Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds

Abstract
:
1. Introduction
2. Results and Discussion
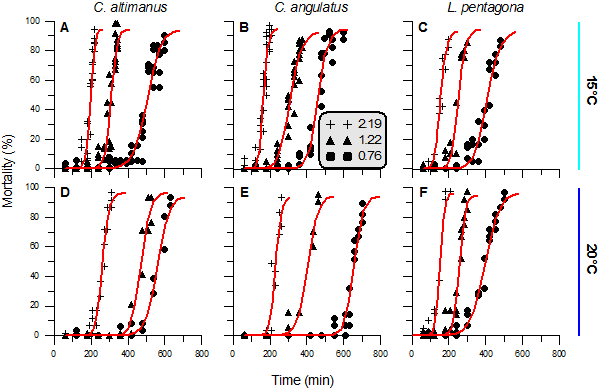
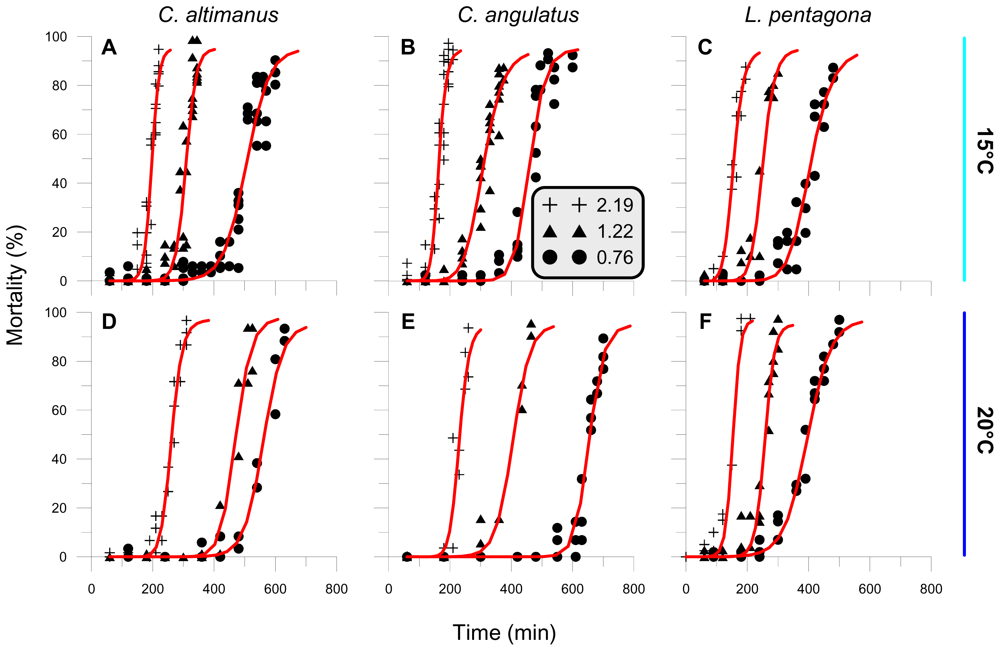
2.1. General responses of crab larvae to radiation and temperature
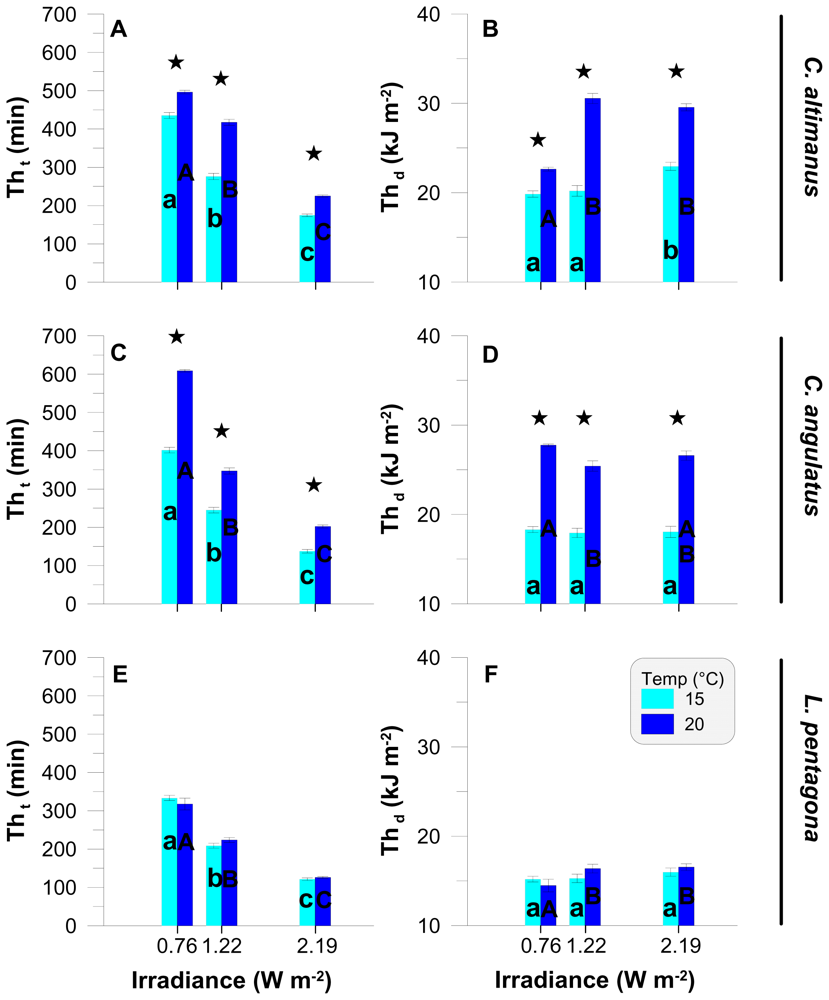
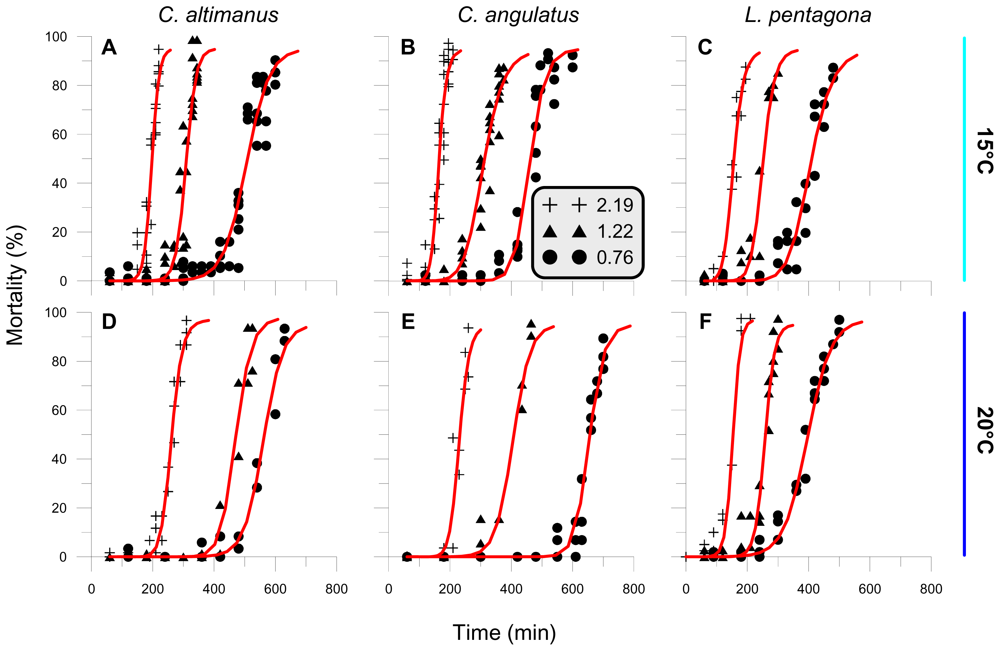
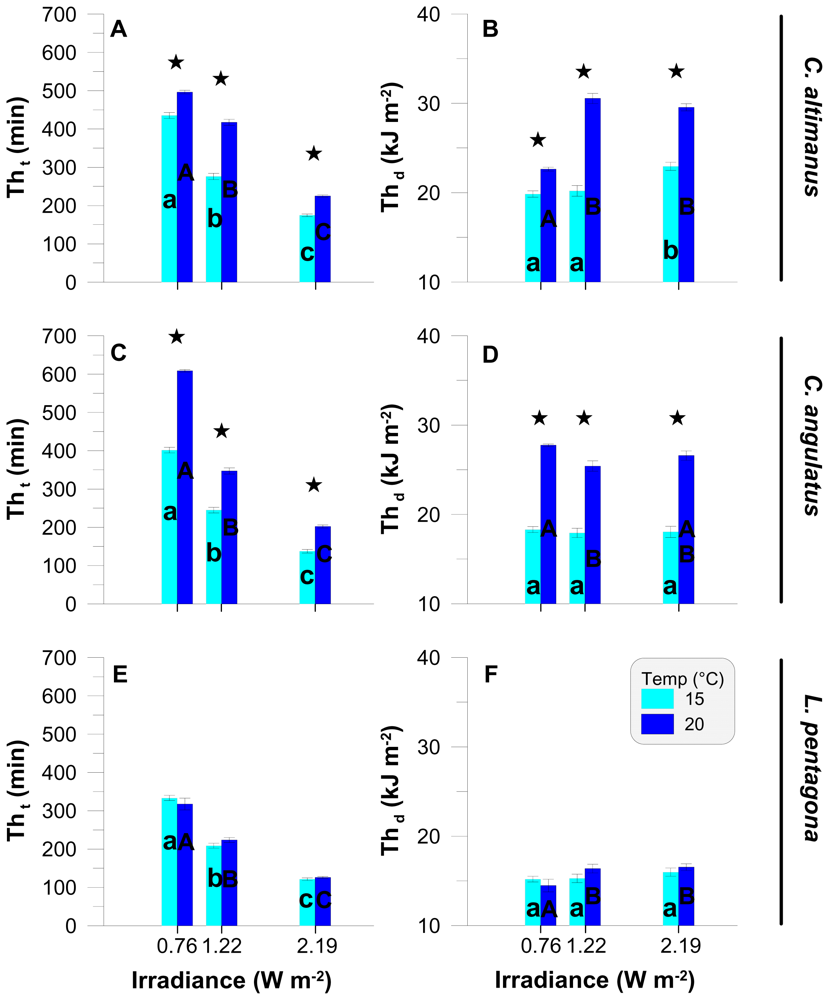
2.2. Threshold
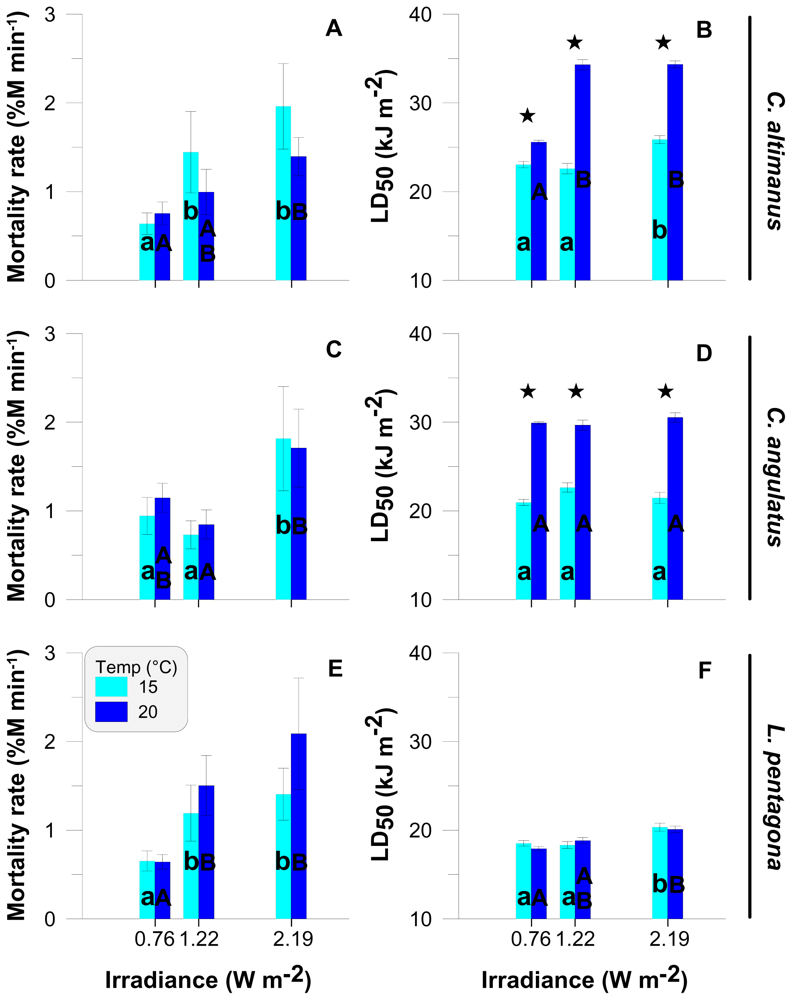
2.3. Rate of mortality and LD50
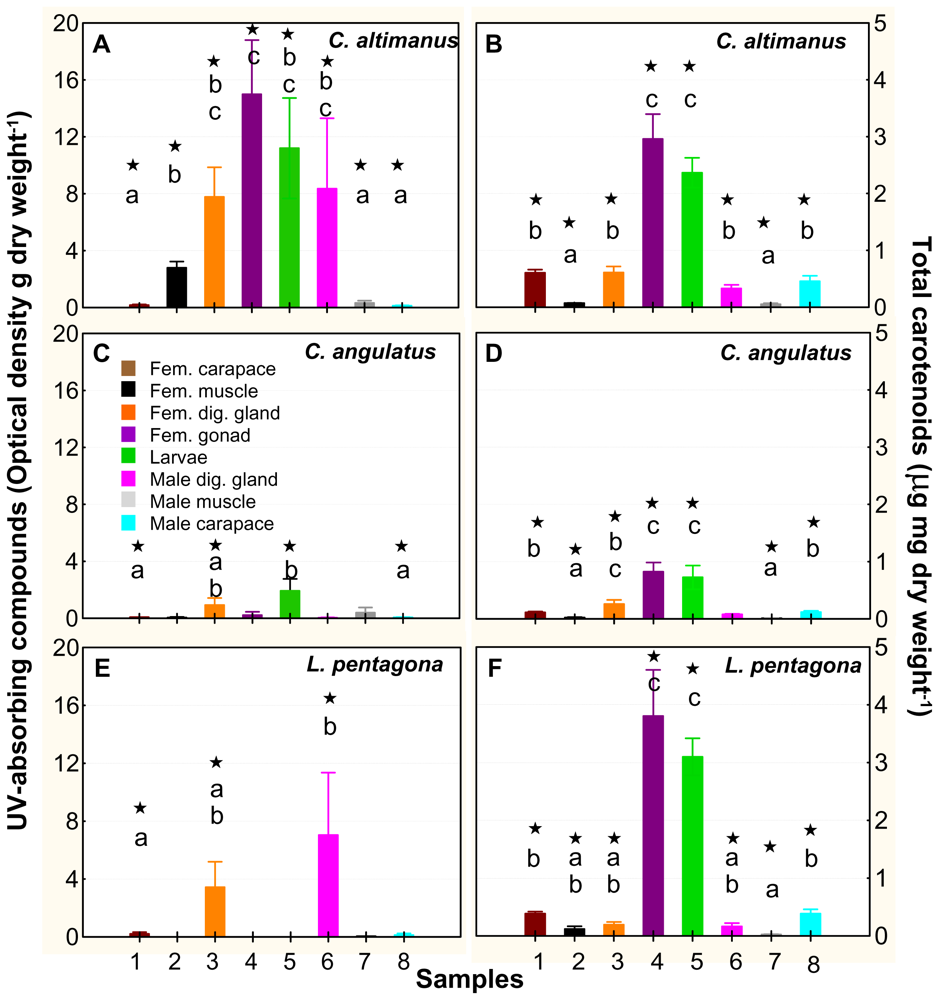
2.4. UV-absorbing compounds and carotenoids
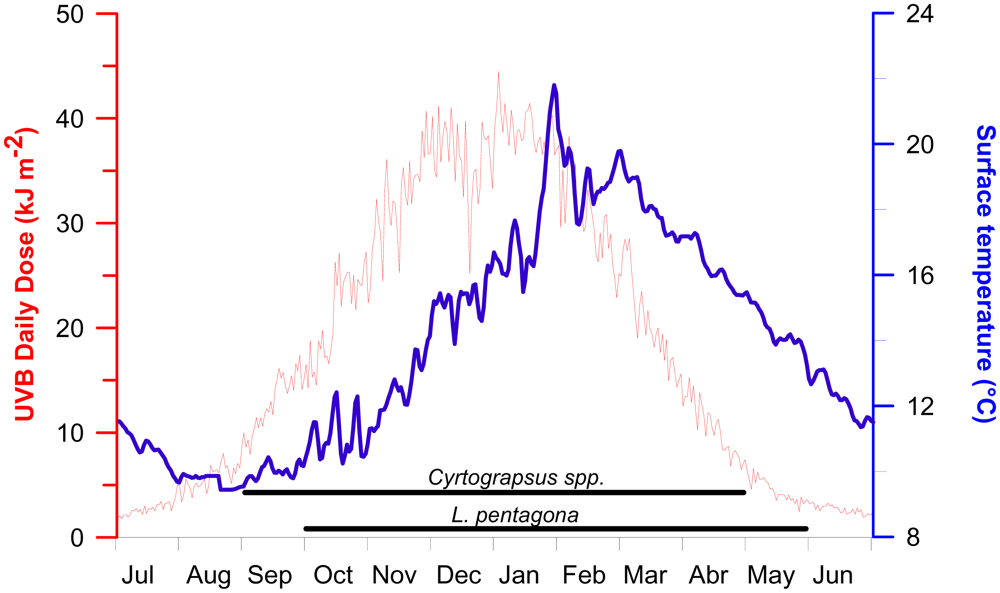
2.5. Ecological implications
3. Experimental Section
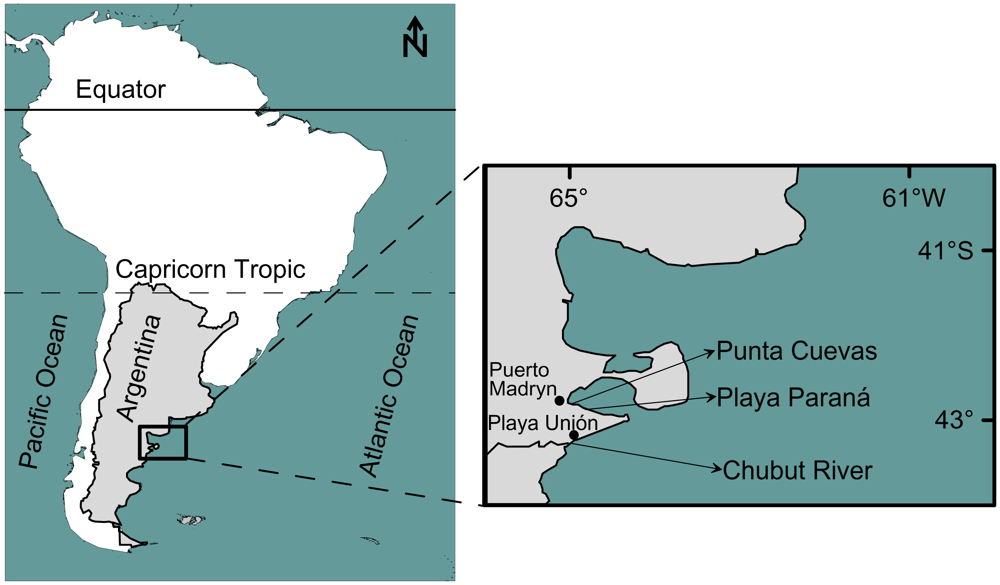
3.1. Sampling/maintenance of specimens
3.2. Experimental set up
3.3. Mortality determinations
3.4. Analysis of UV absorbing compounds
3.5. Analysis of carotenoids
3.6. Radiation/temperature data
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Harley, CDG; Hughes, AR; Hultgre, KM; Miner, BG; Sorte, CJB; Thornber, CS; Rodriguez, LF; Tomanek, L; Williams, SL. The impacts of climate change in coastal marine systems. Ecol Lett 2006, 9, 228–241. [Google Scholar]
