

In Vitro and In Silico Studies on the Anti-H1N1 Activity of Bioactive Compounds from Marine-Derived Streptomyces ardesiacus
 , , ,
, , , 
Abstract
1. Introduction
2. Results
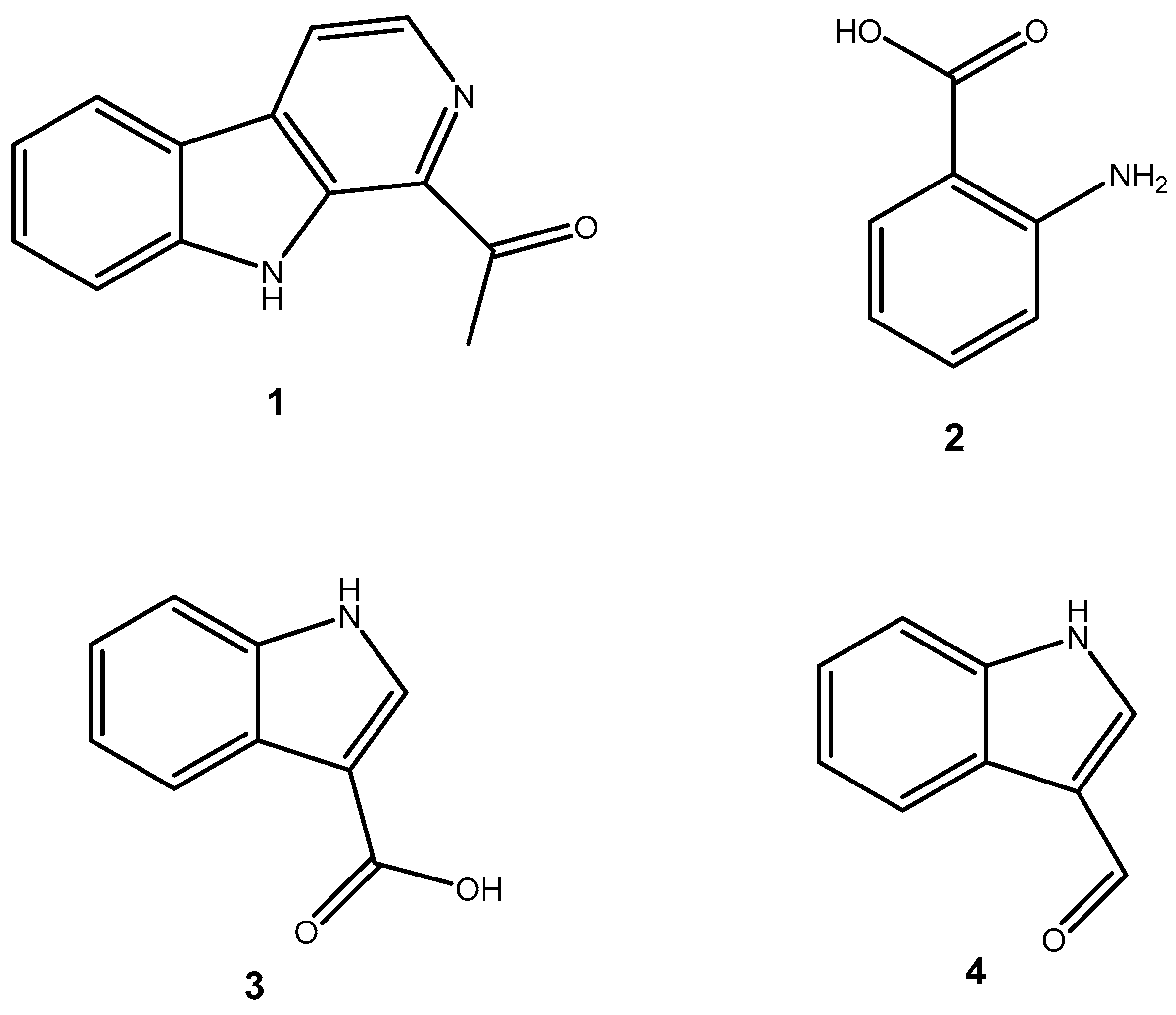
2.1. Structural Elucidation of Compounds Isolated from S. ardesiacus
2.2. Anti-Influenza Activity
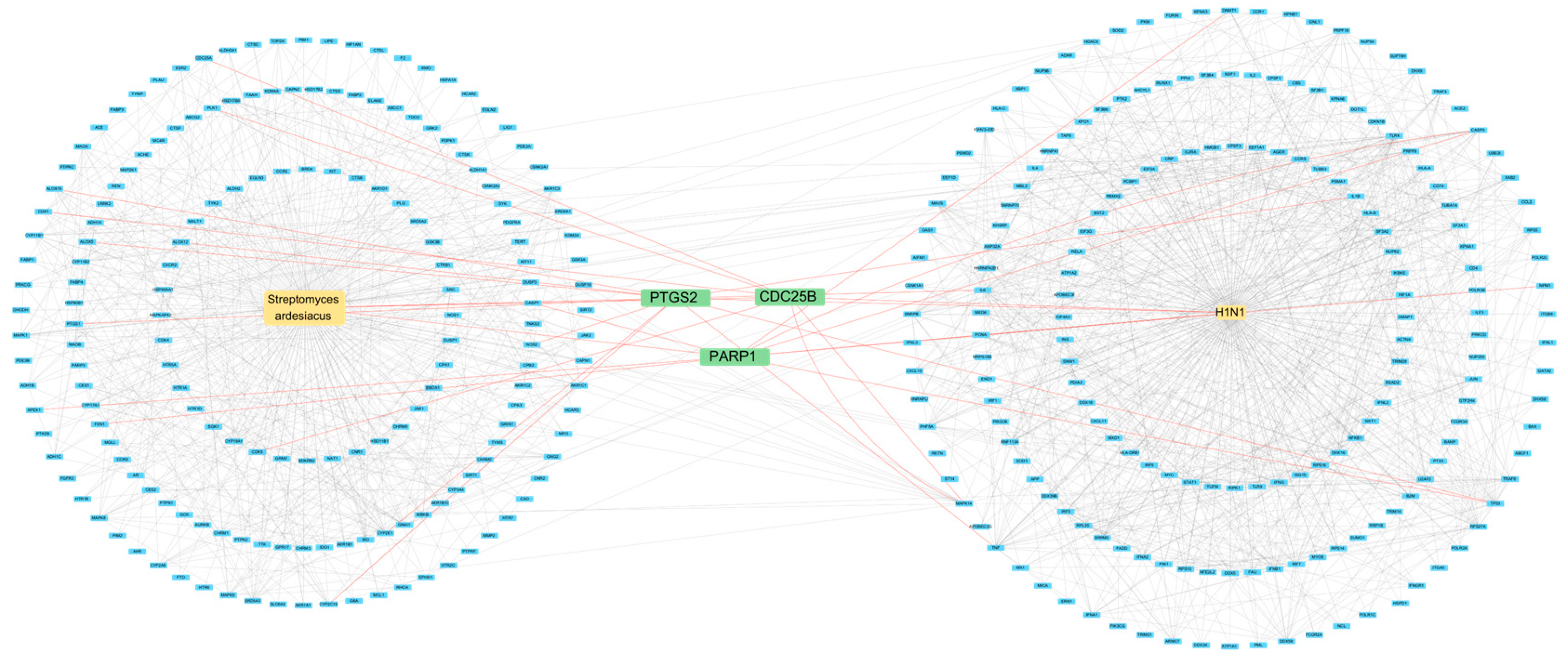
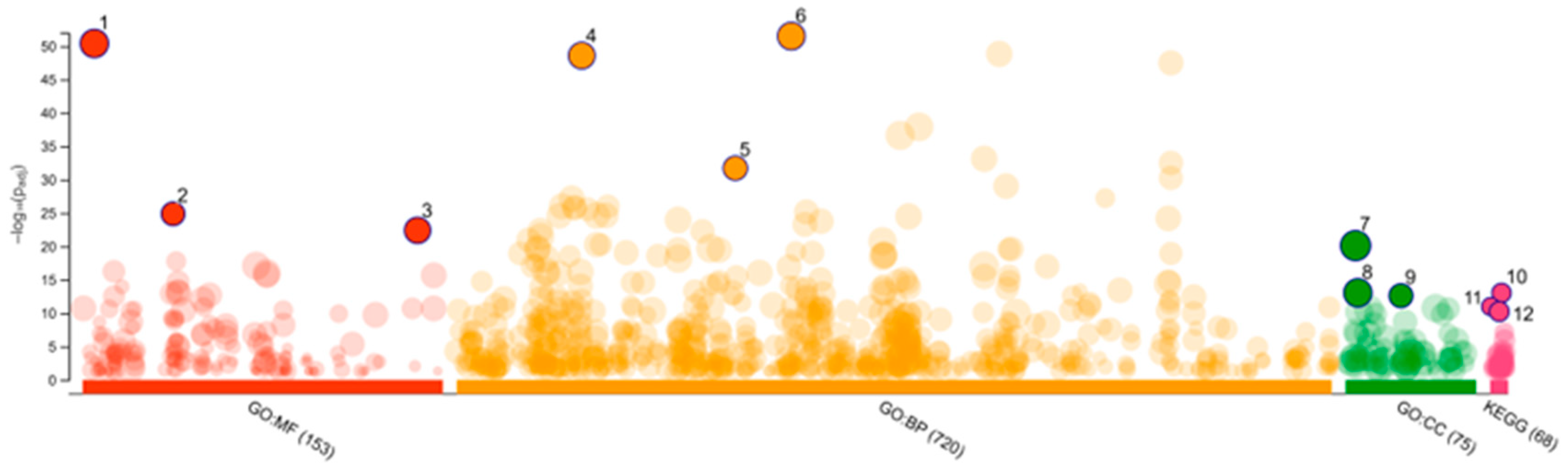
2.3. Network Pharmacology
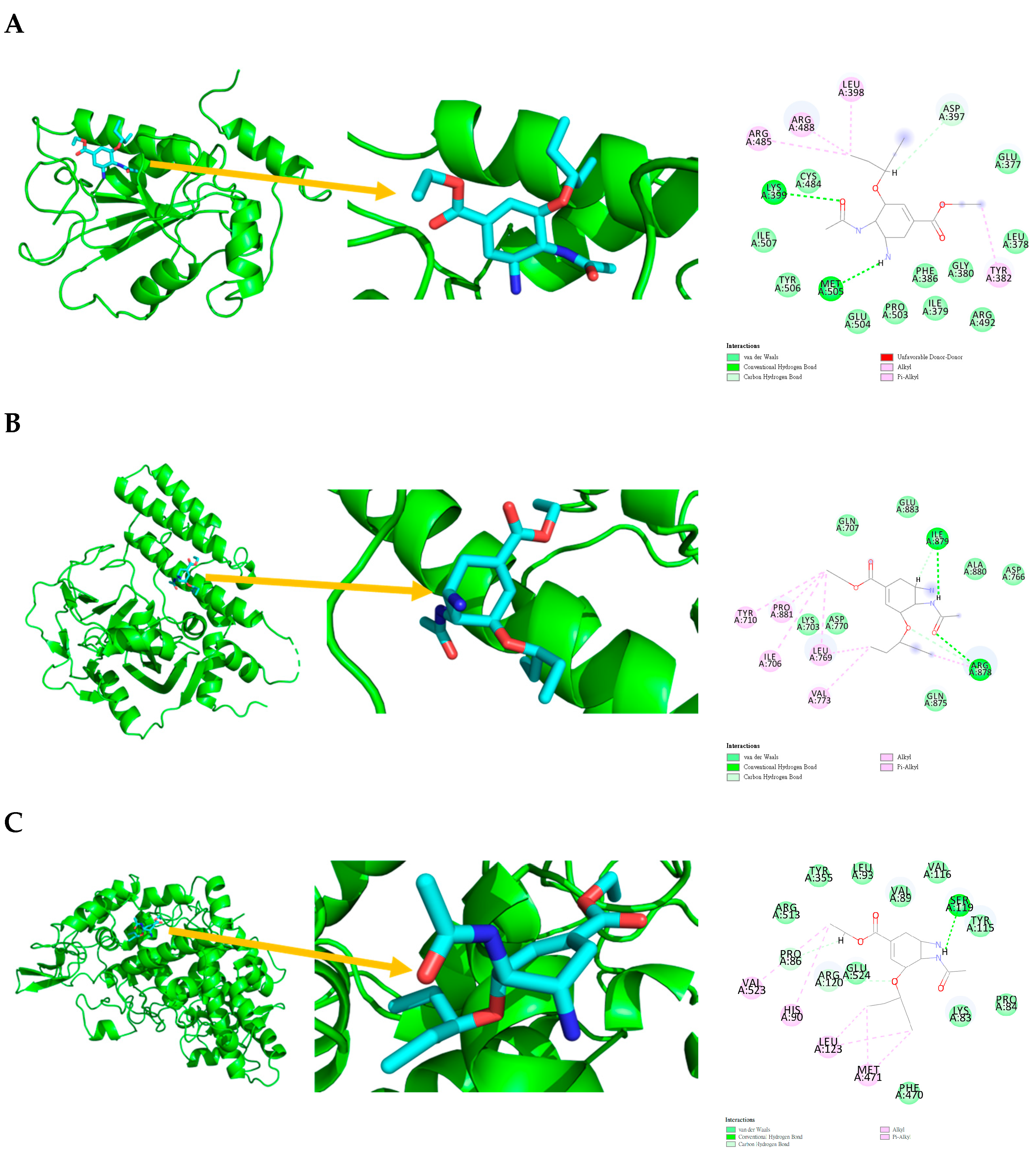
2.4. Molecular Docking
2.5. Molecular Dynamics
2.6. ADMET Analysis
3. Discussion
4. Materials and Methods
4.1. Instruments and Materials
4.1.1. Experimental Materials
Experimental Strains
4.1.2. Main Reagents and Instruments
Main Reagents
Main Instruments
4.2. Isolation Methods
4.2.1. Fermentation Conditions of S. ardesiacus
4.2.2. Extraction and Isolation of Metabolites from S. ardesiacus
Activation of S. ardesiacus
Fermentation Cultivation of S. ardesiacus
Extraction of Fermentation Products
4.3. Culture Conditions for Influenza Virus and MDCK Cells
4.4. Evaluation of Influenza Virus Infection
4.5. Network Pharmacology
4.5.1. Protein–Protein Interaction (PPI)
4.5.2. Gene Ontology Term Enrichment Analysis
4.6. Docking Stimulation
4.6.1. Ligand Preparation
4.6.2. Molecular Preparation and Docking analysis
4.7. Molecular Dynamics
4.7.1. System Preparation
4.7.2. Production Molecular Dynamics
4.8. ADMET Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BCRC | Bioresource Collection and Research Center |
| BP | Biological Process |
| CC | Cellular Component |
| COX-1 | Cyclooxygenase-1 |
| COX-2 | Cyclooxygenase-2 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| TLC | Thin-layer chromatography |
| FBS | Fetal bovine serum |
| FIRDI | Food Industry Research and Development Institute |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MDCK | Madin-Darby Canine Kidney |
| MF | Molecular Function |
| PPI | Protein–Protein Interaction |
| TPCK | L-1-tosylamido-2-phenylethyl chloromethyl ketone |
References
- Wikandari, R.; Hasniah, N.; Taherzadeh, M.J. The Role of Filamentous Fungi in Advancing the Development of a Sustainable Circular Bioeconomy. Bioresour. Technol. 2022, 345, 126531. [Google Scholar] [CrossRef]
- Flärdh, K. Growth Polarity and Cell Division in Streptomyces. Curr. Opin. Microbiol. 2003, 6, 564–571. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.-A. Chapter 11—Streptomyces-Based Cell Factories for Production of Biomolecules and Bioactive Metabolites. In Microbial Cell Factories Engineering for Production of Biomolecules; Singh, V.B.T., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 183–234. ISBN 978-0-12-821477-0. [Google Scholar]
