
Recent Advances on Endocrine Disrupting Effects of UV Filters

Abstract
:1. Introduction
2. Endocrine Disrupting Effects of Typical UV Filters
2.1. Benzophenones
2.1.1. Estrogenic Disrupting Effects
2.1.2. Androgenic Disrupting Effects
2.1.3. Disrupting Effects toward Other Nuclear Receptors
2.2. Camphor Derivatives
2.2.1. Disrupting Effects toward Estrogen Receptor
2.2.2. Disrupting Effects toward Androgen Receptor
2.2.3. Disrupting Effects toward Progesterone Receptor
2.3. Cinnamate Derivatives
2.3.1. Disrupting Effects toward Estrogen Receptor
2.3.2. Disrupting Effects toward Thyroid Hormone Receptor
2.3.3. Disrupting Effects toward Other Nuclear Receptors
3. Perspectives
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morabito, K.; Shapley, N.C.; Steeley, K.G.; Tripathi, A. Review of sunscreen and the emergence of non-conventional absorbers and their applications in ultraviolet protection. Int. J. Cosmet. Sci. 2011, 33, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Diffey, B.L. Sources and measurement of ultraviolet radiation. Methods 2002, 28, 4–13. [Google Scholar] [CrossRef]
- Marie, C.; Cabut, S.; Vendittelli, F.; Sauvant-Rochat, M.P. Changes in cosmetics use during pregnancy and risk perception by women. Int. J. Environ. Res. Public Health 2016, 13, 383. [Google Scholar] [CrossRef] [PubMed]
- Wahie, S.; Lloyd, J.J.; Farr, P.M. Sunscreen ingredients and labelling: A survey of products available in the UK. Clin. Exp. Dermatol. 2007, 32, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Cruz, M.S.; Barceló, D. Chemical analysis and ecotoxicological effects of organic UV-absorbing compounds in aquatic ecosystems. TrAC Trends Anal. Chem. 2009, 28, 708–717. [Google Scholar] [CrossRef]
- Balmer, M.E.; Buser, H.R.; Müller, M.D.; Poiger, T. Occurrence of the organic UV-filter compounds BP-3,4-MBC, EHMC, and OC in wastewater, surface waters, and in fish from Swiss lakes. In Agroscope; Swiss Federal Research Station for Horticulture, Plant Protecting Chemistry: Wädenswil, Switzerland, 2004. [Google Scholar]
- Zhang, N.S.; Liu, Y.S.; Van den Brink, P.J.; Price, O.R.; Ying, G.G. Ecological risks of home and personal care products in the riverine environment of a rural region in South China without domestic wastewater treatment facilities. Ecotoxicol. Environ. Saf. 2015, 122, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Schlumpf, M.; Kypke, K.; Wittassek, M.; Angerer, J.; Mascher, H.; Mascher, D.; Vokt, C.; Birchler, M.; Lichtensteiger, W. Exposure patterns of UV filters, fragrances, parabens, phthalates, organochlor pesticides, PBDEs, and PCBs in human milk: Correlation of UV filters with use of cosmetics. Chemosphere 2010, 81, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Valle-Sistac, J.; Molins-Delgado, D.; Díaz, M.; Ibáñez, L.; Barceló, D.; Silvia Díaz-Cruz, M. Determination of parabens and benzophenone-type UV filters in human placenta. First description of the existence of benzyl paraben and benzophenone-4. Environ. Int. 2016, 88, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Groz, M.P.; Bueno, M.M.; Rosain, D.; Fenet, H.; Casellas, C.; Pereira, C.; Maria, V.; Bebianno, M.J.; Gomez, E. Detection of emerging contaminants (UV filters, UV stabilizers and musks) in marine mussels from Portuguese coast by QuEChERS extraction and GC–MS/MS. Sci. Total Environ. 2014, 493, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Gago-Ferrero, P.; Alonso, M.B.; Bertozzi, C.P.; Marigo, J.; Barbosa, L.; Cremer, M.; Secchi, E.R.; Azevedo, A.; Lailson-Brito, J., Jr.; Torres, J.P.M.; et al. First determination of UV Filters in marine mammals. Octocrylene levels in Franciscana dolphins. Environ. Sci. Technol. 2013, 47, 5619–5625. [Google Scholar] [CrossRef] [PubMed]