- Nelson, GC; Bennet, E; Berhe, AA; Cassman, K; DeFries, R; Dietz, T; Dobermann, A; Dobson, A; Janetos, A; Levy, M; Marco, D; Nakicenovic, N; O’Neill, B; Norgaard, R; Petschel-Held, G; Ojima, D; Pingali, P; Watson, R; Zurek, M. Anthropogenic drivers of ecosystem change: An overview. Ecol Soc 2006, 11, 29. [Google Scholar]
- De Mora, SJ; Demers, S; Vernet, M. The Effects of UV Radiation on Marine Ecosystems; Cambridge University Press: Cambridge, UK, 2000; pp. 1–324. [Google Scholar]
- Helbling, EW; Zagarese, HE. UV Effects in Aquatic Organisms and Ecosystems; The Royal Society of Chemistry: Cambridge, UK, 2003; pp. 1–575. [Google Scholar]
- Häder, DP; Kumar, HD; Smith, RC; Worrest, RC. Effects of solar UV radiation on aquatic ecosystems and interactions with climate change. Photochem Photobiol Sci 2007, 6, 267–285. [Google Scholar]
- Setlow, RB; Grist, E; Thompson, K; Woodhead, AD. Wavelengths effective in induction of malignant melanoma. Proc Natl Acad Sci USA 1993, 90, 6666–6670. [Google Scholar]
- Vega, MP; Pizarro, RA. Oxidative stress and defence mechanisms of the freshwater cladoceran Daphnia longispina exposed to UV radiation. J Photochem Photobiol B Biol 2000, 54, 121–125. [Google Scholar]