- Engevik, K.A.; Hazzard, A.; Puckett, B.; Hoch, K.M.; Haidacher, S.J.; Haag, A.M.; Spinler, J.K.; Versalovic, J.; Engevik, M.A.; Horvath, T.D. Phylogenetically Diverse Bacterial Species Produce Histamine. Syst. Appl. Microbiol. 2024, 47, 126539. [Google Scholar] [CrossRef] [PubMed]
- Worrall, J.A.R.; Vijgenboom, E. Copper Mining in Streptomyces: Enzymes, Natural Products and Development. Nat. Prod. Rep. 2010, 27, 742–756. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.-S.; Kim, H.U.; Charusanti, P.; Palsson, B.Ø.; Lee, S.Y. Systems Biology and Biotechnology of Streptomyces Species for the Production of Secondary Metabolites. Biotechnol. Adv. 2014, 32, 255–268. [Google Scholar] [CrossRef]
- Yang, Z.; He, J.; Wei, X.; Ju, J.; Ma, J. Exploration and Genome Mining of Natural Products from Marine Streptomyces. Appl. Microbiol. Biotechnol. 2020, 104, 67–76. [Google Scholar] [CrossRef]
- Genilloud, O. Actinomycetes: Still a Source of Novel Antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef]
- Oren, A. Industrial and Environmental Applications of Halophilic Microorganisms. Environ. Technol. 2010, 31, 825–834. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-Mining the Microbial Treasures of the Ocean: New Natural Products. Biotechnol. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial Exopolysaccharides from Extreme Marine Habitats: Production, Characterization and Biological Activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Ventosa, A.; Fernández, A.B.; León, M.J.; Sánchez-Porro, C.; Rodriguez-Valera, F. The Santa Pola Saltern as a Model for Studying the Microbiota of Hypersaline Environments. Extremophiles 2014, 18, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhang, L.; Bazylinski, D.A. Deep-Sea Piezosphere and Piezophiles: Geomicrobiology and Biogeochemistry. Trends Microbiol. 2010, 18, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial Diversity and Biogeochemical Cycling in Soda Lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef]
- Murofushi, N.; Yamane, H.; Sakagami, Y.; Imaseki, H.; Kamiya, Y.; Iwamura, H.; Hirai, N.; Tsuji, H.; Yokota, T.; Ueda, J. 8.02-Plant Hormones. In Comprehensive Natural Products Chemistry; Barton, S.D., Nakanishi, K., Meth-Cohn, O.B.T., Eds.; Pergamon: Oxford, UK, 1999; pp. 19–136. ISBN 978-0-08-091283-7. [Google Scholar]
- Umer, S.M.; Solangi, M.; Khan, K.M.; Saleem, R.S.Z. Indole-Containing Natural Products 2019–2022: Isolations, Reappraisals, Syntheses, and Biological Activities. Molecules 2022, 27, 7586. [Google Scholar] [CrossRef]
