Molecular Verification of Bloom-forming Aphanizomenon flos-aquae and Their Secondary Metabolites in the Nakdong River


Abstract
:1. Introduction
2. Materials and Methods
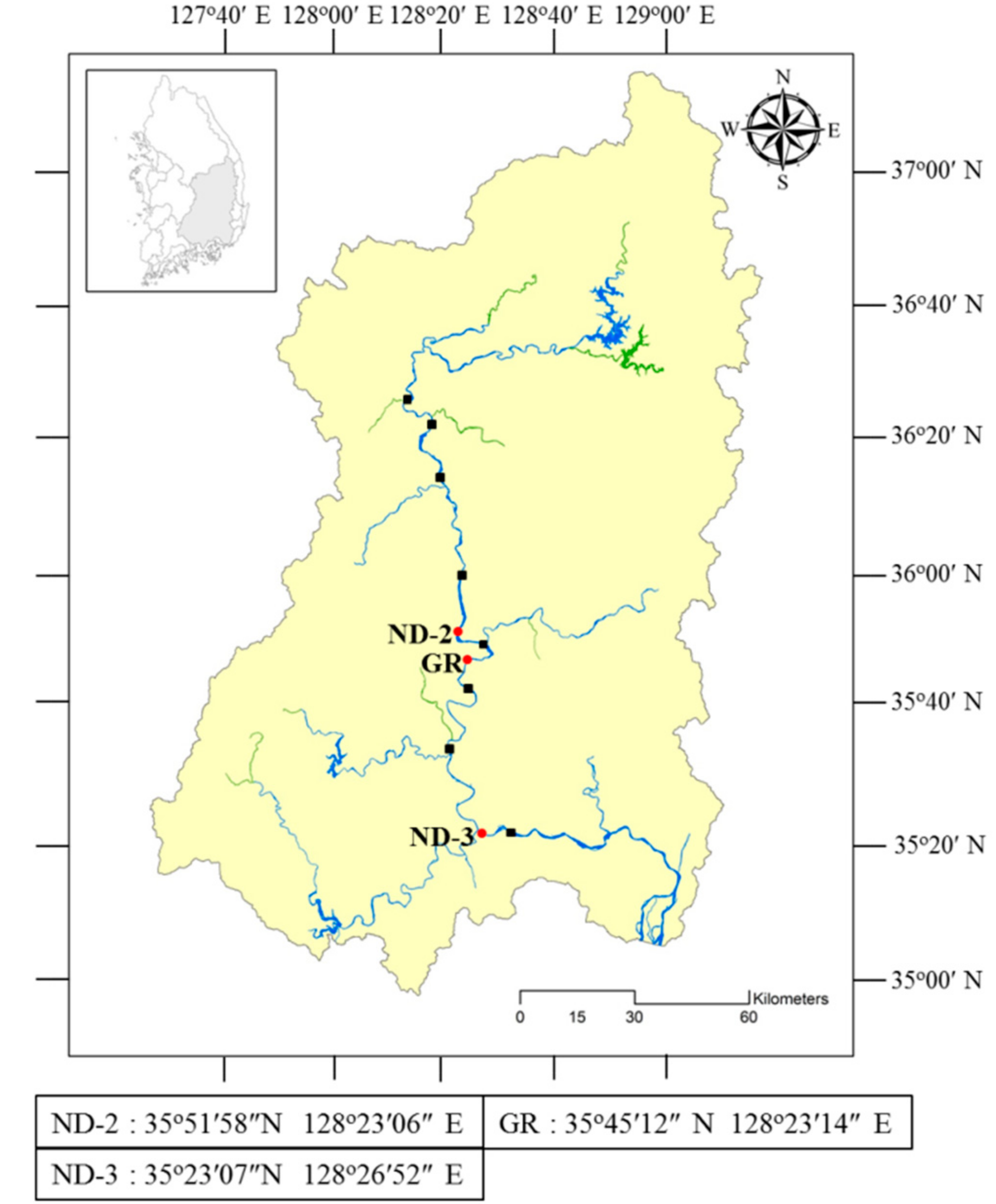
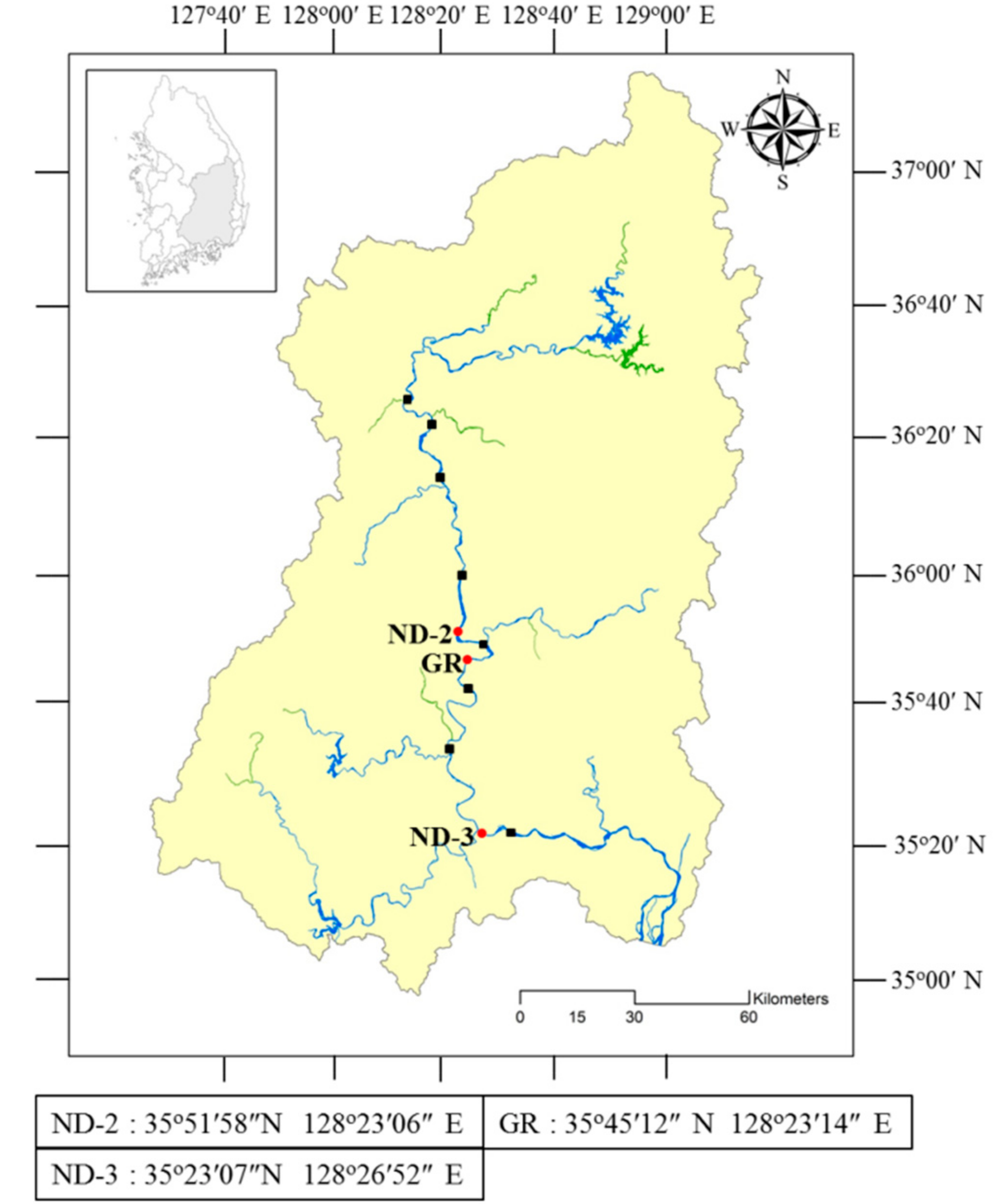
2.1. Sampling and Phytoplankton Analysis
2.2. Isolation and Culture of Aphanizomenon Strains
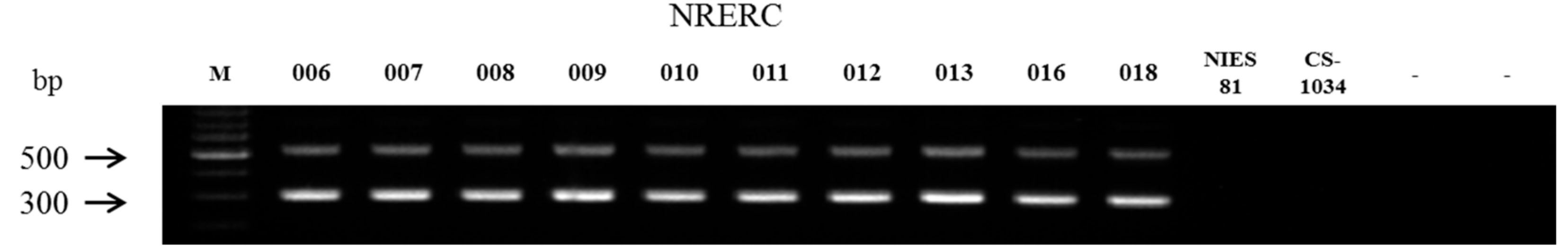
2.3. Isolation of Genomic DNA
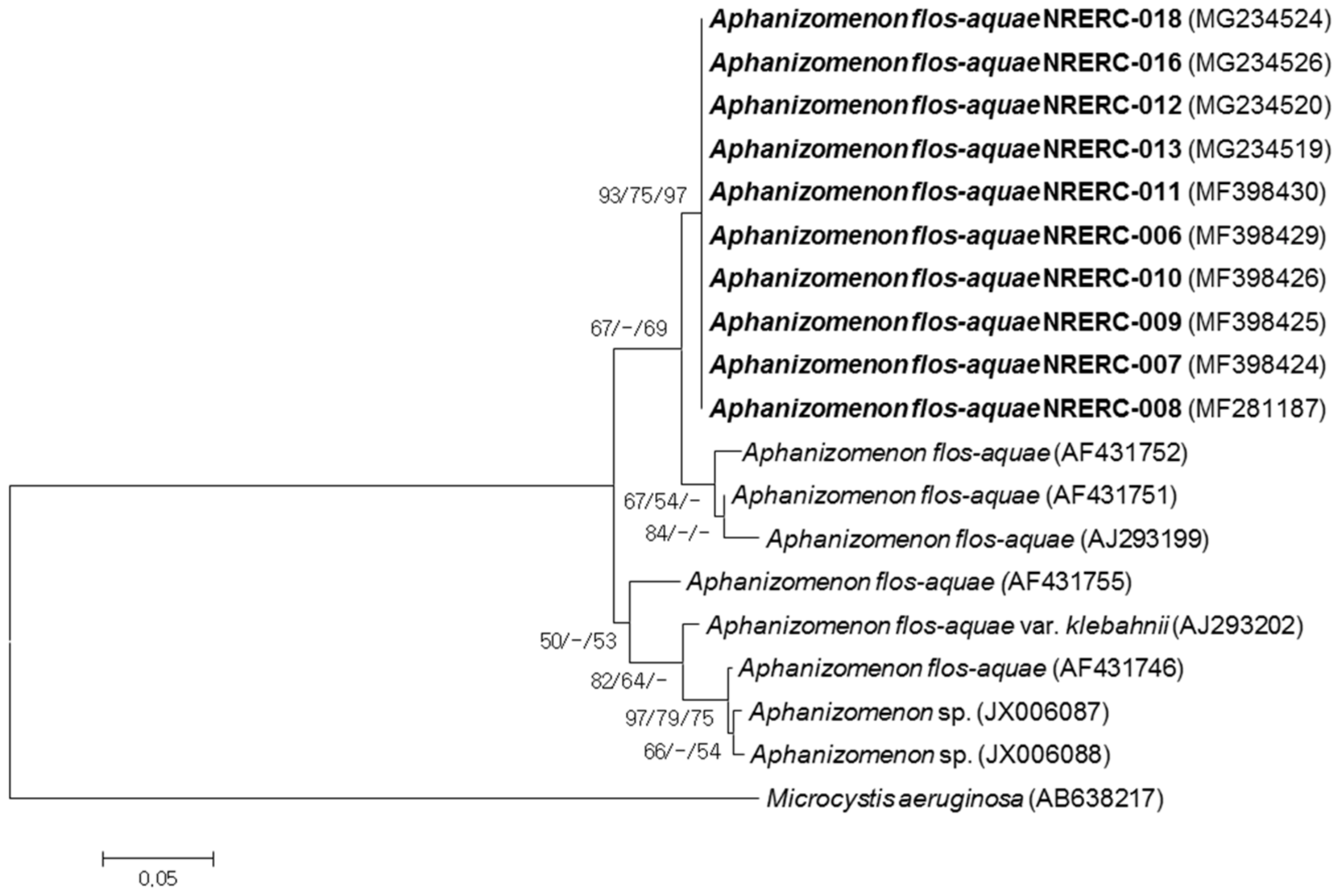
2.4. Phylogenetic Analysis
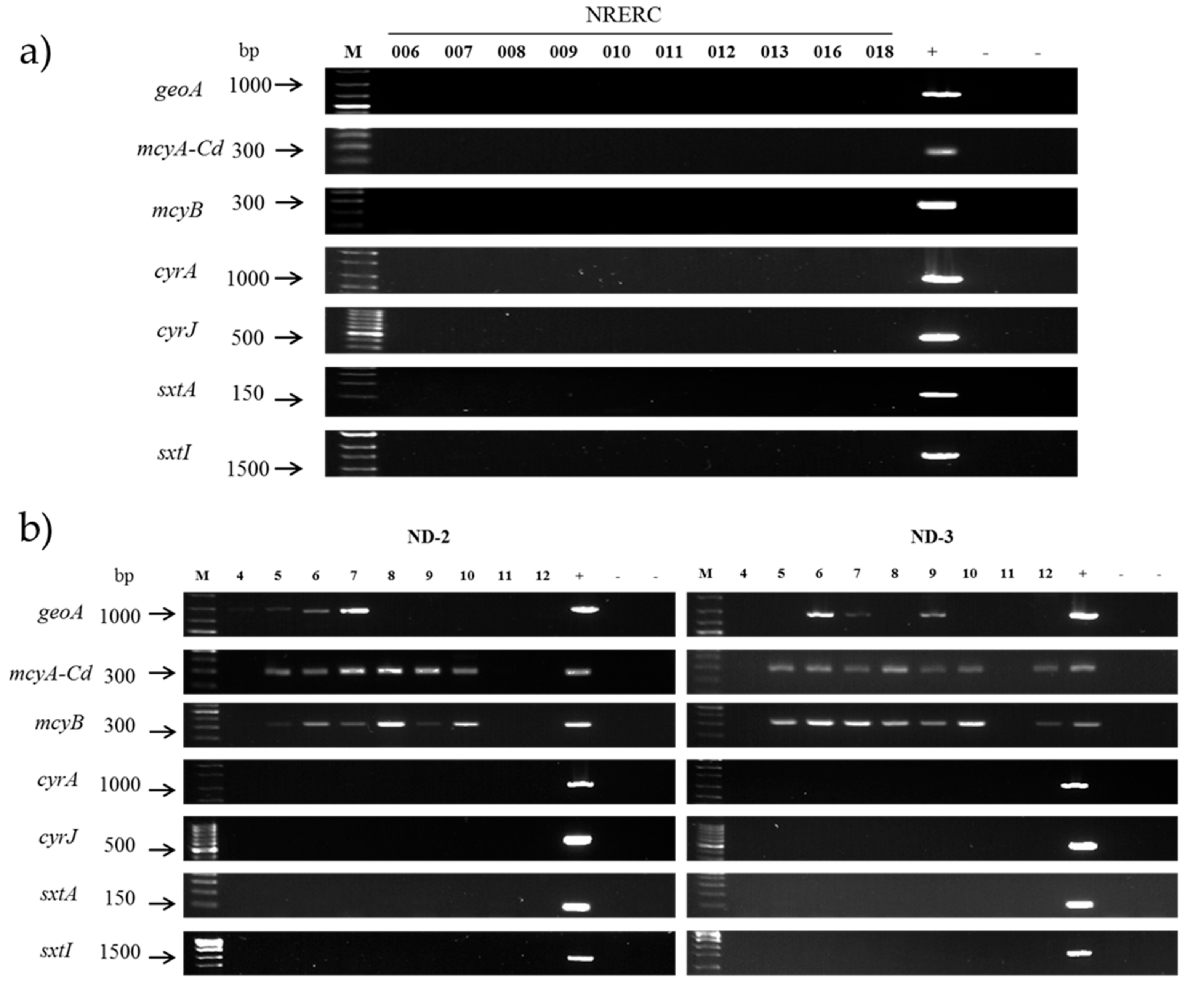
2.5. Molecular Assays for Off-Flavor and Toxin Production
3. Results
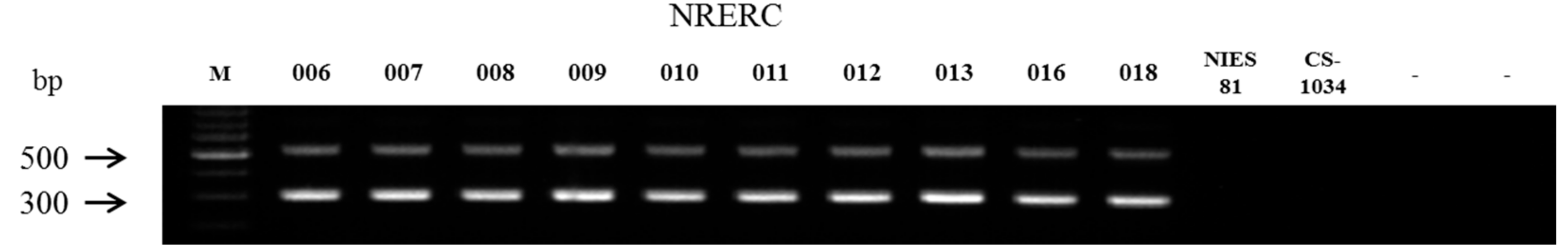
3.1. Identification of Aphanizomenon Isolates by Morphological Characteristics
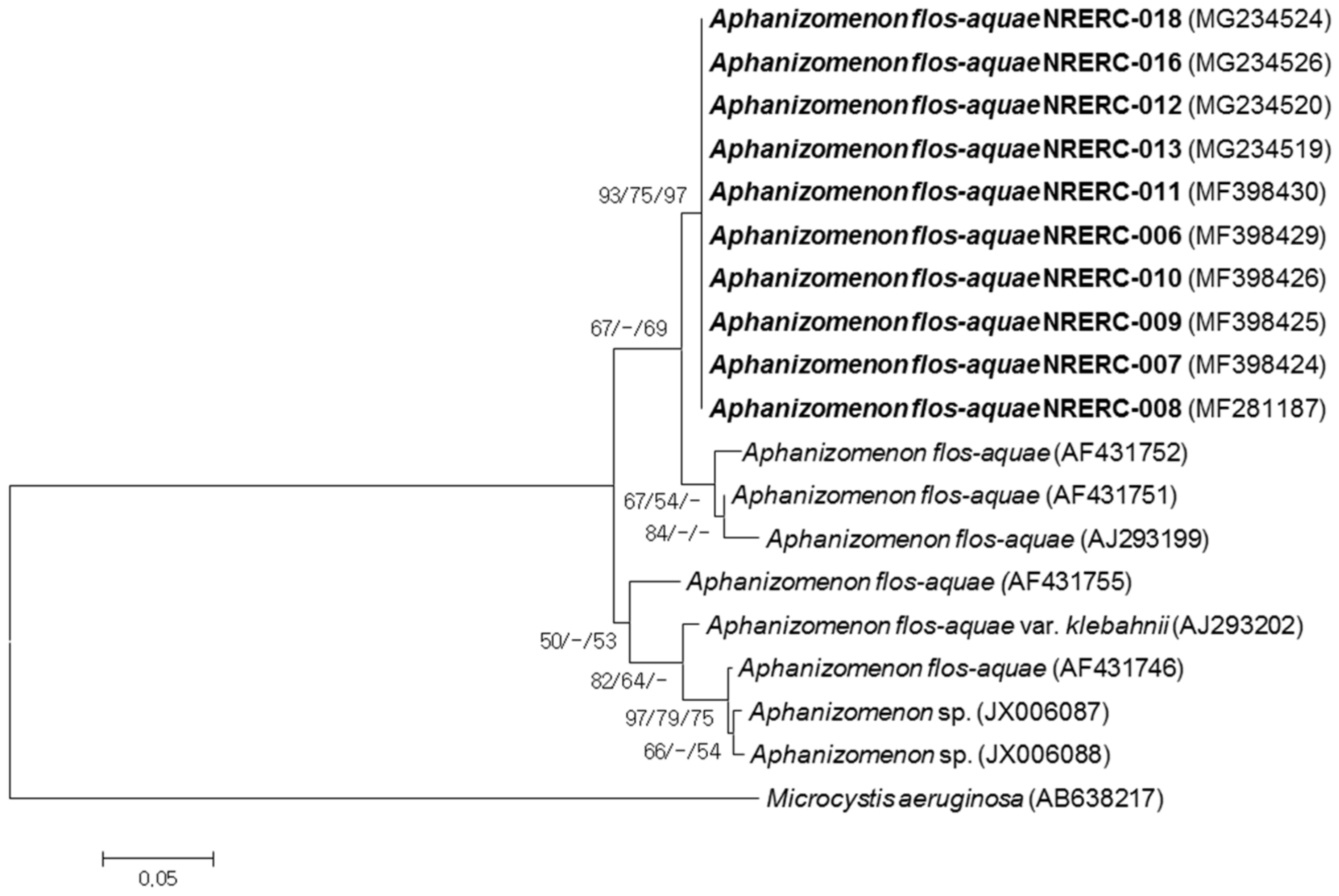
3.2. Phylogenetic Analyses of 16S rRNA and ITS Sequences
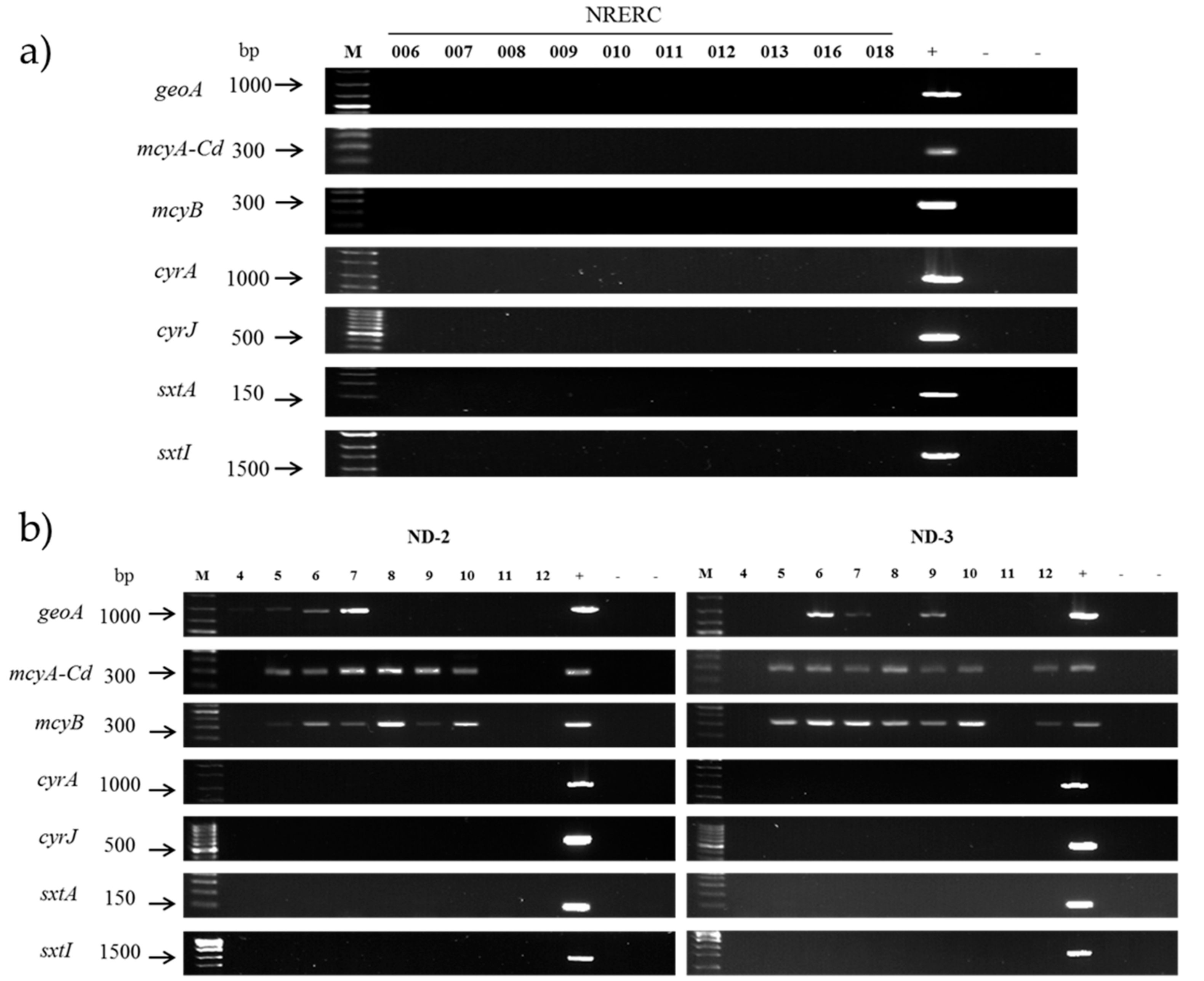
3.3. Evaluation of Secondary Metabolite Production Potential