- Lesser, MP; Farrell, JH; Walker, CW. Oxidative stress, DNA damage and p53 expression in the larvae of Atlantic cod (Gadus morhua) exposed to ultraviolet (290–400 nm) radiation. J Exp Biol 2001, 204, 157–164. [Google Scholar]
- Buma, AGJ; Boelen, P; Jeffrey, WH. Helbling, EW, Zagarese, HE, Eds.; UVR-induced DNA damage in aquatic organisms. In UV Effects in Aquatic Organisms and Ecosystems; The Royal Society of Chemistry: Cambridge, UK, 2003; pp. 291–327. [Google Scholar]
- Villafañe, VE; Sundbäck, K; Figueroa, FL; Helbling, EW. Helbling, EW, Zagarese, HE, Eds.; Photosynthesis in the aquatic environment as affected by UVR. In UV Effects in Aquatic Organisms and Ecosystems; Royal Society of Chemistry: Cambridge, UK, 2003; pp. 357–397. [Google Scholar]
- Anger, K; Thatje, S; Lovrich, G; Calcagno, J. Larval and early juvenile development of Paralomis granulosa reared at different temperatures: tolerance of cold and food limitation in a lithodid crab from high latitudes. Mar Ecol Prog Ser 2003, 253, 243–251. [Google Scholar]
