Abstract
The growing problem of antimicrobial resistance in aquaculture, caused by the excessive and unregulated use of antibiotics, highlights the critical necessity for developing new anti-infective solutions. Based on the characteristics of glycine-rich antimicrobial peptides (AMPs) and transcriptomic data, an antimicrobial peptide, namely AfRgly1, was discovered in this study. Subsequently, the peptide was obtained through heterologous expression in E. coli, and its antibacterial spectrum was determined. Molecular dynamics simulation and molecular biology experiments were conducted to explore the antibacterial target of AfRgly1. Results showed that the mRNA expression level of AfRgly1 was significantly upregulated after Vibrio alginolyticus infection. AfRgly1 has broad-spectrum antibacterial activity targeting on bacterial cell membrane, and it may also interact with bacterial DNA. AfRgly1 displayed low selectivity for fish red blood cells. These results indicate that AfRgly1 is an antimicrobial peptide with considerable potential for application in the development of therapeutic agents.
1. Introduction
Aquaculture serves as a vital source of animal protein for human consumption [1]. However, disease outbreaks present significant challenges to achieving sustainable and eco-friendly development in this sector. The intensive farming methods employed in aquaculture often heighten the environmental stress on aquatic organisms, making them more vulnerable to pathogenic infections [2]. While antimicrobial agents are widely employed for disease prevention and treatment, their excessive application in aquaculture systems may contribute to the emergence of drug-resistant bacterial strains and the spread of resistance genes [2,3]. This phenomenon potentially endangers both aquatic species and humans. In this context, antimicrobial peptides (AMPs), the evolutionarily conserved components of innate immunity, have emerged as promising therapeutic alternatives against drug-resistant pathogens [4].
Invertebrates have evolved sophisticated defense mechanisms against microbial pathogens, primarily through the biosynthesis of diverse AMPs. Extensive research over the last twenty years has identified numerous AMP families across various invertebrate taxa, encompassing arthropods (insects, arachnids, crustaceans, and chelicerates) and mollusks [5]. These bioactive molecules exhibit remarkable structural heterogeneity, which permits their systematic classification into three principal categories based on their conformational characteristics: (1) linear polypeptides that are capable of adopting amphipathic α-helical configurations with distinct hydrophobic domains; (2) cyclic peptides that are stabilized by intramolecular disulfide bridges facilitating β-sheet or mixed α-helix/β-sheet folding patterns; and (3) amino acid-enriched peptides that contain disproportionate high concentrations of specific residues, particularly proline or glycine [6,7,8]
Glycine-rich antimicrobial peptides (AMPs), exemplified by Adepantin-1 (engineered through sequence-based AMP-Designer algorithms), Acanthoscurrin-1 (isolated from Acanthoscurria gomesiana), and Hyastatin (derived from Hyas araneus), exhibit remarkable structural plasticity owing to their distinctive glycine-enriched sequences, which typically comprise over 20% of their amino acid composition [8,9,10]. Since glycine lacks a side chain (containing only a hydrogen atom as the β-carbon substituent), its steric hindrance is significantly reduced, thereby conferring high structural flexibility and conformational diversity [11,12]. Numerous glycine-rich AMPs contain repetitive sequences of (Gly-x)n motifs, with n varying in length; these sequences commonly adopt antiparallel β-sheet conformations or exhibit flexible coiled structures [13]. Leptoglycine (isolated from Leptodactylus pentadactylus skin secretion), Holotricin-3 (found in Holotrichia diomphalia), and Serrulin (found in Tityus serrulatus) are also typical members of glycine-rich AMPs with good antibacterial activity [14,15,16].
Artemia franciscana (brine shrimp) is a small arthropod that inhabits high saline environments such as salt lakes and salt pans [17]. Arthropods solely rely on their innate immunity to resist pathogenic microorganisms, thus becoming one of the most important sources of AMPs [18]. Some confirmed AMPs, such as Cecropin, Melittin, and Crustin, have all been discovered from arthropods [19,20,21]. However, AMPs in A. franciscana have been rarely reported. Based on the characteristics of glycine-rich AMPs and transcriptome data, a novel glycine-rich antimicrobial peptide (named AfRgly1) was discovered from brine shrimp and heterologous produced. Experiments revealed that it has broad-spectrum antibacterial activity. The antibacterial target of AfRgly1 was further analyzed through molecular dynamics simulation and molecular experiments. This research will serve as a novel template for the development of AMPs, offering valuable insights into their design and application.
2. Results
2.1. Screening of AMPs with Rich Glycine
The protein profile derived from the A. franciscana genome revealed the existence of 26,923 predicted proteins [22]. With the fact that AMPs typically consist of fewer than 100 amino acids [23], 1718 predicted proteins shorter than 100 amino acids were screened out. Among them, there are 16 sequences containing more than 20% glycine (Supplementary Table S1) and with extracellular localization. After stimulation with Vibrio alginolyticus, only the gene with NCBI accession number QYM36_006760 was found to be significantly upregulated (Figure 1A and Supplementary Table S2), and it is the coding gene of AfRgly1 (NCBI access number: KAK2718072.1). The sequence alignment analysis using the BLASTP program in the CAMPR3 database (accessed on 7 August 2025) revealed that AfRgly1 is similar to Holotricin-3, with an identity of 38.09% (Figure 1B and Supplementary Table S3).
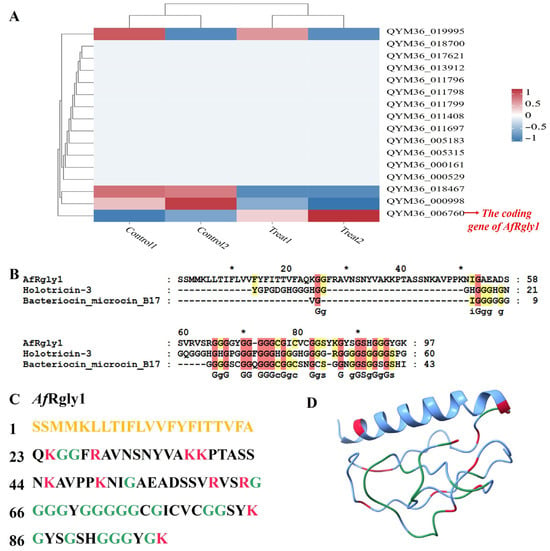
Figure 1.
The response characteristics of glycine-rich antimicrobial peptide coding genes to V. alginolyticus stress and sequence analysis of AfRgly1. (A) The response characteristics of glycine-rich antimicrobial peptide coding genes to V. alginolyticus stress and sequence analysis of AfRgly1. (B) The alignment results between AfRly1 and the two sequences most similar to it in the CAMPR3 database. (C) The amino acid sequence of AfRgly1 is presented, with signal peptide residues indicated in yellow. Cationic amino acids, including lysine and arginine, are displayed in red, while glycine residues are marked in green. (D) The predicted three-dimensional structure of AfRgly1, generated using AlphaFold3, cationic amino acids (lysine and arginine) are shown in red, and glycine residues are labeled in green. * indicates that this is the position of the 10th, 30th, 50th, 70th and 90th amino acids.
2.2. Sequence and Structure Characterization of AfRgly1
The amino acid sequence of AfRgly1 contains 97 residues (Figure 1C), with a calculated molecular weight of 9.79 kDa and a protein isoelectric point value of 9.80 predicted by APD3 [24]. The Grand Average Hydropathy value of AfRgly1 is 0.001, and the total net charge of AfRgly1 is + 9.25. The CDS encoding AfRgly1 is 291 bp in length. AfRgly has an α-helix at its N-terminal, while the rest is linear (Figure 1D). WoLF PSORT predicted its localization as extracellular.
2.3. Recombinant Expression and Purification of AfRgly1
Using pSmartI as the vector template, the mature peptide of AfRgly1 was fused to the C-terminus of the His-SUMO tag. SDS-PAGE analysis revealed marked differences in the banding patterns, with approximately 25 kDa proteins detected in the bacteria after IPTG induction (Figure 2A). This specific band corresponds to the expected size of the His-SUMO-AfRgly1 fusion protein, which consists of a His-SUMO tag (approximately 18 kDa) and AfRgly1 (9.79 kDa). The complex of fusion protein His-SUMO-AfRgly1 can be eluted from the nickel column via gradient elution with imidazole eluent at the optimal concentration of 200 μM (Figure 2B, lane 5). After treatment with the SUMO enzyme, recombinant AfRgly1 (rAfRgly1) without the His-SUMO tag (approximately 10 kDa) was obtained and confirmed by SDS-PAGE (Figure 2C).
