Oxytocin Receptor Gene Polymorphisms and Early Parental Bonding Interact in Shaping Instagram Social Behavior

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Material and Methods
2.1. Participants
2.2. Procedure
2.3. Parental Bonding
2.4. Genetic Assessment
2.5. Instagram Variables
2.5.1. Number of Followings
2.5.2. Number of Posts
2.5.3. Number of Followers
2.5.4. Social Desirability Index
2.6. Statistical Analysis
3. Results
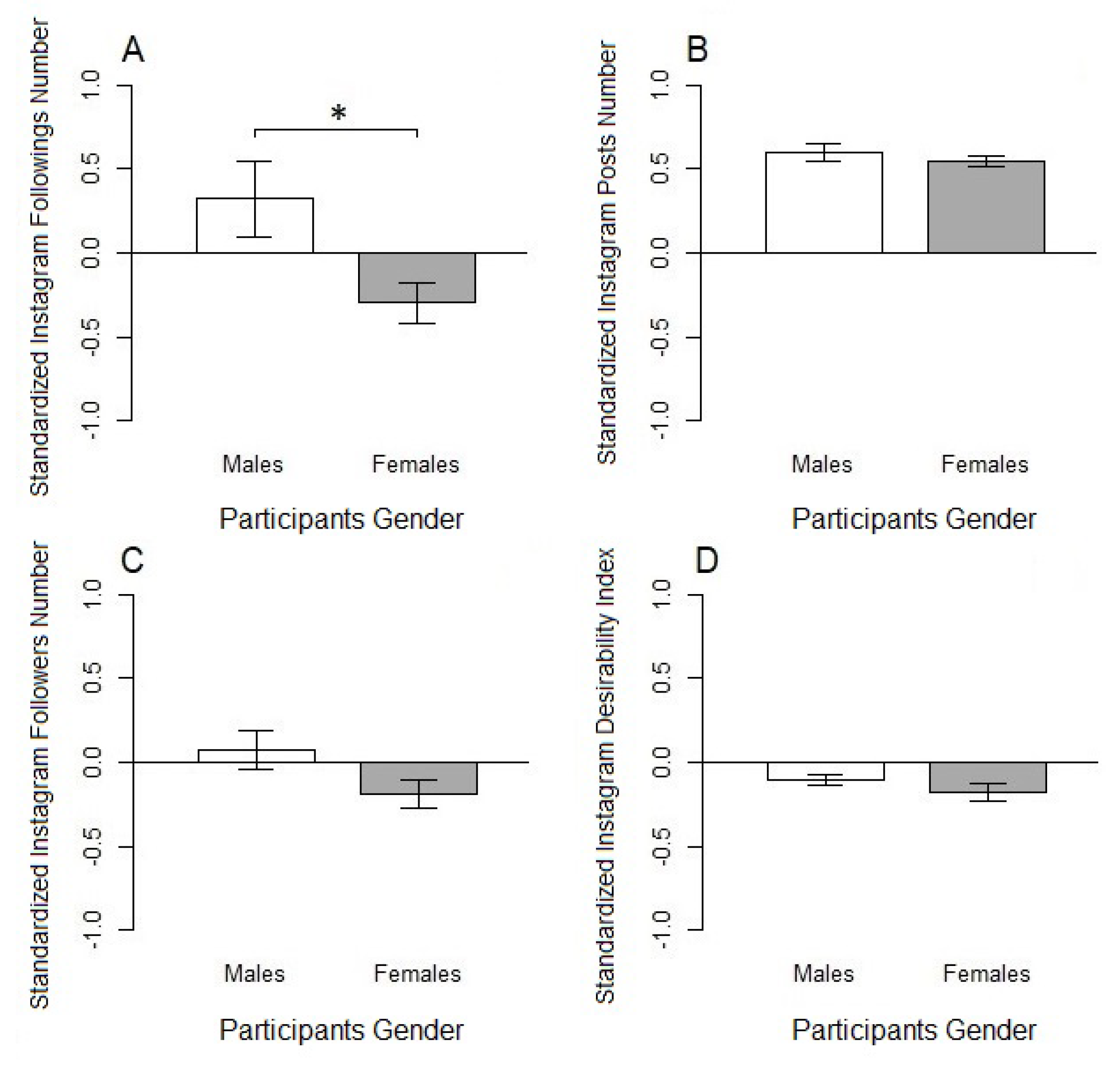
3.1. Instagram Variables: Preliminary Results
3.2. Instagram Effects: OXTr rs2254298
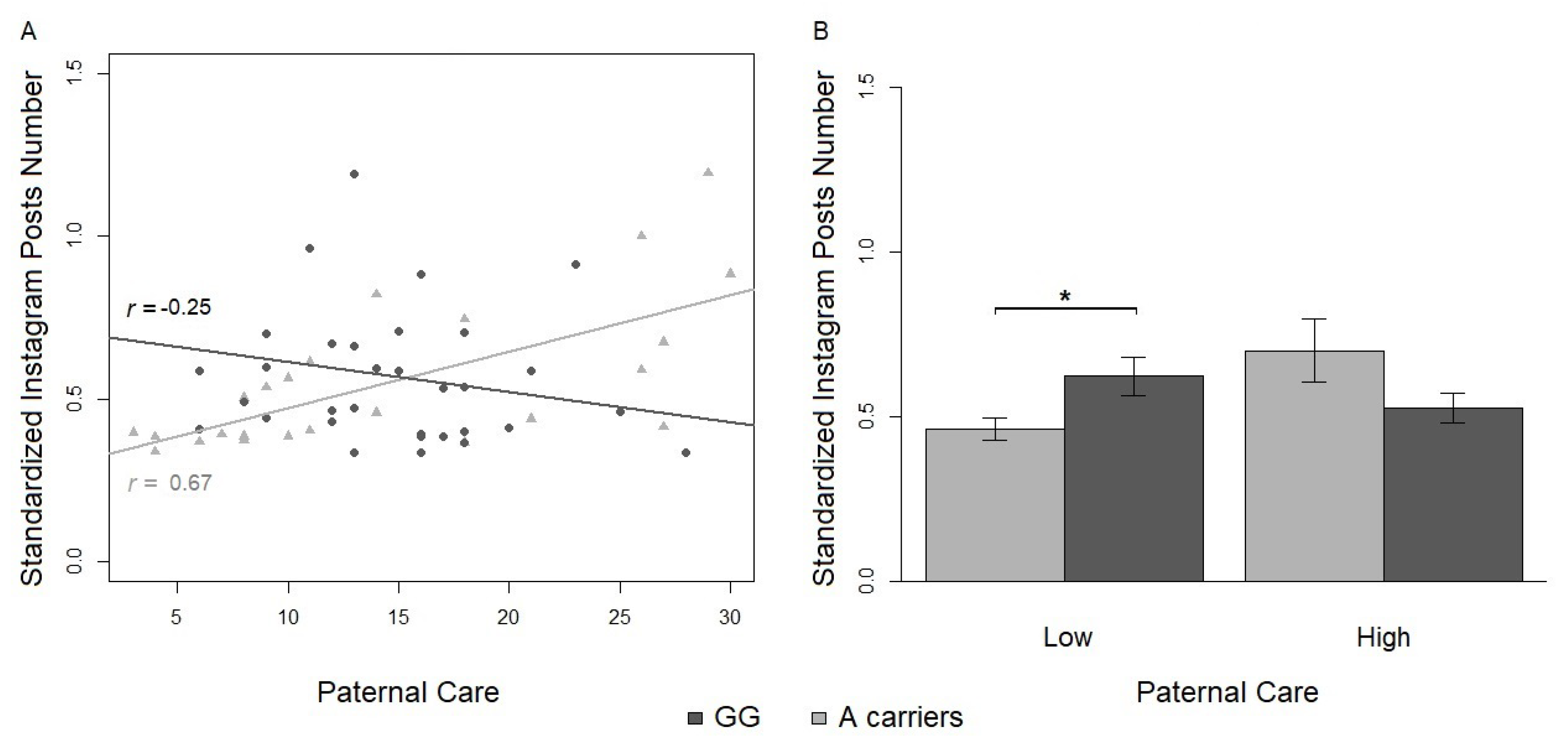
3.2.1. Number of Instagram Posts
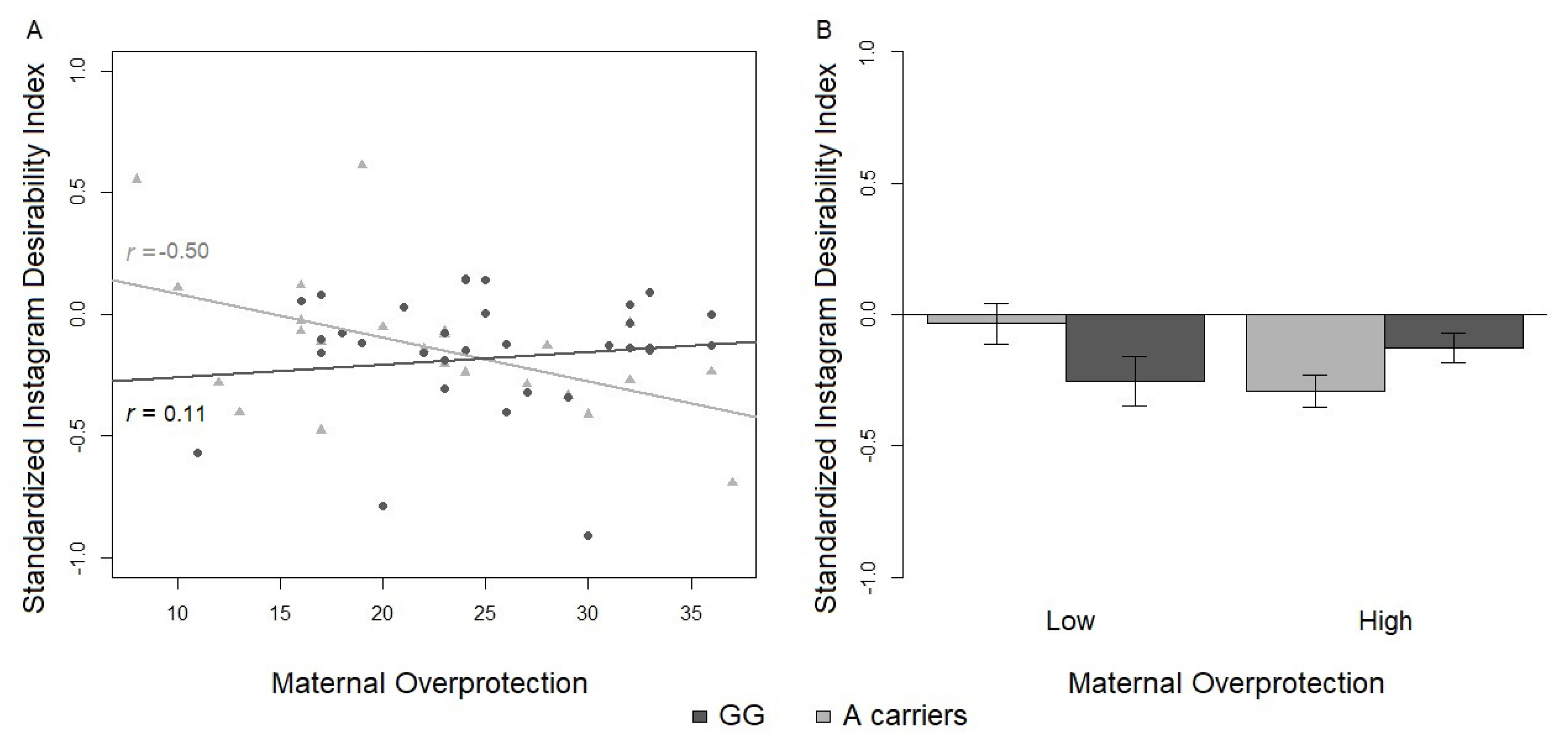
3.2.2. Social Desirability Index
3.2.3. Number of Instagram Followings and Followers
3.3. Instagram Effects: OXTr rs53576
4. Discussion
5. Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SNSs | Social Network Sites |
| OXTr | Oxytocin receptor gene |
| PBI | Parental Bonding Instrument |
| SNP | Single-Nucleotide Polymorphism |
| SDI | Social Desirability Index |
Appendix A. Pre-Registration
Appendix B. Data Repository
Appendix C. Ethics
References
- Jaeggi, A.V.; Burkart, J.M.; Van Schaik, C.P. On the psychology of cooperation in humans and other primates: Combining the natural history and experimental evidence of prosociality. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2723–2735. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, L.; Rousset, F. How life history and demography promote or inhibit the evolution of helping behaviors. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2599–2617. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, S.F.; Bshary, R. Cooperation and deception: From evolution to mechanisms. Philos. Trans. R. Soc. B Biol. Sci. 2010. [Google Scholar] [CrossRef] [PubMed]
- Seifer, R.; Schiller, M. The role of parenting sensitivity, infant temperament, and dyadic interaction in attachment theory and assessment. Monogr. Soc. Res. Child Dev. 1995, 60, 146–174. [Google Scholar] [CrossRef]
- Fagot, B.I. Attachment, parenting, and peer interactions of toddler children. Dev. Psychol. 1997, 33, 489. [Google Scholar] [CrossRef]
- Feeney, J.A. Attachment, caregiving, and marital satisfaction. Pers. Relationsh. 1996, 3, 401–416. [Google Scholar] [CrossRef]
- Griffin, D.W.; Bartholomew, K. Models of the self and other: Fundamental dimensions underlying measures of adult attachment. J. Personal. Soc. Psychol. 1994, 67, 430. [Google Scholar] [CrossRef]
- Esposito, G.; Yoshida, S.; Ohnishi, R.; Tsuneoka, Y.; del Carmen Rostagno, M.; Yokota, S.; Okabe, S.; Kamiya, K.; Hoshino, M.; Shimizu, M.; et al. Infant calming responses during maternal carrying in humans and mice. Curr. Biol. 2013, 23, 739–745. [Google Scholar] [CrossRef]
- Meins, E. Security of Attachment and the Social Development of Cognition; Psychology Press: London, UK, 2013. [Google Scholar]
- Markiewicz, D.; Doyle, A.B.; Brendgen, M. The quality of adolescents’ friendships: Associations with mothers’ interpersonal relationships, attachments to parents and friends, and prosocial behaviors. J. Adolesc. 2001, 24, 429–445. [Google Scholar] [CrossRef]
