflin: Boston, MA, USA, 2003. [Google Scholar]
- Rawat, J.; Saxena, J.; Sanwal, P. Biochar: A Sustainable Approach for Improving Plant Growth and Soil Properties. In Biochar—An Imperative Amendment for Soil and the Environment; Abrol, V., Sharma, P., Eds.; IntechOpen: London, UK, 2019; p. 124. [Google Scholar]
- Ndoung, O.C.N.; Figueiredo, C.C.; Ramos, M.L.G. A scoping review on biochar-based fertilizers: Enrichment techniques and agro-environmental application. Heliyon 2021, 7, e08473. [Google Scholar] [CrossRef]
- Xu, J.; Mohamed, E.; Li, Q.; Lu, T.; Yu, H.; Jiang, W. Effect of humic acid addition on buffering capacity and nutrient storage capacity of soilless substrates. Front. Plant Sci. 2021, 12, 644229. [Google Scholar] [CrossRef]
- Ampong, K.; Thilakaranthna, M.S.; Gorim, L.Y. Understanding the role of humic acids on crop performance and soil health. Front. Agron. 2022, 4, 10. [Google Scholar] [CrossRef]
- Holatko, J.; Hammerschmiedt, T.; Kintl, A.; Danish, S.; Skarpa, P.; Latal, O.; Baltazar, T.; Fahad, S.; Akca, H.; Taban, S.; et al. Effect of carbon-enriched digestate on the microbial soil activity. PLoS ONE 2021, 16, e0252262. [Google Scholar] [CrossRef]
- Gryń, G.; Gryń, G.; Paluszak, Z.; Olszewska, H.; Keutgen, A.J. Chemical and microbiological properties of luvisol after addition of post-fermentation residue. J. Elem. 2020, 25, 701–716. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, J.; Huo, L.; CLi, Y.; Li, X.; Xia, L.; Zhou, Z.; Zhang, M.; Li, B. Humic acids derived from leonardite to improve enzymatic activities and bioavailability of nutrients in a calcareous soil. Int. J. Agric. Biol. Eng. 2020, 13, 200–205. [Google Scholar] [CrossRef]
- Egene, C.E.; Sigurnjak, I.; Regelink, I.C.; Schoumans, O.F.; Adani, F.; Michels, E.; Sleutel, S.; Tack, F.M.G.; Meers, E. Solid fraction of separated digestate as soil improver: Implications for soil fertility and carbon sequestration. J. Soils Sediments 2020, 21, 678–688. [Google Scholar] [CrossRef]
- Holatko, J.; Hammerschmiedt, T.; Datta, R.; Baltazar, T.; Kintl, A.; Latal, O.; Pecina, V.; Sarec, P.; Novak, P.; Balakova, L.; et al. Humic acid mitigates the negative effects of high rates of biochar application on microbial activity. Sustainability 2020, 12, 9524. [Google Scholar] [CrossRef]
- Fascella, G.; Montoneri, E.; Rouphael, Y. Biowaste-derived humic-like substances improve growth and quality of orange jasmine (Murraya paniculata L. Jacq.) plants in soilless potted culture. Resources 2021, 10, 80. [Google Scholar] [CrossRef]
- Tan, F.; Zhu, Q.; Guo, X.; He, L. Effects of digestate on biomass of a selected energy crop and soil properties. J. Sci. Food Agric. 2021, 101, 927–936. [Google Scholar] [CrossRef]
- Allison, S.D. Soil minerals and humic acids alter enzyme stability: Implications for ecosystem processes. Biogeochemistry 2006, 81, 361–373. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, S.-T.; Monreal, C.M.; McLaughlin, N.B.; Zhao, B.-P.; Liu, J.-H.; Hao, G.-C. Bentonite-humic acid improves soil organic carbon, microbial biomass, enzyme activities and grain quality in a sandy soil cropped to maize (Zea mays L.) in a semi-arid region. J. Integr. Agric. 2022, 21, 208–221. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Shah, T.; Khan, S.; Shah, Z. Soil respiration, ph and ec as influenced by biochar. Soil Environ. 2017, 36, 77–83. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Bahar, M.M.; Sarkar, B.; Donne, S.W.; Ok, Y.S.; Palansooriya, K.N.; Kirkham, M.B.; Chowdhury, S.; Bolan, N. Biochar and its importance on nutrient dynamics in soil and plant. Biochar 2020, 2, 379–420. [Google Scholar] [CrossRef]
