Potential for Mycorrhizae-Assisted Phytoremediation of Phosphorus for Improved Water Quality



Abstract
1. Introduction
1.1. Worldwide Freshwater Quality Threats
1.2. Relatively New Field of Myco-Phytoremediation
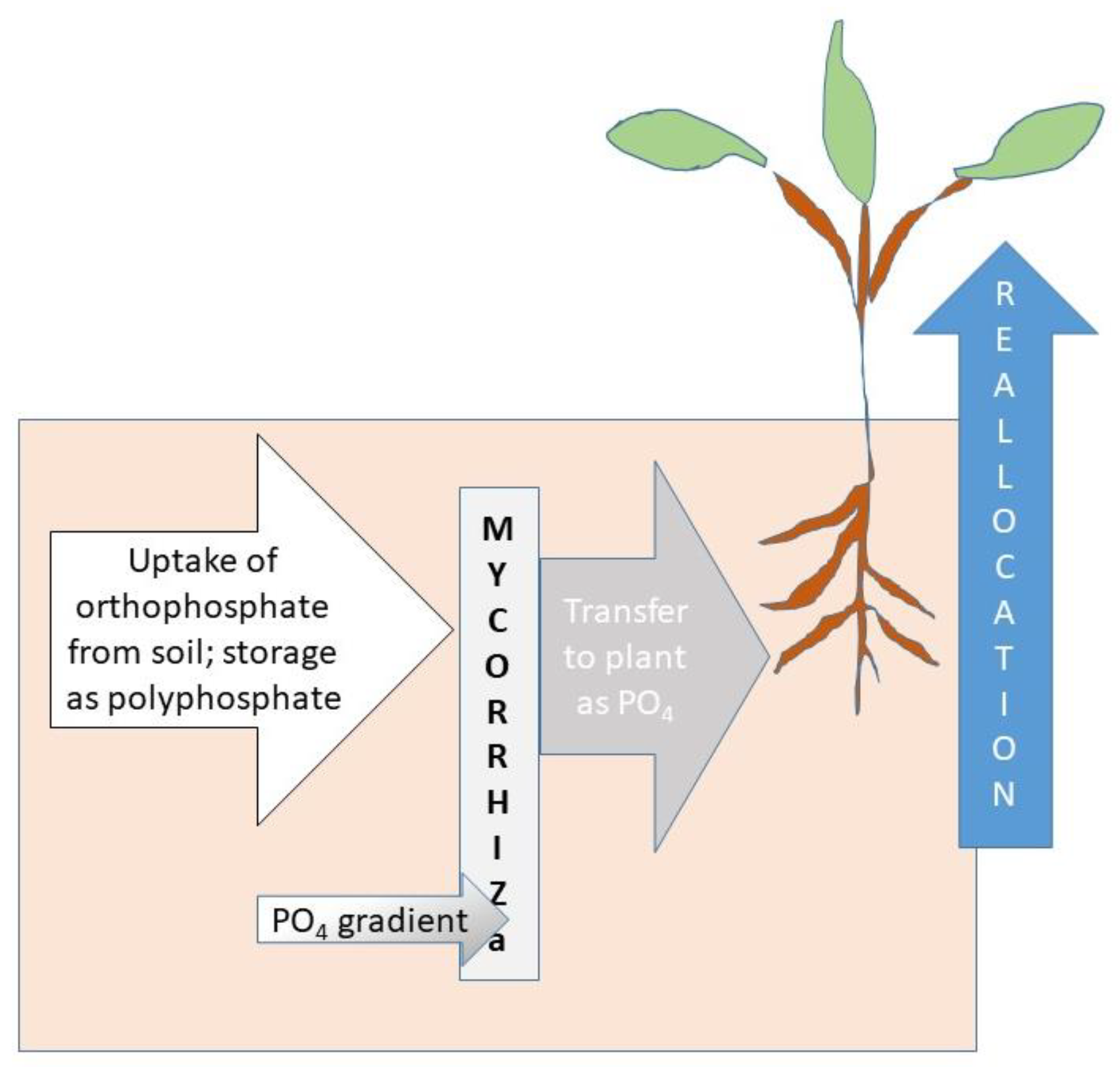
1.3. Mycorrhizae
2. The Phosphorus Problem
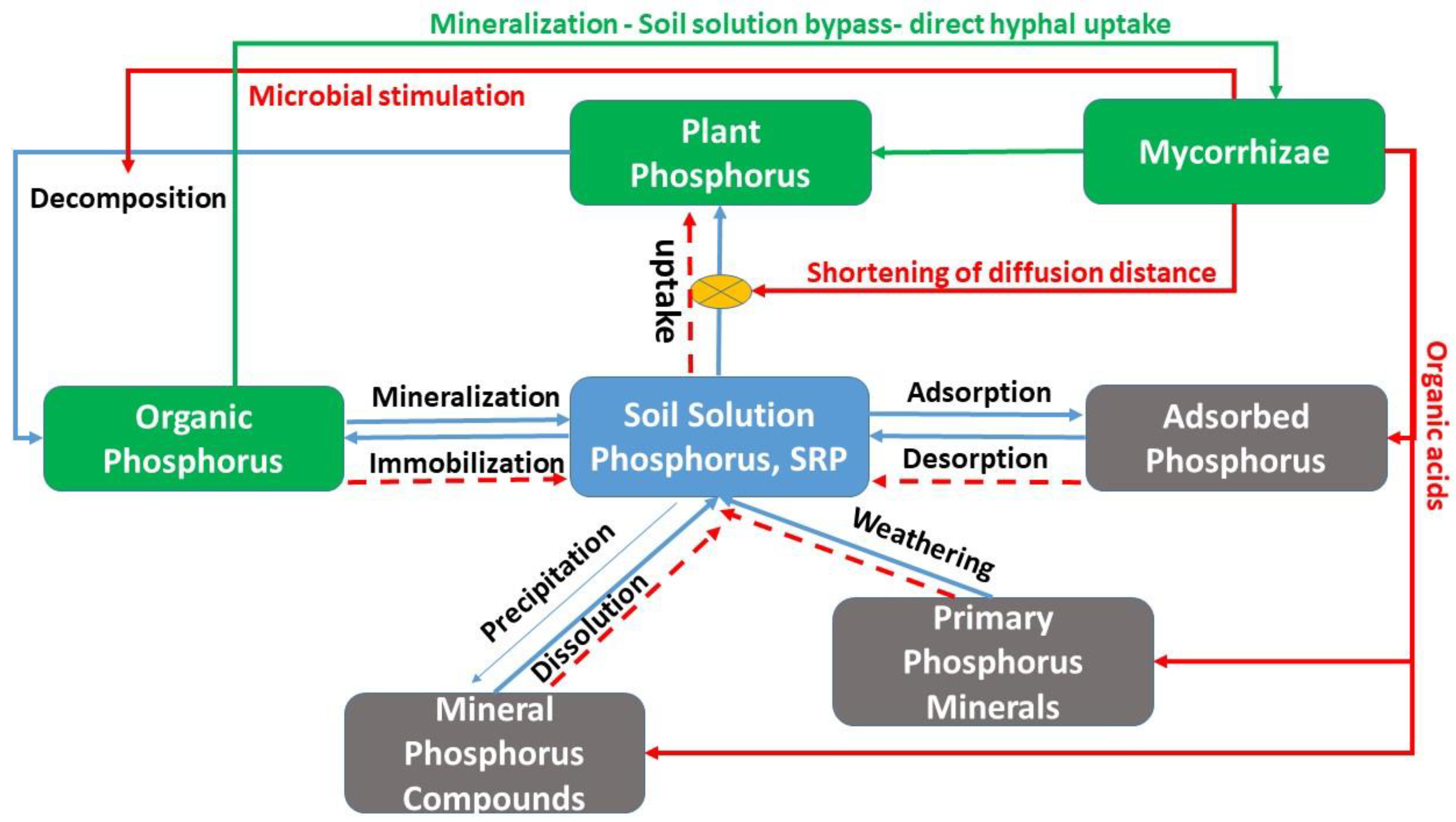
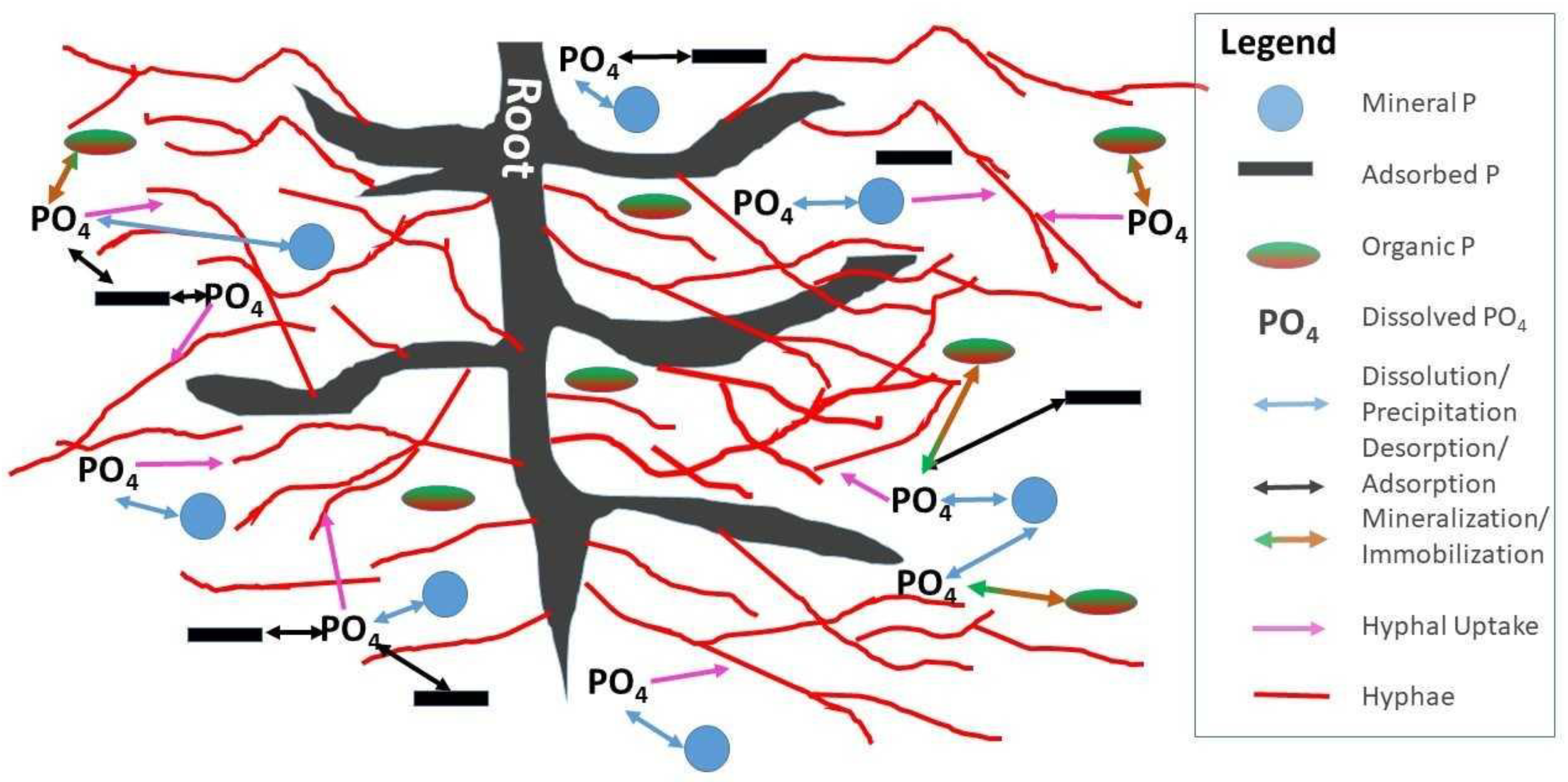
3. Processes in the Phosphorus Cycle Where Mycorrhizae Affect P Availability
4. Mycorrhizae, Landscapes and Soils
5. Riparian Buffers
6. Green Stormwater Infrastructure
7. Summary of Research Results from the Literature
8. Research Needs
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMF | Arbuscular mycorrhizal fungi also known as Endomycorrhizae |
| BMP | Best management practices |
| CW | Constructed wetlands |
| ECM | Ectomycorrhizal fungi |
| NPS | Non-point source pollution |
| P | Phosphorus |
| Pi | Inorganic phosphorus |
| PP | Particulate phosphorus |
| SRP | Soluble reactive phosphorus, orthophosphate |
| TP | Total phosphorus |
References
- Michalak, A.M.; Anderson, E.J.; Beletsky, D.; Boland, S.; Bosch, N.S.; Bridgeman, T.B.; Chaffin, J.D.; Cho, K.; Confesor, R.; Daloglu, I.; et al. Record-setting algal bloom in Lake Erie caused by agricultural and meteorological trends consistent with expected future conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6448–6452. [Google Scholar] [CrossRef]
- Albert, J.S.; Destouni, G.; Duke-Sylvester, S.M.; Magurran, A.E.; Oberdorff, T.; Reis, R.E.; Winemiller, K.O.; Ripple, W.J. Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 2020. [Google Scholar] [CrossRef] [PubMed]
- Qadri, H.; Bhat, R. The Concerns for Global Sustainability of Freshwater Ecosystems. In Freshwater Pollution Dynamics and Remediation, 1st ed.; Qadri, H., Bhat, R., Mehood, M., Dar, G., Eds.; Springer: Singapore, 2020; pp. 1–13. [Google Scholar]
- Tickner, D.; Opperman, J.J.; Abell, R.; Acreman, M.; Arthington, A.H.; Bunn, S.E.; Cooke, S.J.; Dalton, J.; Darwall, W.; Edwards, G.; et al. Bending the Curve of Global Freshwater Biodiversity Loss: An Emergency Recovery Plan. BioScience 2020, 70, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, A.R. Water reuse, food production and public health: Adopting transdisciplinary, systems-based approaches to achieve water and food security in a changing climate. Environ. Res. 2019, 171, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef]
- Mekonnen, M.M.; Hoekstra, A.Y. Global Anthropogenic Phosphorus Loads to Freshwater and Associated Grey Water Footprints and Water Pollution Levels: A High-Resolution Global Study. Water Resour. Res. 2018, 54, 345–358. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Cao, X.; Wang, Y.; He, J.; Luo, X.; Zheng, Z. Phosphorus mobility among sediments, water and cyanobacteria enhanced by cyanobacteria blooms in eutrophic Lake Dianchi. Environ. Pollut. 2016, 219, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; King, K.W.; Williams, M.R. What is causing the harmful algal blooms in Lake Erie? J. Soil Water Conserv. 2015, 70, 27A–29A. [Google Scholar] [CrossRef]
- Troy, A.; Wang, D.; Capen, D.; O’Neil-Dunne, J.; MacFaden, S. Updating the Lake Champlain Basin Land Use Data to Improve Prediction of Phosphorus Loading; Scientific Investigations Report: Burlington, VT, USA, 2017. [Google Scholar]
- Li, C.; Dong, Y.; Lei, Y.; Wu, D.; Xu, P. Removal of low concentration nutrients in hydroponic wetlands integrated with zeolite and calcium silicate hydrate functional substrates. Ecol. Eng. 2015, 82, 442–450. [Google Scholar] [CrossRef]
- Ojoawo, S.O.; Udayakumar, G.; Naik, P. Phytoremediation of Phosphorus and Nitrogen with Canna x generalis Reeds in Domestic Wastewater through NMAMIT Constructed Wetland. Aquat. Procedia 2015, 4, 349–356. [Google Scholar] [CrossRef]
- Hunter, P.D.; Tyler, A.N.; Gilvear, D.J.; Willby, N.J. Using Remote Sensing to Aid the Assessment of Human Health Risks from Blooms of Potentially Toxic Cyanobacteria. Environ. Sci. Technol. 2009, 43, 2627–2633. [Google Scholar] [CrossRef] [PubMed]
- Roy, E.D. Phosphorus recovery and recycling with ecological engineering: A review. Ecol. Eng. 2017, 98, 213–227. [Google Scholar] [CrossRef]
- Li, X.; Zhang, W.; Zhao, C.; Li, H.; Shi, R. Nitrogen interception and fate in vegetated ditches using the isotope tracer method: A simulation study in northern China. Agric. Water Manag. 2020, 228, 105893. [Google Scholar] [CrossRef]
- Anastasi, A.; Tigini, V.; Varese, G.C. The Bioremediation Potential of Different Ecophysiological Groups of Fungi. In Fungi as Bioremediators; Goltapeh, E.M., Danesh, Y.R., Varma, A., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 32, pp. 29–49. [Google Scholar]
- Dogan, I.; Ozyigit, I.I. Plant-Microbe Interactions in Phytoremediation. In Soil Remediation in Plants, Prospects and Challenges, 1st ed.; Hakeem, K.R., Sabir, M., Öztürk, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Zhang, B.Y.; Zheng, J.S.; Sharp, R.G. Phytoremediation in Engineered Wetlands: Mechanisms and Applications. Procedia Environ. Sci. 2010, 2, 1315–1325. [Google Scholar] [CrossRef]
- Gotcher, M.J.; Zhang, H.; Schroder, J.L.; Payton, M.E. Phytoremediation of Soil Phosphorus with Crabgrass. Agron. J. 2014, 106, 528–536. [Google Scholar] [CrossRef]