- Diamond, L.M.; Hicks, A.M. Psychobiological Perspectives on Attachment: Implications for Health over the Lifespan. In Adult Attachment: Theory, Research, and Clinical Implications; Guilford Press: New York, NY, USA, 2004. [Google Scholar]
- Schoenmaker, C.; Huffmeijer, R.; van IJzendoorn, M.H.; Bakermans-Kranenburg, M.J.; van den Dries, L.; Linting, M.; van der Voort, A.; Juffer, F. Attachment and physiological reactivity to infant crying in young adulthood: Dissociation between experiential and physiological arousal in insecure adoptees. Physiol. Behav. 2015, 139, 549–556. [Google Scholar] [CrossRef]
- Robles, T.F.; Kane, H.S. The attachment system and physiology in adulthood: Normative processes, individual differences, and implications for health. J. Personal. 2014, 82, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.; Merrick, S.; Treboux, D.; Crowell, J.; Albersheim, L. Attachment security in infancy and early adulthood: A twenty-year longitudinal study. Child Dev. 2000, 71, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Hutchby, I.; Moran-Ellis, J. Children, Technology and Culture: The Impacts of Technologies in Children’s Everyday Lives; Routledge: Abingdon, UK, 2013. [Google Scholar]
- Espinoza, G.; Juvonen, J. The pervasiveness, connectedness, and intrusiveness of social network site use among young adolescents. Cyberpsychol. Behav. Soc. Netw. 2011, 14, 705–709. [Google Scholar] [CrossRef]
- Vagos, P. Cognitive, affective and behavioral correlates of social anxiety in adolescence. Int. J. Dev. Educ. Psychol. 2009, 1, 293–301. [Google Scholar]
- Blakemore, S.J. The social brain in adolescence. Nat. Rev. Neurosci. 2008, 9, 267. [Google Scholar] [CrossRef] [PubMed]
- Rook, K.S. The negative side of social interaction: Impact on psychological well-being. J. Personal. Soc. Psychol. 1984, 46, 1097. [Google Scholar] [CrossRef]
- Lenhart, A.; Purcell, K.; Smith, A.; Zickuhr, K. Social Media & Mobile Internet Use among Teens and Young Adults. Millennials; Pew Internet & American Life Project: Washington, DC, USA, 2010. [Google Scholar]
- Lin, L.Y.; Sidani, J.E.; Shensa, A.; Radovic, A.; Miller, E.; Colditz, J.B.; Hoffman, B.L.; Giles, L.M.; Primack, B.A. Association between social media use and depression among US young adults. Depress. Anxiety 2016, 33, 323–331. [Google Scholar] [CrossRef]
- Smock, A.D.; Ellison, N.B.; Lampe, C.; Wohn, D.Y. Facebook as a toolkit: A uses and gratification approach to unbundling feature use. Comput. Hum. Behav. 2011, 27, 2322–2329. [Google Scholar] [CrossRef]
- Chen, G.M. Tweet this: A uses and gratifications perspective on how active Twitter use gratifies a need to connect with others. Comput. Hum. Behav. 2011, 27, 755–762. [Google Scholar] [CrossRef]
- Hu, Y.; Manikonda, L.; Kambhampati, S. What we instagram: A first analysis of instagram photo content and user types. In Proceedings of the Eighth International AAAI Conference on Weblogs and Social Media, Ann Arbor, MI, USA, 1–4 June 2014; pp. 595–598. [Google Scholar]
- Archambault, A.; Grudin, J. A longitudinal study of facebook, linkedin, & twitter use. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Austin, TX, USA, 5–10 May 2012; pp. 2741–2750. [Google Scholar]