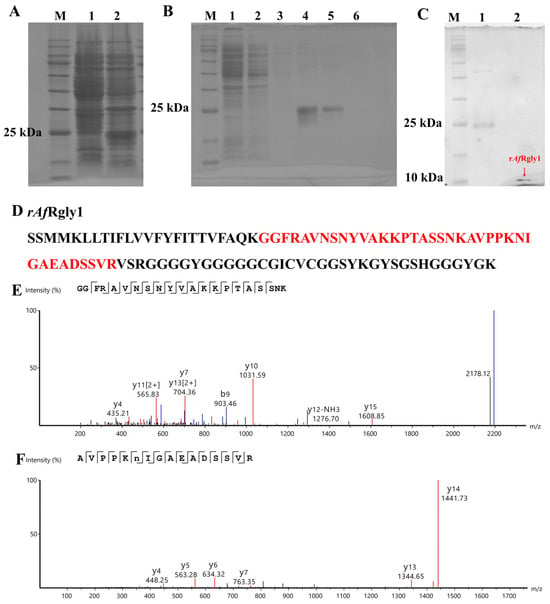
Figure 2.
Acquisition process and MS spectrum analysis of rAfRgly1. (A) SDS-PAGE analysis of recombinant AfRgly1 (rAfRgly1) fused with a His-SUMO tag in E. coli. Lane M, protein marker; lane 1, total protein obtained from E. coli without IPTG induction; lane 2, total protein obtained from E. coli with IPTG induction. (B) His-SUMO-AfRgly1 was purified via nickel column chromatography. Lane M, protein marker; lane 1, protein not caught by the nickel column; lane 2, equilibration buffer; lane 3, eluent with 50 mM imidazole; lane 4, eluent with 100 mM imidazole; lane 5, eluent with 200 mM imidazole; lane 6, eluent with 300 mM imidazole. (C) SDS–PAGE analysis of rAfRgly1 without the SUMO tag. Lane M, protein marker; lane 1, His-SUMO-AfRgly1 before treatment with the SUMO enzyme; lane 2, red arrow points to the band of tag-free rAfRgly1. (D) Alignment of mass spectrometry results with the rAfRgly1 sequence. The red area compares mass spectrometry results with the rAfRgly1 sequence. (E,F) MS spectra of “GGFRAVNSNYVAKKPTASSNK” and “AVPPKNIGAEADSSVR”, respectively.
2.4. Liquid Chromatography and Mass Spectrometry (LC-MS) Identification of AfRgly1
The amino acid sequence of rAfRgly1 was analyzed by LC-MS. As shown in Figure 2E, two peptides were detected in total, whose coverage reached 38.14%.
2.5. Antimicrobial Activity of rAfRgly1
The antibacterial activity of rAfRgly1 against three Gram-positive bacteria and four Gram-negative bacteria was determined by measuring the minimum inhibitory concentration (MIC). As shown in Table 1, rAfRgly1 exhibited significant inhibitory activity against both Gram-positive and Gram-negative bacteria. For Gram-negative bacteria, 64 μM of rAfRgly1 effectively inhibited the growth of V. alginolyticus, Aeromonas hydrophila, Vibrio anguillarum, and Escherichia coli. No growth inhibitory effect of rAfRgly1 on Acinetobacter sp. L32 and Vibrio harveyi was observed at 64 μM. The MIC values of rAfRgly1 against Staphylococcus aureus, Bacillus sp. T2 were lower, which was 32 μM. The antibacterial effect of rAfRgly1 is not as good as that of rPpRcys1 in our previous study.

Table 1.
Minimal inhibitory concentrations (MICs) and minimum bactericidal concentration (MBC)of rAfRgly1 against Gram-positive and Gram-negative bacteria.
2.6. Molecular Dynamics (MD) Simulations of AfRgly1
Before 200 ns, the root mean square distances (RMSD) value continuously increased, indicating significant conformational changes in the protein (Figure 3A,D–F). After 200 ns, the fluctuation range of RMSD gradually decreased and stabilized, suggesting that the system reached a relatively stable conformational state (Figure 3A,F,G). At 50 ns and 200 ns, the radius of gyration (Rg) value suddenly increased and then decreased, indicating that the protein underwent local unfolding and refolding at these two time points (Figure 3B). The root mean square fluctuation (RMSF) values of residues SER41-LYS50 and SER90-LYS97 were relatively high, suggesting that these two regions exhibited strong dynamic fluctuations during the membrane contact process (Figure 3C). Molecular dynamics simulation snapshots showed that the AfRgly1 protein exhibited dynamic behavior of actively approaching the membrane within 0–300 ns (Figure 3D–G). At 100 ns, two positively charged residues, LYS50 and LYS85, were the first to insert into the membrane interface (Figure 3D). At 300 ns, the number of inserted residues increased to four (LYS37, LYS45, LYS50, and LYS85), indicating that the interaction between the protein and the membrane was gradually strengthened over time (Figure 3G).
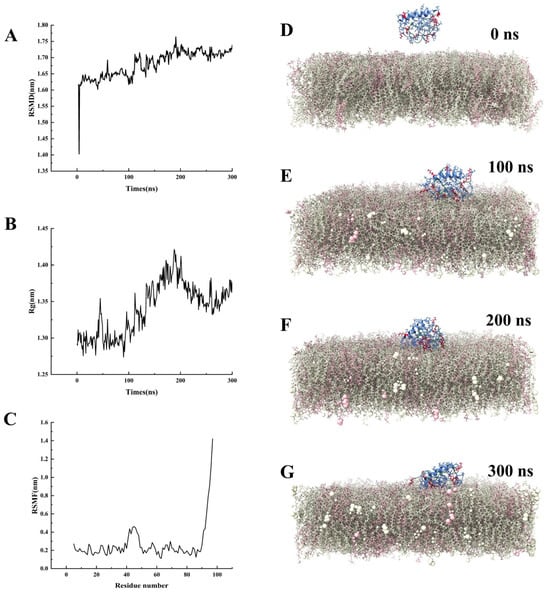
Figure 3.
Molecular dynamics simulation of AfRgly1 binding to membrane. (A) The RSMD of the AfRgly1. (B) The Rg of the AfRgly1. (C) The RSMF of the AfRgly1. (D–G) Time-dependent membrane interaction behavior of AfRgly1 observed throughout the molecular dynamic simulations (0–300 ns). Cationic amino acids (lysine and arginine) are shown in red, and glycine residues are labeled in green.
2.7. Microorganism-Binding and Membrane Mimetic-Binding Activity of rAfRgly1
The His-SUMO-AfRgly1 containing His-SUMO tag was used for the microorganism-binding assay. The negative control His-SUMO tag cannot bind to bacteria. It was found that rAfRgly1 binds to S. Aureus, Bacillus sp. T2, S. agalactiae, A. hydrophila, E. coli, and V. alginolyticus. These findings suggest that rAfRgly1 may interact with bacterial cells (Figure 4A), contributing to its antibacterial activity. The ratio of the membrane mimics is consistent with the membrane components and proportions used in molecular dynamics. rAfRgly1 can combine with the membrane mimics, which is in line with the results of molecular dynamics simulations (Figure 4B).
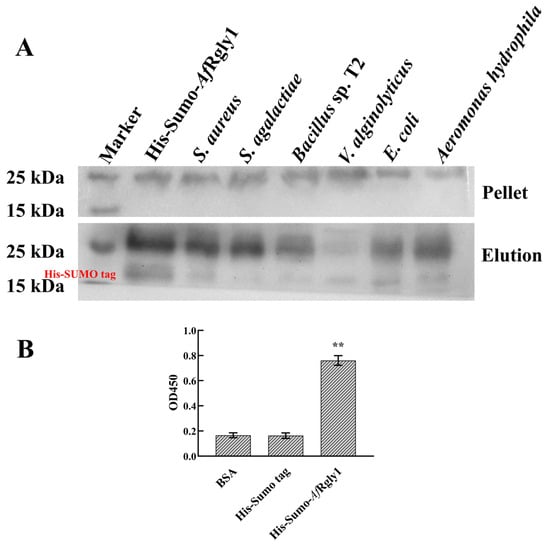
Figure 4.
Microorganism-binding activity and membrane mimetic-binding activity of rAfRgly1. (A) Microorganism-binding activity. His-SUMO-AfRgly1 was detected by Western blot analysis after treatment with bacteria. His-SUMO-AfRgly1 was used as a positive control. Upper panel, final pellet fractions; lower panel, elution fractions. (B) Membrane-mimicking binding activity assay. Bovine serum albumin (BSA) served as the control, while the His-SUMO tag acted as the negative control. The experiment was conducted with three biological repeats, each containing three technical replicates. **, significant difference compared to the control (BSA) at the level of 0.01.
2.8. Effects of rAfRgly 1 on Membrane and Bacterial Morphology
Following the disruption of microbial membrane integrity, intracellular lactate dehydrogenase is released, which serves as an indicator of bacterial cell membrane permeability. After being treated by rAfRgly1 at MICs for 2 h, the membrane permeability of S. aureus, S. agalactiae, V. alginolyticus, and E. coli was measured to be 27.11%, 22.51%, 17.47%, and 16.62%, respectively (Figure 5).