- Khan, A.G. Mycorrhizoremediation—An enhanced form of phytoremediation. J. Zhejiang Univ. Sci. B 2006, 7, 503–514. [Google Scholar] [CrossRef]
- Mäder, P.; Kaiser, F.; Adholeya, A.; Singh, R.; Uppal, H.S.; Sharma, A.K.; Srivastava, R.; Sahai, V.; Aragno, M.; Wiemken, A.; et al. Inoculation of root microorganisms for sustainable wheat–rice and wheat–black gram rotations in India. Soil Biol. Biochem. 2011, 43, 609–619. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Yang, M.; Yan, L.; Kang, Z.; Xiao, Y.; Tang, P.; Ye, L.; Zhang, B.; Zou, J.; et al. Tuber borchii Shapes the Ectomycorrhizosphere Microbial Communities of Corylus avellana. Mycobiology 2019, 47, 180–190. [Google Scholar] [CrossRef]
- Shoaib, A.; Aslam, N.; Aslam, N. Myco and Phyto Remediation of Heavy Metals from Aqueous Solution. Online J. Sci. Technol. 2012, 2, 34–41. [Google Scholar]
- Neagoe, A.; Tenea, G.; Cucu, N.; Ion, S.; Iordache, V. Coupling Nicotiana tabaccum Transgenic Plants with Rhizophagus irregularis for Phytoremediation of Heavy Metal Polluted Areas. Rev. Chim. 2017, 68, 789–795. [Google Scholar] [CrossRef]
- Govarthanan, M.; Mythili, R.; Selvankumar, T.; Kamala-Kannan, S.; Kim, H. Myco-phytoremediation of arsenic- and lead-contaminated soils by Helianthus annuus and wood rot fungi, Trichoderma sp. isolated from decayed wood. Ecotoxicol. Environ. Saf. 2018, 151, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Blagodatsky, S.; Ehret, M.; Rasche, F.; Hutter, I.; Birner, R.; Dzomeku, B.; Neya, O.; Cadisch, G.; Wünsche, J. Myco-phytoremediation of mercury polluted soils in Ghana and Burkina Faso. In Proceedings of the EGU General Assembly Conference, Sharing Geoscience Online Abstracts, Online, 4–8 May 2020. [Google Scholar]
- Ramakrishan, K.G. Bhuvaneswari Influence on Different Types of Mycorrhizal Fungi on Crop Productivity in Ecosystem. Int. Lett. Nat. Sci. 2015, 38, 9–15. [Google Scholar] [CrossRef]
- Sanders, F.E.; Tinker, P.B. Phosphate flow into mycorrhizal roots. Pestic. Sci. 1973, 4, 385–395. [Google Scholar] [CrossRef]
- Rillig, M.C.; Sosa-Hernández, M.A.; Roy, J.; Aguilar-Trigueros, C.A.; Vályi, K.; Lehmann, A. Towards an Integrated Mycorrhizal Technology: Harnessing Mycorrhiza for Sustainable Intensification in Agriculture. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.G.; O’Neill, R.V.; Norby, R.J. Hierarchy theory as a guide to mycorrhizal research on large-scale problems. Environ. Pollut. 1991, 73, 271–284. [Google Scholar] [CrossRef]
- Zalewski, M. Ecohydrology—The scientific background to use ecosystem properties as management tools toward sustainability of water resources. Ecol. Eng. 2000, 16, 1–8. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Michener, W. Win-Win Ecology: How the Earth’s Species Can Survive in the Midst of Human Enterprise. Restor. Ecol. 2004, 12, 306–307. [Google Scholar] [CrossRef]
- Bücking, H.; Liepold, E.; Ambilwade, P. The Role of the Mycorrhizal Symbiosis in Nutrient Uptake of Plants and the Regulatory Mechanisms Underlying These Transport Processes. Plant Sci. 2012. [Google Scholar] [CrossRef]
- Lin, C.; Wang, Y.; Liu, M.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 2020, 10, 12260. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of Arbuscular Mycorrhizas in Plant Phosphorus Nutrition: Interactions between Pathways of Phosphorus Uptake in Arbuscular Mycorrhizal Roots Have Important Implications for Understanding and Manipulating Plant Phosphorus Acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.J.; Jones, M.D.; Kranabetter, J.M. Ectomycorrhizae and tree seedling nitrogen nutrition in forest restoration. New For. 2015, 46, 747–771. [Google Scholar] [CrossRef]
- Becquer, A.; Trap, J.; Irshad, U.; Ali, M.A.; Claude, P. From soil to plant, the journey of P through trophic relationships and ectomycorrhizal association. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.; Durall, D.M.; Tinker, P.B. A comparison of arbuscular and ectomycorrhizal Eucalyptus coccifera: Growth response, phosphorus uptake efficiency and external hyphal production. New Phytol. 1998, 140, 125–134. [Google Scholar] [CrossRef]
- Djighaly, P.I.; Ndiaye, S.; Diarra, A.M.; Dramé, F.A. Inoculation with arbuscular mycorrhizal fungi improves salt tolerance in C. glauca (Sieb). J. Mater. Environ. Sci. 2020, 11, 1616–1625. [Google Scholar]
- Djighaly, P.I.; Ngom, D.; Diagne, N.; Fall, D.; Ngom, M.; Diouf, D.; Hocher, V.; Laplaze, L.; Champion, A.; Farrant, J.M.; et al. Effect of Casuarina Plantations Inoculated with Arbuscular Mycorrhizal Fungi and Frankia on the Diversity of Herbaceous Vegetation in Saline Environments in Senegal. Diversity 2020, 12, 293. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Asmelash, F.; Bekele, T.; Birhane, E. The Potential Role of Arbuscular Mycorrhizal Fungi in the Restoration of Degraded Lands. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Ortaş, I.; Rafique, M. The Mechanisms of Nutrient Uptake by Arbuscular Mycorrhizae. In Mycorrhiza—Nutrient Uptake, Biocontrol, Ecorestoration; Varma, A., Prasad, R., Tuteja, N., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–19. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Policelli, N.; Horton, T.R.; Hudon, A.T.; Patterson, T.; Bhatnagar, J.M. Back to roots: The role of ectomycorrhizal fungi in boreal and temperate forest restoration. Front. For. Glob. Chang. 2020, 3, 97. [Google Scholar] [CrossRef]