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Mantzouki, E. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales Cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Goldyn, R. Domination of invasive Nostocales (Cyanoprokaryota) at 52°N latitude. Phycol. Res. 2017, 65, 322–332. [Google Scholar] [CrossRef]
- Nakdong River Environment Research Center (NRERC). Aquatic Ecosystem Monitoring in Weir of the Nakdong River; Nakdong River Environment Research Center: Goryung, Korea, 2015. [Google Scholar]
- Ha, K.; Cho, E.A.; Kim, H.W.; Joo, G.J. Microcystis bloom formation in the lower Nakdong River, South Korea; Importance of hydrodynamics and nutrient loading. Mar. Freshw. Res. 1999, 50, 89–94. [Google Scholar] [CrossRef]
- Jeong, K.-S.; Kin, D.-K.; Whigham, P.; Joo, G.-J. Modelling Microcystis aeruginosa bloom dynamics in the Nakdong River by means of evolutionary computation and statistical approach. Ecol. Model. 2003, 161, 67–78. [Google Scholar] [CrossRef]
- Park, H.-K.; Shin, R.-Y.; Lee, H.-J.; Lee, K.-L.; Cheon, S.-U. Spatio-temporal characteristics of cyanobacterial communities in the middle-downstream of Nakdong River and Lake Dukdong. J. Korean Soc. Water Environ. 2015, 31, 286–294. [Google Scholar] [CrossRef]
- Ryu, H.-S.; Park, H.-K.; Lee, H.-J.; Shin, R.-Y.; Cheon, S.-U. Occurrence and succession pattern of cyanobacteria in the upper region of the Nakdong River; Factors influencing Aphanizomenon bloom. J. Korean Soc. Water Environ. 2016, 32, 52–59. [Google Scholar] [CrossRef]
- Park, H.-K.; Lee, H.-J.; Park, K.-J.; Cho, H.-Y.; Ryu, H.-S. Characteristics of cyanobacterial occurrence in the large river with consecutive weirs. In Proceedings of the 10th International Conference on Toxic Cyanobacteria—Cyanobacteria and Cyanotoxins; From Research to Risk Management, Wuhan, China, 23–28 October 2016; p. 35. [Google Scholar]