- Santos, L.L.d.; Lacerda, J.J.J.; Zinn, Y.L. Partição de substâncias húmicas em solos brasileiros. Rev. Bras. Ciência Solo 2013, 37, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Kaźmierczak, B.; Wystalska, K.; Malińska, K.; Włodarczyk, R.; Chajczyk, O.; Kutyłowska, M.; Piekarska, K.; Jadwiszczak, P. Effects of pyrolysis parameters on the yield and properties of biochar from pelletized sunflower husk. E3S Web Conf. 2018, 44, 197. [Google Scholar] [CrossRef]
- Wiedner, K.; Glaser, B. Biochar-fungi interactions in soils. In Biochar and Soil Biota; CRC Press: Boca Raton, FL, USA, 2013; p. 10. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Variant | Abbrev. | Digestate per Barrel [L] | Biochar per Barrel [kg] | Humac per Barrel [kg] |
|---|---|---|---|---|
| Digestate | D | 10 | - | - |
| Digestate + Humac | D + H | 10 | - | 0.1 |
| Digestate + biochar | D + B | 10 | 4 | - |
| Digestate + biochar + Humac | D + B + H | 10 | 4 | 0.1 |
| Variants | DM | Ntot | P | S | K | amoA | nirS | 16S rDNA |
|---|---|---|---|---|---|---|---|---|
| % | % | g/kg | g/kg | g/kg | cps/g | cps/g | cps/g | |
| D | 5.96 | 0.65 | 0.76 | 0.85 | 1.35 | 3.81 × 105 | 3.15 × 106 | 5.66 × 1010 |
| D + H | 6.77 | 0.50 | 0.95 | 1.10 | 1.80 | 2.56 × 105 | 1.67 × 106 | 5.03 × 1010 |
| D + B | 15.00 | 0.68 | 2.09 | 2.27 | 3.63 | 1.21 × 106 | 3.05 × 107 | 4.40 × 1011 |
| D + B + H | 29.38 | 0.76 | 2.32 | 5.14 | 2.00 | 5.06 × 105 | 1.70 × 107 | 2.21 × 1011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holatko, J.; Hammerschmiedt, T.; Latal, O.; Kintl, A.; Mustafa, A.; Baltazar, T.; Malicek, O.; Brtnicky, M. Deciphering the Effectiveness of Humic Substances and Biochar Modified Digestates on Soil Quality and Plant Biomass Accumulation. Agronomy 2022, 12, 1587. https://doi.org/10.3390/agronomy12071587
Holatko J, Hammerschmiedt T, Latal O, Kintl A, Mustafa A, Baltazar T, Malicek O, Brtnicky M. Deciphering the Effectiveness of Humic Substances and Biochar Modified Digestates on Soil Quality and Plant Biomass Accumulation. Agronomy. 2022; 12(7):1587. https://doi.org/10.3390/agronomy12071587
Chicago/Turabian StyleHolatko, Jiri, Tereza Hammerschmiedt, Oldrich Latal, Antonin Kintl, Adnan Mustafa, Tivadar Baltazar, Ondrej Malicek, and Martin Brtnicky. 2022. "Deciphering the Effectiveness of Humic Substances and Biochar Modified Digestates on Soil Quality and Plant Biomass Accumulation" Agronomy 12, no. 7: 1587. https://doi.org/10.3390/agronomy12071587
APA StyleHolatko, J., Hammerschmiedt, T., Latal, O., Kintl, A., Mustafa, A., Baltazar, T., Malicek, O., & Brtnicky, M. (2022). Deciphering the Effectiveness of Humic Substances and Biochar Modified Digestates on Soil Quality and Plant Biomass Accumulation. Agronomy, 12(7), 1587. https://doi.org/10.3390/agronomy12071587