- Kunisue, T.; Chen, Z.; Buck Louis, G.M.; Sundaram, R.; Hediger, M.L.; Sun, L.; Kannan, K. Urinary concentrations of benzophenone-type UV Filters in U.S. women and their association with endometriosis. Environ. Sci. Technol. 2012, 46, 4624–4632. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.; Klit, A.; Jensen, M.B.; Søeborg, T.; Frederiksen, H.; Schlumpf, M.; Lichtensteiger, W.; Skakkebaek, N.E.; Drzewiecki, K.T. Sunscreens: Are they beneficial for health? An overview of endocrine disrupting properties of UV-filters. Int. J. Androl. 2012, 35, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.; Pirot, F.; Bertholle, V.; Roussel, L.; Falson, F.; Padois, K. Commonly used UV filter toxicity on biological functions: Review of last decade studies. Int. J. Cosmet. Sci. 2013, 35, 208–219. [Google Scholar] [CrossRef] [PubMed]
- French, J.E. NTP technical report on the toxicity studies of 2-Hydroxy-4-methoxybenzophenone (CAS No. 131-57-7) Administered topically and in dosed feed to F344/N rats and B6C3F1 mice. Toxic. Rep. Ser. 1992, 21, 1-E14. [Google Scholar] [PubMed]
- Axelstad, M.; Boberg, J.; Hougaard, K.S.; Christiansen, S.; Jacobsen, P.R.; Mandrup, K.R.; Nellemann, C.; Lund, S.P.; Hass, U. Effects of pre- and postnatal exposure to the UV-filter octyl methoxycinnamate (OMC) on the reproductive, auditory and neurological development of rat offspring. Toxicol. Appl. Pharmacol. 2011, 250, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Faass, O.; Schlumpf, M.; Reolon, S.; Henseler, M.; Maerkel, K.; Durrer, S.; Lichtensteiger, W. Female sexual behavior, estrous cycle and gene expression in sexually dimorphic brain regions after pre- and postnatal exposure to endocrine active UV filters. Neurotoxicology 2009, 30, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Kinnberg, K.L.; Petersen, G.I.; Albrektsen, M.; Minghlani, M.; Awad, S.M.; Holbech, B.F.; Green, J.W.; Bjerregaard, P.; Holbech, H. Endocrine-disrupting effect of the ultraviolet filter benzophenone-3 in zebrafish, Danio rerio. Environ. Toxicol. Chem. 2015, 34, 2833–2840. [Google Scholar] [CrossRef] [PubMed]
- Paredes, E.; Perez, S.; Rodil, R.; Quintana, J.B.; Beiras, R. Ecotoxicological evaluation of four UV filters using marine organisms from different trophic levels Isochrysis galbana, Mytilus galloprovincialis, Paracentrotus lividus, and Siriella armata. Chemosphere 2014, 104, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Molins-Delgado, D.; Gago-Ferrero, P.; Díaz-Cruz, M.S.; Barceló, D. Single and joint ecotoxicity data estimation of organic UV filters and nanomaterials toward selected aquatic organisms. Urban groundwater risk assessment. Environ. Res. 2016, 145, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Ping, S.; Liu, H.; Yang, S.; Wang, L.; Wang, Z. Acute toxicity of benzophenone-type UV filters for Photobacterium phosphoreum and Daphnia magna: QSAR analysis, interspecies relationship and integrated assessment. Chemosphere 2015, 135, 182–188. [Google Scholar]
- Schlumpf, M.; Cotton, B.; Conscience, M.; Haller, V.; Steinmann, B.; Lichtensteiger, W. In vitro and in vivo estrogenicity of UV screens. Environ. Health Perspect. 2001, 109, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.Y.; Galicia, H.F.; Fent, K. Assessment of hormonal activity of UV filters in tadpoles of frog Xenopus laevis at environmental concentrations. Mar. Environ. Res. 2004, 58, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, J. Differentiation of Brain and Reproductive Organs in Birds: Effects of Environmental Contaminants. Master’s Thesis, Uppsala University, Uppsala, Sweden, 2008. [Google Scholar]