- Nelson, L.L.; Allen, E.B. Restoration of Stipa pulchra Grasslands: Effects of Mycorrhizae and Competition from Avena barbata. Restor. Ecol. 1993, 1, 40–50. [Google Scholar] [CrossRef]
- Policelli, N.; Horton, T.R.; García, R.A.; Naour, M.; Pauchard, A.; Nuñez, M.A. Native and non-native trees can find compatible mycorrhizal partners in each other’s dominated areas. Plant Soil 2020, 454, 285–297. [Google Scholar] [CrossRef]
- Deguchi, S.; Uozumi, S.; Tuono, E.; Kaneko, M.; Tawraya, K. Arbuscular mycorrhizal colonization increases phosphorus uptake and growth of corn in a white clover living mulch system. Soil Sci. Plant Nutr. 2012, 58, 169–172. [Google Scholar] [CrossRef]
- Ishee, E.R.; Ross, D.S.; Garvey, K.M.; Bourgault, R.R.; Ford, C.R. Phosphorus Characterization and Contribution from Eroding Streambank Soils of Vermont’s Lake Champlain Basin. J. Environ. Qual. 2015, 44, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Hesketh, N. Brookes Development of an indicator for risk of phosphorus leaching. Environ. Qual. 2000, 29, 105–110. [Google Scholar] [CrossRef]
- Rowe, H.; Withers, P.J.A.; Baas, P.; Chan, N.I.; Doody, D.; Holiman, J.; Jacobs, B.; Li, H.; MacDonald, G.K.; McDowell, R.; et al. Integrating legacy soil phosphorus into sustainable nutrient management strategies for future food, bioenergy and water security. Nutr. Cycl. Agroecosystems 2016, 104, 393–412. [Google Scholar] [CrossRef]
- Hamilton, S.K. Biogeochemical time lags may delay responses of streams to ecological restoration. Freshw. Biol. 2012, 57, 43–57. [Google Scholar] [CrossRef]
- Meals, D.W.; Dressing, S.A.; Davenport, T.E. Lag Time in Water Quality Response to Best Management Practices: A Review. J. Environ. Qual. 2010, 39, 85–96. [Google Scholar] [CrossRef]
- Sharpley, A.; Jarvie, H.P.; Buda, A.; May, L.; Spears, B.; Kleinman, P. Phosphorus Legacy: Overcoming the Effects of Past Management Practices to Mitigate Future Water Quality Impairment. J. Environ. Qual. 2013, 42, 1308–1326. [Google Scholar] [CrossRef]
- Jarvie, H.P.; Johnson, L.T.; Sharpley, A.N.; Smith, D.R.; Baker, D.B.; Bruulsema, T.W.; Confesor, R. Increased Soluble Phosphorus Loads to Lake Erie: Unintended Consequences of Conservation Practices? J. Environ. Qual. 2017, 46, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Gruau, G.; Dupas, R.; Rumpel, C.; Crème, A.; Fovet, O.; Gascuel-Odoux, C.; Jeanneau, L.; Humbert, G.; Petitjean, P. Release of dissolved phosphorus from riparian wetlands: Evidence for complex interactions among hydroclimate variability, topography and soil properties. Sci. Total Environ. 2017, 598, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Baker, D.E.; Pionke, H.B.; Kunishi, H.M. Soil Tests for Estimating Labile, Soluble, and Algae-Available Phosphorus in Agricultural Soils. J. Environ. Qual. 1985, 14, 341–348. [Google Scholar] [CrossRef]
- Nezat, C.A.; Blum, J.D.; Yanai, R.D.; Park, B.B. Mineral Sources of Calcium and Phosphorus in Soils of the Northeastern United States. Soil Sci. Soc. Am. J. 2008, 72, 1786–1794. [Google Scholar] [CrossRef]
- Pote, D.H.; Daniel, T.C.; Nichols, D.J.; Moore, P.A.; Miller, D.M.; Edwards, D.R. Seasonal and Soil-Drying Effects on Runoff Phosphorus Relationships to Soil Phosphorus. Soil Sci. Soc. Am. J. 1999, 63, 1006–1012. [Google Scholar] [CrossRef]
- Sharpley, A.N. Soil phosphorus dynamics: Agronomic and environmental impacts. Ecol. Eng. 1995, 5, 261–279. [Google Scholar] [CrossRef]
- Al-Abbas, A.H.; Barber, S.A. A Soil Test for Phosphorus Based Upon Fractionation of Soil Phosphorus: II. Development of the Soil Test. Soil Sci. Soc. Am. J. 1964, 28, 221–224. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; Edwards, M.; Elser, J.J. A transgenic approach to enhance phosphorus use efficiency in crops as part of a comprehensive strategy for sustainable agriculture. Chemosphere 2011, 84, 840–845. [Google Scholar] [CrossRef]
- Sharpley, A.N.S.R. Phosphorus in agriculture and its environmental implications. In Phosphorus Loss from Soil to Water; Tunney, H., Carton, O.T., Brookes, P.C., Johnston, A.E., Eds.; CAB International Press: Cambridge, UK, 1997; pp. 1–54. [Google Scholar]
- Macintosh, K.A.; Doody, D.G.; Withers, P.J.A.; McDowell, R.W.; Smith, D.R.; Johnson, L.T.; Bruulsema, T.W.; O’Flaherty, V.; McGrath, J.W. Transforming soil phosphorus fertility management strategies to support the delivery of multiple ecosystem services from agricultural systems. Sci. Total Environ. 2019, 649, 90–98. [Google Scholar] [CrossRef]
- Jordan-Meille, L.; Rubæk, G.H.; Ehlert, P.A.I.; Genot, V.; Hofman, G.; Goulding, K.; Recknagel, J.; Provolo, G.; Barraclough, P. An overview of fertilizer-P recommendations in Europe: Soil testing, calibration and fertilizer recommendations. Soil Use Manag. 2012, 28, 419–435. [Google Scholar] [CrossRef]
- Pierzynski, G.M.; Logan, T.J. Crop, Soil, and Management Effects on Phosphorus Soil Test Levels: A Review. J. Prod. Agric. 1993, 6, 513–520. [Google Scholar] [CrossRef]