- Williamson, CE; Grad, G; De Lange, HJ; Gilroy, S. Temperature-dependent ultraviolet responses in zooplankton: Implications of climate change. Limnol Oceanogr 2002, 47, 1844–1848. [Google Scholar]
- MacFadyen, EJ; Williamson, CE; Grad, G; Lowery, M; Jeffrey, WH; Mitchell, DL. Molecular response to climate change: Temperature dependence of UV-induced DNA damage and repair in the freshwater crustacean Daphnia pulicaria. Global Change Biol 2004, 10, 408–416. [Google Scholar]
- Lamare, MD; Barker, MF; Lesser, MP; Marshall, C. DNA photorepair in echinoid embryos: Effects of temperature on repair rate in Antarctic and non-Antarctic species. J Exp Biol 2006, 209, 5017–5028. [Google Scholar]
- Hoffman, JR; Hansen, LJ; Klinger, T. Interactions between UV radiation and temperature limit inferences from single-factor experiments. J Phycol 2003, 39, 268–272. [Google Scholar]
- Lesser, MP. Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in symbiotic dinoflagellates. Limnol Oceanogr 1996, 41, 271–283. [Google Scholar]
- Persaud, AD; Williamson, CE. Ultraviolet and temperature effects on planktonic rotifers and crustaceans in northern temperate lakes. Freshwater Biol 2005, 50, 467–476. [Google Scholar]