- Rani, A.; Saini, K.C.; Bast, F.; Varjani, S.; Mehariya, S.; Bhatia, S.K.; Sharma, N.; Funk, C. A Review on Micro-bial Products and Their Perspective Application as Antimicrobial Agents. Biomolecules 2021, 11, 1860. [Google Scholar] [CrossRef]
- Hezaveh, K.; Shinde, R.S.; Klötgen, A.; Halaby, M.J.; Lamorte, S.; Ciudad, M.T.; Quevedo, R.; Neufeld, L.; Liu, Z.Q.; Jin, R.; et al. Tryptophan-Derived Microbial Metabolites Activate the Aryl Hydrocarbon Receptor in Tumor-Associated Macrophages to Suppress Anti-Tumor Immunity. Immunity 2022, 55, 324–340.e8. [Google Scholar] [CrossRef]
- Anh, C.V.; Kwon, J.-H.; Kang, J.S.; Lee, H.-S.; Heo, C.-S.; Shin, H.J. New Angucycline Glycosides from a Marine-Derived Bacterium Streptomyces Ardesiacus. Int. J. Mol. Sci. 2022, 23, 13779. [Google Scholar] [CrossRef]
- Qin, R.; Zhao, Q.; Han, B.; Zhu, H.-P.; Peng, C.; Zhan, G.; Huang, W. Indole-Based Small Molecules as Potential Therapeutic Agents for the Treatment of Fibrosis. Front. Pharmacol. 2022, 13, 845892. [Google Scholar] [CrossRef]
- Wang, H.; Cui, E.; Li, J.; Ma, X.; Jiang, X.; Du, S.; Qian, S.; Du, L. Design and Synthesis of Novel Indole and Indazole-Piperazine Pyrimidine Derivatives with Anti-Inflammatory and Neuroprotective Activities for Ischemic Stroke Treatment. Eur. J. Med. Chem. 2022, 241, 114597. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Lee, J.-H.; Lee, J. Diverse Roles of Microbial Indole Compounds in Eukaryotic Systems. Biol. Rev. Camb. Philos. Soc 2021, 96, 2522–2545. [Google Scholar] [CrossRef] [PubMed]
- Dable-Tupas, G.; Tulika, V.; Jain, V.; Maheshwari, K.; Brakad, D.D.; Naresh, P.N.; Suruthimeenakshi, S. 11-Bioactive Compounds of Nutrigenomic Importance. In Role of Nutrigenomics in Modern-day Healthcare and Drug Discovery; Dable-Tupas, G., Egbuna, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 301–342. ISBN 978-0-12-824412-8. [Google Scholar]
- Faheem; Kumar, B.K.; Sekhar, K.V.G.C.; Kunjiappan, S.; Jamalis, J.; Balaña-Fouce, R.; Sankaranarayanan, M. Recent Update on the Anti-Infective Potential of β-Carboline Analogs. Mini. Rev. Med. Chem. 2021, 21, 398–425. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Yu, H.; Li, Z.; Cao, Z.; Du, Y.; Dai, J.; Zhi, D.; Xu, Y.; Li, N.; Wang, J. β-Carboline Dimers Inhibit the Tumor Proliferation by the Cell Cycle Arrest of Sarcoma through Intercalating to Cyclin-A2. Front. Immunol. 2022, 13, 922183. [Google Scholar] [CrossRef]
- Thatikayala, M.; Wadhwa, P.; Kaur, P.; Singh, P.K.; Yadav, A.; Kaushik, M.; Sahu, S.K. Beta-Carboline as a Promising Heterocyclic Nucleus: Synthetic Aspects, Pharmacological Potential and Structure Activity Relationship. Eur. J. Med. Chem. Rep. 2022, 6, 100096. [Google Scholar] [CrossRef]