- Cirés, S.; Ballot, A. A review of the phylogeny, ecology and toxin production of bloom-forming Aphanizomenon spp. and related species within the Nostocales (cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y. Environmental factors that determine the occurrence and seasonal dynamics of Aphanizomenon flos-aquae. J. Limnol. 2009, 68, 122–132. [Google Scholar] [CrossRef]
- Üveges, V.; Tapolczai, K.; Krienitz, L.; Padisák, J. Photosynthetic characterisitics and physiological plasticity of an Aphanizomenon flos-aquae (Cyanobacteria, Nostocaceae) winter bloom in a deep oligo-mesotrophic lake (Lake Stechlin, Germany). Hydrobiologia 2016, 698, 263–272. [Google Scholar] [CrossRef]
- Suurnäkki, S.; Gomez-Saez, G.V.; Rantala-Ylinen, A.; Jokela, J.; Fewer, D.P.; Sivonen, K. Identification of geosmin and 2-methylisoborneol in cyanobacteria and molecular detection methods for the producers of these compounds. Water Res. 2015, 68, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Euler, K.L. Chemical studies on toxins from the blue-green alga Aphanizomenon flos-aquae. In The Water Environment; Springer: New York, NY, USA, 1981; pp. 405–414. [Google Scholar]
- Mahmood, N.A.; Carmichael, W.W. Paralytic shellfish poisons produced by the freshwater Cyanobacterium Aphanizomenon flos-aquae NH-5. Toxicon 1986, 24, 175–186. [Google Scholar] [CrossRef]
- Jung, J.-M.; Lee, Y.-J.; Pak, H.-K.; Jung, E.-Y.; Joo, G.-J. The analysis of cyanobacterial neurotoxins by high-performance liquid chromatography-mass spectrometry. Algae 2003, 18, 233–238. [Google Scholar] [CrossRef]
- Li, R.H.; Carmichael, W.W.; Liu, Y.D.; Watanabe, M.M. Taxonomic re-evaluation of Aphanizomenon flos-aquae NH-5 based on morphology and 16S rRNA gene sequences. Hydrobiologia 2000, 438, 99–105. [Google Scholar] [CrossRef]
- Li, R.H.; Carmichael, W.W.; Pereira, P. Morphological and 16S rRNA gene evidence for reclassification of the paralytic shellfish toxin producing Aphanizomenon flos-aquae LMECYA 31 as Aphanizomenon issatschenkoi (Cyanophyceae). J. Phycol. 2003, 39, 289–295. [Google Scholar] [CrossRef]
- Komárek, J. Freshwater Flora of Central Europe Vol. 19/3 Cyanoprokaryota, 3rd Part: Heterocytous Genera; Springer: Berlin, Germany, 2013. [Google Scholar]
- Kim, K.; Park, C.; Yoon, Y.; Hwang, S.-J. Harmful Cyanobacterial Material Production in the North Han River (South Korea): Genetic Potential and Temperature-Dependent Propertieds. Int. J. Environ. Res. Public Health 2018, 15, 444. [Google Scholar]
- Water Information System. Available online: Water.nier.go.kr/front/algaeInfo/algaeGuide.jsp (accessed on 15 November 2016).