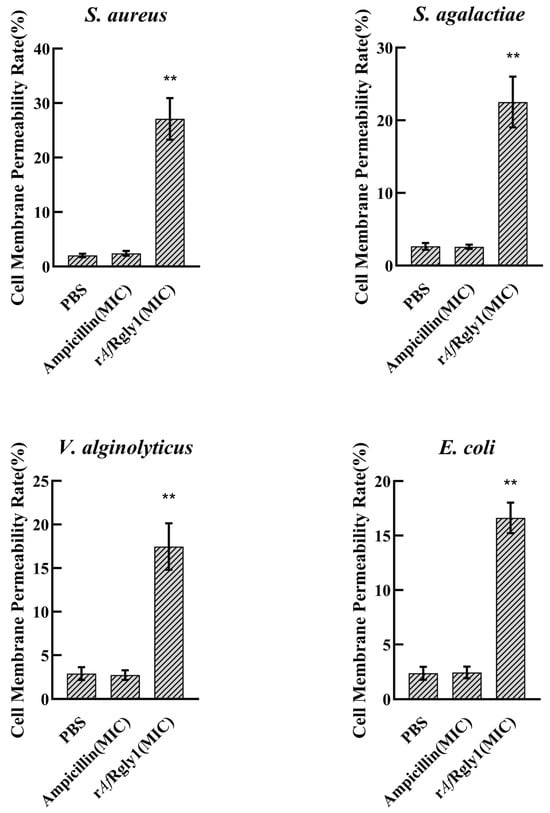
Figure 5.
Effect of rAfRgly1 on membrane permeability. Effect of rAfRgly1 on the membrane permeability rate of S. aureus, Bacillus sp. T2, V. alginolyticus and A. hydrophila, respectively. These assays included three biological replicates, each with three technical replicates. **, significant difference compared to the control (PBS) at the level of 0.01.
Propidium iodide (PI) is unable to penetrate bacteria with intact cell membranes. It can selectively enter bacteria with compromised membrane integrity and subsequently bind to intracellular DNA. PI staining demonstrated that treatment with rAfRgly1 led to substantial penetration of PI into bacterial cells in both S. aureus and V. alginolyticus (Figure 6), suggesting that rAfRgly1 can cause destabilization of bacterial cell membranes.

Figure 6.
The effect of rAfRgly1 on bacterial cell membrane integrity. About 1 × 106 CFU·mL−1 bacteria were incubated with MBC of rAfRgly1 for 2 h. PI: the cells were stained with PI and observed for PI uptake with a fluorescence microscope; BF: the bright field image. The scales are 50 μm.
Scanning electron microscope (SEM) observations revealed that S. aureus and V. alginolyticus displayed abnormal morphology features after exposure to rAfRgly1, including notable cellular shrinkage (Figure 7).
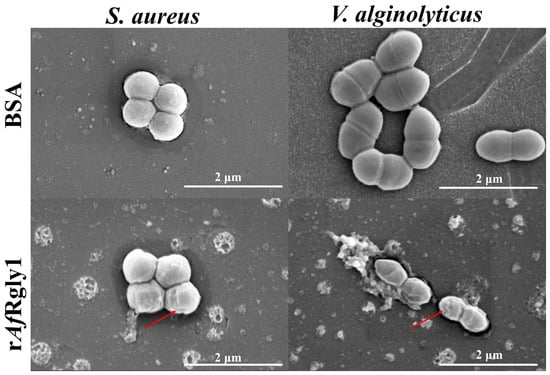
Figure 7.
Morphological alterations in bacterial cells following rAfRgly1 treatment. Bacterial samples (approximately 106 CFU·mL−1) were exposed to the MBC of rAfRgly1 for 2 h and examined using SEM. BSA served as the control group. The scale bars represent 2 μm. The red arrows denote regions where membrane contraction occurs.
2.9. DNA-Binding Activity and Hemolytic Activity of rAfRgly
The SDS-PAGE results showed that after incubation with the plasmid for 2 h, 32 μM and 64 μM of rAfRgly1 could significantly retard the migration of double-stranded DNA. At a concentration of 64 μM, the hemolysis rate of rAfRgly on fish blood cells was 10.26%(Figure 8).
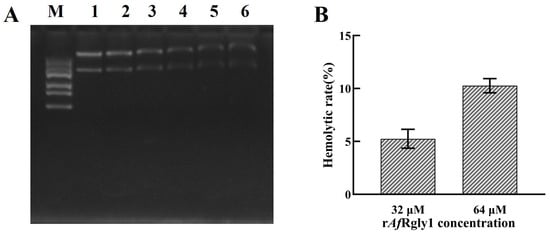
Figure 8.
DNA-binding activity and hemolytic activity of rAfRgly1. (A) Binding activity of rAfRgly1 to plasmid DNA. Lane M, marker; lane 1, 64 μM BSA; lane 2–6, 4 μM, 8 μM, 16 μM, 32 μM, and 64 μM rAfRgly1. (B) Hemolytic effect of rAfRgly1 on red blood cells of fish. The hemolysis percentage was 0% in the 1× PBS (pH 7.4) group, whereas complete lysis (100%) was observed in the 0.2% Triton X-100-treated samples. Experiments were conducted with three biological repeats, each consisting of three technical repeats.
3. Discussion
The escalating crisis of antimicrobial resistance in aquaculture due to the indiscriminate use of antibiotics seriously threatens the sustainable development of the global aquaculture industry [25,26]. AMPs have been considered as ideal candidates to replace traditional antibiotics due to their unique bactericidal mechanism and low risk of drug resistance induction [27,28]. When attacked by pathogenic microorganisms, AMPs are promptly secreted through immune regulatory mechanisms [29,30]. Consistently, the expression levels of genes encoding AMPs are significantly up-regulated [29,30]. Screening AMPs based on transcriptome data serves as a reliable and efficient strategy during the identification of novel AMPs [31]. Moreover, AMPs frequently contain signal peptides that enable their activity outside the cellular environment [32,33]. Specifically, glycine-rich AMPs are typically characterized by a glycine content exceeding 20% [14,15,34]. Based on the transcriptomics data and the above characteristics, we discovered a candidate glycine-rich AMP and named it AfRgly1.
AfRgly1 demonstrates antimicrobial properties and effectively suppresses the growth of various bacterial species. Nevertheless, its antibacterial potency was shown to be less than that of PpRcys1 based on findings from our previous research [35]. AMPs like PpRcys1, which are rich in cysteine, can form stable β-sheet structures through intramolecular disulfide bonds [36,37]. With this rigid conformation, AMPs can effectively penetrate bacterial membranes [38,39]. AfRgly1 contains three cysteine residues, which may theoretically form a disulfide bond. However, AlphaFold3 did not predict the formation of disulfide bonds in AfRgly1. To further investigate the presence of disulfide bonds, circular dichroism spectroscopy will be employed in subsequent experiments. AlphaFold3 predicts that AfRgly1 predominantly consists of a linear peptide (Figure 1D). Molecular dynamics simulations indicate that its structure is highly flexible (Figure 3), potentially contributing to its susceptibility to protease degradation [40]. This structural characteristic may partially explain why its activity is lower compared to PpRcys1. Although both AfRgly1 and PpRcys1 target bacterial cell membranes, AfRgly1 possesses the additional capability of DNA binding, a functional property absent in PpRcys1. Consequently, despite exhibiting a weaker antibacterial effect compared to PpRcys1, AfRgly1 remains a viable candidate as a template for antimicrobial peptide design.
The outer membrane of Gram-negative bacteria consists of lipopolysaccharides, phospholipids, and outer membrane proteins, of which the lipid A moiety of LPS is negatively charged and can attract cationic AMPs through electrostatic interactions [41,42]. However, the high-density lipopolysaccharides and lipoproteins of the outer membrane form a physical barrier that limits the penetration of AMPs [41,42]. In contrast to Gram-negative bacteria, Gram-positive bacteria have no outer membrane but a thick peptidoglycan layer [43,44]. Its negatively charged membrane components, such as teichoic acid, are more likely to bind directly to the cationic region of AMPs [43,44]. This may explain the better antibacterial activity of AfRgly1 against Gram-positive bacteria than against Gram-negative bacteria.
The structural and physicochemical characteristics of AfRgly1 provide important insights into its potential antimicrobial mechanisms and functional properties. With a molecular weight of 9.79 kDa and an isoelectric point of 9.80, AfRgly1 exhibits strong cationic properties (net charge +9.25) that are typical features of many membrane-active AMPs, suggesting that AfRgly1 is likely to interact with negatively charged bacterial membranes [27,33]. The near-neutral Grand Average Hydropathy (GRAVY) value of 0.001 indicates a balanced amphipathic nature, which is crucial for both membrane interaction and water solubility [45]. Therefore, molecular dynamics simulations combined with molecular experiments were conducted to assess its potential for membrane disruption.
The molecular dynamics simulation results reveal the dynamic process of AfRgly1 approaching and interacting with the membrane. RMSD and molecular dynamics simulation snapshots (Figure 3A,D–F) indicate that the AfRgly1 protein undergoes a significant conformational change before 200 ns and subsequently stabilizes to a relatively stable state, suggesting that AfRgly1 requires an initial adaptation period to establish a stable interaction with the membrane. Linear peptides riching in glycine typically exhibit high structural flexibility, as evidenced by the changes in Rg values observed at 50 ns and 200 ns during the process of AfRgly1 binding to the membrane (Figure 3B). These fluctuations suggest dynamic local unfolding and folding events. The sequential insertion pattern of positively charged lysine residues (LYS50 and LYS85 at 100 ns and extended to LYS37 and LYS45 at 300 ns, Figure 3E,G) indicates a time-dependent enhancement of protein–membrane interactions. Cationic residues usually play a key role in the binding of AMPs to bacterial membranes [46,47]. This is also confirmed by our previous molecular dynamics simulation results for PpRcys1 and PpCrus-SWD1 [35,48].