- Cao, R.; Peng, W.; Wang, Z.; Xu, A. Beta-Carboline Alkaloids: Biochemical and Pharmacological Functions. Curr. Med. Chem. 2007, 14, 479–500. [Google Scholar] [CrossRef]
- Xu, T.; Shi, L.; Zhang, Y.; Wang, K.; Yang, Z.; Ke, S. Synthesis and Biological Evaluation of Marine Alkaloid-Oriented β-Carboline Analogues. Eur. J. Med. Chem. 2019, 168, 293–300. [Google Scholar] [CrossRef]
- Dai, J.; Dan, W.; Schneider, U.; Wang, J. β-Carboline Alkaloid Monomers and Dimers: Occurrence, Structural Diversity, and Biological Activities. Eur. J. Med. Chem. 2018, 157, 622–656. [Google Scholar]
- Kim, H.; Sablin, S.O.; Ramsay, R.R. Inhibition of Monoamine Oxidase A by Beta-Carboline Derivatives. Arch Biochem Biophys 1997, 337, 137–142. [Google Scholar] [CrossRef]
- Berlowitz, I.; Egger, K.; Cumming, P. Monoamine Oxidase Inhibition by Plant-Derived β-Carbolines; Implications for the Psychopharmacology of Tobacco and Ayahuasca. Front Pharmacol 2022, 13, 886408. [Google Scholar]
- Yu, L.; Shen, N.; Ren, J.; Xin, H.; Cui, Y. Resource Distribution, Pharmacological Activity, Toxicology and Clinical Drugs of β-Carboline Alkaloids: An Updated and Systematic Review. Fitoterapia 2024, 106326. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, T. β-Carboline Alkaloids in Soy Sauce and Inhibition of Monoamine Oxidase (MAO). Molecules 2023, 28, 2723. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Zuo, Y.; Tu, N.; Guo, J.; Liu, Z.; Cao, S.; Long, S. Diverse Pharmacological Activities of β-Carbolines: Substitution Patterns, SARs and Mechanisms of Action. Eur. J. Med. Chem. 2025, 287, 117350. [Google Scholar] [CrossRef] [PubMed]
- Breine, A.; Van Gysel, M.; Elsocht, M.; Whiteway, C.; Philippe, C.; Quinet, T.; Valcek, A.; Wouters, J.; Ballet, S.; Van der Henst, C. Antimicrobial Activity of a Repurposed Harmine-Derived Compound on Carbapenem-Resistant Acinetobacter Baumannii Clinical Isolates. Front. Cell. Infect. Microbiol. 2021, 11, 789672. [Google Scholar] [CrossRef]
- Zou, R.; Wei, C.; Zhang, X.; Zhou, D.; Xu, J. Alkaloids from Endophytic Fungus Aspergillus Fumigatus HQD24 Isolated from the Chinese Mangrove Plant Rhizophora Mucronata. Nat. Prod. Res. 2022, 36, 5069–5073. [Google Scholar] [CrossRef]
- Khalil, Z.G.; Kankanamge, S.; Capon, R.J. Structure Revision of Penipacids A–E Reveals a Putative New Cryptic Natural Product, N-Aminoanthranilic Acid, with Potential as a Transcriptional Regulator of Silent Secondary Metabolism. Mar. Drugs 2022, 20, 339. [Google Scholar] [CrossRef]