- Ozáez, I.; Morcillo, G.; Martínez-Guitarte, J.L. Ultraviolet filters differentially impact the expression of key endocrine and stress genes in embryos and larvae of Chironomus riparius. Sci. Total Environ. 2016, 557–558, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.Y.; Galicia, H.F.; Fent, K. Comparison of in vitro and in vivo estrogenic activity of UV filters in fish. Toxicol. Sci. 2006, 90, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Coronado, M.; de Haro, H.; Deng, X.; Rempel, M.A.; Lavado, R.; Schlenk, D. Estrogenic activity and reproductive effects of the UV-filter oxybenzone (2-hydroxy-4-methoxyphenyl-methanone) in fish. Aquat. Toxicol. 2008, 90, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, D.; Sieratowicz, A.; Zielke, H.; Oetken, M.; Hollert, H.; Oehlmann, J. Ecotoxicological effect characterisation of widely used organic UV filters. Environ. Pollut. 2012, 163, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Maerkel, K.; Durrer, S.; Henseler, M.; Schlumpf, M.; Lichtensteiger, W. Sexually dimorphic gene regulation in brain as a target for endocrine disrupters: Developmental exposure of rats to 4-methylbenzylidene camphor. Toxicol. Appl. Pharmacol. 2007, 218, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Balázs, A.; Krifaton, C.; Orosz, I.; Szoboszlay, S.; Kovács, R.; Csenki, Z.; Urbányi, B.; Kriszt, B. Hormonal activity, cytotoxicity and developmental toxicity of UV filters. Ecotoxicol. Environ. Saf. 2016, 131, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Soto, L. Evaluation of the estrogenic effects of UV filters on the sergeant major damselfish, Abudefduf saxatilis. Cienc. Mar. 2014, 49, 187–196. [Google Scholar] [CrossRef]
- Morohoshi, K.; Yamamoto, H.; Kamata, R.; Shiraishi, F.; Koda, T.; Morita, M. Estrogenic activity of 37 components of commercial sunscreen lotions evaluated by in vitro assays. Toxicol. in Vitro 2005, 19, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kitamura, S.; Khota, R.; Sugihara, K.; Fujimoto, N.; Ohta, S. Estrogenic and antiandrogenic activities of 17 benzophenone derivatives used as UV stabilizers and sunscreens. Toxicol. Appl. Pharmacol. 2005, 203, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Molina-Molina, J.M.; Escande, A.; Pillon, A.; Gomez, E.; Pakdel, F.; Cavailles, V.; Olea, N.; Ait-Aissa, S.; Balaguer, P. Profiling of benzophenone derivatives using fish and human estrogen receptor-specific in vitro bioassays. Toxicol. Appl. Pharmacol. 2008, 232, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Weisbrod, C.J.; Kunz, P.Y.; Zenker, A.K.; Fent, K. Effects of the UV filter benzophenone-2 on reproduction in fish. Toxicol. Appl. Pharmacol. 2007, 225, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.Y.; Fent, K. Multiple hormonal activities of UV filters and comparison of in vivo and in vitro estrogenic activity of ethyl-4-aminobenzoate in fish. Aquat. Toxicol. 2006, 79, 305–324. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Kojima, H.; Takeuchi, S.; Uramaru, N.; Sanoh, S.; Sugihara, K.; Kitamura, S.; Ohta, S. Metabolism of UV-filter benzophenone-3 by rat and human liver microsomes and its effect on endocrine-disrupting activity. Toxicol. Appl. Pharmacol. 