
Dihydrogeodin from Fennellia flavipes Modulates Platelet Aggregation via Downregulation of Calcium Signaling, αIIbβ3 Integrins, MAPK, and PI3K/Akt Pathways

 , , , ,
, , , , 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Structural Elucidation
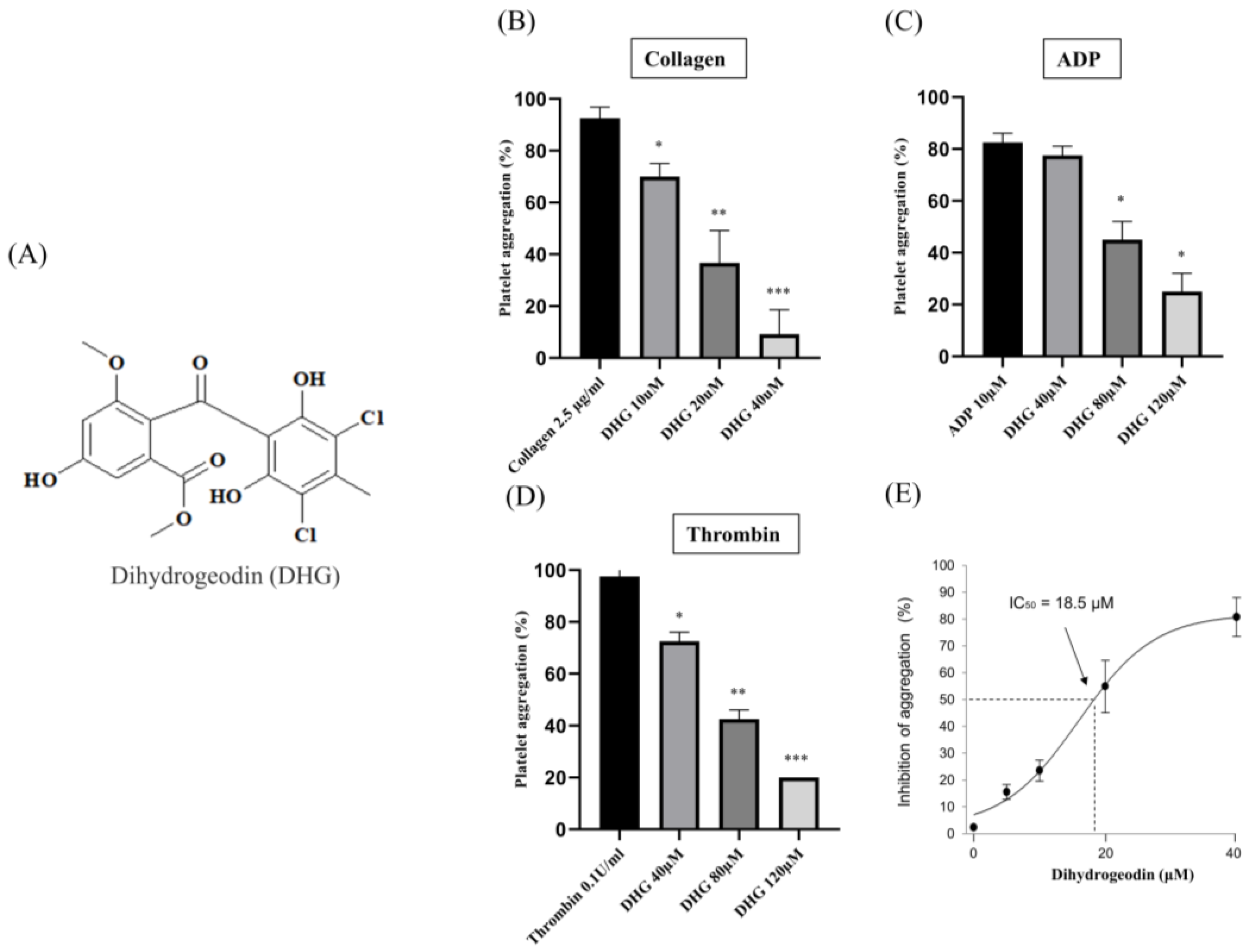
2.2. DHG Inhibits Agonist-Induced Platelet Aggregation
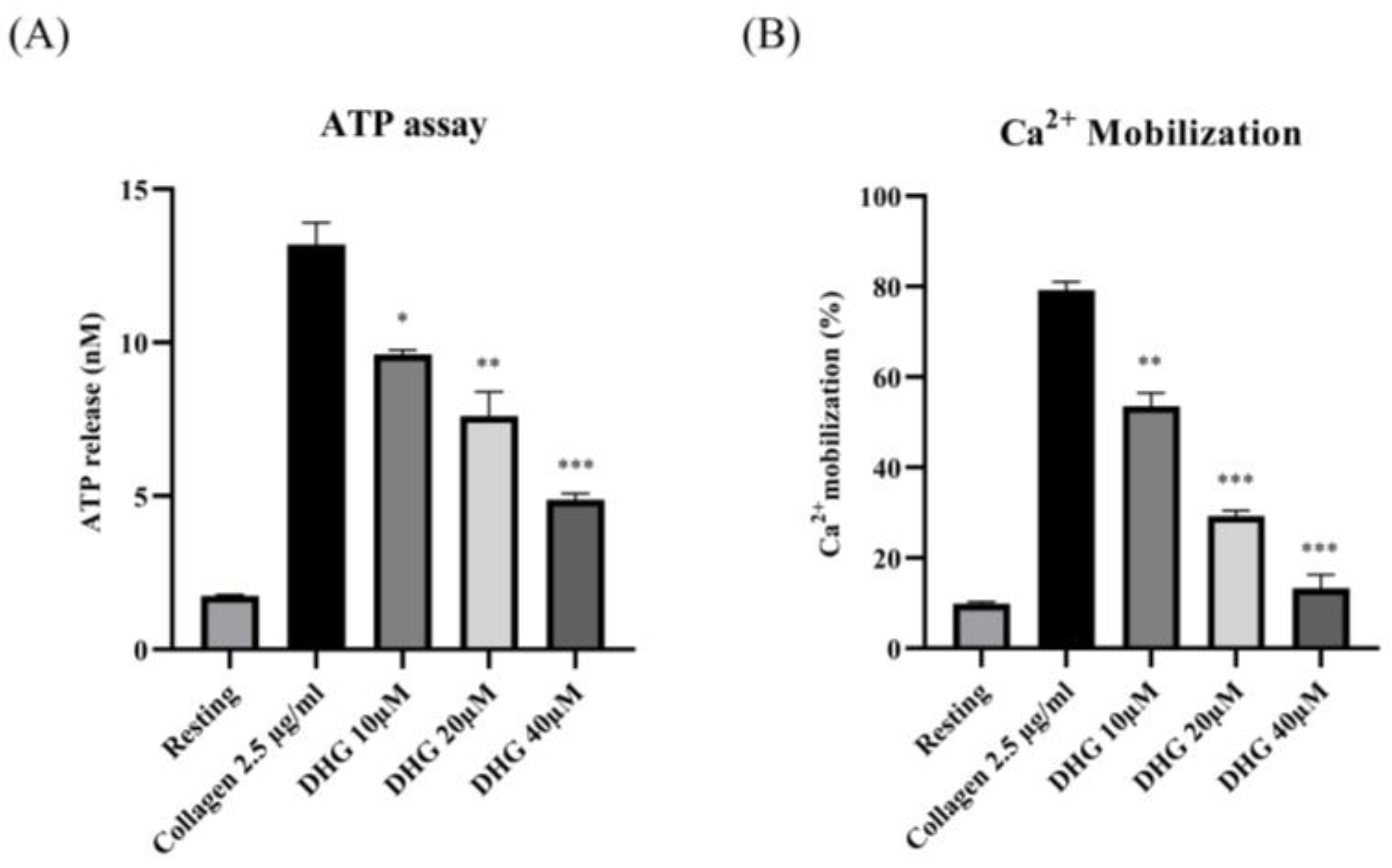
2.3. DHG Inhibits ATP Release and [Ca2+]i Mobilization
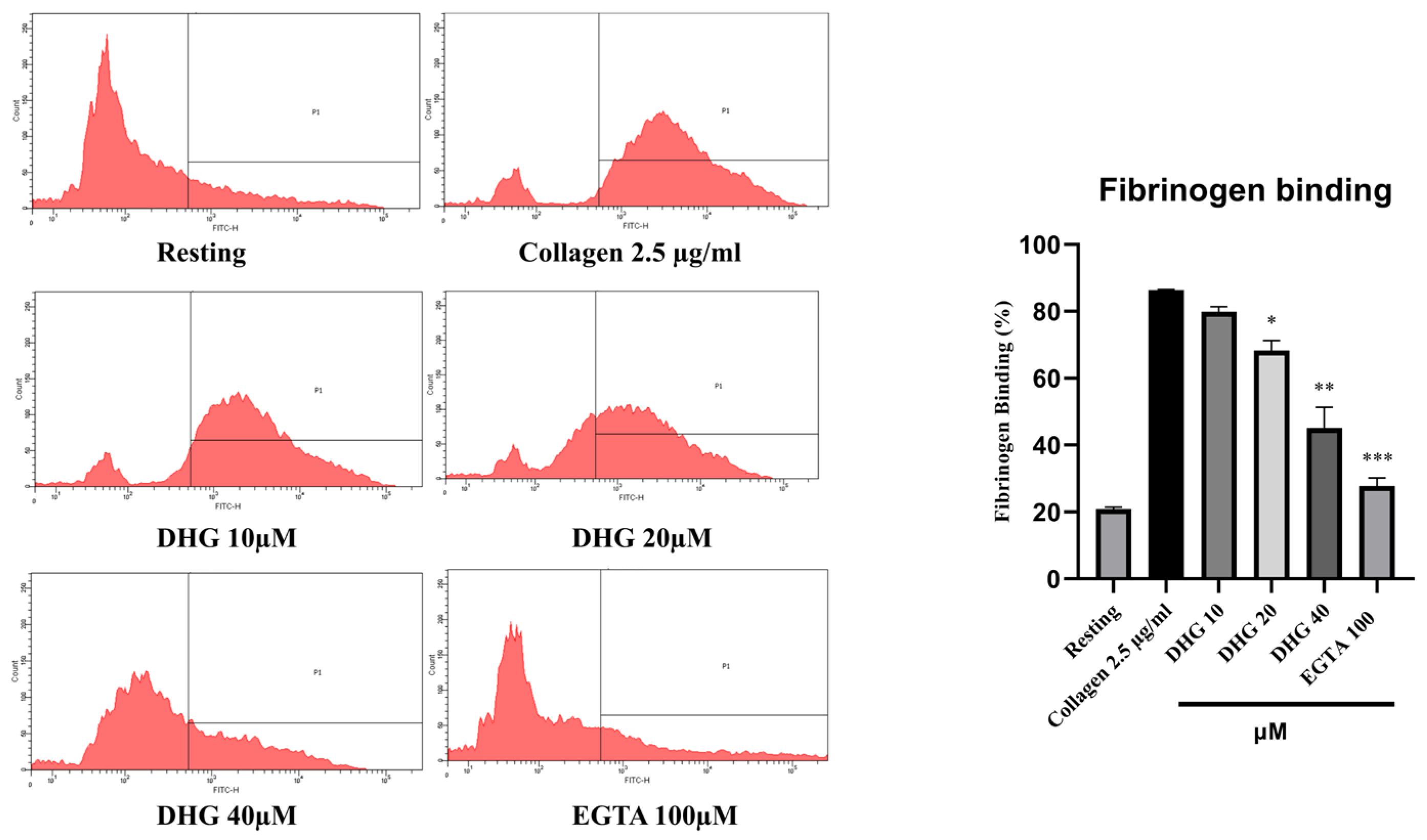
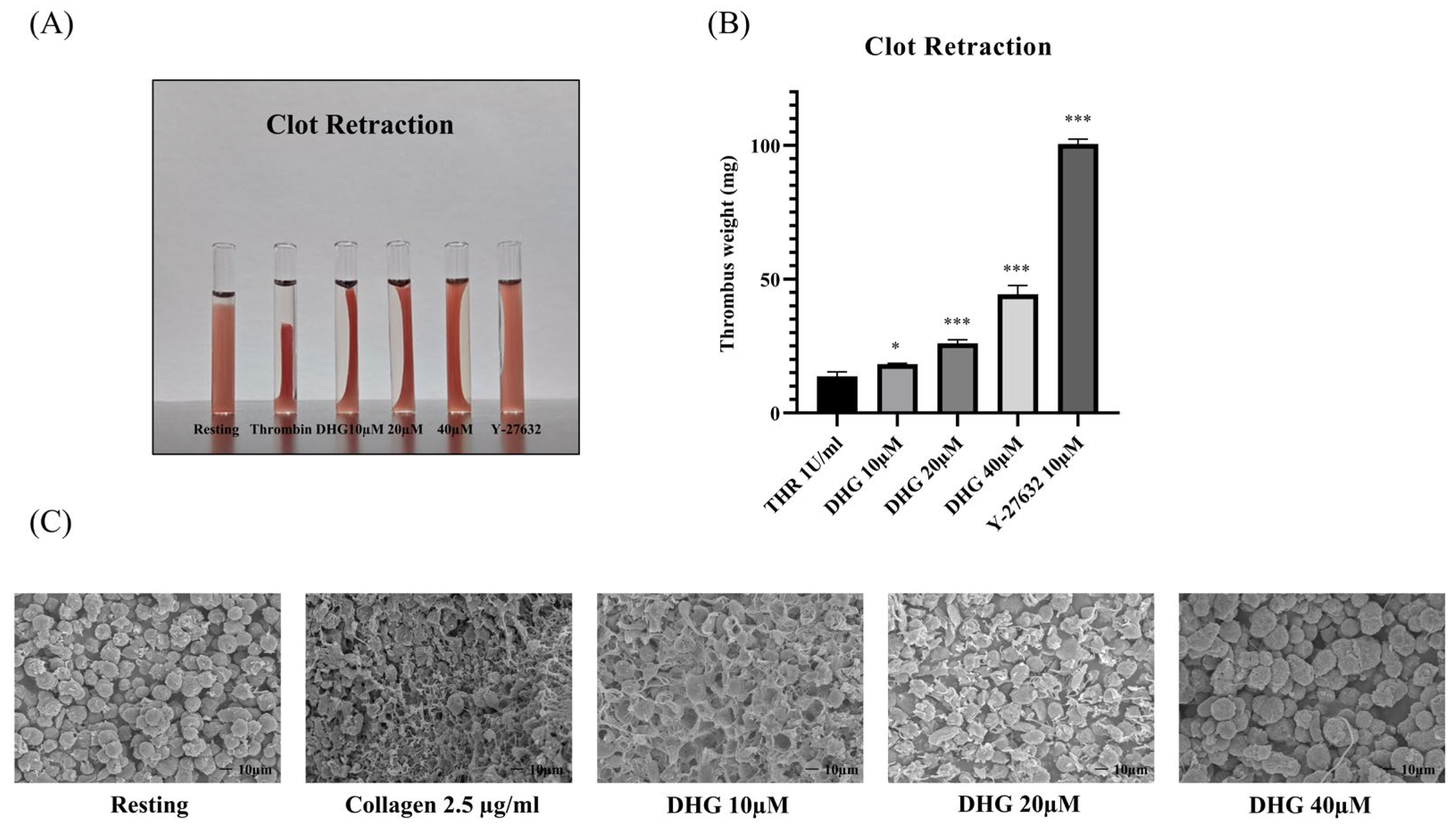
2.4. DHG Downregulates Inside–Out and Outside–In Signaling and Prevents Platelet Shape Change
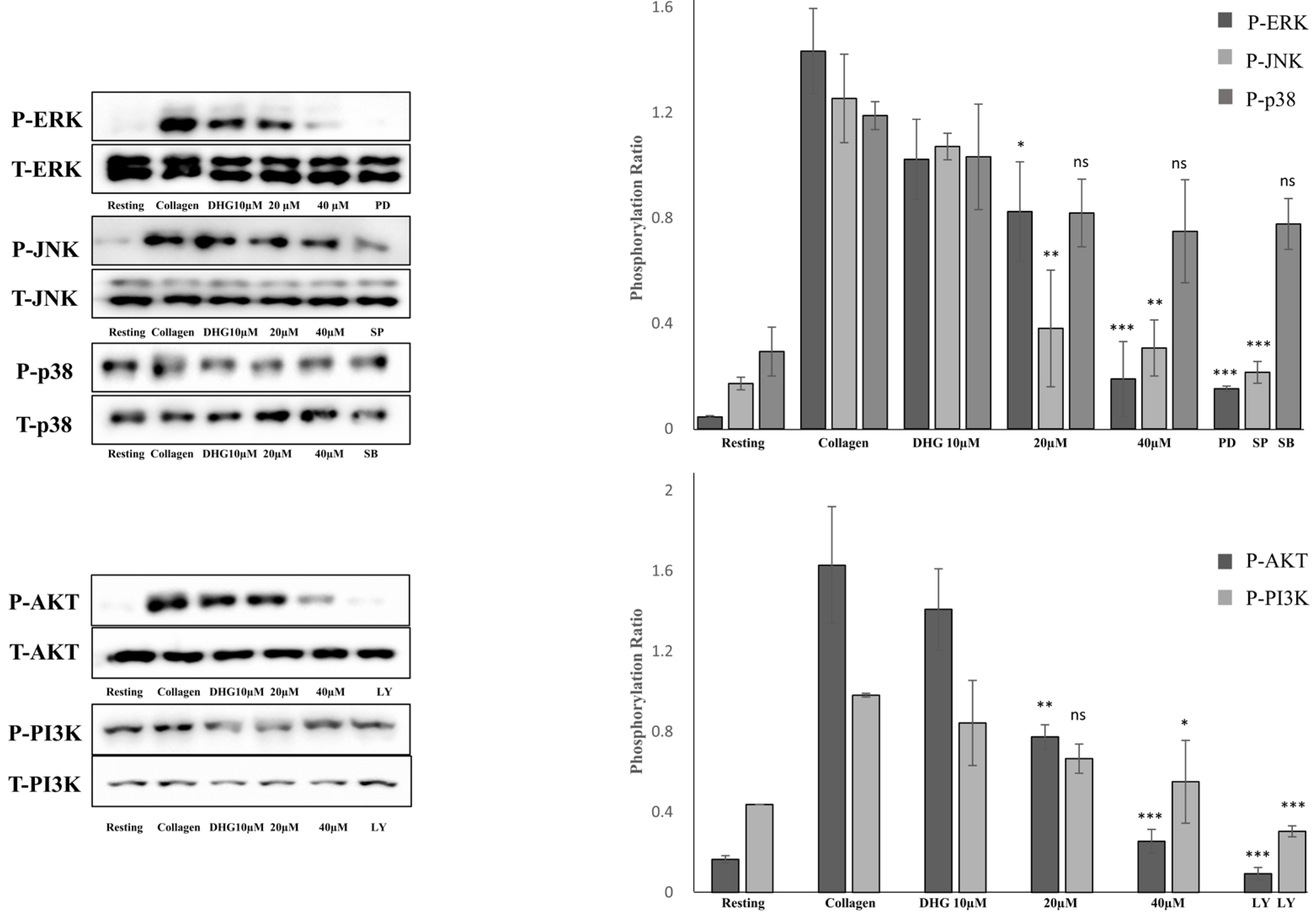
2.5. DHG Attenuates MAPK and PI3K/Akt Phosphorylation
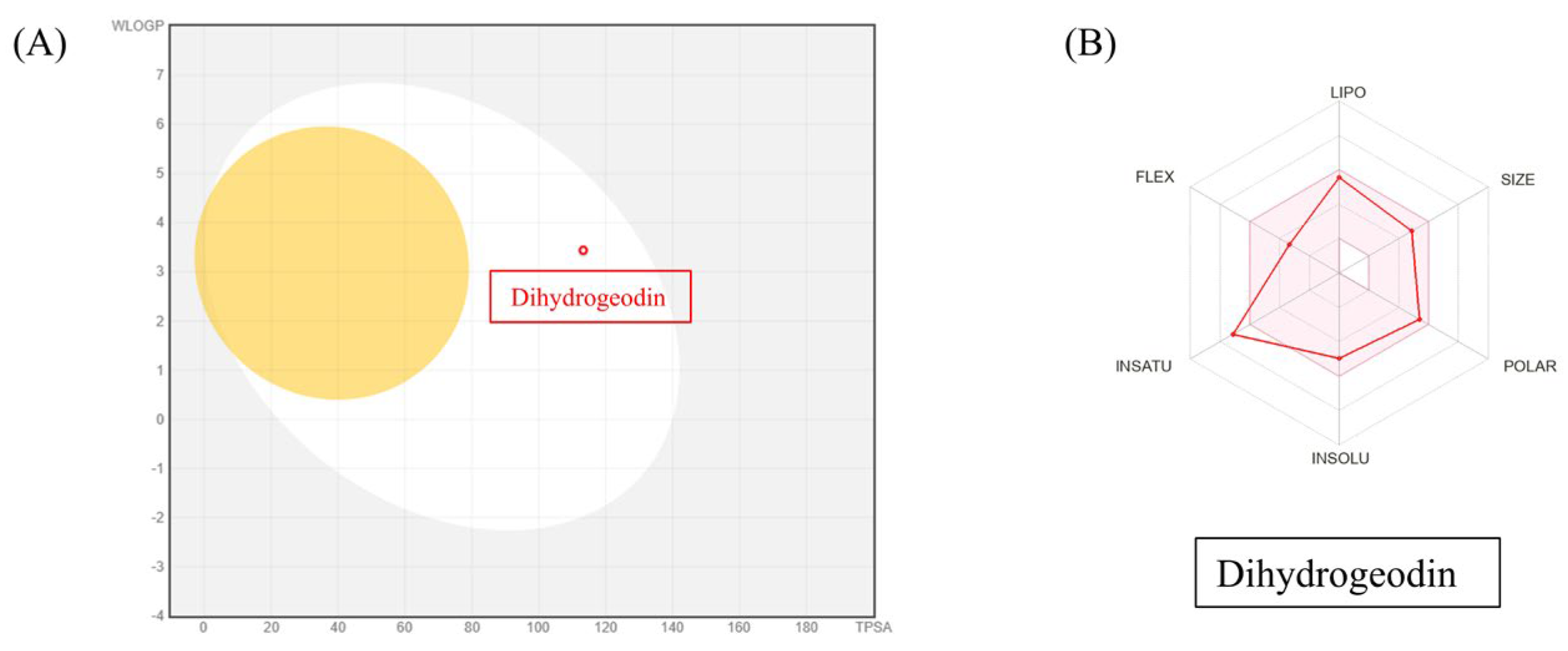
2.6. Pharmacokinetics, Drug-Likeliness Analysis
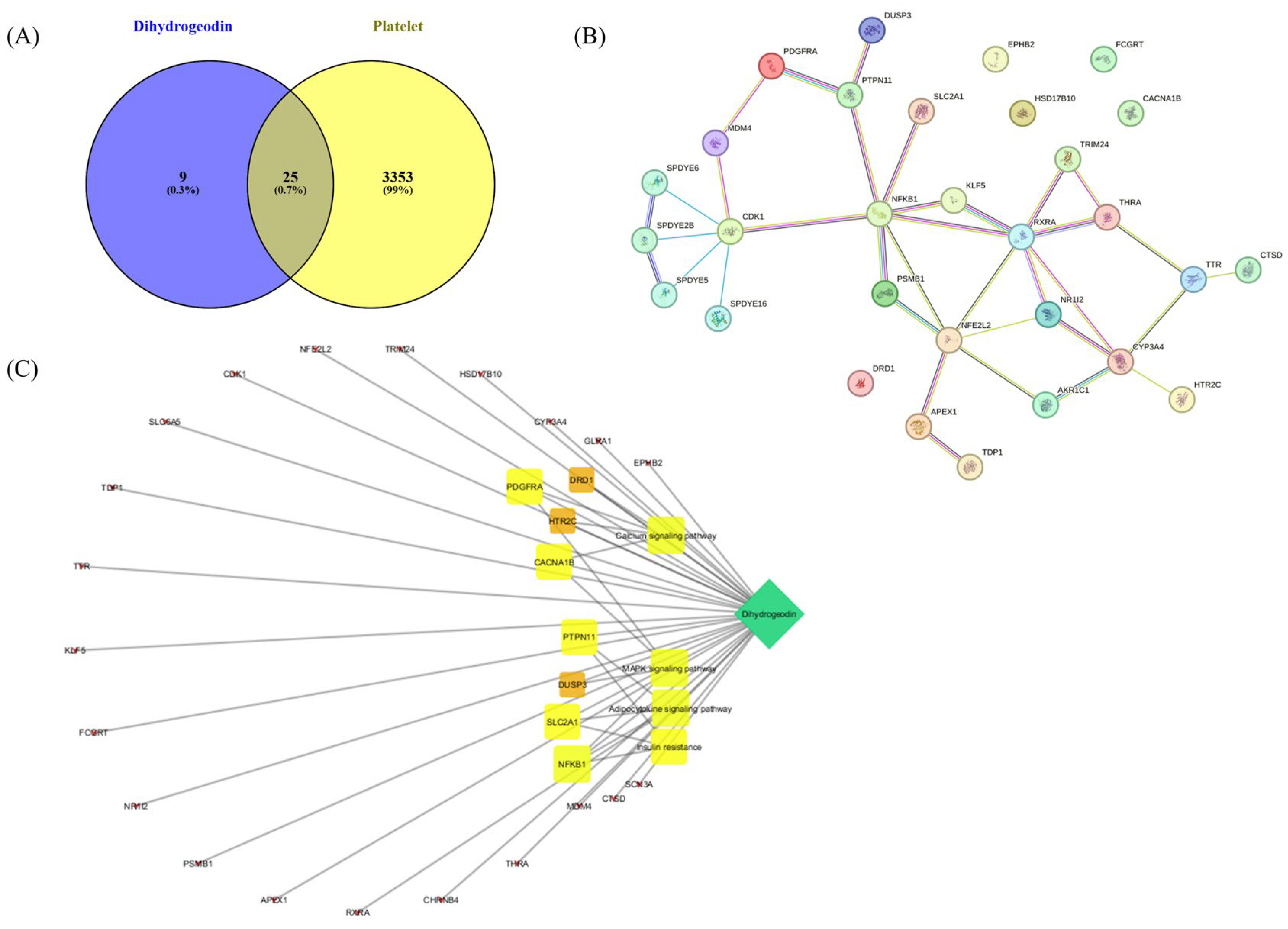
2.7. Network Pharmacology
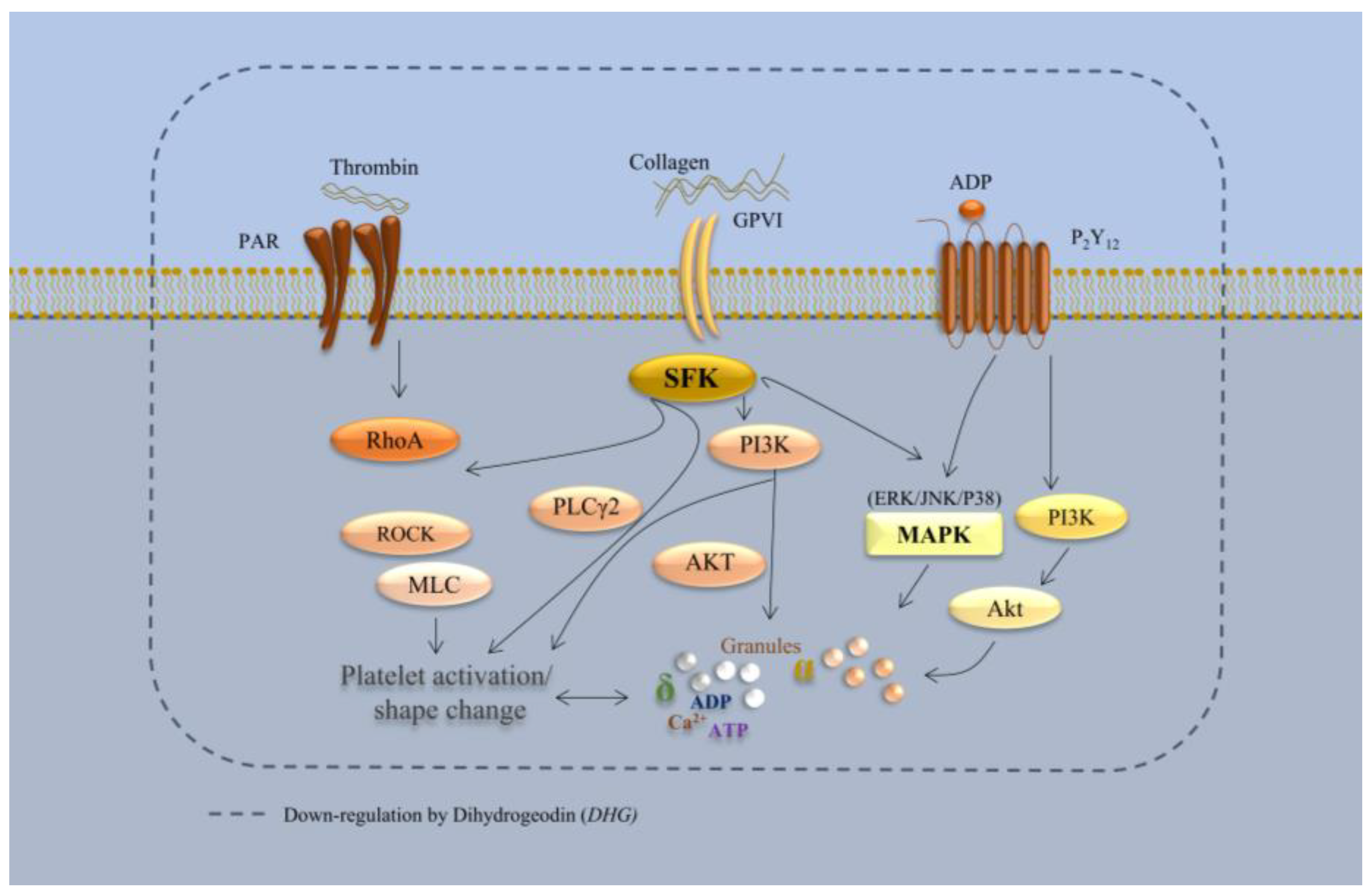
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Fungal Material and Fermentation
4.3. Extraction, Isolation, and Structure Determination
4.4. Experimental Animals
4.5. Light Transmission Aggregometry and Scanning Electron Microscopy (SEM)
4.6. ATP Release Assay, [Ca2+]i Mobilization Assay, and Fibrinogen Binding Assay
4.7. Clot Retraction
4.8. Western Blotting
4.9. Pharmacokinetics, Drug-Likeliness Analysis, and Network Pharmacology
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lockhart, P.B.; Sun, Y.P. Diseases of the cardiovascular system. In Burket’s Oral Medicine; John Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 505–552. [Google Scholar]
- Caffrey, C.; Leamy, A.; O’Sullivan, E.; Zabetakis, I.; Lordan, R.; Nasopoulou, C. Cardiovascular diseases and marine oils: A focus on omega-3 polyunsaturated fatty acids and polar lipids. Mar. Drugs 2023, 21, 549. [Google Scholar] [CrossRef] [PubMed]