- Hagemeier, J.; Schneider, B.; Oldham, N.J.; Hahlbrock, K. Accumulation of Soluble and Wall-Bound Indolic Metabolites in Arabidopsis Thaliana Leaves Infected with Virulent or Avirulent Pseudomonas Syringae Pathovar Tomato Strains. Proc. Natl. Acad. Sci. USA 2001, 98, 753–758. [Google Scholar] [CrossRef]
- Liu, X.-L.; Li, M.-Y. Study on Chemical Constituents of Sceletium tortuosum. Nat. Prod. Res. Dev. 2020, 32, 441–446. [Google Scholar]
- Berendsen, H.J.C.; van der Spoel, D.; van Drunen, R. GROMACS: A Message-Passing Parallel Molecular Dynamics Implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- Chakravarti, I.M.; Laha, R.G.; Roy, J.; Roy, J. Handbook of Methods of Applied Statistics; Wiley: New York, NY, USA, 1967; Volume 1. [Google Scholar]
- Macbeth, G.; Razumiejczyk, E.; Ledesma, R.D. Cliff’s Delta Calculator: A Non-Parametric Effect Size Program for Two Groups of Observations. Univ. Psychol. 2011, 10, 545–555. [Google Scholar]
- Stephen, A.M.M.M.S.; Lawrie, M. Research Methods, Statistics and Evidence-Based Practice. In Companion to Psychiatric Studies E-Book: Companion to Psychiatric Studies E-Book; Churchill Livingstone: London, UK, 2010; Volume 157. [Google Scholar]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Drug Transport across the Blood–Brain Barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1959–1972. [Google Scholar] [PubMed]
- Potts, R.O.; Guy, R.H. Predicting Skin Permeability. Pharm. Res. 1992, 9, 663–669. [Google Scholar] [CrossRef]
- Shin, H.-J.; Lee, H.-S.; Lee, D.-S. The Synergistic Antibacterial Activity of 1-Acetyl-β-Carboline and β-Lactams Against Methicillin-Resistant Staphylococcus Aureus (MRSA). J. Microbiol. Biotechnol. 2010, 20, 501–505. [Google Scholar] [PubMed]
- Loboda, M.; Biliavska, L.; Iutynska, G.; Newitt, J.; Mariychuk, R. Natural Products Biosynthesis by Streptomyces netropsis IMV Ac-5025 under Exogenous Sterol Action. Antibiotics 2024, 13, 146. [Google Scholar] [CrossRef]
- Fang, Q.; Maglangit, F.; Mugat, M.; Urwald, C.; Kyeremeh, K.; Deng, H. Targeted Isolation of Indole Alkaloids from Streptomyces Sp. CT37. Molecules 2020, 25, 1108. [Google Scholar] [CrossRef]
- Aoki, Y.; Yoshida, Y.; Yoshida, M.; Kawaide, H.; Abe, H.; Natsume, M. Anthranilic Acid, a Spore Germination Inhibitor of Phytopathogenic Streptomyces Sp. B-9-1 Causing Root Tumor of Melon. Actinomycetologica 2005, 19, 48–54. [Google Scholar]
