Influence of Algae Age and Population on the Response to TiO2 Nanoparticles


Abstract
1. Introduction
2. Materials and Methods
2.1. Algae
2.2. Nanoparticles
2.3. Test Chambers
2.4. Exposure Regime
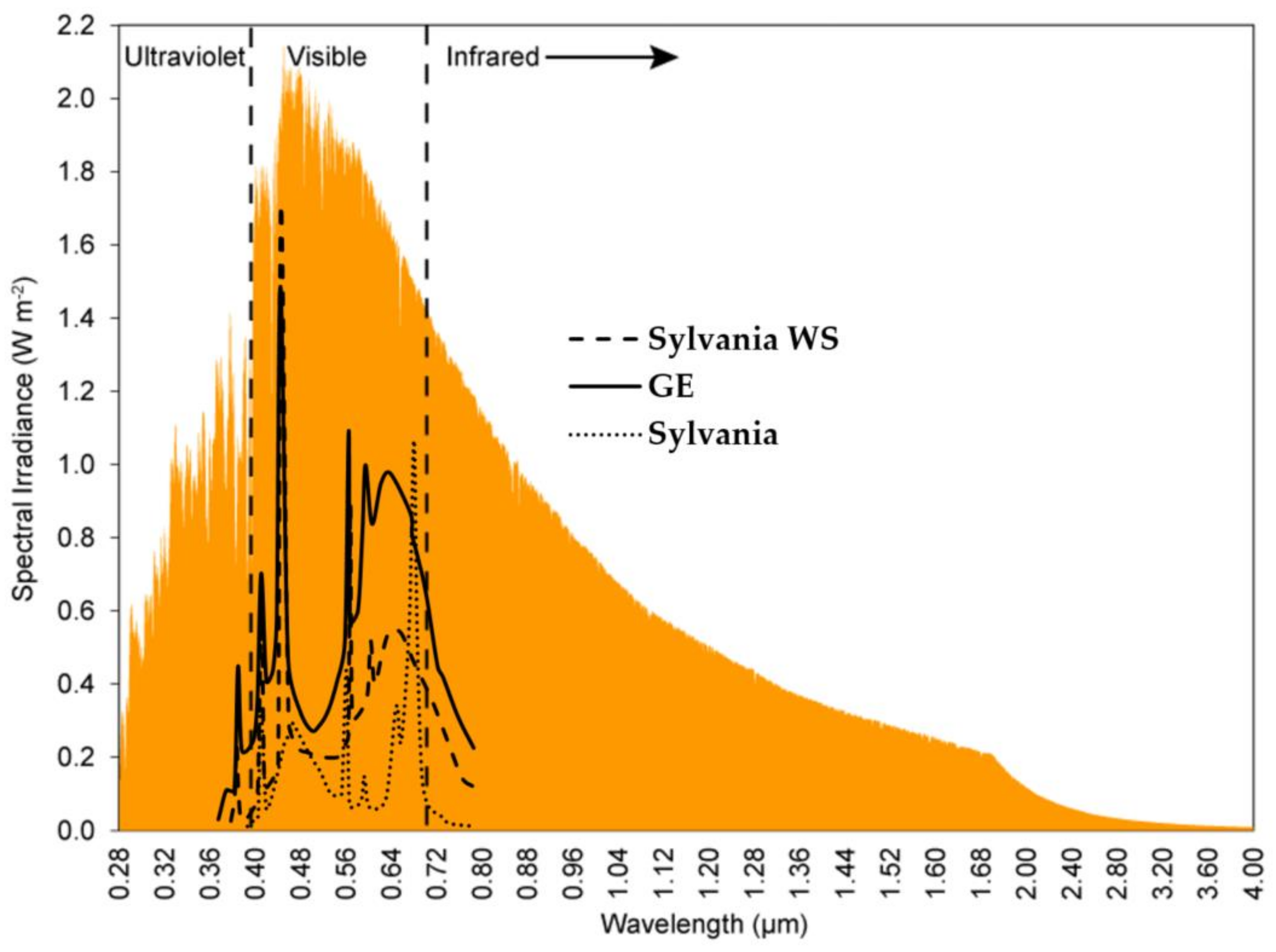
2.5. Measurements
3. Results and Discussions
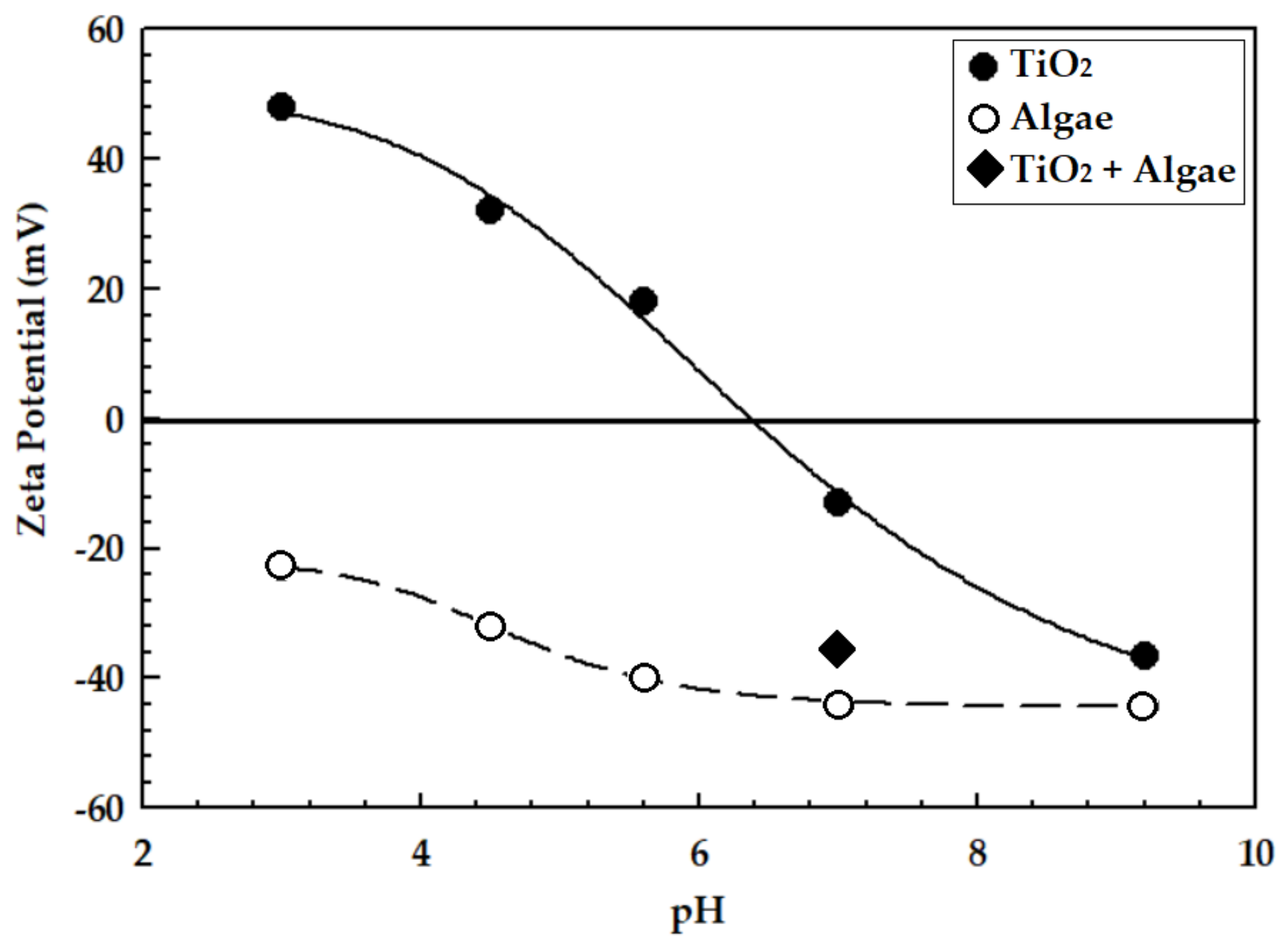
3.1. Nanoparticle Characterization
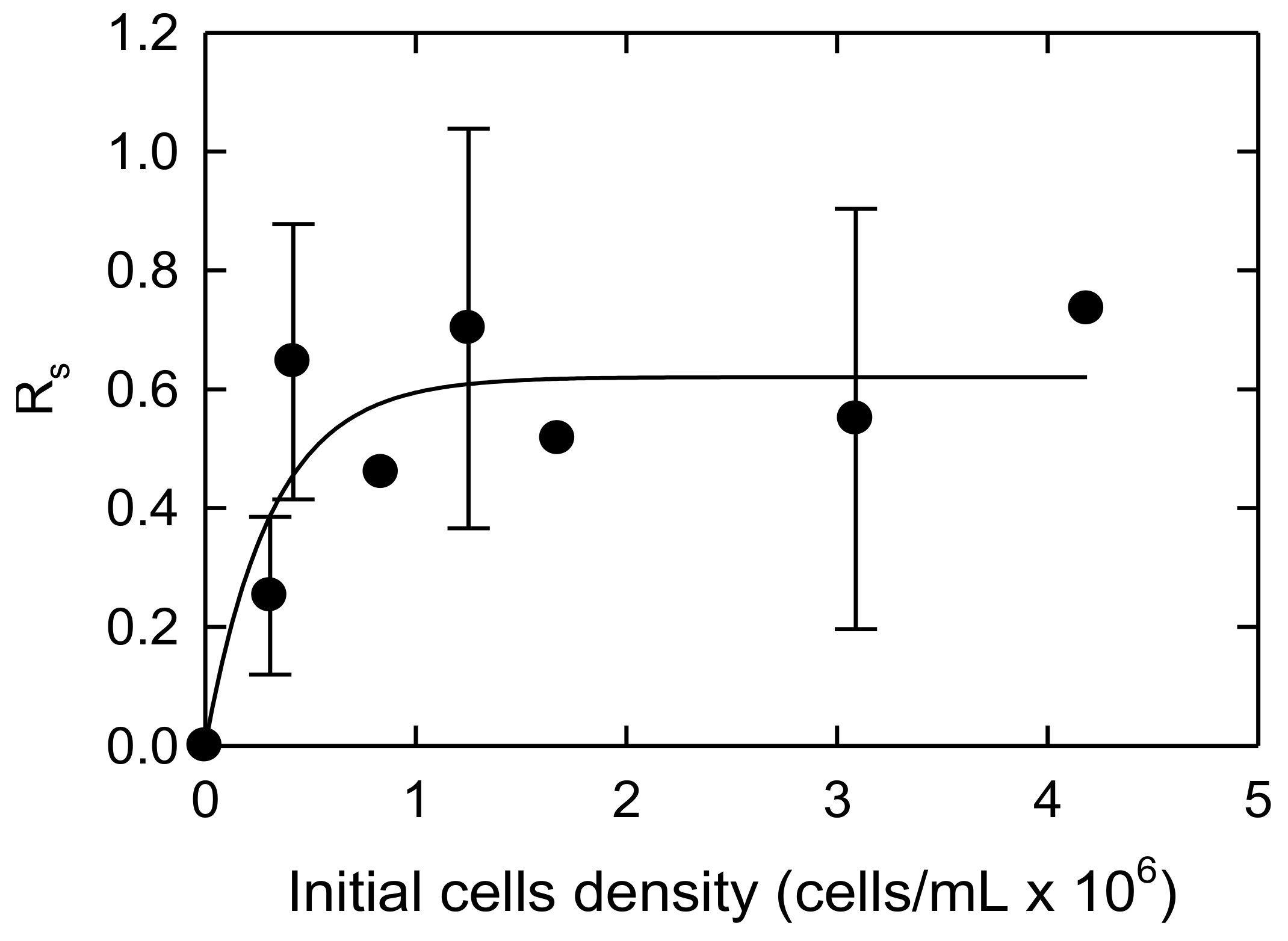
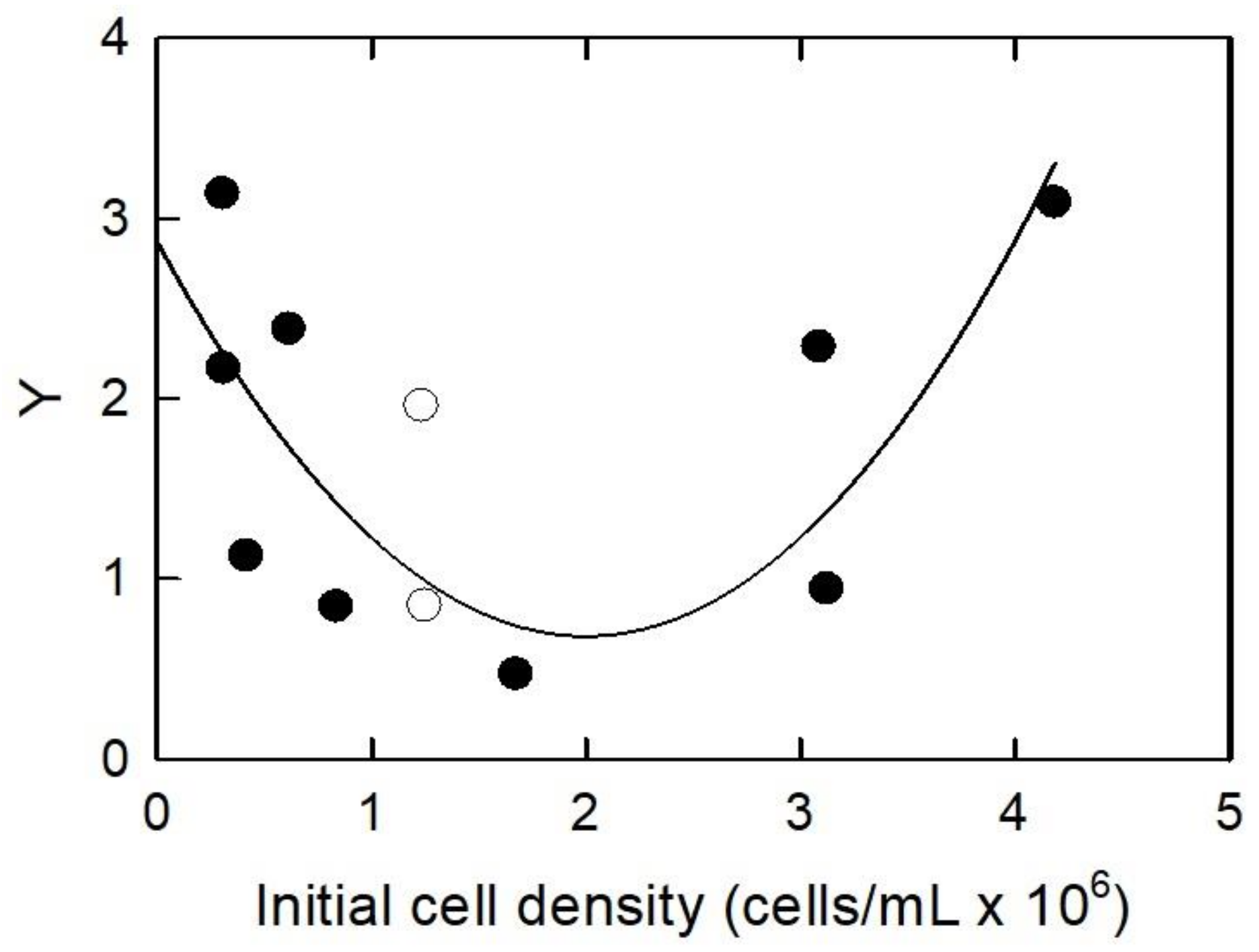
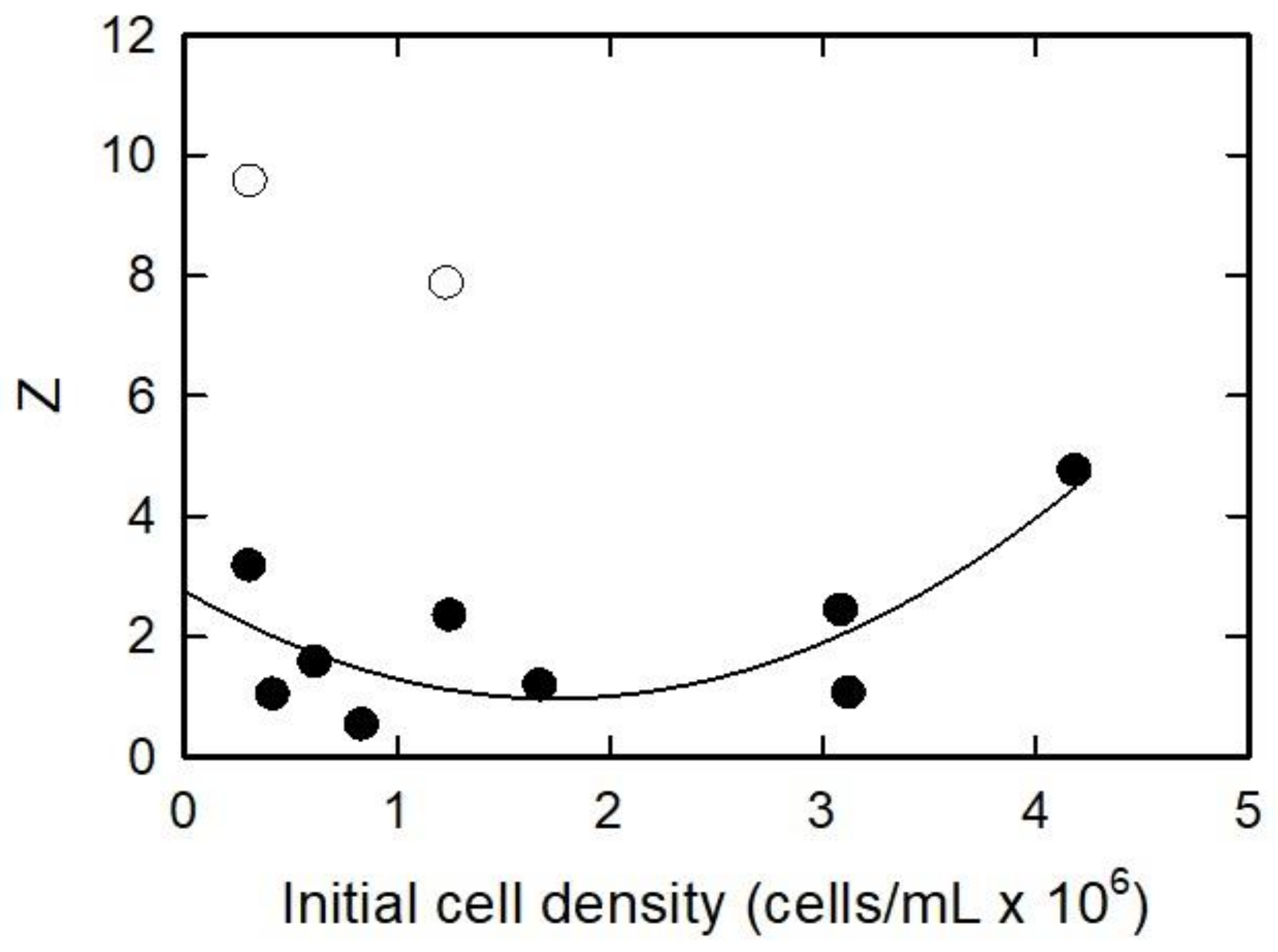
3.2. Initial Algal Population
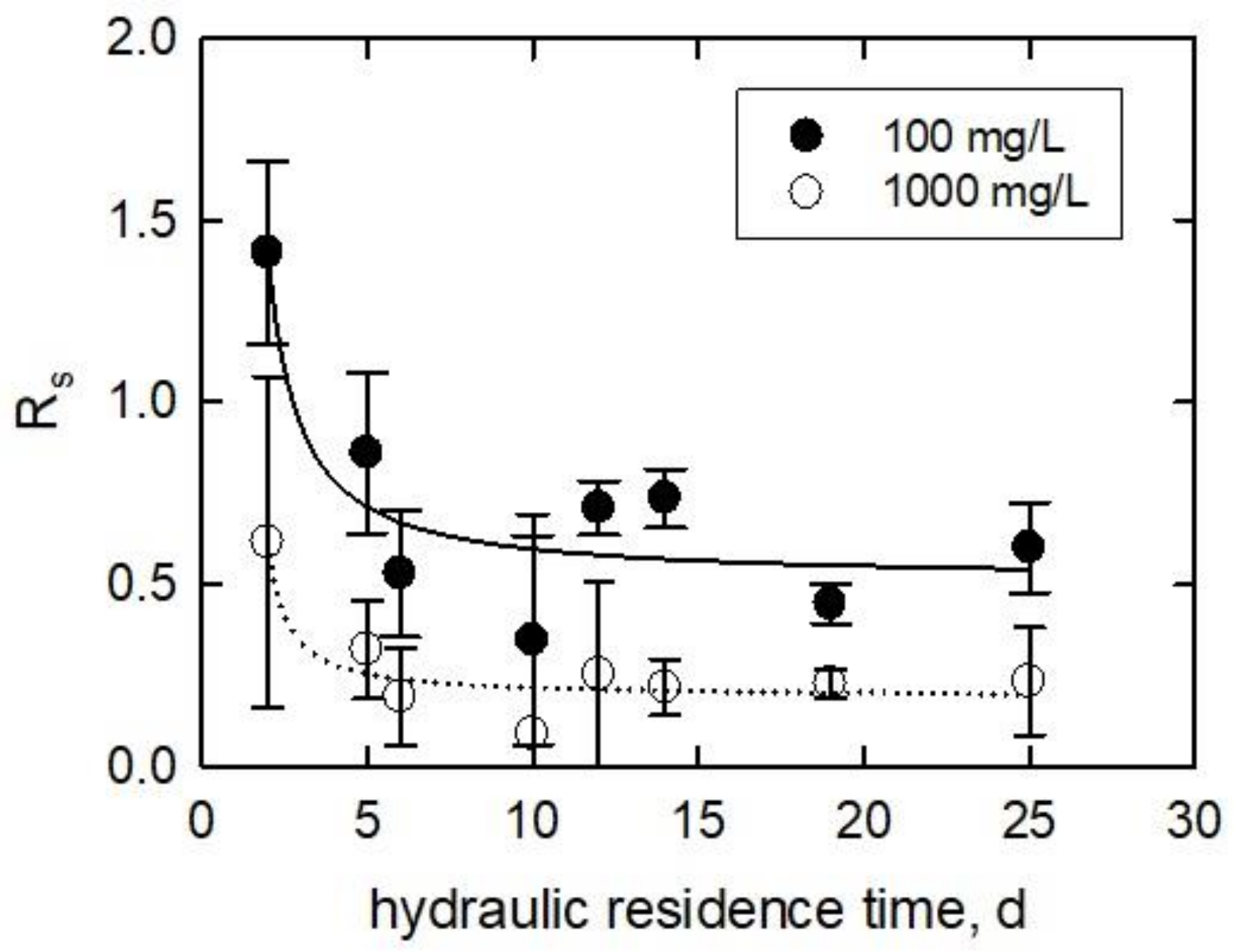
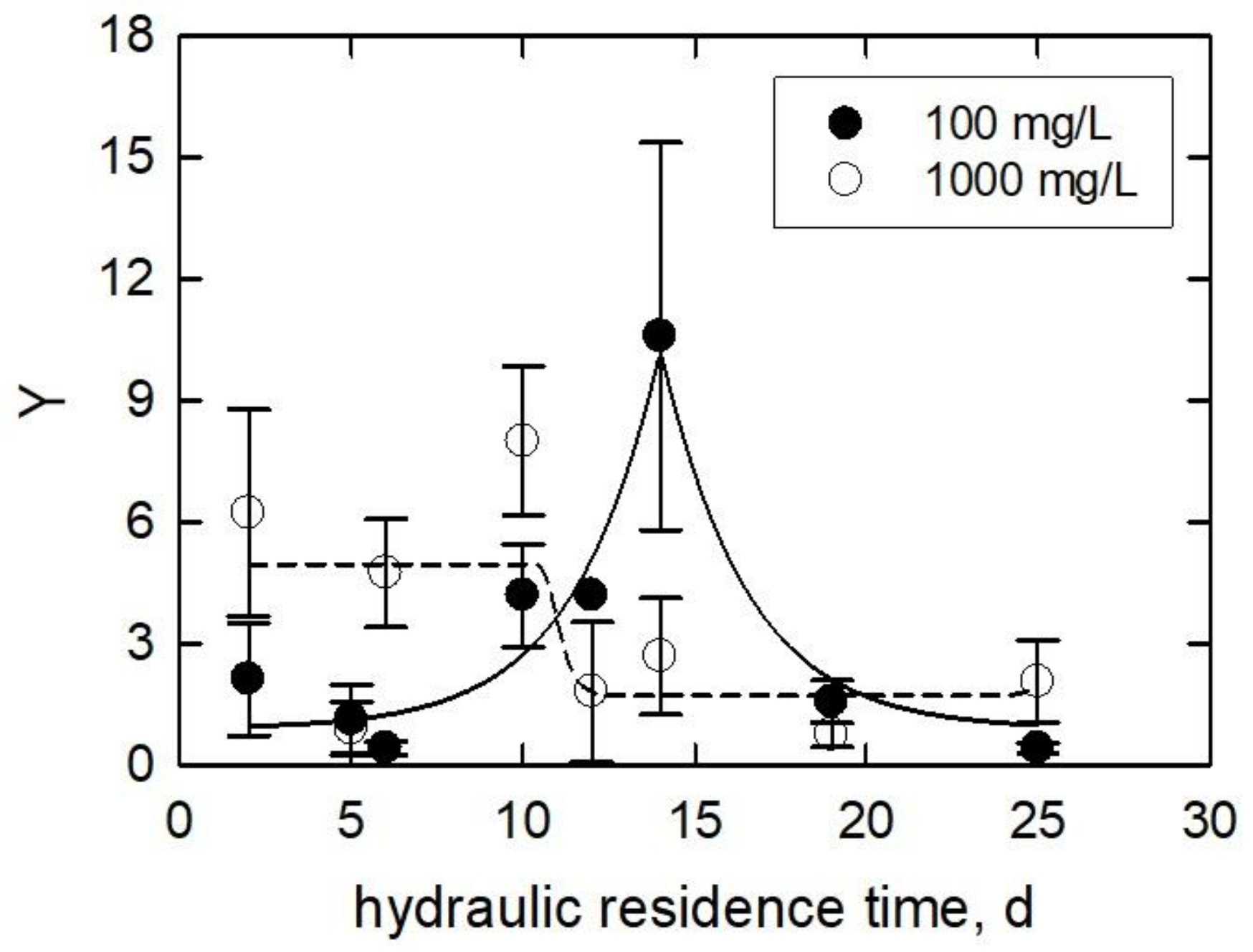
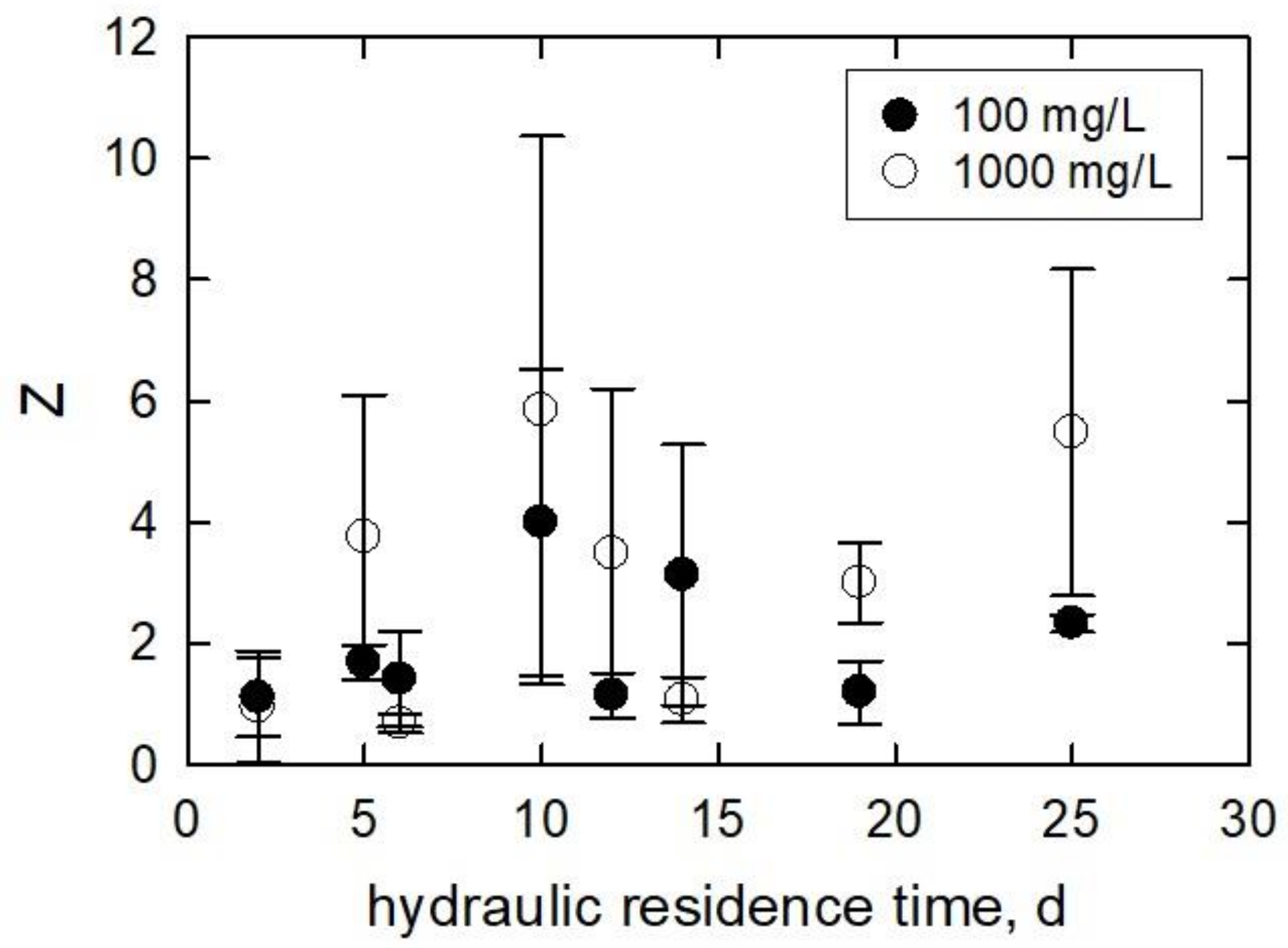
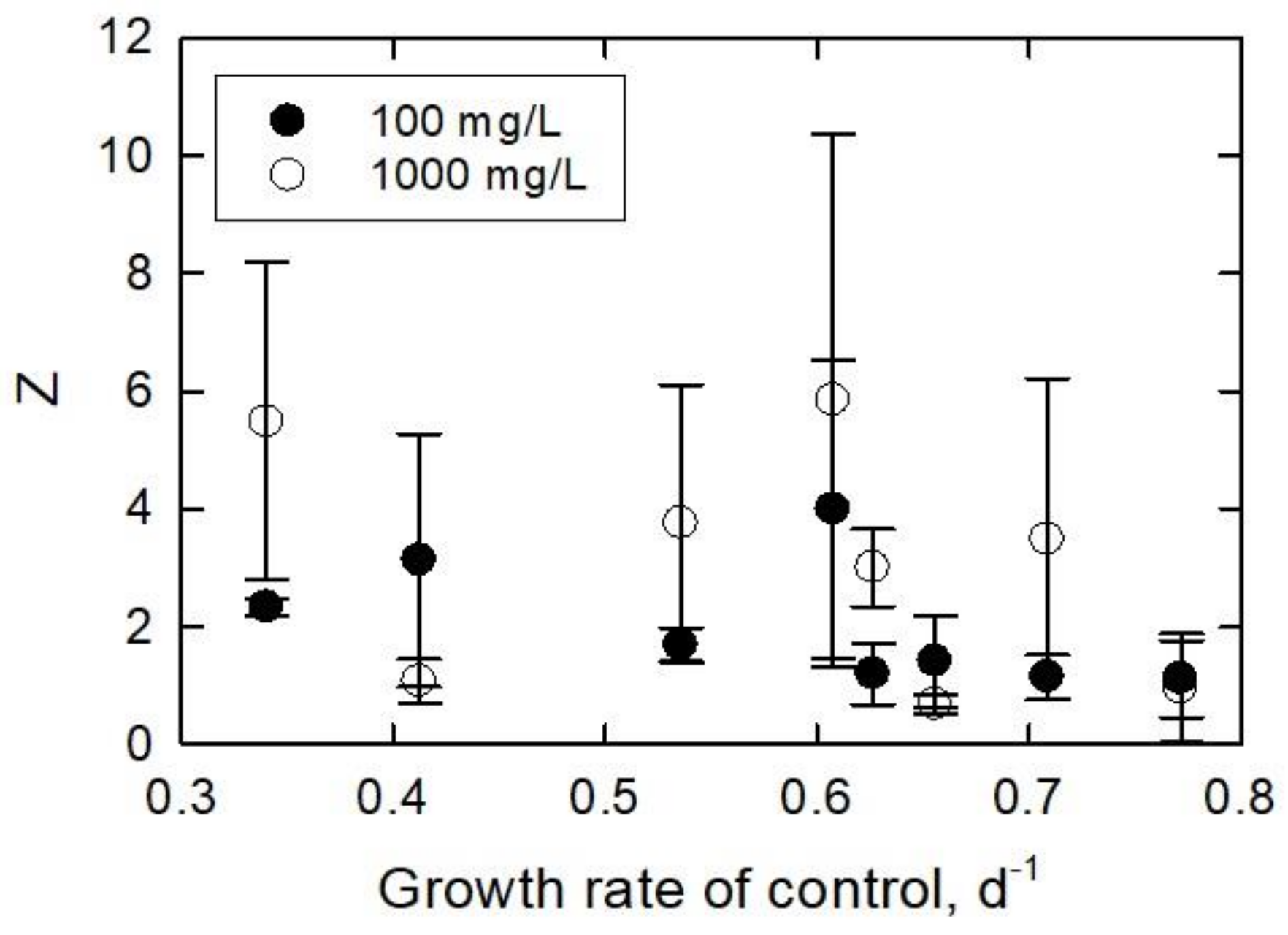
3.3. Algae at Various Hydraulic Residence Times
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Galley, E.; Fardell, N.A. Sunscreen with Coated Microfine Particles. U.S. Patent 5,609,852, 11 March 1997. [Google Scholar]
- Canesi, L.; Fabbri, R.; Gallo, G.; Vallotto, D.; Marcomini, A.; Pojana, G. Biomarkers in Mytilus galloprovincialis exposed to suspensions of selected nanoparticles (Nano carbon black, C60 fullerene, Nano-TiO2, Nano-SiO2). Aquat. Toxicol. 