- Schröder, J.J.; Smit, A.L.; Cordell, D.; Rosemarin, A. Improved phosphorus use efficiency in agriculture: A key requirement for its sustainable use. Chemosphere 2011, 84, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Castán, E.; Satti, P.; González-Polo, M.; Iglesias, M.C.; Mazzarino, M.J. Managing the value of composts as organic amendments and fertilizers in sandy soils. Agric. Ecosyst. Environ. 2016, 224, 29–38. [Google Scholar] [CrossRef]
- Jakobsen, I.; Rosendahl, L. Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytol. 1990, 115, 77–83. [Google Scholar] [CrossRef]
- Li, X.L.; George, E.; Marschner, H. Extension of the phosphorus depletion zone in VA-mycorrhizal white clover in a calcareous soil. Plant Soil 1991, 136, 41–48. [Google Scholar] [CrossRef]
- Bolan, N.S. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1991, 134, 189–207. [Google Scholar] [CrossRef]
- Plassard, C.; Dell, B. Phosphorus nutrition of mycorrhizal trees. Tree Physiol. 2010, 30, 1129–1139. [Google Scholar] [CrossRef]
- Blum, J.D.; Klaue, A.; Nezat, C.A.; Driscoll, C.T.; Johnson, C.E.; Siccama, T.G.; Eagar, C.; Fahey, T.J.; Likens, G.E. Mycorrhizal weathering of apatite as an important calcium source in base-poor forest ecosystems. Nature 2002, 417, 729–731. [Google Scholar] [CrossRef]
- Schneider, K.D.; Martens, J.R.T.; Zvomuya, F.; Reid, D.K.; Fraser, T.D.; Lynch, D.H.; O’Halloran, I.P.; Wilson, H.F. Options for Improved Phosphorus Cycling and Use in Agriculture at the Field and Regional Scales. J. Environ. Qual. 2019, 48, 1247–1264. [Google Scholar] [CrossRef]
- Hamel, C. Mycorrhizae in Crop Production; CRC Press: Boca Rotan, FL, USA, 2007. [Google Scholar]
- Liu, C.; Liu, F.; Ravnskov, S.; Rubæk, G.H.; Sun, Z.; Andersen, M.N. Impact of Wood Biochar and Its Interactions with Mycorrhizal Fungi, Phosphorus Fertilization and Irrigation Strategies on Potato Growth. J. Agron. Crop Sci. 2017, 203, 131–145. [Google Scholar] [CrossRef]
- Funamoto, R.; Saito, K.; Oyaizu, H.; Saito, M.; Aono, T. Simultaneous in situ detection of alkaline phosphatase activity and polyphosphate in arbuscules within arbuscular mycorrhizal roots. Funct. Plant Biol. 2007, 34, 803–810. [Google Scholar] [CrossRef]
- Weidner, S.; Koller, R.; Latz, E.; Kowalchuk, G.; Bonkowski, M.; Scheu, S.; Jousset, A. Bacterial diversity amplifies nutrient-based plant–soil feedbacks. Funct. Ecol. 2015, 29, 1341–1349. [Google Scholar] [CrossRef]
- Sato, T.; Ezawa, T.; Cheng, W.; Tawaraya, K. Release of acid phosphatase from extraradical hyphae of arbuscular mycorrhizal fungus Rhizophagus clarus. Soil Sci. Plant Nutr. 2015, 61, 269–274. [Google Scholar] [CrossRef]
- Battini, F.; Grønlund, M.; Agnolucci, M.; Giovannetti, M.; Jakobsen, I. Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria. Sci. Rep. 2017, 7, 4686. [Google Scholar] [CrossRef] [PubMed]
- Ulén, B.; Aronsson, H.; Bechmann, M.; Krogstad, T.; ØYgarden, L.; Stenberg, M. Soil tillage methods to control phosphorus loss and potential side-effects: A Scandinavian review. Soil Use Manag. 2010, 26, 94–107. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef]
- Rillig, M.C.; Steinberg, P.D. Glomalin production by an arbuscular mycorrhizal fungus: A mechanism of habitat modification? Soil Biol. Biochem. 2002, 34, 1371–1374. [Google Scholar] [CrossRef]
- Tisdall, J.M. Possible role of soil microorganisms in aggregation in soils. Plant Soil 1994, 159, 115–121. [Google Scholar] [CrossRef]
- Caravaca, F.; Garcia, C.; Hernández, M.T.; Roldán, A. Aggregate stability changes after organic amendment and mycorrhizal inoculation in the afforestation of a semiarid site with Pinus halepensis. Appl. Soil Ecol. 2002, 19, 199–208. [Google Scholar] [CrossRef]
- Wubs, E.R.J.; Van Der Putten, W.H.; Bosch, M.; Bezemer, T.M. Soil inoculation steers restoration of terrestrial ecosystems. Nat. Plants 2016, 2, 16107. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Kaul, H.-P.; Vollmann, J.; Eitzinger, J.; Wenzel, W. Developing phosphorus-efficient crop varieties—An interdisciplinary research framework. Field Crops Res. 2014, 162, 87–98. [Google Scholar] [CrossRef]
- Mendes, F.F.; Guimarães, L.J.M.; Souza, J.C.; Guimarães, P.E.O.; Magalhaes, J.V.; Garcia, A.A.F.; Parentoni, S.N.; Guimaraes, C.T. Genetic Architecture of Phosphorus Use Efficiency in Tropical Maize Cultivated in a Low-P Soil. Crop Sci. 2014, 54, 1530–1538. [Google Scholar] [CrossRef]
- Frossard, E.; Bünemann, E.K.; Gunst, L.; Oberson, A.; Schärer, M.; Tamburini, F. Fate of Fertilizer P in Soils—The Organic Pathway. In Phosphorus in Agriculture: 100% Zero; Schnug, E., De Kok, L.J., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 41–61. [Google Scholar]
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Parentoni, S.N.; Mendes, F.F.; Guimarães, L.J.M. Breeding for Phosphorus Use Efficiency. In Plant Breeding for Abiotic Stress Tolerance; Fritsche-Neto, R., Borém, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 67–85. [Google Scholar]
- Dörmann, P. Galactolipids in Plant Membranes. eLS 2013. [Google Scholar] [CrossRef]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Timonen, S.; Marschner, P. Mycorrhizosphere Concept. In Microbial Activity in the Rhizoshere; Mukerji, K.G., Manoharachary, C., Singh, J., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 155–172. [Google Scholar]