- Dunlap, WC; Shick, JM. Ultraviolet radiation-absorbing mycosporine-like amino acids in coral reef organisms: A biochemical and environmental perspective. J Phycol 1998, 34, 418–430. [Google Scholar]
- Banaszak, AT. Helbling, EW, Zagarese, HE, Eds.; Photoprotective physiological and biochemical responses of aquatic organisms. In UV Effects in Aquatic Organisms and Ecosystems; The Royal Society of Chemistry: Cambridge, UK, 2003; pp. 329–356. [Google Scholar]
- Spivak, ED. Los crustáceos decápodos del Atlántico sudoccidental (25°–55°S): Distribución y ciclos de vida. Invest Mar 1997, 25, 69–91. [Google Scholar]
- Arntz, WE; Gorny, M; Soto, R; Lardies, M; Retamal, M; Wehrtmann, IS. Species composition and distribution of decapod crustaceans in the waters off Patagonia and Tierra del Fuego, South America. Sci Mar 1999, 63, 303–314. [Google Scholar]
- Moreno, CA; Jara, HF. Ecological studies on fish fauna associated with Macrocystis pyrifera belts in the south of Fueguian Islands, Chile. Mar Ecol Prog Ser 1984, 15, 99–107. [Google Scholar]
- Delhey, JKV; Carrete, M; Martínez, MM. Diet and feeding behaviour of Olrog’s Gull Larus atlanticus in Bahía Blanca, Argentina. Ardea 2001, 89, 319–329. [Google Scholar]
- Boschi, EE. Las pesquerías de crustáceos decápodos en el litoral de la República Argentina. Invest Mar 1997, 25, 19–40. [Google Scholar]
- Wyngaard, JG; Iorio, MI; Boschi, EE. ¿Es viable el desarrollo de pesquerías de cangrejos en la Argentina? INIDEP 2001, 101, 9. (In Spainish) [Google Scholar]
- Morgan, SG. Young, CM, Sewell, MA, Rice, ME, Eds.; Life and death in the plankton: Larval mortality and adaptation. In Ecology of Marine Invertebrate Larvae; CSC Press: Boca Raton, FL, USA, 1995; pp. 279–321. [Google Scholar]
- Pechenik, JA. Larval experience and latent effects—metamorphosis is not a new beginning. Int Comp Biol 2006, 46, 323–333. [Google Scholar]
- Dellatorre, FG. Influencia de factores ambientales sobre la distribución y el asentamiento de larvas de cangrejos braquiuros y anomuros con potencial pesquero en Golfo Nuevo. Ph D Thesis, Universidad Nacional del Comahue, Bariloche, Argentina, 2009. [Google Scholar]
- IPCC, Climate Change 2007: The Physical Science Basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2007; p. 996.