2010, 100, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Clemente, Z.; Castro, V.L.; Jonsson, C.M.; Fraceto, L.F. Minimal levels of ultraviolet light enhance the toxicity of TiO2 nanoparticles to two representative organisms of aquatic systems. J. Nanopart. Res. 2014, 16, 2559. [Google Scholar] [CrossRef]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed]
- Robichaud, C.O.; Uyar, A.E.; Darby, M.R.; Zucker, L.G.; Wiesner, M.R. Estimates of upper bounds and trends in nano-TiO2 production as a basis for exposure assessment. Environ. Sci. Technol. 2009, 43, 4227–4233. [Google Scholar] [CrossRef] [PubMed]
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J. Nanopart. Res. 2012, 14, 1109. [Google Scholar] [CrossRef]
- Keller, A.A.; McFerran, S.; Lazareva, A.; Suh, S. Global life cycle releases of engineered nanomaterials. J. Nanopart. Res. 2013, 15, 1692–1709. [Google Scholar] [CrossRef]
- Boxall, A.; Chaudhry, Q.; Sinclair, C.; Jones, A.; Aitken, R.; Jefferson, B.; Watts, C. Current and Future Predicted Environmental Exposure to Engineered Nanoparticles; Central Science Laboratory: York, UK, 2007. [Google Scholar]
- Mueller, N.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, F.; Ort, C.; Scholz, R.W.; Nowack, B. Engineered nanomaterials inrivers-exposure scenarios for Switzerland at high spatial and temporal resolution. Environ. Pollut. 2011, 159, 3439–3445. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, F.; Lassen, C.; Kjoelholt, J.; Christensen, F.; Nowack, B. Modeling flows and concentrations of nine engineered nanomaterials in the Danish environment. Int. J. Environ. Res. Public Health 2015, 12, 5581–5602. [Google Scholar] [CrossRef] [PubMed]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry: Chemical Equilibria and Rates in Natural Waters, 3rd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Oomen, A.G.; Bleeker, E.A.J.; Bos, P.M.J.; van Broekhuizen, F.; Gottardo, S.; Groenewold, M.; Hristozov, D.; Hund-Rinke, K.; Irfan, M.A.; Marcomini, A.; et al. Grouping and Read-Across Approaches for Risk Assessment of Nanomaterials. Int. J. Environ. Res. Public Health 2015, 12, 13415–13434. [Google Scholar] [CrossRef] [PubMed]
- Bos, P.M.J.; Gottardo, S.; Scott-Fordsmand, J.J.; van Tongeren, M.; Semenzin, E.; Fernandes, T.F.; Hristozov, D.; Hund-Rinke, K.; Hunt, N.; Irfan, M.A.; et al. The MARINA Risk Assessment Strategy: A Flexible Strategy for Efficient Information Collection and Risk Assessment of Nanomaterials. Int. J. Environ. Res. Public Health 2015, 12, 15007–15021. [Google Scholar] [CrossRef] [PubMed]