- Li, Y.; Reeves, R.M.; Wang, X.; Bassat, Q.; Brooks, W.A.; Cohen, C.; Moore, D.P.; Nunes, M.; Rath, B.; Campbell, H.; et al. Global Patterns in Monthly Activity of Influenza Virus, Respiratory Syncytial Virus, Parainfluenza Virus, and Metapneumovirus: A Systematic Analysis. Lancet Glob Health 2019, 7, e1031–e1045. [Google Scholar] [CrossRef]
- Perwitasari, O.; Torrecilhas, A.C.; Yan, X.; Johnson, S.; White, C.; Tompkins, S.M.; Tripp, R.A. Targeting Cell Division Cycle 25 Homolog B to Regulate Influenza Virus Replication. J. Virol. 2013, 87, 13775–13784. [Google Scholar] [CrossRef]
- Petrova, V.N.; Russell, C.A. The Evolution of Seasonal Influenza Viruses. Nat. Rev. Microbiol. 2018, 16, 47–60. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Kash, J.C. Influenza Virus Evolution, Host Adaptation, and Pandemic Formation. Cell. Host Microbe 2010, 7, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Galaktionov, K.; Beach, D. Specific Activation of Cdc25 Tyrosine Phosphatases by B-Type Cyclins: Evidence for Multiple Roles of Mitotic Cyclins. Cell 1991, 67, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Mahesutihan, M.; Zheng, W.; Meng, L.; Fan, W.; Li, J.; Ye, X.; Liu, W.; Sun, L. CDC25B Promotes Influenza A Virus Replication by Regulating the Phosphorylation of Nucleoprotein. Virology 2018, 525, 40–47. [Google Scholar] [CrossRef]
- Kotova, E.; Jarnik, M.; Tulin, A. V Uncoupling of the Transactivation and Transrepression Functions of PARP1 Protein. Proc. Natl. Acad. Sci. USA 2010, 107, 6406–6411. [Google Scholar] [CrossRef]
- Westera, L.; Jennings, A.M.; Maamary, J.; Schwemmle, M.; García-Sastre, A.; Bortz, E. Poly-ADP Ribosyl Polymerase 1 (PARP1) Regulates Influenza A Virus Polymerase. Adv. Virol. 2019, 2019, 8512363. [Google Scholar]
- Bonfiglio, J.J.; Leidecker, O.; Dauben, H.; Longarini, E.J.; Colby, T.; San Segundo-Acosta, P.; Perez, K.A.; Matic, I. An HPF1/PARP1-Based Chemical Biology Strategy for Exploring ADP-Ribosylation. Cell 2020, 183, 1086–1102.e23. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Kraus, W.L. The Expanding Universe of PARP1-Mediated Molecular and Therapeutic Mechanisms. Mol. Cell. 2022, 82, 2315–2334. [Google Scholar] [CrossRef]
- Huang, Z.; Kang, S.G.; Li, Y.; Zak, J.; Shaabani, N.; Deng, K.; Shepherd, J.; Bhargava, R.; Teijaro, J.R.; Xiao, C. IFNAR1 Signaling in NK Cells Promotes Persistent Virus Infection. Sci. Adv. 2021, 7, eabb8087. [Google Scholar] [CrossRef]
- Yaugel-Novoa, M.; Bourlet, T.; Longet, S.; Botelho-Nevers, E.; Paul, S. Association of IFNAR1 and IFNAR2 with COVID-19 Severity. Lancet Microbe 2023, 4, e487. [Google Scholar]
- Zhang, Z.; Uribe, I.; Davis, K.A.; McPherson, R.L.; Larson, G.P.; Badiee, M.; Tran, V.; Ledwith, M.P.; Feltman, E.; Yú, S.; et al. Global Remodeling of ADP-Ribosylation by PARP1 Suppresses Influenza A Virus Infection. bioRxiv 2024. [Google Scholar] [CrossRef]