- Van Dijck, J. ‘You have one identity’: Performing the self on Facebook and LinkedIn. Media Cult. Soc. 2013, 35, 199–215. [Google Scholar] [CrossRef]
- Greenwood, S.; Perrin, A.; Duggan, M. Social media update 2016. Pew Res. Cent. 2016, 11, 1–18. [Google Scholar]
- Ferrara, E.; Interdonato, R.; Tagarelli, A. Online popularity and topical interests through the lens of instagram. In Proceedings of the 25th ACM Conference on Hypertext and Social Media, Santiago, Chile, 1–4 September 2014; pp. 24–34. [Google Scholar]
- Choi, G.Y.; Lewallen, J. “Say Instagram, Kids!”: Examining Sharenting and Children’s Digital Representations on Instagram. Howard J. Commun. 2018, 29, 144–164. [Google Scholar] [CrossRef]
- Antheunis, M.L.; Schouten, A.P. The effects of other-generated and system-generated cues on adolescents’ perceived attractiveness on social network sites. J. Comput. Mediat. Commun. 2011, 16, 391–406. [Google Scholar] [CrossRef]
- Valkenburg, P.M.; Peter, J.; Schouten, A.P. Friend networking sites and their relationship to adolescents’ well-being and social self-esteem. CyberPsychol. Behav. 2006, 9, 584–590. [Google Scholar] [CrossRef]
- Rosenquist, J.N.; Fowler, J.H.; Christakis, N.A. Social network determinants of depression. Mol. Psychiatry 2011, 16, 273. [Google Scholar] [CrossRef]
- Pittman, M.; Reich, B. Social media and loneliness: Why an Instagram picture may be worth more than a thousand Twitter words. Comput. Hum. Behav. 2016, 62, 155–167. [Google Scholar] [CrossRef]
- Ridgway, J.L.; Clayton, R.B. Instagram unfiltered: Exploring associations of body image satisfaction, Instagram# selfie posting, and negative romantic relationship outcomes. Cyberpsychol. Behav. Soc. Netw. 2016, 19, 2–7. [Google Scholar]
- Sheldon, P.; Bryant, K. Instagram: Motives for its use and relationship to narcissism and contextual age. Comput. Hum. Behav. 2016, 58, 89–97. [Google Scholar] [CrossRef]
- Lee, E.; Lee, J.A.; Moon, J.H.; Sung, Y. Pictures speak louder than words: Motivations for using Instagram. Cyberpsychol. Behav. Soc. Netw. 2015, 18, 552–556. [Google Scholar] [CrossRef]
- Le Moignan, E.; Lawson, S.; Rowland, D.A.; Mahoney, J.; Briggs, P. Has Instagram Fundamentally Altered the ‘Family Snapshot’? In Proceedings of the 2017 CHI Conference on Human Factors in Computing Systems, Denver, CO, USA, 6–11 May 2017; pp. 4935–4947. [Google Scholar]
- Kross, E.; Verduyn, P.; Demiralp, E.; Park, J.; Lee, D.S.; Lin, N.; Shablack, H.; Jonides, J.; Ybarra, O. Facebook use predicts declines in subjective well-being in young adults. PLoS ONE 2013, 8, e69841. [Google Scholar] [CrossRef]
- Junco, R. Too much face and not enough books: The relationship between multiple indices of Facebook use and academic performance. Comput. Hum. Behav. 2012, 28, 187–198. [Google Scholar] [CrossRef]
- Junco, R. The relationship between frequency of Facebook use, participation in Facebook activities, and student engagement. Comput. Educ. 2012, 58, 162–171. [Google Scholar] [CrossRef]