- Teske, S.S.; Detweiler, C.S. The Biomechanisms of Metal and Metal-Oxide Nanoparticles’ Interactions with Cells. Int. J. Environ. Res. Public Health 2015, 12, 1112–1134. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.K.; Lyon, D.Y.; Alvarez, P.J.J. Comparative eco-toxicity of nanoscale TiO2, SiO2, and ZnO water suspensions. Water Res. 2006, 40, 3527–3532. [Google Scholar] [CrossRef] [PubMed]
- Amiano, I.; Olabarrieta, J.; Vitorica, J.; Zorita, S. Acute toxicity of nanosized TiO2 to Daphnia magna under UVA irradiation. Environ. Toxicol. Chem. 2012, 31, 2564–2566. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; An, Y.J. Effects of zinc oxide and titanium dioxide nanoparticles on green algae under visible, UVA, and UVB irradiations: No evidence of enhanced algal toxicity under UV pre-irradiation. Chemosphere 2013, 91, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Sendra, M.; Moreno-Garrido, I.; Yeste, M.P.; Gatica, J.M.; Blasco, J. Toxicity of TiO2, in nanoparticle or bulk form to freshwater and marine microalgae under visible light and UV-A radiation. Environ. Pollut. 2017, 227, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Metzler, D.; Li, M.; Erdem, A.; Huang, C.P. Responses of algae to photocatalytic nano-TiO2 particles with an emphasis on the effect of particle size. Chem. Eng. J. 2011, 170, 538–546. [Google Scholar] [CrossRef]