2015, 282, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, R.H.M.M.; Sonneveld, E.; Jansen, J.H.J.; Seinen, W.; Burg, B.V.D. Interaction of polycyclic musks and UV Filters with the estrogen receptor (ER), androgen receptor (AR), and progesterone receptor (PR) in reporter gene bioassays. Toxicol. Sci. 2004, 83, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Nashev, L.G.; Schuster, D.; Laggner, C.; Sodha, S.; Langer, T.; Wolber, G.; Odermatt, A. The UV-filter benzophenone-1 inhibits 17β-hydroxysteroid dehydrogenase type 3: Virtual screening as a strategy to identify potential endocrine disrupting chemicals. Biochem. Pharmacol. 2010, 79, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Schmutzler, C.; Bacinski, A.; Gotthardt, I.; Huhne, K.; Ambrugger, P.; Klammer, H.; Schlecht, C.; Hoang-Vu, C.; Grüters, A.; Wuttke, W.; et al. The ultraviolet filter benzophenone 2 interferes with the thyroid hormone axis in rats and is a potent in vitro Inhibitor of human recombinant thyroid peroxidase. Endocrinology 2007, 148, 2835–2844. [Google Scholar] [CrossRef] [PubMed]
- Jarry, H.; Christoffel, J.; Rimoldi, G.; Koch, L.; Wuttke, W. Multi-organic endocrine disrupting activity of the UV screen benzophenone 2 (BP2) in ovariectomized adult rats after 5 days treatment. Toxicology 2004, 205, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Schmutzler, C.; Gotthardt, I.; Hofmann, P.J.; Radovic, B.; Kovacs, G.; Stemmler, L.; Nobis, I.; Bacinski, A.; Mentrup, B.; Ambrugger, P.; et al. Endocrine disruptors and the thyroid gland—A combined in vitro and in vivo analysis of potential new biomarkers. Environ. Health Perspect. 2007, 115, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, R.; Lanser, P.; Seinen, W.; Van der Burg, B. Estrogenic activity of UV filters determined by an in vitro reporter gene assay and an in vivo transgenic zebrafish assay. Arch. Toxicol. 2002, 76, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Tinwell, H.; Lefevre, P.A.; Moffat, G.J.; Burns, A.; Odum, J.; Spurway, T.D.; Orphanides, G.; Ashby, J. Confirmation of uterotrophic activity of 3-(4-methylbenzylidine)camphor in the immature rat. Environ. Health. Perspect. 2002, 110, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Pillon, A.; Fenet, H.; Rosain, D.; Duchesne, M.J.; Nicolas, J.C.; Balaguer, P.; Casellas, C. Estrogenic activity of cosmetic components in reporter cell lines: Parabens, UV Screens, and musks. J. Toxicol. Environ. Health A 2005, 68, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Holbech, H.; Norum, U.; Korsgaard, B.; Poul, B. The chemical UV-filter 3-benzylidene camphor causes an oestrogenic effect in an in vivo fish assay. Pharmacol. Toxicol. 2002, 91, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.O.; Kling, M.; Firzani, P.A.; Mecky, A.; Duranti, E.; Shields-Botella, J.; Delansorne, R.; Broschard, T.; Kramer, P.J. Activation of estrogen receptor α and ERβ by 4-methylbenzylidene-camphor in human and rat cells: Comparison with phyto- and xenoestrogens. Toxicol. Lett. 2003, 142, 89–101. [Google Scholar] [CrossRef]
- Inui, M.; Adachi, T.; Takenaka, S.; Inui, H.; Nakazawa, M.; Ueda, M.; Watanabe, H.; Mori, C.; Lguchi, T.; Miyatake, K. Effect of UV screens and preservatives on vitellogenin and choriogenin production in male medaka (Oryzias latipes). Toxicology 2003, 194, 43–50. [Google Scholar] [CrossRef]