- Thimm, C.; Nehls, P. Sharing grief and mourning on Instagram: Digital patterns of family memories. Communications 2017, 42, 327–349. [Google Scholar] [CrossRef]
- York, C. A regression approach to testing genetic influence on communication behavior: Social media use as an example. Comput. Hum. Behav. 2017, 73, 100–109. [Google Scholar] [CrossRef]
- Bonassi, A.; Cataldo, I.; Gabrieli, G.; Tandiono, M.; Foo, J.N.; Lepri, B.; Esposito, G. The interaction between serotonin transporter allelic variation and maternal care modulates sociability on Instagram. PsyArXiv 2020. [Google Scholar] [CrossRef]
- Belsky, J.; Jonassaint, C.; Pluess, M.; Stanton, M.; Brummett, B.; Williams, R. Vulnerability genes or plasticity genes? Mol. Psychiatry 2009, 14, 746. [Google Scholar] [CrossRef]
- Belsky, J.; Pluess, M. The nature (and nurture?) of plasticity in early human development. Perspect. Psychol. Sci. 2009, 4, 345–351. [Google Scholar] [CrossRef]
- Boyce, W.T.; Ellis, B.J. Biological sensitivity to context: I. An evolutionary–developmental theory of the origins and functions of stress reactivity. Dev. Psychopathol. 2005, 17, 271–301. [Google Scholar] [CrossRef]
- Belsky, J.; Newman, D.A.; Widaman, K.F.; Rodkin, P.; Pluess, M.; Fraley, R.C.; Berry, D.; Helm, J.L.; Roisman, G.I. Differential susceptibility to effects of maternal sensitivity? A study of candidate plasticity genes. Dev. Psychopathol. 2015, 27, 725–746. [Google Scholar] [CrossRef]
- Esposito, G.; Setoh, P.; Shinohara, K.; Bornstein, M.H. The development of attachment: Integrating genes, brain, behavior, and environment. Behav. Brain Res. 2017, 325, 87–89. [Google Scholar] [CrossRef][Green Version]
- Ellis, B.J.; Boyce, W.T.; Belsky, J.; Bakermans-Kranenburg, M.J.; Van Ijzendoorn, M.H. Differential susceptibility to the environment: An evolutionary–neurodevelopmental theory. Dev. Psychopathol. 2011, 23, 7–28. [Google Scholar] [CrossRef] [PubMed]
- Truzzi, A.; Poquérusse, J.; Setoh, P.; Shinohara, K.; Bornstein, M.H.; Esposito, G. Oxytocin receptor gene polymorphisms (rs53576) and early paternal care sensitize males to distressing female vocalizations. Dev. Psychobiol. 2018, 60, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Pluess, M.; Belsky, J. Vantage sensitivity: Individual differences in response to positive experiences. Psychol. Bull. 2013, 139, 901. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, I.; Bonassi, A.; Lepri, B.; Foo, J.N.; Setoh, P.; Esposito, G. Recalled parental bonding interacts with oxytocin receptor gene polymorphism in modulating anxiety and avoidance in adult relationships. bioRxiv 2020. [Google Scholar] [CrossRef]
- Belsky, J.; Steinberg, L.; Draper, P. Childhood experience, interpersonal development, and reproductive strategy: An evolutionary theory of socialization. Child Dev. 1991, 62, 647–670. [Google Scholar] [CrossRef]
- Belsky, J. Etiology of child maltreatment: A developmental ecological analysis. Psychol. Bull. 1993, 114, 413. [Google Scholar] [CrossRef]
- Pluess, M.; Belsky, J. Differential susceptibility to parenting and quality child care. Dev. Psychol. 2010, 46, 379. [Google Scholar] [CrossRef]