- Xia, C.; Wolf, J.J.; Sun, C.; Xu, M.; Studstill, C.J.; Chen, J.; Ngo, H.; Zhu, H.; Hahm, B. PARP1 Enhances Influenza A Virus Propagation by Facilitating Degradation of Host Type I Interferon Receptor. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Wang, X.; Jeschke, U.; von Schönfeldt, V. COX-2-PGE(2)-EPs in Gynecological Cancers. Arch. Gynecol. Obstet. 2020, 301, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Darwish, I.; Mubareka, S.; Liles, W.C. Immunomodulatory Therapy for Severe Influenza. Expert. Rev. Anti. Infect. Ther. 2011, 9, 807–822. [Google Scholar] [CrossRef]
- Liu, L.; Cao, Z.; Chen, J.; Li, R.; Cao, Y.; Zhu, C.; Wu, K.; Wu, J.; Liu, F.; Zhu, Y. Influenza A Virus Induces Interleukin-27 through Cyclooxygenase-2 and Protein Kinase A Signaling. J. Biol. Chem. 2012, 287, 11899–11910. [Google Scholar] [CrossRef] [PubMed]
- Chun, K.-S.; Kim, E.-H.; Kim, D.-H.; Song, N.-Y.; Kim, W.; Na, H.-K.; Surh, Y.-J. Targeting Cyclooxygenase-2 for Chemoprevention of Inflammation-Associated Intestinal Carcinogenesis: An Update. Biochem. Pharmacol. 2024, 228, 116259. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. Prostaglandins and Cancer. Gut 2006, 55, 115–122. [Google Scholar] [CrossRef]
- Hu, H.; Hu, C.; Peng, J.; Ghosh, A.K.; Khan, A.; Sun, D.; Luyten, W. Bioassay-Guided Interpretation of Antimicrobial Compounds in Kumu, a TCM Preparation from Picrasma Quassioides’ Stem via UHPLC-Orbitrap-Ion Trap Mass Spectrometry Combined with Fragmentation and Retention Time Calculation. Front. Pharmacol. 2021, 12, 761751. [Google Scholar] [CrossRef]
- Ribas, J.; Cubero, E.; Luque, F.J.; Orozco, M. Theoretical Study of Alkyl-π and Aryl-π Interactions. Reconciling Theory and Experiment. J. Org. Chem. 2002, 67, 7057–7065. [Google Scholar] [CrossRef]
- Chang, T.H.; Zink, J.I. The.Sigma. and.Pi. Interactions of the Carbonyl Ligand Determined from Single-Crystal Polarized Electronic Spectroscopy and Ligand Field Theory. J. Am. Chem. Soc. 1987, 109, 692–698. [Google Scholar] [CrossRef]
- Wan, J.; Luo, C. Accumulation of Hydrogen Bonds and van Der Waals Interactions Determines Force Response between Two Parallel Cellulose Chains: Steered Molecular Dynamics Simulations. J. Phys. Chem. B 2024, 128, 6742–6750. [Google Scholar] [CrossRef]
- Le, D.-N.; Rodriguez-Lopez, P.; Woods, L.M. Nonlinear Effects in Manybody van Der Waals Interactions. Phys. Rev. Res. 2024, 6, 13289. [Google Scholar] [CrossRef]