- Sandoz, F.A.; Bindschedler, S.; Dauphin, B.; Farinelli, L.; Grant, J.R.; Hervé, V. Biotic and abiotic factors shape arbuscular mycorrhizal fungal communities associated with the roots of the widespread fern Botrychium lunaria (Ophioglossaceae). Environ. Microbiol. Rep. 2020, 12, 342–354. [Google Scholar] [CrossRef]
- Lambert, D.H.; Weidensaul, T.C. Element Uptake by Mycorrhizal Soybean from Sewage-Sludge-Treated Soil. Soil Sci. Soc. Am. J. 1991, 55, 393–398. [Google Scholar] [CrossRef]
- Asghari, H.R.; Chittleborough, D.J.; Smith, F.A.; Smith, S.E. Influence of Arbuscular Mycorrhizal (AM) Symbiosis on Phosphorus Leaching through Soil Cores. Plant Soil 2005, 275, 181–193. [Google Scholar] [CrossRef]
- Liu, A.; Hamel, C.; Begna, S.H.; Ma, B.L.; Smith, D.L. Soil phosphorus depletion capacity of arbuscular mycorrhizae formed by maize hybrids. Can. J. Soil Sci. 2003, 83, 337–342. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi—Current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil Dordr. 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Cui, L.-H.; Zhu, X.-Z.; Ouyang, Y.; Chen, Y.; Yang, F.-L. Total Phosphorus Removal from Domestic Wastewater with Cyperus Alternifolius in Vertical-Flow Constructed Wetlands at the Microcosm Level. Int. J. Phytoremediation 2011, 13, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Torit, J.; Siangdung, W.; Thiravetyan, P. Phosphorus removal from domestic wastewater by Echinodorus cordifolius L. J. Environ. Sci. Health Part A 2012, 47, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Komada, M.; Ookuma, A.; Itahashi, S.; Banzai, K. Purification performance of a shallow free-water-surface constructed wetland receiving secondary effluent for about 5 years. Ecol. Eng. 2014, 69, 126–133. [Google Scholar] [CrossRef]
- Kochian, L.V.; Hoekenga, O.A.; Piñeros, M.A. How Do Crop Plants Tolerate Acid Soils? Mechanisms of Aluminum Tolerance and Phosphorous Efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef] [PubMed]
- Bünemann, E.K. Assessment of gross and net mineralization rates of soil organic phosphorus—A review. Soil Biol. Biochem. 2015, 89, 82–98. [Google Scholar] [CrossRef]
- Bolduc, A.; Hijri, M. The Use of Mycorrhizae to Enhance Phosphorus Uptake: A Way Out the Phosphorus Crisis. J. Biofertilizers Biopestic. 2011, 2. [Google Scholar] [CrossRef]
- Cao, H.-X.; Zhang, Z.-B.; Sun, C.-X.; Shao, H.-B.; Song, W.-Y.; Xu, P. Chromosomal Location of Traits Associated with Wheat Seedling Water and Phosphorus Use Efficiency under Different Water and Phosphorus Stresses. Int. J. Mol. Sci. 2009, 10, 4116–4136. [Google Scholar] [CrossRef]
- Abbott, L.K.; Robson, A.D. Colonization of the Root System of Subterranean Clover by Three Species of Vesicular-Arbuscular Mycorrhizal Fungi. New Phytol. 1984, 96, 275–281. [Google Scholar] [CrossRef]
- Asghari, H.R.; Cavagnaro, T.R. Arbuscular mycorrhizas enhance plant interception of leached nutrients. Funct. Plant Biol. 2011, 38, 219–226. [Google Scholar] [CrossRef]
- Smith, F.A.; Jakobsen, I.; Smith, S.E. Spatial differences in acquisition of soil phosphate between two arbuscular mycorrhizal fungi in symbiosis with Medicago truncatula. New Phytol. 2000, 147, 357–366. [Google Scholar] [CrossRef]
- Jansa, J.; Smith, F.A.; Smith, S.E. Are there benefits of simultaneous root colonization by different arbuscular mycorrhizal fungi? New Phytol. 2008, 177, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Dighton, J. Acquisition of nutrients from organic resources by mycorrhizal autotrophic plants. Experientia 1991, 47, 362–369. [Google Scholar] [CrossRef]
- Bunn, R.A.; Simpson, D.T.; Bullington, L.S.; Lekberg, Y.; Janos, D.P. Revisiting the ‘direct mineral cycling’ hypothesis: Arbuscular mycorrhizal fungi colonize leaf litter, but why? ISME J. 2019, 13, 1891–1898. [Google Scholar] [CrossRef]
- Azcón-Aguilar, C.; Barea, J.M. Nutrient cycling in the mycorrhizosphere. J. Soil Sci. Plant Nutr. 2015, 15, 372–396. [Google Scholar] [CrossRef]
- Koide, R.T.; Kabir, Z. Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate. New Phytol. 2000, 148, 511–517. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, X.; Yun, W.; Xia, Y.; You, Z.; Rillig, M.C. Arbuscular mycorrhiza contributes to the control of phosphorus loss in paddy fields. Plant Soil 2020, 447, 623–636. [Google Scholar] [CrossRef]
- Bender, S.F.; Conen, F.; Van der Heijden, M.G.A. Mycorrhizal effects on nutrient cycling, nutrient leaching and N2O production in experimental grassland. Soil Biol. Biochem. 2015, 80, 283–292. [Google Scholar] [CrossRef]
- Heijden, M.G.A. van der Mycorrhizal fungi reduce nutrient loss from model grassland ecosystems. Ecology 2010, 91, 1163–1171. [Google Scholar] [CrossRef]
- Martinez-Garcia, L.B.; de Deyn, G.B.; Pugnaire, F.I.; Kothamasi, D.; van der Heijden, M.G.A. Symbiotic soil fungi enhance ecosystem resilience to climate change. Glob. Chang. Biol. 2017, 23, 5228–5236. [Google Scholar] [CrossRef]
- Easton, Z.M.; Faulkner, J.W. Communicating Climate Change to Agricultural Audiences; Virginia Cooperative Extension, Virginia Tech.: Blacksburg, VA, USA, 2016. [Google Scholar]
- Melillo, J.M.; Richmond, T.; Yohe, G.W. Climate Change Impacts in the United States: The Third National Climate Assessment; U.S. Global Change Research Program: Washington, DC, USA, 2014.