To verify the binding ability of AfRgly1 to bacterial membranes, membrane mimics identical to those used in the molecular dynamics simulations were constructed, and ELISA experiments were employed. The results demonstrate that AfRgly1 is capable of binding to these membrane mimics, which corroborates the findings from the molecular dynamics simulations. Furthermore, Western blot analysis confirmed that AfRgly1 interacts with intact bacteria, which is also corroborated by the ELISA results. These results indicate that AfRgly1 can interact with bacterial cell membranes and has the potential to disrupt them. SEM revealed that the bacterial cell surface shrank after treatment with AfRgly1. Additionally, PI staining and membrane permeability tests confirmed that AfRgly1 altered the permeability of the bacterial cytoplasmic membrane. Hence, it can be concluded that the bacterial cell membrane is one of the targets of AfRgly1.
Most AMPs carry positive charges, while the phosphate backbone of DNA is negatively charged; thus, they may non-specifically bind through electrostatic attraction. For instance, the linear isomer of melittin can wrap around DNA in a flexible conformation; similarly, the linear AMP indolicidin is disordered in solution, and its conformational flexibility may facilitate the binding to DNA [49]. Based on these findings, we hypothesized that DNA might be a potential intracellular target of AfRgly1 in bacteria. To verify it, electrophoretic mobility shift assays were conducted. Results showed that when 32 μM of AfRgly1 was co-incubated with plasmid DNA, the migration rate of the plasmid DNA in agarose gel was significantly decreased, indicating the existence of binding of plasmid DNA with AfRgly1. Therefore, bacterial DNA may be another target of AfRgly1.
In vitro hemolysis testing, as outlined by Ketan Amin and Rose-Marie Dannenfelser in their guidance for pharmaceutical scientists, indicates that formulations with a hemolysis value of less than 10% are considered non-hemolytic, whereas values exceeding 25% are deemed to pose a potential risk for hemolysis [50]. In this study, the hemolysis rate of rAfRgly1 at 64 μM was 10.26%. Therefore, we will add cell experiments in the subsequent period to evaluate its biological toxicity. The membrane activity characteristics of recombinant peptides, especially those related to cell membranes, may be directly attributed to the presence of uncleared signal peptides in their structure [51]. Therefore, the presence of signal peptides may be one of the factors leading to the hemolysis rate of AfRgly1 for FBCs exceeding 10% at 64 μM. The further research direction is to optimize AfRgly1 to reduce its hemolytic toxicity and enhance its antibacterial activity.
The N-terminal region of AfRgly1 contains a signal peptide, and its N-terminal segment is incomplete. When organisms secrete antimicrobial peptides, they usually remove their signaling peptides. But signal peptides may therefore represent a validated target for drug design [52], and thus were retained. We employed an 18 kDa His-SUMO tag and fused it with AfRgly1, starting from SER1, to facilitate protein expression. The use of a larger His-SUMO tag may provide steric protection to the potentially unstable N-terminal region, thereby preserving the native N-terminal structure and preventing possible cleavage by signal peptidases [53]. And the His-SUMO fusion system was used to produce AMPs in E. coli BL21(DE3) and can reduce its toxicity to the host [54,55]. By optimizing the induction conditions (such as reducing the IPTG concentration) [49,56], we successfully decreased the unit yield of the AMP and ultimately obtained recombinant AfRgly1 (rAfRgly1). But these operations contribute to the structural differences between rAfRgly1 and its natural counterpart, AfRgly1, which may result in variations in their antibacterial activities. In the future, we plan to construct a mutant of rAfRgly1 that lacks the signal peptide in order to obtain a structure closer to its native form. Subsequently, the antibacterial activity of this mutant will be assessed to evaluate the influence of the signal peptide on the antibacterial properties of AfRgly1.
Recombinant antimicrobial peptides may exhibit reduced antibacterial activity compared to their natural counterparts when expressed in the E. coli system, primarily due to the limited capacity of this system for post-translational modifications, such as incomplete or absent glycosylation, phosphorylation and disulfide bond formation [57]. Recombinant AfRgly1 without undergoing appropriate oxidation and renaturation processes is unlikely to form correct disulfide bonds [57]. In contrast, naturally occurring AfRgly1 is expected to possess a defined disulfide bond structure. Therefore, differences in antibacterial activity may exist between rAfRgly1 and native AfRgly1 [38,57]. To further investigate this, we plan to extract native AfRgly1 and determine its disulfide bond connectivity using techniques such as mass spectrometry or diagonal electrophoresis. Concurrently, oxidative-reductive treatments will be applied to rAfRgly1 to facilitate proper disulfide bond formation. Ultimately, the functional impact of disulfide bonds will be assessed by comparing the antibacterial activities of native AfRgly1, rAfRgly1, and refolded rAfRgly1.
The inherent limitations of LC-MS technology in the analysis of antimicrobial peptides include reduced detection sensitivity for hydrophilic or low-molecular-weight peptides, such as those containing continuous glycine residues or polar regions, which is attributed to poor ionization efficiency and weak chromatographic retention [58]. Furthermore, the application of trypsin in this study to cleave lysine or arginine residues may hinder the precise identification of certain peptides [59]. It is likely that these factors contributed to the LC-MS identification results covering only 38.14% of the complete peptide sequence of AfRgly1.
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
The experimental bacteria included three Gram-positive strains—S. aureus (ATCC 6538), S. agalactiae (ATCC 51487), and Bacillus sp. T2—as well as six Gram-negative strains—Aeromonas hydrophila (ATCC 35654), V. alginolyticus (ATCC 17749), V. anguillarum (ATCC 14181), E. coli (ATCC 8739), V. harveyi (ATCC 43516), and Acinetobacter sp. L32, which were kindly provided by Professor Chaogang Wang and Xiaohui Cai [60,61]. Bacteria were initially preserved as glycerol stocks and then cryopreserved at –80 °C for long-term storage [62]. Prior to experimentation, the bacterial stock (50 μL) was thawed in 2 mL of growth medium and incubated at 37 °C with shaking at 200 rpm for 12 h to ensure their revival and proper activation. Specifically, S. aureus, Bacillus sp. T2, Acinetobacter sp. L32, E. coli, and A. hydrophila were cultured in Luria–Bertani (LB) medium (ST163, Beyotime, Shanghai, China), while V. harveyi, V. alginolyticus, and V. anguillarum were propagated in Zobell Marine Agar 2216E (HB0132, Haibo, Qingdao, China).
4.2. Quantitative Analysis of AfRglys Expression
Sixty A. franciscana, originally collected from the Penglai Mozhikou Marine Aquaculture Facility in Penglai, China, were acclimatized in sterile seawater supplemented with ampicillin (100 μg/mL) for 48 h. This study employed the double parallel experimental design, dividing A. franciscana into a pathogen-stressed treatment group (n = 2) and a PBS-treated control group (n = 2). Each group consisted of 15 individuals of A. franciscana, which were treated with 106 CFU/mL pathogen suspension and an equal volume of sterile PBS, respectively. After 6 h, four groups of A. franciscana were immediately frozen in liquid nitrogen. Total RNA was extracted from these four samples simultaneously using the Trizol method (KL-0016, Shanghai Kanglang Biology Technology Co., Ltd., Shanghai, China). After digestion with DNase I, the RNA quality was strictly checked, with the requirement that the OD260/280 ratio detected by Nanodrop was between 1.8 and 2.0 and the RIN value detected by Agilent 2100 was ≥7.0. Consequently, the Hieff NGS® ds-cDNA Synthesis Kit (13488ES08, Yease, Shanghai, China) was used to construct cDNA libraries for these four samples simultaneously, and PE150 sequencing was performed on the NovaSeq 6000 platform. During the data analysis, Hisat2 (T2.2.1)was used for sequence alignment [63] and StringTie was used for transcript assembly [64]. The coefficient of gene expression between the two groups of parallel samples was required to be >0.9. Differential expression analysis was conducted using the edge R package (3.3.5)(p value < 0.05) [65].
4.3. Prediction and Identification of Glycine-Rich AMP
The genomic data of A. franciscana was obtained from the NCBI database (accession: GCA_032884065.1). Using a screening pipeline, we selected sequences meeting the following criteria: length < 100 amino acids, glycine content > 20%, subcellular localization outside the cell, and having a signal peptide. In addition, the expression level should be significantly upregulated after stimulation by pathogenic microorganisms. Signal peptide prediction was performed using SignalP 6.0 [66], whereas WoLF PSORT (https://wolfpsort.hgc.jp/, accessed on 14 August 2025) was used to determine subcellular localization [67]. Structural modeling of the homologous protein AfRgly1 was performed using AlphaFold3 [68], while physicochemical properties were predicted via APD3 (http://aps.unmc.edu/AP/ (accessed on 1 January 2025)). A search for AMPs with sequence similarity to AfRgly1 was conducted using the CAMPR3 database [69].