- Metzler, D.; Erdem, A.; Tseng, Y.H.; Huang, C.P. Responses of algal cells to engineered nanoparticles measured as algal cell population, chlorophyll a, and lipid peroxidation: Effect of particle size and type. J. Nanotechnol. 2012, 2012, 237284. [Google Scholar] [CrossRef]
- Ma, H.; Brennan, A.; Diamond, S.A. Phototoxicity of TiO2 nanoparticles under solar radiation to two aquatic species: Daphnia magna and Japanese medaka. Environ. Toxicol. Chem. 2012, 31, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wallis, L.K.; Diamond, S.A.; Ma, H.; Hoff, D.J. Species sensitivity and dependence on exposure conditions impacting the phototoxicity of TiO2 nanoparticles to benthic organisms. Environ. Toxicol. Chem. 2014, 33, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wallis, L.K.; Ma, H.; Diamond, S.A. Phototoxicity of TiO2 nanoparticles to a freshwater benthic amphipod: Are benthic systems at risk? Sci. Total Environ. 2014, 466–467, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Faria, M.; Navas, J.M.; Soares, A.M.; Barata, C. Oxidative stress effects of titanium dioxide nanoparticle aggregates in zebrafish embryos. Sci. Total Environ. 2014, 470–471, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Ozkaleli, M.; Erdem, A. Biotoxicity of TiO2 nanoparticles on Raphidocelis subcapitata microalgae exemplified by membrane deformation. Int. J. Environ. Res. Public Health 2018, 15, 416. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, C.M.; Alloy, M.M.; Hamilton, J.; Verbeck, G.F.; Newton, K.; Klaine, S.J.; Roberts, A.P. Photo-induced toxicity of titanium dioxide nanoparticles to Daphnia magna under natural sunlight. Chemosphere 2015, 120, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.; Peter, H.; Ciesielski, T.M.; Thomas, K.V.; Sommaruga, R.; Salvenmoser, W.; Weyhenmeyer, G.A.; Tranvik, L.J.; Jenssen, B.M. Impact of TiO2 nanoparticles on freshwater bacteria from three Swedish lakes. Sci. Total Environ. 2015, 535, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A.; Zaidi, W.; Samadani, M.; Dewez, D. Toxicity of nickel oxide nanoparticles on a freshwater green algal strain of Chlorella vulgaris. Biomed. Res. Int. 2017, 2017, 9528180. [Google Scholar] [CrossRef] [PubMed]
- OECD. Freshwater Alga and Cyanobacteria, Growth Inhibition Test; Organization for Economic Co-operation and Development: Paris, France, 2006. [Google Scholar]
- USEPA. Short-Term Methods for Estimating the Chronic Toxicity of Effluents and Receiving Waters to Freshwater Organisms, 4th ed.; EPA-821-R-02-013; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 2002.