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi—Potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Wallander, H. Uptake of P from apatite by Pinus sylvestris seedlings colonised by different ectomycorrhizal fungi. Plant Soil 2000, 218, 249–256. [Google Scholar] [CrossRef]
- Tawaraya, K.; Hirose, R.; Wagatsuma, T. Inoculation of arbuscular mycorrhizal fungi can substantially reduce phosphate fertilizer application to Allium fistulosum L. and achieve marketable yield under field condition. Biol. Fertil. Soils 2012, 48, 839–843. [Google Scholar] [CrossRef]
- Broadmeadow, S.; Nisbet, T.R. The effects of riparian forest management on the freshwater environment: A literature review of best management practice. Hydrol. Earth Syst. Sci. Discuss. 2004, 8, 286–305. [Google Scholar] [CrossRef]
- Heckrath, G.; Brookes, P.C.; Poulton, P.R.; Goulding, K.W.T. Phosphorus leaching from soils containing different phosphorus concentrations in the Broadbalk Experiment. J. Environ. Qual. 1995, 24, 904–910. [Google Scholar] [CrossRef]
- Holste, E.K.; Kobe, R.K.; Gehring, C.A. Plant species differ in early seedling growth and tissue nutrient responses to arbuscular and ectomycorrhizal fungi. Mycorrhiza 2017, 27, 211–223. [Google Scholar] [CrossRef]
- Khalil, S.; Loynachan, T.E. Soil drainage and distribution of VAM fungi in two toposequences. Soil Biol. Biochem. 1994, 26, 929–934. [Google Scholar] [CrossRef]
- Ellis, J.R. Post Flood Syndrome and Vesicular-Arbuscular Mycorrhizal Fungi. J. Prod. Agric. 1998, 11, 200–204. [Google Scholar] [CrossRef]
- Stevens, K.J.; Wellner, M.R.; Acevedo, M.F. Dark septate endophyte and arbuscular mycorrhizal status of vegetation colonizing a bottomland hardwood forest after a 100 year flood. Aquat. Bot. 2010, 92, 105–111. [Google Scholar] [CrossRef]
- Shenker, M.; Seitelbach, S.; Brand, S.; Haim, A.; Litaor, M.I. Redox reactions and phosphorus release in re-flooded soils of an altered wetland. Eur. J. Soil Sci. 2005, 56, 515–525. [Google Scholar] [CrossRef]
- Rubæk, G.H.; Kristensen, K.; Olesen, S.E.; Østergaard, H.S.; Heckrath, G. Phosphorus accumulation and spatial distribution in agricultural soils in Denmark. Geoderma 2013, 209–210, 241–250. [Google Scholar] [CrossRef]
- Fornara, D.A.; Flynn, D.; Caruso, T. Improving phosphorus sustainability in intensively managed grasslands: The potential role of arbuscular mycorrhizal fungi. Sci. Total Environ. 2020, 706, 135744. [Google Scholar] [CrossRef] [PubMed]
- Ngosong, C.; Jarosch, M.; Raupp, J.; Neumann, E.; Ruess, L. The impact of farming practice on soil microorganisms and arbuscular mycorrhizal fungi: Crop type versus long-term mineral and organic fertilization. Appl. Soil Ecol. 2010, 46, 134–142. [Google Scholar] [CrossRef]
- Sheng, M.; Lalande, R.; Hamel, C.; Ziadi, N. Effect of long-term tillage and mineral phosphorus fertilization on arbuscular mycorrhizal fungi in a humid continental zone of Eastern Canada. Plant Soil 2013, 369, 599–613. [Google Scholar] [CrossRef]
- Schneider, K.D.; Voroney, R.P.; Lynch, D.H.; Oberson, A.; Frossard, E.; Bünemann, E.K. Microbially-mediated P fluxes in calcareous soils as a function of water-extractable phosphate. Soil Biol. Biochem. 2017, 106, 51–60. [Google Scholar] [CrossRef]
- Thirkell, T.J.; Charters, M.D.; Elliott, A.J.; Sait, S.M.; Field, K.J. Are mycorrhizal fungi our sustainable saviours? Considerations for achieving food security. J. Ecol. 2017, 105, 921–929. [Google Scholar] [CrossRef]
- Oka, N.; Karasawa, T.; Okazaki, K.; Takebe, M. Maintenance of soybean yield with reduced phosphorus application by previous cropping with mycorrhizal plants. Soil Sci. Plant Nutr. 2010, 56, 824–830. [Google Scholar] [CrossRef]
- Grant, C.; Bittman, S.; Montreal, M.; Plenchette, C.; Morel, C. Soil and fertilizer phosphorus: Effects on plant P supply and mycorrhizal development. Can. J. Plant Sci. 2005, 85, 3–14. [Google Scholar] [CrossRef]
- Kabir, Z. Tillage or no-tillage: Impact on mycorrhizae. Can. J. Plant Sci. 2005, 85, 23–29. [Google Scholar] [CrossRef]
- Köhl, L.; Van Der Heijden, M.G. Arbuscular mycorrhizal fungal species differ in their effect on nutrient leaching. Soil Biol. Biochem. 2016, 94, 191–199. [Google Scholar] [CrossRef]
- Djodjic, F. Phosphorus Leaching in Relation to Soil Type and Soil Phosphorus Content. J. Environ. Qual. 2004, 33, 7. [Google Scholar] [CrossRef] [PubMed]
- Landry, C.P.; Hamel, C.; Vanasse, A. Influence of arbuscular mycorrhizae on soil P dynamics, corn P-nutrition and growth in a ridge-tilled commercial field. Can. J. Soil Sci. 2008, 88, 283–294. [Google Scholar] [CrossRef]
- Hoffmann, C.C.; Kjaergaard, C.; Uusi-Kämppä, J.; Hansen, H.C.B.; Kronvang, B. Phosphorus Retention in Riparian Buffers: Review of Their Efficiency. J. Environ. Qual. 2009, 38, 1942–1955. [Google Scholar] [CrossRef] [PubMed]
- Turunen, J.; Markkula, J.; Rajakallio, M.; Aroviita, J. Riparian forests mitigate harmful ecological effects of agricultural diffuse pollution in medium-sized streams. Sci. Total Environ. 2019, 649, 495–503. [Google Scholar] [CrossRef]
- Knopf, F.L.; Johnson, R.R.; Rich, T.; Samson, F.B.; Szaro, R.C. Conservation of Riparian Ecosystems in the United States. Wilson Bull. 1988, 100, 272–284. [Google Scholar]
- Tanaka, M.O.; de Souza, A.L.T.; Moschini, L.E.; Oliveira, A.K. de Influence of watershed land use and riparian characteristics on biological indicators of stream water quality in southeastern Brazil. Agric. Ecosyst. Environ. 2016, 216, 333–339. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; Rodríguez Osuna, V.; Cak, A.D.; Bhaduri, A.; Bunn, S.E.; Corsi, F.; Gastelumendi, J.; Green, P.; Harrison, I.; Lawford, R.; et al. Ecosystem-based water security and the Sustainable Development Goals (SDGs). Ecohydrol. Hydrobiol. 2018, 18, 317–333. [Google Scholar] [CrossRef]
- Kelly, J.M.; Kovar, J.L.; Sokolowsky, R.; Moorman, T.B. Phosphorus uptake during four years by different vegetative cover types in a riparian buffer. Nutr. Cycl. Agroecosystems 2007, 78, 239–251. [Google Scholar] [CrossRef]
- Kiedrzyńska, E.; Wagner, I.; Zalewski, M. Quantification of phosphorus retention efficiency by floodplain vegetation and a management strategy for a eutrophic reservoir restoration. Ecol. Eng. 2008, 33, 15–25. [Google Scholar] [CrossRef]
- Volk, T.A.; Abrahamson, L.P.; Nowak, C.A.; Smart, L.B.; Tharakan, P.J.; White, E.H. The development of short-rotation willow in the northeastern United States for bioenergy and bioproducts, agroforestry and phytoremediation. Biomass Bioenergy 2006, 30, 715–727. [Google Scholar] [CrossRef]
- Lu, S.Y.; Wu, F.C.; Lu, Y.F.; Xiang, C.S.; Zhang, P.Y.; Jin, C.X. Phosphorus removal from agricultural runoff by constructed wetland. Ecol. Eng. 2009, 35, 402–409. [Google Scholar] [CrossRef]
- Maestre, A.; Pitt, R.E.; Williamson, D. University of Alabama Nonparametric Statistical Tests Comparing First Flush and Composite Samples from the National Stormwater Quality Database. J. Water Manag. Model. 2004. [Google Scholar] [CrossRef]
- Oberndorfer, E.; Lundholm, J.; Bass, B.; Coffman, R.R.; Doshi, H.; Dunnett, N.; Gaffin, S.; Köhler, M.; Liu, K.K.Y.; Rowe, B. Green Roofs as Urban Ecosystems: Ecological Structures, Functions, and Services. BioScience 2007, 57, 823–833. [Google Scholar] [CrossRef]
- John, J.; Kernaghan, G.; Lundholm, J. The potential for mycorrhizae to improve green roof function. Urban Ecosyst. 2017, 20, 113–127. [Google Scholar] [CrossRef]
- Kye-Han, L.; Isenhart, T.M.; Schultz, R.C.; Mickelson, S.K. Multispecies riparian buffers trap sediment and nutrients during rainfall simulations. J. Environ. Qual. Madison 2000, 29, 1200. [Google Scholar]
- Koerselman, W.; Bakker, S.A.; Blom, M. Nitrogen, Phosphorus and Potassium Budgets for Two Small Fens Surrounded by Heavily Fertilized Pastures. J. Ecol. 1990, 78, 428–442. [Google Scholar] [CrossRef]
- Fillion, M.; Brisson, J.; Guidi, W.; Labrecque, M. Increasing phosphorus removal in willow and poplar vegetation filters using arbuscular mycorrhizal fungi. Ecol. Eng. 2011, 37, 199–205. [Google Scholar] [CrossRef]
- Kieta, K.A.; Owens, P.N.; Lobb, D.A.; Vanrobaeys, J.A.; Flaten, D.N. Phosphorus dynamics in vegetated buffer strips in cold climates: A review. Environ. Rev. 2018, 26, 255–272. [Google Scholar] [CrossRef]
- Mejía, A.; Miguel, N.H.; Enrique, R.S.; Miguel, D. The United Nations World Water Development Report—N° 4—Water and Sustainability (A Review of Targets, Tools and Regional Cases); UNESCO: Paris, France, 2012. [Google Scholar]
- Sato, T.; Qadir, M.; Yamamoto, S.; Endo, T.; Zahoor, A. Global, regional, and country level need for data on wastewater generation, treatment, and use. Agric. Water Manag. 2013, 130, 1–13. [Google Scholar] [CrossRef]
- Maltais-Landry, G.; Frossard, E. Similar phosphorus transfer from cover crop residues and water-soluble mineral fertilizer to soils and a subsequent crop. Plant Soil 2015, 393, 193–205. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Stormwater Technology Fact Sheet: Bioretention; USEPA 832 F 99 102: Washington, DC, USA, 1999.