- Scridon, A. Platelets and their role in hemostasis and thrombosis—From physiology to pathophysiology and therapeutic implications. Int. J. Mol. Sci. 2022, 23, 12772. [Google Scholar] [CrossRef] [PubMed]
- Młynarska, E.; Czarnik, W.; Fularski, P.; Hajdys, J.; Majchrowicz, G.; Stabrawa, M.; Rysz, J.; Franczyk, B. From atherosclerotic plaque to myocardial infarction—The leading cause of coronary artery occlusion. Int. J. Mol. Sci. 2024, 25, 7295. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, D.; Tan, P.; Xian, B.; Jiang, H.; Wu, Q.; Huang, X.; Zhang, P.; Xiao, X.; Pei, J. Mechanism of platelet activation and potential therapeutic effects of natural drugs. Phytomedicine 2023, 108, 154463. [Google Scholar] [CrossRef]
- Akram, A.W.; Saba, E.; Rhee, M.H. Antiplatelet and antithrombotic activities of Lespedeza cuneata via Pharmacological inhibition of integrin αIIbβ3, MAPK, and PI3K/AKT pathways and FeCl3-induced murine thrombosis. Evid.-Based Complement. Altern. Med. 2024, 2024, 9927160. [Google Scholar] [CrossRef]
- Chen, C.; Yang, F.-Q.; Zhang, Q.; Wang, F.-Q.; Hu, Y.-J.; Xia, Z.-N. Natural products for antithrombosis. Evid.-Based Complement. Altern. Med. 2015, 2015, 876426. [Google Scholar] [CrossRef]
- Li, D.; Li, Y.; Yang, S.; Yu, Z.; Xing, Y.; Wu, M. Mechanism and potential target of blood-activating Chinese botanical drugs combined with anti-platelet drugs: Prevention and treatment of atherosclerotic cardiovascular diseases. Front. Pharmacol. 2022, 13, 811422. [Google Scholar] [CrossRef]
- Krishnan, K.; Nguyen, T.N.; Appleton, J.P.; Law, Z.K.; Caulfied, M.; Cabrera, C.P.; Lenthall, R.; Hewson, D.; England, T.; McConachie, N. Antiplatelet resistance: A review of concepts, mechanisms, and implications for management in acute ischemic stroke and transient ischemic attack. Stroke Vasc. Interv. Neurol. 2023, 3, e000576. [Google Scholar] [CrossRef]