- Cataldo, I.; Azhari, A.; Esposito, G. A review of oxytocin and arginine-vasopressin receptors and their modulation of autism spectrum disorder. Front. Mol. Neurosci. 2018, 11, 27. [Google Scholar] [CrossRef]
- Bartz, J.A.; Zaki, J.; Bolger, N.; Ochsner, K.N. Social effects of oxytocin in humans: Context and person matter. Trends Cogn. Sci. 2011, 15, 301–309. [Google Scholar] [CrossRef]
- Feldman, R.; Monakhov, M.; Pratt, M.; Ebstein, R.P. Oxytocin pathway genes: Evolutionary ancient system impacting on human affiliation, sociality, and psychopathology. Biol. Psychiatry 2016, 79, 174–184. [Google Scholar] [CrossRef]
- Ebstein, R.P.; Knafo, A.; Mankuta, D.; Chew, S.H.; San Lai, P. The contributions of oxytocin and vasopressin pathway genes to human behavior. Horm. Behav. 2012, 61, 359–379. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Li, Z.; Su, Y. The association between oxytocin receptor gene polymorphism (OXTR) and trait empathy. J. Affect. Disord. 2012, 138, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, I.; Azhari, A.; Lepri, B.; Esposito, G. Oxytocin receptors (OXTR) and early parental care: An interaction that modulates psychiatric disorders. Res. Dev. Disabil. 2018, 82, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Truzzi, A.; Setoh, P.; Putnick, D.L.; Shinohara, K.; Bornstein, M.H. Genetic predispositions and parental bonding interact to shape adults’ physiological responses to social distress. Behav. Brain Res. 2017, 325, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Krueger, F.; Parasuraman, R.; Iyengar, V.; Thornburg, M.; Weel, J.; Lin, M.; Clarke, E.; McCabe, K.; Lipsky, R. Oxytocin receptor genetic variation promotes human trust behavior. Front. Hum. Neurosci. 2012, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.E.; Porges, E.C.; Norman, G.J.; Connelly, J.J.; Decety, J. Oxytocin receptor gene variation predicts empathic concern and autonomic arousal while perceiving harm to others. Soc. Neurosci. 2014, 9, 1–9. [Google Scholar] [CrossRef]
- Rodrigues, S.M.; Saslow, L.R.; Garcia, N.; John, O.P.; Keltner, D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 21437–21441. [Google Scholar] [CrossRef]
- Li, J.; Zhao, Y.; Li, R.; Broster, L.S.; Zhou, C.; Yang, S. Association of oxytocin receptor gene (OXTR) rs53576 polymorphism with sociality: A meta-analysis. PLoS ONE 2015, 10, e0131820. [Google Scholar] [CrossRef]
- Chen, F.S.; Kumsta, R.; von Dawans, B.; Monakhov, M.; Ebstein, R.P.; Heinrichs, M. Common oxytocin receptor gene (OXTR) polymorphism and social support interact to reduce stress in humans. Proc. Natl. Acad. Sci. USA 2011, 108, 19937–19942. [Google Scholar] [CrossRef]
- Parker, G.; Tupling, H.; Brown, L.B. A parental bonding instrument. Br. J. Med Psychol. 1979, 52, 1–10. [Google Scholar] [CrossRef]
- Dalsant, A.; Truzzi, A.; Setoh, P.; Esposito, G. Maternal bonding in childhood moderates autonomic responses to distress stimuli in adult males. Behav. Brain Res. 2015, 292, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.S.; Garrity-Rokous, F.E.; Chazan-Cohen, R.; Little, C.; Briggs-Gowan, M.J. Maternal depression and comorbidity: Predicting early parenting, attachment security, and toddler social-emotional problems and competencies. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Bonassi, A.; Ghilardi, T.; Truzzi, A.; Cataldo, I.; Azhari, A.; Setoh, P.; Shinohara, K.; Esposito, G. Dataset on genetic and physiological adults’ responses to social distress. Data Brief 2017, 13, 742–748. [Google Scholar] [CrossRef]