4.4. Expression and Purification of Recombinant AfRgly1
The codon-optimized AfRgly1 (NCBI: KAK2718072.1) according to the E. coli preference was fused to the His-SUMO tag and expressed in E. coli BL21(DE3). The synthetic 348 bp gene (with additional BamHI/XhoI sites) was cloned into the pSmartI vector (5850 bp; Supplementary Figure S1) and transformed into competent cells. Positive clones (kanamycin-resistant) were verified by PCR/sequencing using general primers of pSmartI (EF:5′-TTA AGA TTC TTG TAC GAC GG-3′, ER: 5′-TGC TAG TTA TTG CTC AGC GG-3′).
Established positive clones were then inoculated into LB medium supplemented with kanamycin (100 μg/mL) and incubated overnight at 37 °C with shaking at 200 rpm. On the following day, the overnight bacterial culture was then inoculated into fresh LB medium using a 1:100 dilution ratio. The suspension was subsequently incubated at 37 °C with continuous shaking at 200 rpm until reaching mid-log phase (OD600 ≈ 0.6). Protein expression was induced by the addition of isopropyl-β-d-thiogalactopyranoside (IPTG), which was adjusted to a final concentration of 0.2 mM, followed by a 12 h induction period at 37 °C.
Bacterial cell collection and lysis were conducted as previously described [48]. The cells were disrupted using the TieChui E. coli lysis buffer, followed by centrifugation at 4 °C for 30 min at a speed of 10,000 rpm to isolate the supernatant from the cellular debris. Crude proteins were extracted from uninduced and induced cells and analyzed via 12% SDS-PAGE. The recombinant His-SUMO-AfRgly1 peptide was then purified from the clarified cell lysate using nickel-affinity column chromatography. Post-purification, the target protein was subjected to buffer exchange dialysis against 1× PBS for 24 h at 4 °C. The His-SUMO tag was then completely cleaved with an additional 1 U of SUMO protease (General Biosystems, Chuzhou, China) and incubated overnight at 4 °C. The reaction mixture was then passed through the column again, with the cleaved His-SUMO tags being bound to the Ni-NTA resin, allowing tag-free rAfRgly1 to be collected in the flow-through. Purified protein samples were then subjected to SDS-PAGE analysis for quality assessment, with protein quantification performed using a BCA-based assay kit (Beyotime) according to standard protocols. Obtained protein solutions were divided into aliquots, freeze-dried, and preserved at −80 °C in powdered form. To confirm the identity, integrity, and purity of the obtained recombinant protein, rAfRgly1 samples were analyzed by Wininnovate Bio Company (Shenzhen, China) using liquid chromatography–mass spectrometry (LC-MS) analysis.
4.5. Antimicrobial Activity Determination
The antimicrobial efficacy of recombinant AfRgly1 (rAfRgly1) was assessed employing a microdilution technique based on the CLSI-recommended protocol. Bacterial suspensions with an OD600 of 0.4 were standardized to a concentration of 104 CFU/mL in Mueller–Hinton broth [37]. Two-fold serial dilutions of rAfRgly1, ranging from 64 to 1 μM in PBS, were prepared in 96-well microtiter plates, with each well containing 20 μL of peptide solution and 80 μL of bacterial inoculum. Phosphate-buffered saline (PBS) was used as the negative control. Following an 18 h incubation at 37 °C, microbial growth was monitored by measuring optical density at 560 and 590 nm (OD560 and OD590). The minimum inhibitory concentration (MIC) was defined as the lowest peptide concentration that prevented visible color change, as detected by the resazurin-based viability indicator.
To further investigate the growth inhibition kinetics, bacterial growth curves were generated by measuring OD600 at time intervals of 0, 4, 8, 12, and 24 h under MIC and half-MIC (0.5× MIC) conditions. Bovine serum albumin (BSA) was used as the control protein. The minimum bactericidal concentration (MBC) was determined through a colony-counting assay. After 18 h of incubation, 10 μL aliquots from wells showing no visible growth (sample OD600 ≤ control OD600) were transferred onto Mueller–Hinton agar (MHA, HB0128, Haibo, Qingdao, China) plates and incubated at 37 °C for 24 h. The MBC was identified as the lowest peptide concentration resulting in ≥99.9% bacterial reduction, confirmed by the absence of colony formation. Ampicillin and recombinant PpRcys1 (rPpRcys1) were used as positive controls, while 1× PBS served as the negative control.
All experiments were performed in triplicate to ensure reproducibility and data consistency [35].
4.6. Molecular Dynamics (MD) Simulations Analysis
To investigate the interaction between AfRgly1 and bacterial membranes, a series of molecular dynamics (MD) simulations was conducted, emphasizing the significance of peptide-membrane binding in computational models. The CHARMM36 force field was employed to describe the structural and dynamic properties of AfRgly1 [70]. The membrane system was constructed using 324 POPE lipids and 162 POPG lipids per leaflet, based on previously established membrane compositions [71,72]. Solvation was achieved using the TIP3P water model [73]. A cubic simulation box with dimensions of 12 × 12 × 12 nm3 was set up for systems. To ensure electrostatic neutrality, appropriate numbers of Na+ and Cl− ions were introduced into the system. All molecular dynamics simulations were performed using GROMACS version 2023.3 [74]. VMD 1.9.3 [75] was used for trajectory visualization.
Non-bonded interactions were calculated with a cutoff distance of 1.2 nm for both the Lennard–Jones and short-range electrostatic interactions. For long-range electrostatic interactions, the particle mesh Ewald (PME) method was applied, with a grid spacing of 0.16 nm and a fourth-order spline interpolation [76,77]. Temperature regulation was achieved using the V-rescale thermostat at 310 K, which is above the phase transition temperature of POPG, with a coupling time constant of 1.0 ps [78]. Pressure was maintained semi-isotropically at 1 bar using the C-rescale method, with a time constant of 5 ps and a compressibility of 4.5 × 10−5 (kJ·mol−1·nm−3)−1 [79]. Bond constraints were applied using the LINCS algorithm [80], allowing for a stable integration time step of 2 fs. Prior to production runs, the system underwent energy minimization, followed by a 500 ps equilibration phase under both NVT (constant number of particles, volume, and temperature) and NPT (constant number of particles, pressure, and temperature) conditions. Finally, a 500 ns production simulation was conducted to analyze the behavior of AfRgly1.
4.7. Electron Microscopy
Electron microscopy analysis was performed as established protocols with modifications [81]. Mid-log phase cultures of S. aureus and V. alginolyticus were collected and standardized to 106 CFU/mL using PBS. Bacterial suspensions were treated with rAfRgly1 at MBC level on coverslips (24-well plate, 2 h incubation), with BSA-treated samples as controls. Post-treatment specimens underwent sequential processing: primary fixation in 5% glutaraldehyde (4 °C, 10 h), PBS rinses (3×), gradient ethanol dehydration (30–100%, 10 min/step at 4 °C), critical point drying (Hitachi-HCP system, Hitachi, Tokyo, Japan), and gold sputter-coating (Hitachi MC1000, Hitachi, Tokyo, Japan). Imaging was performed using a Thermo Fisher APREO S SEM(Thermo Fisher, Waltham, MA, USA).
4.8. Microorganism-Binding Assay
Western blot analysis was employed to evaluate the rAfRgly1-bacterial binding ability. Briefly, bacterial suspensions at a concentration of 1 × 108 CFU/mL were incubated with 200 μL of a mixed solution containing His-SUMO-AfRgly1 and His-SUMO tag, where each component was present at a concentration of 10 μM, under rotary agitation at room temperature for 1 h. Washed thrice with TBS, the microbial pellets and supernatants were separated by centrifugation (10,000 rpm for 5 min) and then subjected to SDS-PAGE analysis. His-SUMO-AfRgly1 and His-SUMO tag served as positive/negative controls, respectively. Polyvinylidene fluoride (PVDF) membranes were blocked with 5% skim milk/TBST and then probed with HRP-anti-His antibody (Boyi Biotech, Boyi, China, 1:30,000). Protein bands were visualized using BeyoECL Plus chemiluminescent substrate (Beyotime, Shanghai, China) and chemiluminescent detection system (WD-9423B/C, Liuyi, Beijing, China) following standard protocols, and triplicate biological replicates were performed.
4.9. Binding Assay for Membrane Mimetic
To determine the optimal lipid composition, a molar ratio of POPE to POPG (7.5 nmol: 2.5 nmol) was selected within a total lipid concentration of 100 μM. The lipid mixture was initially dissolved in chloroform, followed by solvent evaporation under a nitrogen stream and further drying under high vacuum for 1 h to ensure complete removal of residual organic solvent. Liposome preparation was initiated by rehydrating the dried lipid film with preheated HEPES buffer (20 mM HEPES, 150 mM NaCl, pH 7.4). The resulting suspension underwent vortex mixing and sonication—either through pulsed probe sonication on ice (10 s bursts for 10 cycles) or bath sonication at 55 °C for 30 min—to generate small unilamellar vesicles.