- Mussbacher, M.; Kral-Pointner, J.B.; Salzmann, M.; Schrottmaier, W.C.; Assinger, A. Mechanisms of hemostasis: Contributions of platelets, coagulation factors, and the vessel wall. In Fundamentals of Vascular Biology; Springer: Berlin/Heidelberg, Germany, 2024; pp. 167–203. [Google Scholar]
- Shehwar, D.; Barki, S.; Aliotta, A.; Calderara, D.B.; Veuthey, L.; Portela, C.P.; Alberio, L.; Alam, M.R. Platelets and mitochondria: The calcium connection. Mol. Biol. Rep. 2025, 52, 276. [Google Scholar] [CrossRef]
- Irfan, M.; Kwon, T.-H.; Kwon, H.-W.; Rhee, M.H. Pharmacological actions of dieckol on modulation of platelet functions and thrombus formation via integrin αIIbβ3 and cAMP signaling. Pharmacol. Res. 2022, 177, 106088. [Google Scholar] [CrossRef]
- Kanchanawong, P.; Calderwood, D.A. Organization, dynamics and mechanoregulation of integrin-mediated cell–ECM adhesions. Nat. Rev. Mol. Cell Biol. 2023, 24, 142–161. [Google Scholar] [CrossRef] [PubMed]
- Ngo, A.T.P.; Parra-Izquierdo, I.; Aslan, J.E.; McCarty, O.J.T. Rho GTPase regulation of reactive oxygen species generation and signalling in platelet function and disease. Small GTPases 2021, 12, 440–457. [Google Scholar] [CrossRef]
- Akram, A.W.; Shin, J.-H.; Batmunkh, U.; Saba, E.; Kang, Y.-M.; Jung, S.; Han, J.E.; Kim, S.D.; Kwak, D.; Kwon, H.-W. Ginsenoside Rg5 inhibits platelet aggregation by regulating GPVI signaling pathways and ferric chloride-induced thrombosis. J. Ginseng Res. 2025. [CrossRef]
- Li, Z.; Wang, Z.; Shen, B.; Chen, C.; Ding, X.; Song, H. Effects of aspirin on the gastrointestinal tract: Pros vs. cons. Oncol. Lett. 2020, 20, 2567–2578. [Google Scholar] [CrossRef]
- Bebars, A.E.; Alhazmi, A.A.; Aldajan, M.Y.; Alhujeri, N.A.F.; Alhaidari, R.M. Gastrointestinal side effects of antiplatelet drugs (Aspirin and Clopidogrel) in Medina, Saudi Arabia. Int. J. Med. Dev. Ctries. 2021, 5, 210. [Google Scholar] [CrossRef]