- Seidlovawuttke, D.; Jarry, H.; Christoffel, J.; Rimoldi, G.; Wuttke, W. Comparison of effects of estradiol (E2) with those of octylmethoxycinnamate (OMC) and 4-methylbenzylidene camphor (4MBC)—2 filters of UV light—On several uterine, vaginal and bone parameters. Toxicol. Appl. Pharmacol. 2006, 210, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Cotton, B.; Lichtensteiger, W.; Schlumpf, M. UV Filters with antagonistic action at androgen receptors in the MDA-kb2 Cell transcriptional-activation assay. Toxicol. Sci. 2003, 74, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ozáez, I.; Aquilino, M.; Morcillo, G.; Martínez-Guitarte, J.L. UV filters induce transcriptional changes of different hormonal receptors in Chironomus riparius embryos and larvae. Environ. Pollut. 2016, 214, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Maerkel, K.; Lichtensteiger, W.; Durrer, S.; Conscience, M.; Schlumpf, M. Sex- and region-specific alterations of progesterone receptor mRNA levels and estrogen sensitivity in rat brain following developmental exposure to the estrogenic UV filter 4-methylbenzylidene camphor. Environ. Toxicol. Pharmacol. 2005, 19, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Schmutzler, C.; Hamann, I.; Hofmann, P.J.; Kovacs, G.; Stemmler, L.; Mentrup, B.; Schomburg, L.; Ambrugger, P.; Grüters, A.; Seidlova-Wuttke, D.; et al. Endocrine active compounds affect thyrotropin and thyroid hormone levels in serum as well as endpoints of thyroid hormone action in liver, heart and kidney. Toxicology 2004, 205, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Klammer, H.; Schlecht, C.; Wuttke, W.; Schmutzler, C.; Gotthardt, I.; Kohrle, J.; Jarry, H. Effects of a 5-day treatment with the UV-filter octyl-methoxycinnamate (OMC) on the function of the hypothalamo-pituitary–thyroid function in rats. Toxicology 2007, 238, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ren, N.; Li, Y.F.; Kunisue, T.; Gao, D.; Kannan, K. Determination of benzotriazole and benzophenone UV Filters in sediment and sewage sludge. Environ. Sci. Technol. 2011, 45, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Tsui, M.M.P.; Leung, H.W.; Wai, T.C.; Yamashita, N.; Taniyasu, S.; Liu, W.; Lam, P.K.S.; Murphy, M.B. Occurrence, distribution and ecological risk assessment of multiple classes of UV filters in surface waters from different countries. Water Res. 2014, 67, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kannan, K. Characteristic profiles of benzonphenone-3 and its derivatives in urine of children and adults from the United States and China. Environ. Sci. Technol. 2013, 47, 12532–12538. [Google Scholar] [CrossRef] [PubMed]
- Loraine, G.A.; Pettigrove, M.E. Seasonal variations in concentrations of pharmaceuticals and personal care products in drinking water and reclaimed wastewater in Southern California. Environ. Sci. Technol. 2006, 40, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Schlumpf, M.; Schmid, P.; Durrer, S.; Conscience, M.; Maerkel, K.; Henseler, M.; Gruetter, M.; Herzog, I.; Reolon, S.; Ceccatelli, R.; et al. Endocrine activity and developmental toxicity of cosmetic UV filters—An update. Toxicology 2004, 205, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Ozáez, I.; Martínez-Guitarte, J.L.; Morcillo, G. Effects of in vivo exposure to UV filters (4-MBC, OMC, BP-3, 4-HB, OC, OD-PABA) on endocrine signaling genes in the insect Chironomus riparius. Sci. Total Environ. 2013, 456–457, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Fuentes, G.; Sandoval-Gío, J.J.; Arroyo-Silva, A.; Noreña-Barroso, E.; Escalante-Herrera, K.S.; Olvera-Espinosa, F. Evaluation of the estrogenic and oxidative stress effects of the UV filter 3-benzophenone in zebrafish (Danio rerio) eleuthero-embryos. Ecotoxicol. Environ. Saf. 2015, 115, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Kerdivel, G.; Le Guevel, R.; Habauzit, D.; Brion, F.; Ait-Aissa, S.; Pakdel, F. Estrogenic potency of benzophenone UV filters in breast cancer cells: Proliferative and transcriptional activity substantiated by docking analysis. PLoS ONE 2013, 8, e60567. [Google Scholar] [CrossRef] [PubMed]