- Komárek, J.; Anagnostidis, K. Freshwater Flora of Central Europe Vol. 19/1 Cyanoprokaryota, 1st Part: Chroococcales; Gustav Fischer Verlag: Jena, Germany, 1999. [Google Scholar]
- John, D.M.; Whitton, B.M.; Brook, A.J. The Freshwater Algal Flora of the British Isles; An Identification Guide to Freshwater and Terrestrial Algae, 2nd ed.; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Watanabe, M.M.; Nozaki, H. NIES-Collection List of Strains, 4th ed.; The National Institute for Environmental Studies, The Environmental Agency: Tsukuba, Japan, 1994. [Google Scholar]
- Hisbergues, M.; Christiansen, G.; Rouhiainen, L.; Sivonen, K.; Börner, T. PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch. Microbiol. 2003, 180, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Nonneman, D.; Zimba, D.V. A PCR-based test to assess the potential for microcystin occurrence in channel catfish production ponds. J. Phycol. 2002, 38, 230–233. [Google Scholar] [CrossRef]
- Kellmann, R.; Mills, T.; Neilan, B.A. Functional modeling and phylogenetic distribution of putative cylindrospermopsin biosynthesis enzymes. J. Mol. Evol. 2006, 62, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Tebrineh, J.A.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [PubMed]
- Gkelis, S.; Papadimitriou, T.; Zaoutsos, N.; Leonardos, I. Anthropogenic and climate-induced change favors toxic cyanobacteria blooms: Evidence from monitoring a highly eutrophic, urban mediterranean lake. Harmful Algae 2014, 39, 322–333. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Steche, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Park, H.-K.; Hong, D.-G.; Choi, S.-K. Characteristics of geosmin production by Dolichospermum planctonicum isolated from the Nakdong River, Korea. In Proceedings of the 2017 International Symposium in Commemoration of the 50th Anniversary of Korean Society of Limnoly, Harmful Algal Blooms: Understanding and Management, Gwangju, Korea, 18 October 2017; p. 35. [Google Scholar]
- Laamanen, M.J.; Forsström, L.; Sivonen, K. Diversity of Aphanizomenon flos-aquae (Cyanobacterium) populations along a Baltic Sea salinity gradient. Appl. Environ. Microbiol. 2002, 68, 5296–5303. [Google Scholar] [CrossRef] [PubMed]
- Janse, I.; Meima, M.; Kardinaal, W.E.A.; Zwart, G. High-resolution differentiation of cyanobacteria by using rRNA-internal transcribed spacer denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2003, 69, 6634–6643. [Google Scholar] [CrossRef] [PubMed]
- Nelissen, B.; Wilmotte, A.; Neefs, J.M.; De Wachter, R. Phylogenetic relationships among filamentous helical cyanobacteria investigated on the basis of 16S ribosomal RNA gene sequence analysis. Syst. Appl. Microb. 1994, 17, 206–210. [Google Scholar] [CrossRef]
- Omur-Ozbek, P. Global Taste and Odor Survey of Water Utilities, Final Report (October 12); The American Water Works Association Taste and Odor Committee: Denver, CO, USA, 2012; p. 43. [Google Scholar]
- You, K.-A.; Byeon, M.-S.; Youn, S.-J.; Hwang, S.-J.; Rhew, D.-H. Growth characteristics of blue-green algae (Anabaena spiroides) causing tastes and odors in the North-Han River, Korea. Korean J. Ecol. Environ. 2013, 46, 135–144. [Google Scholar] [CrossRef]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]
- Hoff-Risseti, C.; Dörr, F.A.; Schaker, P.D.C.; Pinto, E.; Werner, V.R.; Fiore, M.F. Cylindrospermopsin and saxitoxin synthetase genes in Cylindrospermopsis raciborskii strains from Brazilian freshwater. PLoS ONE 2013, 8, e74238. [Google Scholar] [CrossRef] [PubMed]
- Mihali, T.K.; Carmichael, W.W.; Neilan, B.A. A putative gene cluster from a Lyngbya wollei bloom that encodes paralytic shellfish toxin biosynthesis. PLoS ONE 2011, 6, e14657. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, P.J.; Gentile, J.H.; Sasner, J.J. Demonstration of a toxin from Aphanizomenon flos-aquae (L.) Ralfs. Can. J. Microbiol. 1968, 14, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, methods of detection and toxicology. J. Appl. Microbiol. 2012, 114, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xiao, P.; Yu, G.; Shao, J.; Liu, D.; Azevedo, S.M.F.; Li, R. Sporadic distribution and distinctive variations of cylindrospermopsin genes in cyanobacterial strains and environmental samples from Chinese freshwater bodies. Appl. Environ. Microbiol. 2014, 80, 5219–5230. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Ramm, J.; Eundberget, T.; Kaplan-Levy, R.N.; Hada, O.; Sukenik, A.; Wiedner, C. Occurrence of non-cylindrospermopsin-producing Aphanizomenon ovalisporum and Anabaena bergii in Lake Kinneret (Israel). J. Plankton Res. 2011, 33, 1736–1746. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Set | Sequence (5′—3′) | Fragment Length (bp) | References |