- Ferreira, M.; Freitas-Silva, M.; Assis, J.; Pinto, R.; Nunes, J.P.; Medeiros, R. The emergent phenomenon of aspirin resistance: Insights from genetic association studies. Pharmacogenomics 2020, 21, 125–140. [Google Scholar] [CrossRef]
- Hidayat, R.; Nabilah, R.A.; Rasyid, A.; Harris, S.; Harahap, A.R.; Louisa, M.; Listyaningsih, E.; Rambe, A.S.; Loho, T. Clopidogrel resistance among ischemic stroke patients and its risk factors in Indonesia. Acta Medica Acad. 2022, 51, 29. [Google Scholar] [CrossRef]
- Chaachouay, N.; Zidane, L. Plant-derived natural products: A source for drug discovery and development. Drugs Drug Candidates 2024, 3, 184–207. [Google Scholar] [CrossRef]
- Aware, C.B.; Patil, D.N.; Suryawanshi, S.S.; Mali, P.R.; Rane, M.R.; Gurav, R.G.; Jadhav, J.P. Natural bioactive products as promising therapeutics: A review of natural product-based drug development. South Afr. J. Bot. 2022, 151, 512–528. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Hashmi, H.A.; Muzammal, U.; Akram, A.W.; Alvi, M.A.; Talib, M.T.; Basharat, A.; Rauf, U.; Rahman, H.M.S.; ur Rahman, H.M.H. Use of natural feed additives as a remedy for diseases in veterinary medicine. In Complementary and Alternative Medicine: Feed Additives; Unique Scientific Publisher: Faisalabad, Pakistan, 2024; p. 104. [Google Scholar] [CrossRef]
- Wang, H.; He, D.; Duan, L.; Lv, L.; Gao, Q.; Wang, Y.; Yang, S.; Lv, Z. In Vivo anticoagulant and antithrombic activity of depolymerized glycosaminoglycan from Apostichopus japonicus and dynamic effect–exposure relationship in rat plasma. Mar. Drugs 2022, 20, 631. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, C.; Xia, Q.; Li, P.; Liu, K.; Zhang, Y. Brevianamide F exerts antithrombotic effects by modulating the MAPK signaling pathway and coagulation cascade. Mar. Drugs 2024, 22, 439. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhu, G.; Li, S.; Li, P.; Zhang, J.; Yin, R.; Yuan, L.; Gao, N.; Zhao, J. Fucosylated glycosaminoglycan oligosaccharide HS14, derived from Sea Cucumbers, is a novel inhibitor of platelet Toll-like receptor 2. Mar. Drugs 2025, 23, 110. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Y.; Chang, C.-C.; Tsai, Y.-H.; El-Shazly, M.; Wu, C.-C.; Wang, S.-W.; Hwang, T.-L.; Wei, C.-K.; Hohmann, J.; Yang, Z.-J. Anti-inflammatory, antiplatelet aggregation, and antiangiogenesis polyketides from Epicoccum sorghinum: Toward an understating of its biological activities and potential applications. ACS Omega 2020, 5, 11092–11099. [Google Scholar] [CrossRef]
- Zhao, L.; Lin, X.; Fu, J.; Zhang, J.; Tang, W.; He, Z. A novel bi-functional fibrinolytic enzyme with anticoagulant and thrombolytic activities from a marine-derived fungus Aspergillus versicolor ZLH-1. Mar. Drugs 2022, 20, 356. [Google Scholar] [CrossRef]
- Papikinou, M.-A.; Pavlidis, K.; Cholidis, P.; Kranas, D.; Adamantidi, T.; Anastasiadou, C.; Tsoupras, A. Marine fungi bioactives with anti-inflammatory, antithrombotic and antioxidant health-promoting properties against inflammation-related chronic diseases. Mar. Drugs 2024, 22, 520. [Google Scholar] [CrossRef]
- Ji, Y.B.; Chen, W.J.; Shan, T.Z.; Sun, B.Y.; Yan, P.C.; Jiang, W. Antibacterial diphenyl ether, benzophenone and xanthone derivatives from Aspergillus flavipes. Chem. Biodivers. 2020, 17, e1900640. [Google Scholar] [CrossRef]
- Peterson, S.W. Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 2008, 100, 205–226. [Google Scholar] [CrossRef]
- Choi, D.-C.; Ki, D.-W.; Kim, J.-Y.; Lee, I.-K.; Yun, B.-S. New dimeric 1, 3-dihydroisobenzofuran from culture broth of Fennellia flavipes. Phytochem. Lett. 2025, 65, 141–144. [Google Scholar] [CrossRef]
- Hamed, A.; Ismail, M.; El-Metwally, M.M.; Frese, M.; Stammler, H.G.; Sewald, N.; Shaaban, M. X-ray, structural assignment and molecular docking study of dihydrogeodin from Aspergillus Terreus TM8. Nat. Prod. Res. 2019, 33, 117–121. [Google Scholar] [CrossRef]