- Damkaer, DM; Dey, DB; Heron, GA. Dose/dose rate responses of shrimp larvae to UV-B radiation. Oecologia 1981, 48, 178–182. [Google Scholar]
- Damkaer, DM; Dey, DB. UV damage and photoreactivation potentials of larval shrimp, Pandalus platyceros, and adult euphausiids, Thysanoessa raschii. Oecologia 1983, 60, 169–175. [Google Scholar]
- Hovel, KA; Morgan, SG. Susceptibility of estuarine crab larvae to ultraviolet radiation. J Exp Mar Biol Ecol 1999, 237, 107–125. [Google Scholar]
- Grad, G; Williamson, CE; Karapelou, DM. Zooplankton survival and reproduction responses to damaging UV radiation: A test of reciprocity and photoenzymatic repair. Limnol Oceanogr 2001, 46, 584–591. [Google Scholar]
- Roy, S. De Mora, SJ, Demers, S, Vernet, M, Eds.; Strategies for the minimization of UV-induced damage. In The Effects of UV Radiation in the Marine Environment; Cambridge University Press: Cambridge, UK, 2000; pp. 177–205. [Google Scholar]
- Carroll, AK; Shick, JM. Dietary accumulation of UV-absorbing mycosporine-like aminoacids (MAAs) by the gree sea urchin (Strongylocentrotus droebachiensis). Mar Biol 1996, 124, 561–569. [Google Scholar]
- Newman, SJ; Dunlap, WC; Nicol, S; Ritz, D. Antarctic krill (Euphausia superba) acquire a UV-absorbing mycosporine-like amino acid from dietary algae. J Exp Mar Biol Ecol 2000, 255, 93–110. [Google Scholar]
- Gonçalves, RJ; Villafañe, VE; Helbling, EW. Photorepair activity and protective compounds in two freshwater zooplankton species (Daphnia menucoensis and Metacyclops mendocinus) from Patagonia, Argentina. Photochem Photobiol Sci 2002, 1, 996–1000. [Google Scholar]
- Helbling, EW; Menchi, CF; Villafañe, VE. Bioaccumulation and role of UV-absorbing compounds in two marine crustacean species from Patagonia, Argentina. Photochem Photobiol Sci 2002, 1, 820–825. [Google Scholar]
- Moeller, RE; Gilroy, S; Williamson, CE; Grad, G; Sommaruga, R. Dietary acquisition of photoprotective compounds (mycosporine-like amino acids, carotenoids) and acclimation to ultraviolet radiation in a freshwater copepod. Limnol Oceanogr 2005, 50, 427–439. [Google Scholar]
- Riemer, U; Lamare, MD; Peake, BM. Temporal concentrations of sunscreen compounds (Mycosporine-like Amino Acids) in phytoplankton and in the New Zealand krill, Nyctiphanes australis G.O. Sars. J Plankton Res 2007, 29, 1077–1086. [Google Scholar]
- Persaud, AD; Moeller, RE; Williamson, CE; Burns, CW. Photoprotective compounds in weakly and strongly pigmented copepods and co-occurring cladocerans. Freshwater Biol 2007, 52, 2121–2133. [Google Scholar]
- Villafañe, VE; Marcoval, MA; Helbling, EW. Photosynthesis versus irradiance characteristics in phytoplankton assemblages off Patagonia (Argentina): Temporal variability and solar UVR effects. Mar Ecol Prog Ser 2004, 284, 23–34. [Google Scholar]
- Gao, K; Li, G; Helbling, EW; Villafañe, VE. Variability of UVR effects on photosynthesis of summer phytoplankton assemblages from a tropical coastal area of the South China Sea. Photochem Photobiol 2007, 83, 802–809. [Google Scholar]
- Helbling, EW; Villafañe, VE; Holm-Hansen, O. Weiler, CS, Penhale, PA, Eds.; Effects of ultraviolet radiation on Antarctic marine phytoplankton photosynthesis with particular attention to the influence of mixing. In Ultraviolet Radiation in Antarctica: Measurements and Biological Effects; American Geophysical Union: Washington, DC, USA, 1994; pp. 207–227. [Google Scholar]