- ISO. Water Quality–Fresh Water Algal Growth Inhibition Test with Unicellular Green Algae; International Organization for Standardization: Geneva, Switzerland, 2002. [Google Scholar]
- Creswell, L. Phytoplankton culture for aquaculture feed. SRAC 2010, 5004, 1–16. Available online: http://www2.ca.uky.edu/wkrec/PhytoplanktonAlgaeCulture.pdf (accessed on 8 January 2018).
- Hoang, T.C.; Klaine, S.J. Influence of organism age on metal toxicity to Daphnia magna. Environ. Toxicol. Chem. 2007, 26, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Rogevich, E.C.; Hoang, T.C.; Rand, G.M. The effects of water quality and age on the acute toxicity of copper to the florida apple snail, Pomacea paludosa. Arch. Environ. Contam. Toxicol. 2008, 54, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P. Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in symbiotic dinoflagellates. Limnol. Oceanogr. 1996, 41, 271–283. [Google Scholar] [CrossRef]
- Hoang, T.C.; Tomasso, J.R.; Klaine, S.J. Influence of water quality and age on nickel toxicity to fathead minnows (Pimephales promelas). Environ. Toxicol. Chem. 2004, 23, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, L.M.; Dickson, H.; Klanjscek, T.; Keller, A.A.; McCauley, E.; Nisbet, R.M. Environmental feedbacks and engineered nanoparticles: Mitigation of silver nanoparticle toxicity to Chlamydomonas reinhardtii by algal-produced organic compounds. PLoS ONE 2013, 8, e74456. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Garrido, I.; Lubián, L.M.; Soares, A.M.V.M. Influence of cellular density on determination of EC50 in microalgal growth inhibition tests. Ecotoxicol. Environ. Saf. 2000, 47, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Warheit, D.B.; Hoke, R.A.; Finlay, C.; Donner, E.M.; Reed, K.L.; Sayes, C.M. Development of a base set of toxicity tests using ultrafine TiO2 particles as a component of nanoparticle risk. Toxicol. Lett. 2007, 17, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, N.B.; Von der Kammer, F.; Hofmann, T.; Baalousha, M.; Ottofuelling, S.; Baun, A. Algal testing of titanium dioxide nanoparticles∔Testing considerations, inhibitory effects and modification of cadmium bioavailability. Toxicology 2010, 269, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Aruoja, V.; Dubourguier, H.C.; Kasemets, K.; Kahru, A. Toxicity of nanoparticles of CuO, ZnO and TiO2 to microalgae Pseudokirchneriella subcapitata. Sci. Total Environ. 2009, 407, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Metzler, D. Effect of Food-Borne Cadmium on the Reproduction of Ceriodaphnia Dubia. Master’s Thesis, University of Delaware, Newark, DE, USA, 2003. [Google Scholar]
- National Renewable Energy Laboratory (NREL). Best Practices Handbook for the Collection and Use of Solar Resource Data for Solar Energy Applications, 2nd ed.; Sengupta, M., Habte, A., Gueymard, C., Wilbert, S., Renné, D., Stoffel, T.U.S., Eds.; Department of Energy Office of Energy Efficiency & Renewable Energy Operated by the Alliance for Sustainable Energy: Alexandria, VA, USA, 2017; p. 233.
- Weber, C.I.; Fay, L.A.; Collins, G.B.; Rathke, D.E.; Tobin, J. Review of Methods for the Analysis of Chlorophyll in Periphyton and Plankton of Marine and Freshwater Systems; Ohio State University Sea Grant Program Technical Bulletin, OHSU-TB-15, 1–71; Ohio State University: Columbus, OH, USA, 1986. [Google Scholar]
- Maness, P.C.; Smolinski, S.; Blake, D.M.; Huang, Z.; Wolfrum, E.J.; Jacoby, W.A. Bactericidal activity of photocatalytic TiO2 reaction: Toward an understanding of its killing mechanism. Appl. Environ. Microbiol. 1999, 65, 4094–4098. [Google Scholar] [PubMed]
- Ottofuelling, S.; Von der Kammer, F.; Hofmann, T. Commercial titanium dioxide nanoparticles in both natural and synthetic water: Comprehensive multidimensional testing and prediction of aggregation behavior. Environ. Sci. Technol. 2011, 45, 10045–10052. [Google Scholar] [CrossRef] [PubMed]
- Raza, G.; Amjad, M.; Kaur, I.; Wen, D. Stability and aggregation kinetics of titania nanomaterials under environmentally realistic conditions. Environ. Sci. Technol. 2016, 50, 8462–8472. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Tseng, Y.H.; Huang, C.P. Interactions between nano-TiO2 particles and algal cells at moderate particle concentration. Front. Chem. Sci. Eng. 2015, 9, 242–257. [Google Scholar] [CrossRef]
- Hoffmann, M.R.; Martin, S.T.; Choi, W.; Bahnemann, D.W. Environmental Applications of Semiconductor Photocatalysis. Chem. Rev. 1995, 95, 69–96. [Google Scholar] [CrossRef]
- Kosmulski, M. pH-dependent surface charging and points of zero charge. IV. Update and new approach. J. Colloid Interface Sci. 2009, 337, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Guzman, K.A.; Finnegan, M.P.; Banfield, J.F. Influence of surface potential on aggregation and transport of titania nanoparticles. Environ. Sci. Technol. 2006, 40, 7688–7693. [Google Scholar] [CrossRef] [PubMed]
- Othman, S.H.; Rashid, S.A.; Ghazi, T.I.M.; Abdullah, N. Dispersion and stabilization of photocatalytic TiO2 nanoparticles in aqueous suspension for coatings applications. J. Nanomater. 2012, 2012, 2. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, H.; Liu, C.; Deng, Y. Growth, pigment composition, chlorophyll fluorescence and antioxidant defenses in red alga Gracilaria. S. Afr. J. Bot. 2012, 100, 27–32. [Google Scholar] [CrossRef]