- Sato, S.; Okusa, N.; Ogawa, A.; Ikenoue, T.; Seki, T.; Tsuji, T. Identification and preliminary SAR studies of (+)-geodin as a glucose uptake stimulator for rat adipocytes. J. Antibiot. 2005, 58, 583–589. [Google Scholar] [CrossRef]
- Lemmens, T.P.; Luo, Q.; Wielders, S.J.H.; Scheijen, J.; Al-Nasiry, S.; Koenen, R.R.; Wenzel, P.; Cosemans, J. Platelet collagen receptors and their role in modulating platelet adhesion patterns and activation on alternatively processed collagen substrates. Thromb. Res. 2024, 244, 109201. [Google Scholar] [CrossRef] [PubMed]
- Gachet, C.; Hechler, B. Platelet purinergic receptors in thrombosis and inflammation. Hämostaseologie 2020, 40, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Wu, J.; Roest, M.; Heemskerk, J.W.M. Long-term platelet priming after glycoprotein VI stimulation in comparison to protease-activating receptor (PAR) stimulation. PLoS ONE 2021, 16, e0247425. [Google Scholar] [CrossRef]
- Huang, W.-C.; Lin, K.-C.; Hsia, C.-W.; Hsia, C.-H.; Chen, T.-Y.; Bhavan, P.S.; Sheu, J.-R.; Hou, S.-M. The antithrombotic agent pterostilbene interferes with integrin αIIbβ3-mediated inside-out and outside-in signals in human platelets. Int. J. Mol. Sci. 2021, 22, 3643. [Google Scholar] [CrossRef]
- Xiang, B.; Zhang, G.; Zhang, Y.; Wu, C.; Joshi, S.; Morris, A.J.; Ware, J.; Smyth, S.S.; Whiteheart, S.W.; Li, Z. Calcium ion chelation preserves platelet function during cold storage. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 234–249. [Google Scholar] [CrossRef]
- Lichota, A.; Szewczyk, E.M.; Gwozdzinski, K. Factors affecting the formation and treatment of thrombosis by natural and synthetic compounds. Int. J. Mol. Sci. 2020, 21, 7975. [Google Scholar] [CrossRef]