- Hurley, S.; Shrestha, P.; Cording, A. Nutrient Leaching from Compost: Implications for Bioretention and Other Green Stormwater Infrastructure. J. Sustain. Water Built Environ. 2017, 3, 04017006. [Google Scholar] [CrossRef]
- Poor, C.; Balmes, C.; Freudenthaler, M.; Martinez, A. Role of Mycelium in Bioretention Systems: Evaluation of Nutrient and Metal Retention in Mycorrhizae-Inoculated Mesocosms. J. Environ. Eng. 2018, 144, 04018034. [Google Scholar] [CrossRef]
- Polomski, R.F.; Taylor, M.D.; Bielenberg, D.G.; Bridges, W.C.; Klaine, S.J.; Whitwell, T. Nitrogen and Phosphorus Remediation by Three Floating Aquatic Macrophytes in Greenhouse-Based Laboratory-Scale Subsurface Constructed Wetlands. Water. Air. Soil Pollut. 2009, 197, 223–232. [Google Scholar] [CrossRef]
- Hinsinger, P.; Brauman, A.; Devau, N.; Gérard, F.; Jourdan, C.; Laclau, J.; Le Cadre, E.; Jaillard, B.; Plassard, C. Acquisition of phosphorus and other poorly mobile nutrients by roots. Where do plant nutrition models fail? Plant Soil Dordr. 2011, 348, 29–61. [Google Scholar] [CrossRef]
- Bao, X.; Wang, Y.; Olsson, P.A. Arbuscular mycorrhiza under water—Carbon‒phosphorus exchange between rice and arbuscular mycorrhizal fungi under different flooding regimes. Soil Biol. Biochem. 2019, 129, 169–177. [Google Scholar] [CrossRef]
- Xu, Z.; Ban, Y.; Jiang, Y.; Zhang, X.; Liu, X. Arbuscular Mycorrhizal Fungi in Wetland Habitats and Their Application in Constructed Wetland: A Review. Pedosphere 2016, 26, 592–617. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J.; Klironomos, J.N. Plant coexistence mediated by arbuscular mycorrhizal fungi. Trends Ecol. Evol. 2003, 18, 418–423. [Google Scholar] [CrossRef]
- Doolette, A.; Armstrong, R.; Tang, C.; Guppy, C.; Mason, S.; McNeill, A. Phosphorus uptake benefit for wheat following legume break crops in semi-arid Australian farming systems. Nutr. Cycl. Agroecosystems 2019, 113, 247–266. [Google Scholar] [CrossRef]
- Pavinato, P.S.; Rodrigues, M.; Soltangheisi, A.; Sartor, L.R.; Withers, P.J.A. Effects of Cover Crops and Phosphorus Sources on Maize Yield, Phosphorus Uptake, and Phosphorus Use Efficiency. Agron. J. 2017, 109, 1039–1047. [Google Scholar] [CrossRef]
- Arcand, M.M.; Lynch, D.H.; Voroney, R.P.; van Straaten, P. Residues from a buckwheat (Fagopyrum esculentum) green manure crop grown with phosphate rock influence bioavailability of soil phosphorus. Can. J. Soil Sci. 2010, 90, 257–266. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Giles, C.; Darch, T.; George, T.S.; Blackwell, M.; Stutter, M.; Shand, C.; Lumsdon, D.; Cooper, P.; Wendler, R.; et al. Opportunities for mobilizing recalcitrant phosphorus from agricultural soils: A review. Plant Soil 2018, 427, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Withers, P.J.A.; Sylvester-Bradley, R.; Jones, D.L.; Healey, J.R.; Talboys, P.J. Feed the Crop Not the Soil: Rethinking Phosphorus Management in the Food Chain. Environ. Sci. Technol. 2014, 48, 6523–6530. [Google Scholar] [CrossRef] [PubMed]
- Ruckli, R.; Rusterholz, H.-P.; Baur, B. Invasion of an annual exotic plant into deciduous forests suppresses arbuscular mycorrhiza symbiosis and reduces performance of sycamore maple saplings. For. Ecol. Manag. 2014, 318, 285–293. [Google Scholar] [CrossRef]
- Zubek, S.; Majewska, M.L.; Błaszkowski, J.; Stefanowicz, A.M.; Nobis, M.; Kapusta, P. Invasive plants affect arbuscular mycorrhizal fungi abundance and species richness as well as the performance of native plants grown in invaded soils. Biol. Fertil. Soils 2016, 52, 879–893. [Google Scholar] [CrossRef]
- Lekberg, Y.; Gibbons, S.M.; Rosendahl, S.; Ramsey, P.W. Severe plant invasions can increase mycorrhizal fungal abundance and diversity. ISME J. 2013, 7, 1424–1433. [Google Scholar] [CrossRef]
- Bunn, R.A.; Ramsey, P.W.; Lekberg, Y. Do native and invasive plants differ in their interactions with arbuscular mycorrhizal fungi? A meta-analysis. J. Ecol. 2015, 103, 1547–1556. [Google Scholar] [CrossRef]
- Orion, T. Beyond the War on Invasive Species; Chelsea Green Publishing: White River Junction, VT, USA, 2015. [Google Scholar]
- Meisner, A.; Gera Hol, W.H.; de Boer, W.; Krumins, J.A.; Wardle, D.A.; van der Putten, W.H. Plant–soil feedbacks of exotic plant species across life forms: A meta-analysis. Biol. Invasions 2014, 16, 2551–2561. [Google Scholar] [CrossRef]
- Nikolić, L.; Džigurski, D.; Ljevnaić-Mašić, B. Nutrient removal by Phragmites australis (Cav.) Trin. ex Steud. In the constructed wetland system. Contemp. Probl. Ecol. 2014, 7, 449–454. [Google Scholar] [CrossRef]
- Moore, M.T.; Locke, M.A.; Kröger, R. Using aquatic vegetation to remediate nitrate, ammonium, and soluble reactive phosphorus in simulated runoff. Chemosphere 2016, 160, 149–154. [Google Scholar] [CrossRef]
- El Amrani, A.; Dumas, A.-S.; Wick, L.Y.; Yergeau, E.; Berthomé, R. “Omics” Insights into PAH Degradation toward Improved Green Remediation Biotechnologies. Environ. Sci. Technol. 2015, 49, 11281–11291. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study Context | Study Conditions | Phosphorus Quantity Measured | % Change with Mycorrhiza # | Location | Ref. # |
|---|---|---|---|---|---|
| Crop uptake | Agro ecosystem Triticum aestivum, AMF | Phosphorus use efficiency | +85–102% | Uttar Pradesh, Haryana, India | [22] |
| Growth of native grasses | Field ecosystem and pots in greenhouse, Stipa pulchra Avena barbata, fungicide/no fungicide *** | Shoot P concentration [mg/g] | San Diego CA, USA | [49] | |
| Field | |||||
| S. pulchra, | +22% | ||||
| A. barbata | +68% | ||||
| Greenhouse | |||||
| Shoot P concentration | |||||
| S. pulchra | +1.6% | ||||
| A. barbata | −11.8% | ||||
| Root concentration | |||||
| S. pulchra | +24% | ||||
| A. barbata | −15% | ||||
| Mulch Experiment | Pots, greenhouse Trifolium repens Zea Mays Fungicide/no fungicide *** | Plant P concentrations (%) | Morioka, Japan | [51] | |
| No Mulch | +28% | ||||
| Living Mulch | +135% | ||||
| Plant P (mg P/plant) | |||||
| No mulch | +17% | ||||
| Living mulch | +709% | ||||