- Correa, B.A.; Goncalves, A.S.; de Souza, A.M.; Freitas, C.A.; Cabral, L.M.; Albuquerque, M.G.; Castro, H.C.; dos Santos, E.P.; Rodrigues, C.R. Molecular modeling studies of the structural, electronic, and UV absorption properties of benzophenone derivatives. J. Phys. Chem. A 2012, 116, 10927–10933. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, S.; Zhang, C.; Liu, W. Atomic insights into distinct hormonal activities of bisphenol A analogues toward PPARγ and ERα receptors. Chem. Res. Toxicol. 2014, 27, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, J.; Tong, C.; Chen, Y.; Zhuang, S.; Liu, W. Molecular interactions of benzophenone UV filters with human serum albumin revealed by spectroscopic techniques and molecular modeling. J. Hazard. Mater. 2013, 263, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, S.; Zhang, J.; Wen, Y.; Zhang, C.; Liu, W. Distinct mechanisms of endocrine disruption of DDT-related pesticides toward estrogen receptor α and estrogen-related receptor γ. Environ. Toxicol. Chem. 2012, 31, 2597–2605. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.K.; Sarma, S.N.; Kim, Y.J.; Ryu, J.C. Toxicokinetics and metabolisms of benzophenone-type UV filters in rats. Toxicology 2008, 248, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Okereke, C.S.; Abdel-Rhaman, M.S.; Friedman, M.A. Disposition of benzophenone-3 after dermal administration in male rats. Toxicol. Lett. 1994, 73, 113–122. [Google Scholar] [CrossRef]
- Hayashi, T.; Okamoto, Y.; Ueda, K.; Kojima, N. Formation of estrogenic products from benzophenone after exposure to sunlight. Toxicol. Lett. 2006, 167, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geissler, W.M.; Davis, D.L.; Wu, L.; Bradshaw, K.D.; Patel, S.; Mendonca, B.B.; Elliston, K.O.; Wilson, J.D.; Russell, D.W.; Andersson, S. Male pseudohermaphroditism caused by mutations of testicular 17 beta-hydroxysteroid dehydrogenase 3. Nat. Genet. 1994, 7, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Taurog, A.; Dorris, M.L.; Doerge, D.R. Mechanism of simultaneous iodination and coupling catalyzed by thyroid peroxidase. Arch. Biochem. Biophys. 1996, 330, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Søeborg, T.; Ganderup, N.C.; Kristensen, J.H.; Bjerregaard, P.; Pedersen, K.L.; Bollen, P.; Hansen, S.H.; Halling-Sørensen, B. Distribution of the UV filter 3-benzylidene camphor in rat following topical application. J. Chromatogr. B 2006, 834, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Buser, H.R.; Balmer, M.E.; Schmid, P.; Kohler, M. Occurrence of UV filters 4-methylbenzylidene camphor and octocrylene in fish from various Swiss rivers with inputs from wastewater treatment plants. Environ. Sci. Technol. 2006, 40, 1427–1431. [Google Scholar] [CrossRef] [PubMed]
- Schlumpf, M.; Kypke, K.; Vökt, C.C.; Birchler, M.; Durrer, S.; Faass, O.; Ehnes, C.; Fuetsch, M.; Gaille, C.; Henseler, M. Endocrine active UV filters: Developmental toxicity and exposure through breast milk. Chimia 2008, 62, 345–351. [Google Scholar] [CrossRef]