- Pan, N.; Li, Z.-C.; Li, Z.-H.; Chen, S.-H.; Jiang, M.-H.; Yang, H.-Y.; Liu, Y.-S.; Hu, R.; Zeng, Y.-W.; Dai, L.-H. Antiplatelet and antithrombotic effects of isaridin E isolated from the marine-derived fungus via downregulating the PI3K/Akt signaling pathway. Mar. Drugs 2021, 20, 23. [Google Scholar] [CrossRef]
- Vasconcelos, A.A.; Sucupira, I.D.; Guedes, A.L.; Queiroz, I.N.; Frattani, F.S.; Fonseca, R.J.; Pomin, V.H. Anticoagulant and antithrombotic properties of three structurally correlated sea urchin sulfated glycans and their low-molecular-weight derivatives. Mar. Drugs 2018, 16, 304. [Google Scholar] [CrossRef]
- Cao, S.; He, X.; Qin, L.; He, M.; Yang, Y.; Liu, Z.; Mao, W. Anticoagulant and antithrombotic properties in vitro and in vivo of a novel sulfated polysaccharide from marine green alga Monostroma nitidum. Mar. Drugs 2019, 17, 247. [Google Scholar] [CrossRef]
- Moura, L.d.A.; Marqui de Almeida, A.C.; Francielle Souza Domingos, T.; Ortiz-Ramirez, F.; Negrão Cavalcanti, D.; Laneuville Teixeira, V.; Lopes Fuly, A. Antiplatelet and anticoagulant effects of diterpenes isolated from the marine alga, Dictyota menstrualis. Mar. Drugs 2014, 12, 2471–2484. [Google Scholar] [CrossRef]
- Gholkar, A.A.; Nikam, Y.P.; Zambare, K.K.; Reddy, K.V.; Ghorpade, A.D. Potential anticoagulant herbal plants: A review. Asian J. Pharm. Sci. 2020, 10, 51–55. [Google Scholar] [CrossRef]
- Gao, H.; Huang, J.; Huang, X.; Lin, X.; Li, X.; Deng, H.; Zhou, Y.; Wu, L.; Xi, X.; Jin, J. Tryptanthrin impairs platelet function and thrombus formation by reducing Gp1bα expression. Eur. J. Pharmacol. 2025, 991, 177332. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, R.; Hua, C.; Wu, M.; Yuan, Y.; Zhang, L.; Guo, F.; Liu, J.; Yang, Z.; Liu, G. P2Y12 receptor-independent antiplatelet mechanism of cryptotanshinone: Network pharmacology and experimental validation of multi-target signaling pathways. J. Ethnopharmacol. 2025, 341, 119321. [Google Scholar] [CrossRef] [PubMed]