- Sherlock, M.; Wagstaff, D.L. Exploring the relationship between frequency of Instagram use, exposure to idealized images, and psychological well-being in women. Psychol. Pop. Media Cult. 2018, 8, 482. [Google Scholar] [CrossRef]
- Araújo, C.S.; Corrêa, L.P.D.; da Silva, A.P.C.; Prates, R.O.; Meira, W. It is not just a picture: Revealing some user practices in instagram. In Proceedings of the 2014 9th Latin American Web Congress, Ouro Preto, Brazil, 22–24 October 2014; pp. 19–23. [Google Scholar]
- Jang, J.Y.; Han, K.; Shih, P.C.; Lee, D. Generation like: Comparative characteristics in Instagram. In Proceedings of the 33rd Annual ACM Conference on Human Factors in Computing Systems, Seoul, Korea, 18–23 April 2015; pp. 4039–4042. [Google Scholar]
- Frison, E.; Eggermont, S. Browsing, posting, and liking on Instagram: The reciprocal relationships between different types of Instagram use and adolescents’ depressed mood. Cyberpsychol. Behav. Soc. Netw. 2017, 20, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Li, P.; Loh, R.S.M.; Chua, T.H.H. A study of Singapore adolescent girls’ selfie practices, peer appearance comparisons, and body esteem on Instagram. Body Image 2019, 29, 90–99. [Google Scholar] [CrossRef]
- Lup, K.; Trub, L.; Rosenthal, L. Instagram# instasad?: Exploring associations among instagram use, depressive symptoms, negative social comparison, and strangers followed. Cyberpsychol. Behav. Soc. Netw. 2015, 18, 247–252. [Google Scholar]
- Chancellor, S.; Lin, Z.J.; De Choudhury, M. This post will just get taken down: Characterizing removed pro-eating disorder social media content. In Proceedings of the 2016 CHI Conference on Human Factors in Computing Systems, San Jose, CA, USA, 7–12 May 2016; pp. 1157–1162. [Google Scholar]
- Holland, G.; Tiggemann, M. “Strong beats skinny every time”: Disordered eating and compulsive exercise in women who post fitspiration on Instagram. Int. J. Eat. Disord. 2017, 50, 76–79. [Google Scholar] [CrossRef]
- Barry, C.T.; McDougall, K.H.; Anderson, A.C.; Perkins, M.D.; Lee-Rowland, L.M.; Bender, I.; Charles, N.E. ‘Check Your Selfie before You Wreck Your Selfie’: Personality ratings of Instagram users as a function of self-image posts. J. Res. Personal. 2019, 82, 103843. [Google Scholar] [CrossRef]
- Sannon, S.; Murnane, E.I.; Bazarova, N.N.; Gay, G. “I was really, really nervous posting it”: Communicating about Invisible Chronic Illnesses across Social Media Platforms. In Proceedings of the 2019 CHI Conference on Human Factors in Computing Systems, Glasgow, UK, 4–9 May 2019; p. 353. [Google Scholar]
- Cheung, T.T. A Study on Motives, Usage, Self-Presentation and Number of Followers on Instagram. CityU Institutional Repository. 2014. Available online: http://dspace.cityu.edu.hk/handle/2031/7521 (accessed on 3 October 2020).