The prepared liposomes were diluted to a working concentration of 10–20 μg/mL and added to a 96-well microplate at 100 μL per well. The plate was incubated overnight at 4 °C to allow liposome adsorption onto the well surface. Following this, each well was rinsed three times with 1× phosphate-buffered saline containing Tween 20 (PBST, pH 7.4). To block nonspecific binding sites, 100 μL of 5% skim milk in 1× PBST (pH 7.4) was added to each well, and the plate was incubated at 37 °C for 2 h. Afterward, the wells were gently washed three times with 1× PBST (pH 7.4) to remove excess blocking agent.
His-SUMO-AfRgly1 was prepared at a concentration of 10 μM in 1× PBS (pH 7.4) and added to the respective wells. For comparative analysis, 10 μM bovine serum albumin (BSA) served as the positive control, while 10 μM His-SUMO tag alone was used as the negative control. The plate was incubated at 37 °C for 1 h, followed by a single wash with 1× PBST (pH 7.4). Subsequently, 100 μL of horseradish peroxidase (HRP)-conjugated anti-His antibody, diluted 1:5000 in 1× PBST (pH 7.4) (Boyi, Changzhou, China), was added to each well. The plate was then incubated at 37 °C for another hour, followed by five washes with 1× PBST (pH 7.4) to remove unbound antibody.
To detect binding, 100 μL of TMB substrate solution was added to each well to initiate color development. The reaction was terminated immediately by adding 200 μL of ELISA stop solution per well. Absorbance at 450 nm was recorded using a microplate reader (SynergyTM LX, BioTek, Kaysville, UT, USA). To ensure experimental reliability, the assay was performed with three biological replicates and three technical replicates.
4.10. Lactate Dehydrogenase (LDH) Release Assay
The membrane-disruption activity of AfRgly1 against S. aureus, S. agalactiae, V. alginolyticus, and E. coli was assessed through an LDH release assay. In brief, bacterial cultures in the mid-logarithmic growth phase (OD600 ≈ 0.5) were harvested, rinsed, and resuspended in phosphate-buffered saline (PBS). Aliquots of 100 μL of the bacterial suspension were then exposed to AfRgly1 at its minimum inhibitory concentration (MIC) in a 96-well plate format for 2 h. Following centrifugation at 12,000× g for 2 min, 50 μL of the supernatant was combined with 50 μL of reaction solution containing 50 mM sodium phosphate buffer (pH 7.5), 0.6 mM pyruvate, and 0.2 mM NADH, and the mixture was allowed to react at ambient temperature for 10 min. The reaction was terminated by adding 50 μL of 1 M acetic acid, and the absorbance was recorded at 340 nm. To determine total LDH content, bacterial cells lysed with 1% Triton X-100 were used as a reference [82,83]. The percentage of LDH released was subsequently calculated as follows:
with statistical significance assessed by one-way ANOVA (triplicate, p < 0.05). Controls included bacteria treated by BSA (negative) and 1% Triton X-100 (positive).
4.11. PI Staining
S. aureus and V. alginolyticus were cultured as described above at their MBC for 2 h. Samples were stained with a PI staining kit (Sangon, Shanghai, China) according to the manufacturer’s instructions. Cells were observed under a fluorescence microscope (BX51, Olympus, Tokyo, Japan).
4.12. DNA-Binding Assay
The V. parahaemolyticus PirA virulence gene (GenBank: MH410659.1) was synthesized with additional BamHI/XhoI flanking sites and cloned into the pSmartI vector to construct pSmart-PirA (Supplementary Figure S2). DNA–protein binding reactions (20 μL) containing 400 ng plasmid DNA and serially diluted rAfRgly1 (16–64 μM) in Tris-based buffer (10 mM of Tris-HCl, pH of 8.0, 1 mM of EDTA, 20 mM of KCl, 1 mM of DTT, 5% glycerol, 50 μg/mL of BSA) were incubated at 37 °C for 1 h. BSA at 64 μM was used as the negative control. Samples mixed with loading buffer were electrophoresed on 1% agarose gel at 120 V for 20 min. The DNA retardation patterns were used to indicate binding activity.
4.13. Hemolytic Activity Assay
The hemolytic potential of recombinant proteins was evaluated using carp fish erythrocytes. Fish red blood cells (FRBCs, HQ80080, Hongqian Bio, Guangzhou, China) were washed with sterile saline (0.85%) and resuspended to a final 4% (v/v) suspension. Aliquots (150 μL) were incubated with equal volumes of test protein (16–64 μM), PBS (the negative control), or 0.2% Triton X-100 (the positive control) at 37 °C for 1 h. After centrifugation, supernatant absorbance at 570 nm (OD570) was measured. Hemolysis percentage was calculated as
Triplicate biological and technical replicates were performed to ensure data reliability.
4.14. Statistical Analysis
Statistical analysis was conducted using GraphPad Prism 10.0 (GraphPad, San Diego, CA, USA). Significance was assessed with a one-way analysis of variance (ANOVA), and all data were reported as mean ± SD from three biological replicates. p-value < 0.05 was deemed to be statistically significant.
5. Conclusions
In this study, a novel AMP, AfRgly1, was successfully identified and hetero-expressed, according to the features of glycine-rich AMPs and transcriptomic analysis of brine shrimp. AfRgly1 exhibits broad-spectrum antibacterial activity and is capable of exerting its effects by compromising the integrity of bacterial cell membranes. Furthermore, it may also interact with bacterial DNA. Within the effective antibacterial concentration range, AfRgly1 exhibits low selectivity for fish red blood cells. These findings indicate that AfRgly1 has substantial potential for pharmaceutical development.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/md23080330/s1, Figure S1: pSmartI-AfRgly1 Plasmid map; Figure S2: pSmart-PirA Plasmid map; Figure S3: original Western blot; Table S1: screening of rich glycine peptides from Artemia franciscana genome and transcriptome; Table S2: results of transcriptome gene differential expression analysis. Table S3: the comparison result of AfRgly1 in the CAMPR3 database.
Author Contributions
Conceptualization, W.Z. and D.C.; methodology, M.T.; validation, H.Y., G.Y. and A.S.; formal analysis, H.S.; investigation, H.Y.; data curation, M.T.; writing—original draft preparation, M.T.; writing—review and editing, W.Z.; visualization, H.S.; supervision, D.C.; project administration, W.Z.; funding acquisition, M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32303034) and the Marine Science Young Scientists Cultivation Program of the Guangxi Beibu Gulf Marine Science and Technology Innovation Cooperation Base (24-03190010).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article, and further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors would like to express their gratitude to the Biosciences Central Research Facility at Shenzhen University for providing technical support, especially in the LC-MS experiments. Additionally, we are thankful to the Instrument Analysis Center at Shenzhen University for their help with the SEM analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
Correction Statement
This article has been republished with a minor correction to the existing affiliation information. This change does not affect the scientific content of the article.