|---|---|---|---|---|
| 16S rRNA | CY16S-F1 | CAGGATGAACGCTGGCGG | 840 | This study |
| CY16S-R2 | GAATGGGATTAGATACCCCA | |||
| CY16S-F3 | AAAGGAGGTGATCCAGCCAC | 763 | This study | |
| CY16S-R1 | TCCTTTGAGTTTCACAGTTG | |||
| ITS | APHITS-F | ACAAGGTAGCCGTACCGGAA | 312/541 | This study |
| APHITS-R | ACAATTTCTTTTGCTTCCAC | |||
| geoA | geo78F | GCATTCCAAAGCCTGGGCTTA | 905 | [14] |
| geo982R | ATCGCATGTGCCACTCGTGAC | |||
| mcyA-Cd | mcyAF | AAAAGTGTTTTATTAGCGGCTCAT | 303 | [26] |
| mcyAR | AAAATTAAAAGCCGTATCAAA | |||
| mcyB | mcyB2959F | TGGGAAGATGTTCTTCAGGTATCCAA | 320 | [27] |
| mcyB3278R | AGAGTGGAAACAATATGATAAGCT AC | |||
| cyrA | CYLAT-F | ATTGTAAATAGCTGGAATGAGTGG | 1105–1179 | [28] |
| CYLAT-R | TTAGGGAAGTAATCTTCACAG | |||
| cyrJ | cynsulfF | ACTTCTCTCCTTTCCCTATC | 578 | [29] |
| cylnamR | GAGTGAAAATGCGTAGAACTTG | |||
| sxtA | sxtA-F | GATGACGGAGTATTTGAAGC | 125 | [30] |
| sxtA-R | CTGCATCTTCTGGACGGTAA | |||
| sxtI | sxtI-F | GCTTACTACCACGATAGTGCTGCCG | 1669 | [31] |
| sxtI-R | GGTTCGCCGCGGACATTAAA |
| Strain No. (NRERC-) | Scientific Name | Collection Date | Isolation Source | NCBI GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| 16S rRNA | ITS Type-1 | ITS Type-2 | ||||
| 006 | Aphanizomenon flos-aquae | 26 November 2016 | ND-2 | MF362173 | MF398429 | MF398428 |
| 007 | Aphanizomenon flos-aquae | 28 November 2016 | ND-3 | MF362174 | MF398424 | MF398427 |
| 008 | Aphanizomenon flos-aquae | 26 November 2016 | ND-2 | KY952630 | MF281187 | MF322523 |
| 009 | Aphanizomenon flos-aquae | 28 November 2016 | ND-3 | MF362175 | MF398425 | MF398423 |
| 010 | Aphanizomenon flos-aquae | 28 November 2016 | ND-3 | MF362177 | MF398426 | MF398422 |
| 011 | Aphanizomenon flos-aquae | 26 November 2016 | ND-2 | MF362176 | MF398430 | MF422681 |
| 012 | Aphanizomenon flos-aquae | 28 November 2016 | ND-3 | MG234522 | MG234520 | MG234525 |
| 013 | Aphanizomenon flos-aquae | 28 November 2016 | ND-3 | MG234521 | MG234519 | MG234527 |
| 016 | Aphanizomenon flos-aquae | 8 June 2017 | GR | MG234523 | MG234526 | MG234528 |
| 018 | Aphanizomenon flos-aquae | 8 June 2017 | GR | MG234518 | MG234524 | MG234529 |
| Sampling Date | ND-2 | ND-3 | ||||
|---|---|---|---|---|---|---|
| Aphanizomenon spp. | Microcystis spp. | Dolichospermum spp. | Aphanizomenon spp. | Microcystis spp. | Dolichospermum spp. | |
| 25 April 2016 | 19 | ND | ND | 43 | ND | ND |
| 30 May 2016 | ND | 1525 | ND | 700 | 17,280 | ND |
| 20 June 2016 | 600 | 1868 | 314 | 15,080 | 7143 | 602 |
| 25 July 2016 | ND | 12 | 23 | 365 | 3630 | 375 |
| 22 August 2016 | ND | 3270 | ND | 2184 | 71,625 | ND |
| 26 September 2016 | 35 | 83 | ND | 202 | 89 | 80 |
| 24 October 2016 | ND | 18 | ND | 64 | 51 | ND |
| 28 November 2016 | ND | ND | ND | 1532 | ND | ND |
| 26 December 2016 | ND | ND | ND | 227 | 50 | ND |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-K.; Kwon, M.-A.; Lee, H.-J.; Oh, J.; Lee, S.-H.; Kim, I.-S. Molecular Verification of Bloom-forming Aphanizomenon flos-aquae and Their Secondary Metabolites in the Nakdong River. Int. J. Environ. Res. Public Health 2018, 15, 1739. https://doi.org/10.3390/ijerph15081739
Park H-K, Kwon M-A, Lee H-J, Oh J, Lee S-H, Kim I-S. Molecular Verification of Bloom-forming Aphanizomenon flos-aquae and Their Secondary Metabolites in the Nakdong River. International Journal of Environmental Research and Public Health. 2018; 15(8):1739. https://doi.org/10.3390/ijerph15081739
Chicago/Turabian StylePark, Hae-Kyung, Mi-Ae Kwon, Hae-Jin Lee, Jonghee Oh, Su-Heon Lee, and In-Soo Kim. 2018. "Molecular Verification of Bloom-forming Aphanizomenon flos-aquae and Their Secondary Metabolites in the Nakdong River" International Journal of Environmental Research and Public Health 15, no. 8: 1739. https://doi.org/10.3390/ijerph15081739
APA StylePark, H.-K., Kwon, M.-A., Lee, H.-J., Oh, J., Lee, S.-H., & Kim, I.-S. (2018). Molecular Verification of Bloom-forming Aphanizomenon flos-aquae and Their Secondary Metabolites in the Nakdong River. International Journal of Environmental Research and Public Health, 15(8), 1739. https://doi.org/10.3390/ijerph15081739