- Dietz, K.J.; Turkan, I.; Krieger-Liskay, A. Redox-and reactive oxygen species–dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Muyssen, B.T.A.; Janssen, C.R. Age and exposure duration as a factor influencing Cu and Zn toxicity toward Daphnia magna. Ecotoxicol. Environ. Saf. 2007, 68, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.Q.; Wang, W.X. Kinetic uptake of bioavailable cadmium, selenium, and zinc by Daphnia magna. Environ. Toxicol. Chem. 2002, 21, 2348–2355. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, T.H.; Solbe’, J.; Kloepper-Sams, P.J. Analysis of the ecetoc aquatic toxicity (EAT) database III —Comparative toxicity of chemical substances to different life stages of aquatic organisms. Chemosphere 1998, 36, 129–142. [Google Scholar] [CrossRef]
- Stuhlbacher, A.; Bradley, M.C.; Naylor, C.; Calow, P. Variation in the development of cadmium resistance in Daphnia magna straus; Effect of temperature, nutrition, age and genotype. Environ. Pollut. 1993, 80, 153–158. [Google Scholar] [CrossRef]
- Kulandaivelu, G.; Senger, H. Changes in the reactivity of the photosynthetic apparatus in heterotrophic ageing cultures of Scenedesmus obliquus. I. Changes in the Photochemical Activities. Physiol. Plant. 1976, 36, 157–164. [Google Scholar] [CrossRef]
- Samuelsson, G.; Öquist, G. A method for studying photosynthetic capacities of unicellular algae based on in vivo chlorophyll fluorescence. Physiol. Plant. 1977, 40, 315–319. [Google Scholar] [CrossRef]
- Cheregi, O.; Kotabová, E.; Prášil, O.; Schröder, W.P.; Kaňa, R.; Funk, C. Presence of state transitions in the cryptophyte alga Guillardia theta. J. Exp. Bot. 2015, 66, 6461–6470. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A. Change in photosystem II photochemistry during algal growth phases of Chlorella vulgaris and Scenedesmus obliquus. Curr. Microbiol. 2016, 72, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.; Curtis, T.P.; Egerton, T.A.; Kosa, S.A.M.; Tinlin, J.R. Photoelectrocatalytic and photocatalytic disinfection of E. coli suspensions by titanium dioxide. Appl. Catal. B Environ. 2003, 41, 376–386. [Google Scholar] [CrossRef]
- Schneider, J.; Matsuoka, M.; Takeuchi, M.; Zhang, J.; Horiuchi, Y.; Anpo, M.; Bahnemann, D.W. Understanding TiO2 photocatalysis: Mechanisms and materials. Chem. Rev. 2014, 114, 9919–9986. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.A.; Havrilla, C.M.; Brady, T.C.; Abramo, K.H.; Levin, E.D. Oxidative stress in toxicology: Established mammalian and emerging piscine model systems. Environ. Health Perspect. 1998, 106, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Klaine, S.J.; Cho, J.; Kim, S.H.; Kim, S.D. Oxidative stress responses of Daphnia magna exposed to TiO2 nanoparticles according to size fraction. Sci. Total Environ. 2010, 408, 2268–2272. [Google Scholar] [CrossRef] [PubMed]
- Long, T.C.; Saleh, N.; Tilton, R.D.; Lowry, G.V.; Veronesi, B. Titanium Dioxide (P25) produces reactive oxygen species in immortalized brain microglia (BV2): Implications for nanoparticle neurotoxicity. Environ. Sci. Technol. 2006, 40, 4346–4352. [Google Scholar] [CrossRef] [PubMed]
- Kaduková, J.; Velgosová, O.; Mražíková, A.; Marcinčáková, R. The effect of culture age and initial Silver concentration on biosynthesis of Ag nanoparticles. Nova Biotechnol. Chim. 2014, 13, 28–37. [Google Scholar] [CrossRef]
- Adeleye, A.S.; Stevenson, L.M.; Su, Y.; Nisbet, R.M.; Zhang, Y.; Keller, A.A. Influence of phytoplankton on fate and effects of modified zerovalent iron nanoparticles. Environ. Sci. Technol. 2016, 50, 5597–5605. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Algae Population (Cell/mL) | Test Media Type | Calculated Ionic Strength (mM) | Particle Size (nm) | Half Maximal Effect Concentration (EC50, mg/L) | Reference No |
|---|---|---|---|---|---|
| 104 | Algal medium | 1.5 | 21 | 2.53 | [18] |
| 1.4 ± 0.9 × 106 | Algal medium | 640 | 42 | 25.5 | [21] |
| 2.6 ± 1.1 × 105 | Synthetic freshwater solutions | 0.5 | 212 ± 19 | 4.16 ± 0.05 | [26] |
| 1 | 287 ± 25 | 3.58 ± 0.16 | |||
| 2 | 546 ± 71 | 9.32 ± 0.11 | |||
| 8 | 1428 ± 202 | 12.14 ± 0.09 | |||
| 105 | Algal medium | 640 | 140 | 87 | [40] |
| 104 | Algal medium | 1.5 | ~30 | 71.1 | [41] |
| 3.5 × 106 | Algal medium | 640 | 25–70 | 5.38 | [42] |
| 3 × 106 | Algal medium | 620 | 140 and 380 | 87 and 61 | [43] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metzler, D.M.; Erdem, A.; Huang, C.P. Influence of Algae Age and Population on the Response to TiO2 Nanoparticles. Int. J. Environ. Res. Public Health 2018, 15, 585. https://doi.org/10.3390/ijerph15040585
Metzler DM, Erdem A, Huang CP. Influence of Algae Age and Population on the Response to TiO2 Nanoparticles. International Journal of Environmental Research and Public Health. 2018; 15(4):585. https://doi.org/10.3390/ijerph15040585
Chicago/Turabian StyleMetzler, David M., Ayca Erdem, and Chin Pao Huang. 2018. "Influence of Algae Age and Population on the Response to TiO2 Nanoparticles" International Journal of Environmental Research and Public Health 15, no. 4: 585. https://doi.org/10.3390/ijerph15040585
APA StyleMetzler, D. M., Erdem, A., & Huang, C. P. (2018). Influence of Algae Age and Population on the Response to TiO2 Nanoparticles. International Journal of Environmental Research and Public Health, 15(4), 585. https://doi.org/10.3390/ijerph15040585