- Hofkamp, L.; Bradley, S.; Tresguerres, J.; Lichtensteiger, W.; Schlumpf, M.; Timms, B. Region-specific growth effects in the developing rat prostate following fetal exposure to estrogenic ultraviolet filters. Environ. Health. Perspect. 2008, 116, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Schlumpf, M.; Jarry, H.; Wuttke, W.; Ma, R.; Lichtensteiger, W. Estrogenic activity and estrogen receptor beta binding of the UV filter 3-benzylidene camphor. Comparison with 4-methylbenzylidene camphor. Toxicology 2004, 199, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Durrer, S.; Maerkel, K.; Schlumpf, M.; Lichtensteiger, W. Estrogen target gene regulation and coactivator expression in rat uterus after developmental exposure to the ultraviolet filter 4-methylbenzylidene camphor. Endocrinology 2005, 146, 2130–2139. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Díaz, I.; Molina-Molina, J.M.; Zafra-Gómez, A.; Ballesteros, O.; Navalón, A.; Real, M.; Sáenz, J.M.; Fernández, M.F.; Olea, N. Simultaneous determination of the UV-filters benzyl salicylate, phenyl salicylate, octyl salicylate, homosalate, 3-(4-methylbenzylidene) camphor and 3-benzylidene camphor in human placental tissue by LC–MS/MS. Assessment of their in vitro endocrine activity. J. Chromatogr. B 2013, 936, 80–87. [Google Scholar]
- Giokas, D.L.; Salvador, A.; Chisvert, A. UV filters: From sunscreens to human body and the environment. TrAC Trends Anal. Chem. 2007, 26, 360–374. [Google Scholar] [CrossRef]
- Balmer, M.E.; Buser, H.R.; Müller, M.D.; Poiger, T. Occurrence of some organic UV filters in wastewater, in surface waters, and in fish from Swiss lakes. Environ. Sci. Technol. 2005, 39, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Kupper, T.; Plagellat, C.; Brandli, R.C.; de Alencastro, L.F.; Grandjean, D.; Tarradellas, J. Fate and removal of polycyclic musks, UV filters and biocides during wastewater treatment. Water Res. 2006, 40, 2603–2612. [Google Scholar] [CrossRef] [PubMed]
- Janjua, N.R.; Kongshoj, B.; Andersson, A.M.; Wulf, H.C. Sunscreens in human plasma and urine after repeated whole-body topical application. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Hamann, I.; Hofmann, P.; Schmutzler, C.; Mentrup, B.; Huhne, K.; Jarry, H.; Seidlová-Wuttke, D.; Wuttke, W.; KöHrle, J. 4MBC and OMC, components of UV-sunscreens, exert organ specific alterations on type I 5′-Deiodinase activity and expression in female rats. Exp. Clin. Endocrinol. Diabetes 2005, 113–138. [Google Scholar] [CrossRef]
- Zhuang, S.; Wang, H.; Ding, K.; Wang, J.; Pan, L.; Lu, Y.; Liu, Q.; Zhang, C. Interactions of benzotriazole UV stabilizers with human serum albumin: Atomic insights revealed by biosensors, spectroscopies and molecular dynamics simulations. Chemosphere 2016, 144, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Zhang, H.; Wang, H.; Lv, X.; Pan, L.; Zhang, W.; Zhuang, S. Atomic-scale investigation of the interactions between tetrabromobisphenol A, tetrabromobisphenol S and bovine trypsin by spectroscopies and molecular dynamics simulations. J. Hazard. Mater. 2015, 299, 486–494. [Google Scholar] [CrossRef] [PubMed]

| Compound | CAS No. | Chemical Structure | Kp (cm/h) * |
|---|---|---|---|
| BP-1 | 131-56-6 | 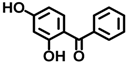 | 0.00917 |
| BP-2 | 131-55-5 | 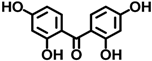 | 0.00458 |
| BP-3 | 131-57-7 | 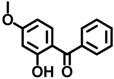 | 0.0271 |
| BP-4 | 4065-45-6 | 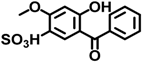 | 0.0000511 |
| 4-MBC | 36861-47-9 | 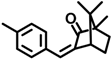 | 0.504 |