- Suades, R.; Padró, T.; Vilahur, G.; Badimon, L. Platelet-released extracellular vesicles: The effects of thrombin activation. Cell. Mol. Life Sci. 2022, 79, 190. [Google Scholar] [CrossRef]
- Zou, J.; Swieringa, F.; de Laat, B.; de Groot, P.G.; Roest, M.; Heemskerk, J.W.M. Reversible platelet integrin αIIbβ3 activation and thrombus instability. Int. J. Mol. Sci. 2022, 23, 12512. [Google Scholar] [CrossRef]
- Quah, Y.; Lee, Y.Y.; Lee, S.-J.; Kim, S.D.; Rhee, M.H.; Park, S.-C. In silico investigation of Panax ginseng lead compounds against COVID-19 associated platelet activation and thromboembolism. J. Ginseng Res. 2023, 47, 283–290. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akram, A.W.; Choi, D.-C.; Chae, H.-K.; Kim, S.D.; Kwak, D.; Yun, B.-S.; Rhee, M.H. Dihydrogeodin from Fennellia flavipes Modulates Platelet Aggregation via Downregulation of Calcium Signaling, αIIbβ3 Integrins, MAPK, and PI3K/Akt Pathways. Mar. Drugs 2025, 23, 212. https://doi.org/10.3390/md23050212
Akram AW, Choi D-C, Chae H-K, Kim SD, Kwak D, Yun B-S, Rhee MH. Dihydrogeodin from Fennellia flavipes Modulates Platelet Aggregation via Downregulation of Calcium Signaling, αIIbβ3 Integrins, MAPK, and PI3K/Akt Pathways. Marine Drugs. 2025; 23(5):212. https://doi.org/10.3390/md23050212
Chicago/Turabian StyleAkram, Abdul Wahab, Dae-Cheol Choi, Hyung-Kyu Chae, Sung Dae Kim, Dongmi Kwak, Bong-Sik Yun, and Man Hee Rhee. 2025. "Dihydrogeodin from Fennellia flavipes Modulates Platelet Aggregation via Downregulation of Calcium Signaling, αIIbβ3 Integrins, MAPK, and PI3K/Akt Pathways" Marine Drugs 23, no. 5: 212. https://doi.org/10.3390/md23050212
APA StyleAkram, A. W., Choi, D.-C., Chae, H.-K., Kim, S. D., Kwak, D., Yun, B.-S., & Rhee, M. H. (2025). Dihydrogeodin from Fennellia flavipes Modulates Platelet Aggregation via Downregulation of Calcium Signaling, αIIbβ3 Integrins, MAPK, and PI3K/Akt Pathways. Marine Drugs, 23(5), 212. https://doi.org/10.3390/md23050212