- Villafañe, VE; Barbieri, ES; Helbling, EW. Annual patterns of ultraviolet radiation effects on temperate marine phytoplankton off Patagonia, Argentina. J Plankton Res 2004, 26, 167–174. [Google Scholar]
- Helbling, EW; Barbieri, ES; Marcoval, MA; Gonçalves, RJ; Villafañe, VE. Impact of solar ultraviolet radiation on marine phytoplankton of Patagonia, Argentina. Photochem Photobiol 2005, 81, 807–818. [Google Scholar]
- Alonso, C; Zagarese, HE; Rocco, V; Barriga, JP; Battini, MA. Surface avoidance by freshwater zooplankton: Field evidence on the role of ultraviolet radiation. Limnol Oceanogr 2004, 49, 225–232. [Google Scholar]
- Queiroga, H; Blanton, J. Interactions between behaviour and physical forcing in the control of horizontal transport of Decapod Crustacean larvae. Adv Mar Biol 2005, 47, 107–214. [Google Scholar]
- Hessen, D; De Lange, HJ; Van Donk, E. UV-induced changes in phytoplankton cells and its effects on grazers. Freshwater Biol 1997, 38, 513–524. [Google Scholar]
- Williamson, CE; Hargreaves, BR; Orr, PS; Lovera, PA. Does UV play a role in changes in predation and zooplankton community structure in acidified lakes? Limnol Oceanogr 1999, 44, 774–783. [Google Scholar]
- Fischer, JM; Fields, PA; Pryzbylkowski, PG; Nicolai, JL; Neale, PJ. Sublethal exposure to UV radiation affects respiration rates of the freshwater cladoceran Daphnia catawba. Photochem Photobiol 2006, 82, 547–550. [Google Scholar]
- Alemanni, ME; Lozada, M; Zagarese, HE. Assessing sublethal effects of ultraviolet radiation in juvenile rainbow trout (Oncorhynchus mykiss). Photochem Photobiol Sci 2003, 2, 867–870. [Google Scholar]
- Tartarotti, B; Sommaruga, R. The effect of different methanol concentrations and temperatures on the extraction of mycosporine-like amino acids (MAAs) in algae and zooplankton. Arch Hydrobiol 2002, 154, 691–703. [Google Scholar]
- Helbling, EW; Chalker, BE; Dunlap, WC; Holm-Hansen, O; Villafañe, VE. Photoacclimation of antarctic marine diatoms to solar ultraviolet radiation. J Exp Mar Biol Ecol 1996, 204, 85–101. [Google Scholar]
- Hairston, NG, Jr. The adaptive significance of color polymorphism in two species of Diaptomus (Copepoda). Limnol Oceanogr 1979, 24, 15–37. [Google Scholar]
- Helbling, EW; Pérez, DE; Medina, CD; Lagunas, MG; Villafañe, VE. Phytoplankton distribution and photosynthesis dynamics in the Chubut River estuary (Patagonia, Argentina) throughout tidal cycles. Limnol Oceanogr 2010, 55, 55–65. [Google Scholar]
- Zar, JH. Biostatistical Analysis, 4th ed; Prentice Hall: Englewood Cliffs, NJ, USA, 1999. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance to the solar simulator (m) | PAR (W m−2) | UV-A (W m−2) | UV-B (W m−2) |
|---|---|---|---|
| 0.56 | 223.5 | 89.4 | 2.19 |
| 0.85 | 125.8 | 49.1 | 1.22 |
| 1.09 | 84.5 | 30 | 0.76 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moresino, R.D.H.; Helbling, E.W. Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds. Mar. Drugs 2010, 8, 1681-1698. https://doi.org/10.3390/md8051681
Moresino RDH, Helbling EW. Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds. Marine Drugs. 2010; 8(5):1681-1698. https://doi.org/10.3390/md8051681
Chicago/Turabian StyleMoresino, Rodrigo D. Hernández, and E. Walter Helbling. 2010. "Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds" Marine Drugs 8, no. 5: 1681-1698. https://doi.org/10.3390/md8051681
APA StyleMoresino, R. D. H., & Helbling, E. W. (2010). Combined Effects of UVR and Temperature on the Survival of Crab Larvae (Zoea I) from Patagonia: The Role of UV-Absorbing Compounds. Marine Drugs, 8(5), 1681-1698. https://doi.org/10.3390/md8051681