| 3-BC | 15087-24-8 | 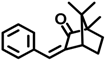 | 0.261 |
| OMC | 5466-77-3 | 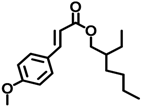 | 0.264 |
| IMC | 71617-10-2 | 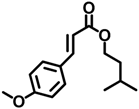 | 0.0477 |
| OC | 6197-30-4 | 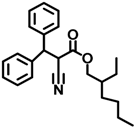 | 0.549 |
| UV Filters | Endocrine Disrupting Effects | References | |
|---|---|---|---|
| Benzophenones | Estrogenic disrupting effects | Activation of ERα, ERβ; Inhibition of the activity of 17β-Estradiol; Induction of proliferation of MCF-7 cell; Induction of VTG in fathead minnows; Reduce of the uterine weight in immature Long-Evans rats | [22,26,32,33,34,35,36,37] |
| Androgenic disrupting effects | Antagonists of human AR transactivation; Repression of 4,5-dihydrotestosterone-induced transactivational activity; Inhibition of testosterone formation in mice and rats | [34,36,37,38,39] | |
| Disrupting effects toward other nuclear receptors | Inhibition of human recombinant TPO; Interference with THR; Inhibition of TPO activity in rats; Antagonists of PR | [38,40,41,42] | |
| Camphor derivatives | Disrupting effects toward estrogen receptor | Activation of ERα, ERβ; Inhibition of the activity of 17β-Estradiol; Induction of proliferation of MCF-7 cell; Induction of pS2 protein in MCF-7 cells; Reduce of the uterine weight in rats; Induction of VTG in fish | [22,26,43,44,45,46,47,48,49] |
| Disrupting effects toward androgen receptor | Repression of 4,5-dihydrotestosterone-induced transactivational activity; Inhibition of testosterone formation in HEK-293 cells; Antagonists of Human AR | [36,38,39,50] | |
| Disrupting effects toward progesterone receptor | Antagonists of PR; Increase of PR mRNA levels in rats; Inhibition of the expression of PR protein in rats; Disturbance of the expression of membrane-associate PR in insects | [38,47,51,52] | |
| Cinnamate derivatives | Disrupting effects toward estrogen receptor | Activation of ERα; Inhibition of the activity of 17β-Estradiol; Induction of proliferation of MCF-7 cell; Reduce of the uterine weight in rats; Induction of VTG in fish | [22,36,43,45,48,49] |
| Disrupting effects toward thyroid hormone receptor | Decrease of T4 level; Inhibition of the conversion of T4 to triiodothyronine in rats | [16,53,54] | |
| Disrupting effects toward other nuclear receptors | Antagonists of PR and AR; Inhibition of 4,5-dihydrotestosterone activity; Reduce of the prostate and testicular weight in rats | [16,36,38] | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Pan, L.; Wu, S.; Lu, L.; Xu, Y.; Zhu, Y.; Guo, M.; Zhuang, S. Recent Advances on Endocrine Disrupting Effects of UV Filters. Int. J. Environ. Res. Public Health 2016, 13, 782. https://doi.org/10.3390/ijerph13080782
Wang J, Pan L, Wu S, Lu L, Xu Y, Zhu Y, Guo M, Zhuang S. Recent Advances on Endocrine Disrupting Effects of UV Filters. International Journal of Environmental Research and Public Health. 2016; 13(8):782. https://doi.org/10.3390/ijerph13080782
Chicago/Turabian StyleWang, Jiaying, Liumeng Pan, Shenggan Wu, Liping Lu, Yiwen Xu, Yanye Zhu, Ming Guo, and Shulin Zhuang. 2016. "Recent Advances on Endocrine Disrupting Effects of UV Filters" International Journal of Environmental Research and Public Health 13, no. 8: 782. https://doi.org/10.3390/ijerph13080782
APA StyleWang, J., Pan, L., Wu, S., Lu, L., Xu, Y., Zhu, Y., Guo, M., & Zhuang, S. (2016). Recent Advances on Endocrine Disrupting Effects of UV Filters. International Journal of Environmental Research and Public Health, 13(8), 782. https://doi.org/10.3390/ijerph13080782