Abbreviations
The following abbreviations are used in this manuscript:
| AMP | antimicrobial peptide |
| Rg | gyration |
| RMSD | root mean square distances |
| RMSF | root mean square fluctuations |
| POPE | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine |
| POPG | phosphatidylglycerol |
References
- Boyd, C.E.; McNevin, A.A.; Davis, R.P. The Contribution of Fisheries and Aquaculture to the Global Protein Supply. Food Secur. 2022, 14, 805–827. [Google Scholar] [CrossRef]
- Chen, X.; Liu, H.; Liu, S.; Mao, J. Impact of Bacteriocins on Multidrug-Resistant Bacteria and Their Application in Aquaculture Disease Prevention and Control. Rev. Aquac. 2024, 16, 1286–1307. [Google Scholar] [CrossRef]
- Dayie, N.T.K.D.; Nathan-Mensah, F.N.N.; Kotey, F.C.N.; Tabi, B.K.A.; Kabotso, D.E.K.; Odoom, A.; Hotor, P.; Dayie, A.D.; Tetteh-Quarcoo, P.B.; Egyir, B. Multidrug-Resistant Bacteria in Aquaculture Systems in Accra, Ghana. Environ. Health Insights 2024, 18, 11786302241299368. [Google Scholar] [CrossRef]
- Chen, P.; Ye, T.; Li, C.; Praveen, P.; Hu, Z.; Li, W.; Shang, C. Embracing the Era of Antimicrobial Peptides with Marine Organisms. Nat. Prod. Rep. 2024, 41, 331–346. [Google Scholar] [CrossRef]
- Bulet, P.; Charlet, M.; Hetru, C. Antimicrobial Peptides in Insect Immunity. In Innate Immunity; Springer: Berlin/Heidelberg, Germany, 2003; pp. 89–107. [Google Scholar]
- Koehbach, J.; Craik, D.J. The Vast Structural Diversity of Antimicrobial Peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Li, W.; Tailhades, J.; O’Brien-Simpson, N.M.; Separovic, F.; Otvos, L.; Hossain, M.A.; Wade, J.D. Proline-Rich Antimicrobial Peptides: Potential Therapeutics against Antibiotic-Resistant Bacteria. Amino Acids 2014, 46, 2287–2294. [Google Scholar] [CrossRef]
- Lorenzini, D.M.; da Silva, P.I.; Fogaça, A.C.; Bulet, P.; Daffre, S. Acanthoscurrin: A Novel Glycine-Rich Antimicrobial Peptide Constitutively Expressed in the Hemocytes of the Spider Acanthoscurria Gomesiana. Dev. Comp. Immunol. 2003, 27, 781–791. [Google Scholar] [CrossRef]
- Ilić, N.; Novković, M.; Guida, F.; Xhindoli, D.; Benincasa, M.; Tossi, A.; Juretić, D. Selective Antimicrobial Activity and Mode of Action of Adepantins, Glycine-Rich Peptide Antibiotics Based on Anuran Antimicrobial Peptide Sequences. Biochim. Et Biophys. Acta (BBA) Biomembr. 2013, 1828, 1004–1012. [Google Scholar] [CrossRef]
- Sperstad, S.V.; Haug, T.; Vasskog, T.; Stensvåg, K.; Hyastatin, A. Glycine-Rich Multi-Domain Antimicrobial Peptide Isolated from the Spider Crab (Hyas Araneus) Hemocytes. Mol. Immunol. 2009, 46, 2604–2612. [Google Scholar] [CrossRef]
- Öten, A.M.; Atak, E.; Taktak Karaca, B.; Fırtına, S.; Kutlu, A. Discussing the Roles of Proline and Glycine from the Perspective of Cold Adaptation in Lipases and Cellulases. Biocatal. Biotransformation 2023, 41, 243–260. [Google Scholar] [CrossRef]
- Rončević, T.; Vukičević, D.; Ilić, N.; Krce, L.; Gajski, G.; Tonkić, M.; Goić-Barišić, I.; Zoranić, L.; Sonavane, Y.; Benincasa, M.; et al. Antibacterial Activity Affected by the Conformational Flexibility in Glycine–Lysine Based α-Helical Antimicrobial Peptides. J. Med. Chem. 2018, 61, 2924–2936. [Google Scholar] [CrossRef]
- Mousavi, A.; Hotta, Y. Glycine-Rich Proteins. Appl. Biochem. Biotechnol. 2005, 120, 169–174. [Google Scholar] [CrossRef]
- Sousa, J.C.; Berto, R.F.; Gois, E.A.; Fontenele-Cardi, N.C.; Honório-Júnior, J.E.R.; Konno, K.; Richardson, M.; Rocha, M.F.G.; Camargo, A.A.C.M.; Pimenta, D.C.; et al. Leptoglycin: A New Glycine/Leucine-Rich Antimicrobial Peptide Isolated from the Skin Secretion of the South American Frog Leptodactylus Pentadactylus (Leptodactylidae). Toxicon 2009, 54, 23–32. [Google Scholar] [CrossRef]
- Nunes, L.G.P.; Reichert, T.; Machini, M.T. His-Rich Peptides, Gly- and His-Rich Peptides: Functionally Versatile Compounds with Potential Multi-Purpose Applications. Int. J. Pept. Res. Ther. 2021, 27, 2945–2963. [Google Scholar] [CrossRef]
- Lee, S.; Moon, H.; Kurata, S.; Natori, S.; Lee, B. Purification and CDNA Cloning of an Antifungal Protein from the Hemolymph of Holotrichia Diomphalia Larvae. Biol. Pharm. Bull. 1995, 18, 1049–1052. [Google Scholar] [CrossRef]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Roque, A.; Lizarraga-Partida, M.L.; Guerra-Flores, A.L. Virulence of Luminous Vibrios to Artemia Franciscana Nauplii. Dis. Aquat. Organ. 2003, 53, 231–240. [Google Scholar] [CrossRef]
- Saucedo-Vázquez, J.P.; Gushque, F.; Vispo, N.S.; Rodriguez, J.; Gudiño-Gomezjurado, M.E.; Albericio, F.; Tellkamp, M.P.; Alexis, F. Marine Arthropods as a Source of Antimicrobial Peptides. Mar. Drugs 2022, 20, 501. [Google Scholar] [CrossRef]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect Cecropins, Antimicrobial Peptides with Potential Therapeutic Applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef]
- Dempsey, C.E. The Actions of Melittin on Membranes. Biochim. Et Biophys. Acta (BBA) Rev. Biomembr. 1990, 1031, 143–161. [Google Scholar] [CrossRef]
- Smith, V.J.; Fernandes, J.M.O.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP Domain-Containing Antibacterial Proteins from Crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef]
- Bett, V.K.; Macon, A.; Vicoso, B.; Elkrewi, M. Chromosome-Level Assembly of Artemia Franciscana Sheds Light on Sex Chromosome Differentiation. Genome Biol. Evol. 2024, 16, evae006. [Google Scholar] [CrossRef]
- Seyfi, R.; Kahaki, F.A.; Ebrahimi, T.; Montazersaheb, S.; Eyvazi, S.; Babaeipour, V.; Tarhriz, V. Antimicrobial Peptides (AMPs): Roles, Functions and Mechanism of Action. Int. J. Pept. Res. Ther. 2020, 26, 1451–1463. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The Antimicrobial Peptide Database as a Tool for Research and Education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Smith, R.; Coast, J. The True Cost of Antimicrobial Resistance. BMJ Br. Med. J. 2013, 346, f1493. [Google Scholar] [CrossRef]
- Majumder, M.A.A.; Rahman, S.; Cohall, D.; Bharatha, A.; Singh, K.; Haque, M.; Gittens-St Hilaire, M. Antimicrobial Stewardship: Fighting Antimicrobial Resistance and Protecting Global Public Health. Infect. Drug Resist. 2020, 13, 4713–4738. [Google Scholar] [CrossRef]
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. Int. J. Mol. Sci. 2021, 22, 11401. [Google Scholar] [CrossRef]
- Ramzah, N.H.H.L.; Yenn, T.W.; Lee, W.-H.; Loo, C.-Y.; Tan, W.-N.; Ring, L.C. Antimicrobial Peptides, An Alternative Antimicrobial Agent Against Multi-Drug-Resistant Microbes: Source, Application, and Potential. Adv. Mater. Sci. Technol. Led Women 2023, 165, 235–259. [Google Scholar]
- Wang, D.; Li, F.; Li, S.; Wen, R.; Xiang, J. Expression Profiles of Antimicrobial Peptides (AMPs) and Their Regulation by Relish. Chin. J. Oceanol. Limnol. 2012, 30, 611–619. [Google Scholar] [CrossRef]
- Amparyup, P.; Kondo, H.; Hirono, I.; Aoki, T.; Tassanakajon, A. Molecular Cloning, Genomic Organization and Recombinant Expression of a Crustin-like Antimicrobial Peptide from Black Tiger Shrimp Penaeus Monodon. Mol. Immunol. 2008, 45, 1085–1093. [Google Scholar] [CrossRef]
- Jhong, J.-H.; Chi, Y.-H.; Li, W.-C.; Lin, T.-H.; Huang, K.-Y.; Lee, T.-Y. DbAMP: An Integrated Resource for Exploring Antimicrobial Peptides with Functional Activities and Physicochemical Properties on Transcriptome and Proteome Data. Nucleic Acids Res. 2019, 47, D285–D297. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Gallo, R.L. Antimicrobial Peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Rahman, M.S.; Choi, Y.H.; Choi, Y.S.; Yoo, J.C. Glycin-Rich Antimicrobial Peptide YD1 from B. Amyloliquefaciens, Induced Morphological Alteration in and Showed Affinity for Plasmid DNA of E. Coli. AMB Express 2017, 7, 8. [Google Scholar] [CrossRef]
- He, Z.; Fei, Z.; Shi, H.; Huang, M.; Wei, L.; Wang, J.; He, P.; Zhang, W. Heterologous Expression and Antimicrobial Mechanism of a Cysteine-Rich Peptide from Barnacle Pollicipes Pollicipes. Microorganisms 2025, 13, 1381. [Google Scholar] [CrossRef]
- Ma, H.; Feng, Y.; Cao, Q.; Jia, J.; Ali, M.; Shah, D.; Meyers, B.C.; He, H.; Zhang, Y. Evolution of Antimicrobial Cysteine-Rich Peptides in Plants. Plant Cell Rep. 2023, 42, 1517–1527. [Google Scholar] [CrossRef]