| Crop uptake | Pots, AMF, Allium fistolosum | Plant P concentration [mg/g] | +194% | Haguromachi, Japan | [82] |
| Plant uptake [mg P/pot] | +1525% | ||||
| Effect of mycorrhizosphere bacteria on plant uptake | Pots, corn (Zea Mays), AMF | P plant uptake [mg P/pot] | Denmark | [83] | |
| Shoots | +168% | ||||
| Roots | +234% | ||||
| Effect of sewage sludge P on plant uptake | Pot, greenhouse Glycine max AMF | Shoot biomass P [mg/shoot] | Ohio, USA | [99] | |
| No P addition | +144% | ||||
| 150 mg P/kg addition | +125% | ||||
| 270 mg P/kg addition | −0.8% | ||||
| 420 mg P/kg addition | −16.9% | ||||
| Effect of AMF on P leaching | Packed columns, greenhouse, Trifolium subterraneum AMF | Leachate P [mg] | South Australia | [100] | |
| without added P | −60% | ||||
| with added P. | 0% | ||||
| Plant P [mg] | |||||
| without added P | +251% | ||||
| with added P | −23% | ||||
| Effect of mycorrhizae on crop uptake and extractable soil P | Pot, greenhouse, corn (Zea Mays), AMF | Plant uptake (mg P/plant) | Quebec Canada | [101] | |
| Hybrid | |||||
| P3979 | +8.4% | ||||
| LRS | +19.1% | ||||
| LNS | +19.8% | ||||
| Mehlich 3 extractable Soil P Concentration [mg/kg] | |||||
| Hybrids, no P fertilizer | |||||
| P3979 | −5.1% | ||||
| LRS | −14.4% | ||||
| LNS | −10.5% | ||||
| Mehlich 3 extractable Soil P Concentration [mg/kg], | |||||
| Hybrids, P fertilizer applied | ns | ||||
| Leaching mitigation | Pots, greenhouses, Phalaris aquatic, AMF | Shoot P content (mg) | +150% | Southeastern Australia | [112] |
| Root P content (mg) | +168% | ||||
| Spatial differences in P uptake between AMF species | Pots, Medicago trunculata, AMF | Plant P concentrations | Roskilde, Denmark | [113] | |
| Glomus caledonium | |||||
| Shoot | |||||
| 35 days | +39% | ||||
| 49 days | −17% | ||||
| Roots | |||||
| 35 days | +61% | ||||
| 49 days | +10% | ||||
| Scutetllospora calosporia | |||||
| Shoot | |||||
| 35 days | +39% | ||||
| 49 days | −12% | ||||
| Roots | |||||
| 35 days | +84% | ||||
| 49 days | +40% | ||||
| Differential effect of AMF species | Pots, Medicago tranculata, AMF ## | P uptake [mg/plant] | Mallala, South Australia | [114] | |
| Glomus mossae | |||||
| 4 weeks | +1425% | ||||
| 8 weeks | +314% | ||||
| Glomus claroideum | |||||
| 4 weeks | +625% | ||||
| 8 weeks | +193% | ||||
| Glomus intraradices | |||||
| 4 weeks | +925% | ||||
| 8 weeks | +357% | ||||
| P losses from field | Microcosms Orya sativa L AMF | Leachate [kg P/ha] ### | Jiangsu, China | [119] | |
| Particulate P | −11.1% | ||||
| Dissolved Organic P | −14.4% | ||||
| SRP (PO4) * | −81% | ||||
| Runoff [kg P/ha] | |||||
| Particulate P | −11.1% | ||||
| Dissolved Organic P | −4.95% | ||||
| SRP (PO4) * | −11% | ||||
| Nutrient cycling in presence of mycorrhizae | Microcosms, Heath and Pasture communities, AMF | P in leachate [mg] ### | Switzerland | [120] | |
| Pasture | |||||
| Added NH4 | −14.2% | ||||
| Added NO3 | −38.5% | ||||
| Heath | |||||
| Added NH4 | −68.4% | ||||
| Added NO3 | −63.4% | ||||
| Leaching from grasslands | Mesocosms, grassland, AMF | Reduction in leaching | [121] | ||
| Low nutrient availability | ~ 60% | ||||
| High nutrient availability | ns | ||||
| Climate Change Resilience | Mesocosms, grassland communities, AMF | Leachate P [ug] ### | The Netherlands | [122] | |
| Moderate rain | −149% | ||||
| High rain | −58% | ||||
| Crop Uptake | Pots, Allium fistulosum (Welsh Onion) AMF | Shoot concentration | +88% | Tozawa, Japan | [127] |
| Crop uptake | Agroecosystem Zea Mays AMF | Plant P [mg/plant] ** | Quebec, Canada | [128] | |
| Year 1 Sample days | |||||
| 22 | +26.5% | ||||
| 48 | +46.5% | ||||
| 72 | +18.7 | ||||
| Year 2 Sample days | |||||
| 22 | +19.4% | ||||
| 48 | +14.2% | ||||
| 72 | +41.8% | ||||
| Nutrient Leaching | Laboratory mesocosms. Lolium multiflorum, Trifolium pratense, sterilized soils AMF | Leachate Loss SRP [mg] | Zürich, Switzerland | [129] | |
| Lolium multiflora | |||||
| Claroideoglomus claroideum | +14.2% | ||||
| Funnelformis mosseae | −19.5% | ||||
| Rhizoglomus irregular | +45.0% | ||||
| Trifolium pretense | |||||
| Claroideoglomus claroideum | ns | ||||
| Funnelformis mosseae | ns | ||||
| Rhizoglomus irregular | ns | ||||
| Unreactive P | |||||
| Lolium multiflora | |||||
| Claroideoglomus claroideum | −10.8% | ||||
| Funnelformis mosseae | +3.9% | ||||
| Rhizoglomus irregular | ns | ||||
| Trifolium pratense | |||||
| Claroideoglomus claroideum | +29.9% | ||||
| Funnelformis mosseae | +19.1% | ||||
| Rhizoglomus irregular | +62.4% | ||||
| Vegetative buffers | Pot, Salix, Populus AMF | P stem content | +33% | Southern Quebec, Canada | [162] |
| Bioretention | Field mesocosms, Carex stipata, AMF/ECM commercial mix | Leachate mass rate (mg/hour)### | −34% | Portland, Oregon, USA | [169] |
| Crop uptake | Microcosms, Orya sativa L. AMF | Plant P concentrations {mg/g]### | Sweden | [171] | |
| First growth stage | |||||
| Leaf | ns | ||||
| Stem | +66% | ||||
| Continuous flooding | |||||
| No flooding | −19% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubin, J.A.; Görres, J.H. Potential for Mycorrhizae-Assisted Phytoremediation of Phosphorus for Improved Water Quality. Int. J. Environ. Res. Public Health 2021, 18, 7. https://doi.org/10.3390/ijerph18010007
Rubin JA, Görres JH. Potential for Mycorrhizae-Assisted Phytoremediation of Phosphorus for Improved Water Quality. International Journal of Environmental Research and Public Health. 2021; 18(1):7. https://doi.org/10.3390/ijerph18010007
Chicago/Turabian StyleRubin, Jessica A., and Josef H. Görres. 2021. "Potential for Mycorrhizae-Assisted Phytoremediation of Phosphorus for Improved Water Quality" International Journal of Environmental Research and Public Health 18, no. 1: 7. https://doi.org/10.3390/ijerph18010007
APA StyleRubin, J. A., & Görres, J. H. (2021). Potential for Mycorrhizae-Assisted Phytoremediation of Phosphorus for Improved Water Quality. International Journal of Environmental Research and Public Health, 18(1), 7. https://doi.org/10.3390/ijerph18010007