- Agam, D. Followers Ratio on Instagram Affects in Product’s Brand Awareness. Aust. J. Account. Econ. Financ. 2017, 3, 85–89. [Google Scholar]
- Erz, A.; Marder, B.; Osadchaya, E. Hashtags: Motivational drivers, their use, and differences between influencers and followers. Comput. Hum. Behav. 2018, 89, 48–60. [Google Scholar] [CrossRef]
- Ben-Gal, I. Outlier detection. In Data Mining and Knowledge Discovery Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 131–146. [Google Scholar]
- Kwak, S.K.; Kim, J.H. Statistical data preparation: Management of missing values and outliers. Korean J. Anesthesiol. 2017, 70, 407. [Google Scholar] [CrossRef] [PubMed]
- Gravetter, F.; Wallnau, L. Essentials of Statistics for the Behavioral Sciences; Cengage Learning: Belmont, CA, USA, 2005. [Google Scholar]
- Diedenhofen, B.; Musch, J. Cocor: A comprehensive solution for the statistical comparison of correlations. PLoS ONE 2015, 10, e0121945. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G* Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Nielsen, M.; Haun, D.; Kärtner, J.; Legare, C.H. The persistent sampling bias in developmental psychology: A call to action. J. Exp. Child Psychol. 2017, 162, 31–38. [Google Scholar] [CrossRef]
- Belsky, J. Differential susceptibility to environmental influences. Int. J. Child Care Educ. Policy 2013, 7, 15–31. [Google Scholar] [CrossRef]
- Costa, B.; Pini, S.; Gabelloni, P.; Abelli, M.; Lari, L.; Cardini, A.; Muti, M.; Gesi, C.; Landi, S.; Galderisi, S.; et al. Oxytocin receptor polymorphisms and adult attachment style in patients with depression. Psychoneuroendocrinology 2009, 34, 1506–1514. [Google Scholar] [CrossRef]
- McQuaid, R.J.; McInnis, O.A.; Matheson, K.; Anisman, H. Oxytocin and social sensitivity: Gene polymorphisms in relation to depressive symptoms and suicidal ideation. Front. Hum. Neurosci. 2016, 10, 358. [Google Scholar] [CrossRef]
- Belsky, J.; Bakermans-Kranenburg, M.J.; Van IJzendoorn, M.H. For better and for worse: Differential susceptibility to environmental influences. Curr. Dir. Psychol. Sci. 2007, 16, 300–304. [Google Scholar] [CrossRef]
- Cataldo, I.; Neoh, M.J.Y.; Chew, W.F.; Foo, J.N.; Lepri, B.; Esposito, G. Oxytocin receptor gene and parental bonding modulate prefrontal responses to cries: A NIRS Study. Sci. Rep. 2020, 10, 8588. [Google Scholar] [CrossRef]
- Besser, A.; Priel, B. Dependency, self-criticism and negative affective responses following imaginary rejection and failure threats: Meaning-making processes as moderators or mediators. Psychiatry Interpers. Biol. Process. 2011, 74, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Marcus, S. Picturing’ourselves into being: Assessing identity, sociality and visuality on Instagram. In Proceedings of the International Communication Association Conference, San Juan, Puerto Rico, 21–25 May 2015; pp. 21–25. [Google Scholar]
- Mikulincer, M.; Shaver, P.R. Attachment in Adulthood: Structure, Dynamics, and Change; Guilford Press: New York, NY, USA, 2007. [Google Scholar]
- Jackson, C.A.; Luchner, A.F. Self-presentation mediates the relationship between self-criticism and emotional response to Instagram feedback. Personal. Individ. Differ. 2018, 133, 1–6. [Google Scholar] [CrossRef]
- Ershad, Z.S.; Aghajani, T. Prediction of Instagram social network addiction based on the personality, alexithymia and attachment Styles. Sociol. Stud. Youth 2017, 8, 21–34. [Google Scholar]
- Jenkins-Guarnieri, M.A.; Wright, S.L.; Hudiburgh, L.M. The relationships among attachment style, personality traits, interpersonal competency, and Facebook use. J. Appl. Dev. Psychol. 2012, 33, 294–301. [Google Scholar] [CrossRef]
- Laible, D. Attachment with parents and peers in late adolescence: Links with emotional competence and social behavior. Personal. Individ. Differ. 2007, 43, 1185–1197. [Google Scholar] [CrossRef]
- Yates, T.M.; Gregor, M.A.; Haviland, M.G. Child maltreatment, alexithymia, and problematic internet use in young adulthood. Cyberpsychol. Behav. Soc. Netw. 2012, 15, 219–225. [Google Scholar] [CrossRef]
- Lee, S.J.; Chae, Y.G. Children’s Internet use in a family context: Influence on family relationships and parental mediation. Cyberpsychol. Behav. 2007, 10, 640–644. [Google Scholar] [CrossRef]
- Kirwil, L. Parental mediation of children’s internet use in different European countries. J. Child. Media 2009, 3, 394–409. [Google Scholar] [CrossRef]
- van Den Eijnden, R.J.; Spijkerman, R.; Vermulst, A.A.; van Rooij, T.J.; Engels, R.C. Compulsive Internet use among adolescents: Bidirectional parent–child relationships. J. Abnorm. Child Psychol. 2010, 38, 77–89. [Google Scholar] [CrossRef]
- Teo, A.R.; Gaw, A.C. Hikikomori, a Japanese culture-bound syndrome of social withdrawal? A proposal for DSM-V. J. Nerv. Ment. Dis. 2010, 198, 444. [Google Scholar] [CrossRef]