- Ho, T.N.T.; Turner, A.; Pham, S.H.; Nguyen, H.T.; Nguyen, L.T.T.; Nguyen, L.T.; Dang, T.T. Cysteine-Rich Peptides: From Bioactivity to Bioinsecticide Applications. Toxicon 2023, 230, 107173. [Google Scholar] [CrossRef]
- Holzknecht, J.; Marx, F. Navigating the Fungal Battlefield: Cysteine-Rich Antifungal Proteins and Peptides from Eurotiales. Front. Fungal Biol. 2024, 5, 1451455. [Google Scholar] [CrossRef]
- Nagy, K.; Végh, A.G.; Kereszt, A.; Kondorosi, É.; Váró, G.; Szegletes, Z. Interaction of Cysteine-Rich Cationic Antimicrobial Peptides with Intact Bacteria and Model Membranes. Gen. Physiol. Biophys. 2015, 34, 135–144. [Google Scholar] [CrossRef]
- Lai, Z.; Yuan, X.; Chen, H.; Zhu, Y.; Dong, N.; Shan, A. Strategies Employed in the Design of Antimicrobial Peptides with Enhanced Proteolytic Stability. Biotechnol. Adv. 2022, 59, 107962. [Google Scholar] [CrossRef]
- Hiroshi, N. Molecular Basis of Bacterial Outer Membrane Permeability Revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Papo, N.; Shai, Y. A Molecular Mechanism for Lipopolysaccharide Protection of Gram-Negative Bacteria from Antimicrobial Peptides*. J. Biol. Chem. 2005, 280, 10378–10387. [Google Scholar] [CrossRef]
- Chen, N.; Jiang, C. Antimicrobial Peptides: Structure, Mechanism, and Modification. Eur. J. Med. Chem. 2023, 255, 115377. [Google Scholar] [CrossRef]
- Malanovic, N.; Lohner, K. Gram-Positive Bacterial Cell Envelopes: The Impact on the Activity of Antimicrobial Peptides. Biochim. Et Biophys. Acta (BBA) Biomembr. 2016, 1858, 936–946. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Lopez Cascales, J.J.; Zenak, S.; García de La Torre, J.; Lezama, O.G.; Garro, A.; Enriz, R.D. Small Cationic Peptides: Influence of Charge on Their Antimicrobial Activity. ACS Omega 2018, 3, 5390–5398. [Google Scholar] [CrossRef]
- Glukhov, E.; Burrows, L.L.; Deber, C.M. Membrane Interactions of Designed Cationic Antimicrobial Peptides: The Two Thresholds. Biopolym. Orig. Res. Biomol. 2008, 89, 360–371. [Google Scholar] [CrossRef]
- Zhang, W.; Wei, L.; Chen, P.; Ning, B.; Wang, J.; He, P.; Shang, C.; Yu, D. Discovery and Characterization of an Atypical Crustin Antimicrobial Peptide from Pollicipes Pollicipes. Mar. Drugs 2024, 22, 526. [Google Scholar] [CrossRef]
- Hsu, C.-H.; Chen, C.; Jou, M.-L.; Lee, A.Y.-L.; Lin, Y.-C.; Yu, Y.-P.; Huang, W.-T.; Wu, S.-H. Structural and DNA-Binding Studies on the Bovine Antimicrobial Peptide, Indolicidin: Evidence for Multiple Conformations Involved in Binding to Membranes and DNA. Nucleic Acids Res. 2005, 33, 4053–4064. [Google Scholar] [CrossRef]
- Amin, K.; Dannenfelser, R.-M. In Vitro Hemolysis: Guidance for the Pharmaceutical Scientist. J. Pharm. Sci. 2006, 95, 1173–1176. [Google Scholar] [CrossRef]
- Garsin, D.A. Peptide Signals Sense and Destroy Target Cells. Science (1979) 2004, 306, 2202–2203. [Google Scholar] [CrossRef]
- Lumangtad, L.A.; Bell, T.W. The Signal Peptide as a New Target for Drug Design. Bioorganic Med. Chem. Lett. 2020, 30, 127115. [Google Scholar] [CrossRef]
- Heinrich, J.; Drewniok, C.; Neugebauer, E.; Kellner, H.; Wiegert, T. The YoaW Signal Peptide Directs Efficient Secretion of Different Heterologous Proteins Fused to a StrepII-SUMO Tag in Bacillus Subtilis. Microb. Cell Fact. 2019, 18, 31. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Wei, D.; Wang, J.; Shan, A.; Li, Z. Expression of Plectasin in Bacillus Subtilis Using SUMO Technology by a Maltose-Inducible Vector. J. Ind. Microbiol. Biotechnol. 2015, 42, 1369–1376. [Google Scholar] [CrossRef]
- Xu, Y.; Dong, M.; Wang, Q.; Sun, Y.; Hang, B.; Zhang, H.; Hu, J.; Zhang, G. Soluble Expression of Antimicrobial Peptide BSN-37 from Escherichia Coli by SUMO Fusion Technology. Protein J. 2023, 42, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Einsfeldt, K.; Severo Júnior, J.B.; Corrêa Argondizzo, A.P.; Medeiros, M.A.; Alves, T.L.M.; Almeida, R.V.; Larentis, A.L. Cloning and Expression of Protease ClpP from Streptococcus Pneumoniae in Escherichia Coli: Study of the Influence of Kanamycin and IPTG Concentration on Cell Growth, Recombinant Protein Production and Plasmid Stability. Vaccine 2011, 29, 7136–7143. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Recombinant Production of Antimicrobial Peptides in Escherichia Coli: A Review. Protein Expr. Purif. 2011, 80, 260–267. [Google Scholar] [CrossRef]
- Kang, L.; Weng, N.; Jian, W. LC–MS Bioanalysis of Intact Proteins and Peptides. Biomed. Chromatogr. 2020, 34, e4633. [Google Scholar] [CrossRef]
- Lubec, G.; Afjehi-Sadat, L. Limitations and Pitfalls in Protein Identification by Mass Spectrometry. Chem. Rev. 2007, 107, 3568–3584. [Google Scholar] [CrossRef]
- Zhuang, H.; Ou, Y.; Chen, R.; Huang, D.; Wang, C. Comparing the Ability of Secretory Signal Peptides for Heterologous Expression of Anti-Lipopolysaccharide Factor 3 in Chlamydomonas Reinhardtii. Mar. Drugs 2023, 21, 346. [Google Scholar] [CrossRef]
- Yang, S.; Cai, X. Genome-Wide Screening of the Classical Cadherin Gene Family and Cadherin-1 Expression Response Infected with Streptococcus Agalactiae in Nile Tilapia (Oreochromis Niloticus). Aquac. Rep. 2020, 17, 100393. [Google Scholar] [CrossRef]
- Yang, H.; Li, S.; Li, F.; Xiang, J. Structure and Bioactivity of a Modified Peptide Derived from the LPS-Binding Domain of an Anti-Lipopolysaccharide Factor (ALF) of Shrimp. Mar. Drugs 2016, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. Erecipes Res. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. Differential Expression Analysis of Complex RNA-Seq Experiments Using EdgeR; Springer International Publishing: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Nakai, K. Protein Subcellular localization prediction with wolf psort. In Proceedings of the 4th Asia-Pacific Bioinformatics Conference, Taipei, Taiwan, 12–16 February 2006; World Scientific: Singapore, 2006; pp. 39–48. [Google Scholar]
- Oldach, L. AlphaFold 3 to Offer Structure Prediction via Web Browser. Chem. Eng. News 2024, 102, 7. [Google Scholar]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMPR3: A Database on Sequences, Structures and Signatures of Antimicrobial Peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; MacKerell, A.D., Jr. Optimization of the Additive CHARMM All-Atom Protein Force Field Targeting Improved Sampling of the Backbone ϕ, ψ and Side-Chain Χ1 and Χ2 Dihedral Angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef]
- Murzyn, K.; Róg, T.; Pasenkiewicz-Gierula, M. Phosphatidylethanolamine-Phosphatidylglycerol Bilayer as a Model of the Inner Bacterial Membrane. Biophys. J. 2005, 88, 1091–1103. [Google Scholar] [CrossRef]
- Balatti, G.E.; Martini, M.F.; Pickholz, M. A Coarse-Grained Approach to Studying the Interactions of the Antimicrobial Peptides Aurein 1.2 and Maculatin 1.1 with POPG/POPE Lipid Mixtures. J. Mol. Model. 2018, 24, 208. [Google Scholar] [CrossRef]
- Berendsen, H.J.; Postma, J.V.; Van Gunsteren, W.F.; DiNola, A.R.H.J.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Darden, T.; York, D. An N⋅Log (N) Method for Ewald Sums in Large Systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical Sampling through Velocity Rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef]
- Bernetti, M.; Bussi, G. Pressure Control Using Stochastic Cell Rescaling. J. Chem. Phys. 2020, 153, 114107. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, X.; Zhang, J.; Ye, T.; Zhou, Q.; Xu, Y.; Li, W.; Hu, Z.; Shang, C. Discovery and Characterization of a New Crustin Antimicrobial Peptide from Amphibalanus Amphitrite. Pharmaceutics 2022, 14, 413. [Google Scholar] [CrossRef]
- Cornett, J.B.; Shockman, G.D. Cellular Lysis of Streptococcus Faecalis Induced with Triton X-100. J. Bacteriol. 1978, 135, 153–160. [Google Scholar] [CrossRef]
- Sung, K.; Khan, S.A.; Nawaz, M.S.; Khan, A.A. A Simple and Efficient Triton X-100 Boiling and Chloroform Extraction Method of RNA Isolation from Gram-Positive and Gram-Negative Bacteria. FEMS Microbiol. Lett. 2003, 229, 97–101. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).