- Paulus, F.W.; Ohmann, S.; Von Gontard, A.; Popow, C. Internet gaming disorder in children and adolescents: A systematic review. Dev. Med. Child Neurol. 2018, 60, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Hawk, S.T.; van den Eijnden, R.J.; van Lissa, C.J.; ter Bogt, T.F. Narcissistic adolescents’ attention-seeking following social rejection: Links with social media disclosure, problematic social media use, and smartphone stress. Comput. Hum. Behav. 2019, 92, 65–75. [Google Scholar] [CrossRef]
- Marino, C.; Gini, G.; Angelini, F.; Vieno, A.; Spada, M.M. Social norms and e-motions in problematic social media use among adolescents. Addict. Behav. Rep. 2020, 11, 100250. [Google Scholar] [CrossRef] [PubMed]
- Yamasue, H. Function and structure in social brain regions can link oxytocin-receptor genes with autistic social behavior. Brain Dev. 2013, 35, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. Oxytocin receptor (OXTR) and serotonin transporter (5-HTT) genes associated with observed parenting. Soc. Cogn. Affect. Neurosci. 2008, 3, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Munafò, M.R. Reliability and replicability of genetic association studies. Addiction 2009, 104, 1439–1440. [Google Scholar] [PubMed]
- Mitchell, K.J. Neurogenomics-towards a more rigorous science. Eur. J. Neurosci. 2018, 47, 109–114. [Google Scholar] [CrossRef]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and vasopressin in the human brain: Social neuropeptides for translational medicine. Nat. Rev. Neurosci. 2011, 12, 524–538. [Google Scholar] [CrossRef]
- Montag, C.; Reuter, M. Disentangling the molecular genetic basis of personality: From monoamines to neuropeptides. Neurosci. Biobehav. Rev. 2014, 43, 228–239. [Google Scholar] [CrossRef]
- Insel, T.R. The challenge of translation in social neuroscience: A review of oxytocin, vasopressin, and affiliative behavior. Neuron 2010, 65, 768–779. [Google Scholar] [CrossRef]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| PBI Subscale | Lower | Raw | Upper |
|---|---|---|---|
| Paternal care | 0.83 | 0.88 | 0.93 |
| Maternal care | 0.88 | 0.91 | 0.95 |
| Paternal Overprotection | 0.79 | 0.85 | 0.91 |
| Maternal Overprotection | 0.84 | 0.89 | 0.93 |
| Variables | Min | 1st Q | Median | Mean | 3rd Q | Max |
|---|---|---|---|---|---|---|
| Followings Number | −1.54 | −0.77 | −0.12 | −0.12 | 0.52 | 1.80 |
| Posts Number | 0.33 | 0.39 | 0.49 | 0.56 | 0.67 | 1.19 |
| Followers Number | −0.80 | −0.50 | −0.16 | −0.11 | 0.22 | 1.45 |
| Social Desirability Index | −1.16 | −0.28 | −0.13 | −0.16 | −0.03 | 0.61 |
| Maternal care | 0.00 | 4.00 | 8.00 | 9.97 | 14.00 | 28.00 |
| Paternal care | 0.00 | 9.00 | 14.00 | 14.49 | 18.00 | 30.00 |
| Maternal overprotection | 8.00 | 18.00 | 23.00 | 23.82 | 30.00 | 37.00 |
| Paternal overprotection | 11.00 | 23.00 | 29.00 | 27.05 | 32.00 | 37.00 |
| Variables | Skewness | Kurtosis |
|---|---|---|
| Followings Number | 0.29 | −0.72 |
| Posts Number | 1.69 | 2.56 |
| Log-transformed Posts Number | 1.22 | 0.82 |
| Followers Number | 0.79 | 0.17 |
| Social Desirability Index | −0.70 | 2.63 |
| Instagram Variables | Males | Females |
|---|---|---|
| Followings Number | 0.32 (0.23) | −0.28 (0.12) |
| Posts Number | 0.60 (0.05) | 0.55 (0.03) |
| Followers Number | 0.08 (0.12) | −0.19 (0.08) |
| Social Desirability Index | −0.11 (0.03) | −0.18 (0.05) |
| Instagram Variables | Low | High | ||
| Posts Number | 0.54 (0.04) | 0.59 (0.05) | ||
| Social Desirability Index | −0.14 (0.06) | −0.18 (0.05) | ||
| Instagram Variables | Low/A | Low/GG | High/A | High/GG |
| Posts Number | 0.46 (0.03) | 0.62 (0.06) | 0.70 (0.10) | 0.52 (0.05) |
| Social Desirability Index | −0.04 (0.08) | −0.25 (0.10) | −0.29 (0.06) | −0.13 (0.06) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonassi, A.; Cataldo, I.; Gabrieli, G.; Foo, J.N.; Lepri, B.; Esposito, G. Oxytocin Receptor Gene Polymorphisms and Early Parental Bonding Interact in Shaping Instagram Social Behavior. Int. J. Environ. Res. Public Health 2020, 17, 7232. https://doi.org/10.3390/ijerph17197232
Bonassi A, Cataldo I, Gabrieli G, Foo JN, Lepri B, Esposito G. Oxytocin Receptor Gene Polymorphisms and Early Parental Bonding Interact in Shaping Instagram Social Behavior. International Journal of Environmental Research and Public Health. 2020; 17(19):7232. https://doi.org/10.3390/ijerph17197232
Chicago/Turabian StyleBonassi, Andrea, Ilaria Cataldo, Giulio Gabrieli, Jia N. Foo, Bruno Lepri, and Gianluca Esposito. 2020. "Oxytocin Receptor Gene Polymorphisms and Early Parental Bonding Interact in Shaping Instagram Social Behavior" International Journal of Environmental Research and Public Health 17, no. 19: 7232. https://doi.org/10.3390/ijerph17197232
APA StyleBonassi, A., Cataldo, I., Gabrieli, G., Foo, J. N., Lepri, B., & Esposito, G. (2020). Oxytocin Receptor Gene Polymorphisms and Early Parental Bonding Interact in Shaping Instagram Social Behavior. International Journal of Environmental Research and Public Health, 17(19), 7232. https://doi.org/10.3390/ijerph17197232