- Carter-Fenk, K.; Herbert, J.M. Reinterpreting π-Stacking. Phys. Chem. Chem. Phys. 2020, 22, 24870–24886. [Google Scholar] [CrossRef]
- Motherwell, W.B.; Moreno, R.B.; Pavlakos, I.; Arendorf, J.R.T.; Arif, T.; Tizzard, G.J.; Coles, S.J.; Aliev, A.E. Noncovalent Interactions of π Systems with Sulfur: The Atomic Chameleon of Molecular Recognition. Angew. Chem. Int. Ed. Engl. 2018, 57, 1193–1198. [Google Scholar] [CrossRef]
- Zhang, H.; Fang, Y.; Yang, F.; Liu, X.; Lu, X. Aromatic Organic Molecular Crystal with Enhanced π–π Stacking Interaction for Ultrafast Zn-Ion Storage. Energy Environ. Sci. 2020, 13, 2515–2523. [Google Scholar] [CrossRef]
- Deng, J.-H.; Luo, J.; Mao, Y.-L.; Lai, S.; Gong, Y.-N.; Zhong, D.-C.; Lu, T.-B. π-π Stacking Interactions: Non-Negligible Forces for Stabilizing Porous Supramolecular Frameworks. Sci. Adv. 2024, 6, eaax9976. [Google Scholar] [CrossRef]
- Fargher, H.A.; Sherbow, T.J.; Haley, M.M.; Johnson, D.W.; Pluth, M.D. C–H⋯S Hydrogen Bonding Interactions. Chem. Soc. Rev. 2022, 51, 1454–1469. [Google Scholar] [CrossRef]
- Horowitz, S.; Trievel, R.C. Carbon-Oxygen Hydrogen Bonding in Biological Structure and Function. J. Biol. Chem. 2012, 287, 41576–41582. [Google Scholar] [CrossRef]
- De Clercq, E.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A Web Server for Target Prediction of Bioactive Small Molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2025 Update. Nucleic. Acids. Res. 2025, 53, 1516–1525. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.S.; Rohane, S.H. Review on Discovery Studio: An Important Tool for Molecular Docking. Asian J. Chem. 2021, 1, 1–3. [Google Scholar]
- Baroroh, U.; Biotek, M.; Muscifa, Z.S.; Destiarani, W.; Rohmatullah, F.G.; Yusuf, M. Molecular Interaction Analysis and Visualization of Protein-Ligand Docking Using Biovia Discovery Studio Visualizer. Indones. J. Biotechnol. 2023, 2, 22–30. [Google Scholar]
- Andrio, P.; Hospital, A.; Conejero, J.; Jordá, L.; Del Pino, M.; Codo, L.; Soiland-Reyes, S.; Goble, C.; Lezzi, D.; Badia, R.M. BioExcel Building Blocks, a Software Library for Interoperable Biomolecular Simulation Workflows. Sci. Data 2019, 6, 169. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (μg/mL) |
|---|---|
| 1-acetyl-β-carboline (1) | 9.71 |
| 1H-indole-3-carbaldehyde (2) | - |
| Anthranilic acid (3) | 82.06 |
| Indole-3-carboxylic acid (4) | 81.49 |
| Source | NO. | Term Name | Padj * |
|---|---|---|---|
| GO MF | 1. | Catalytic activity | 3.961 × 10−51 |
| 2. | Oxidoreductase activity | 1.328 × 10−25 | |
| 3. | Catalytic activity, acting on a protein | 3.970 × 10−23 | |
| GO BP | 1. | Response to chemical | 3.154 × 10−52 |
| 2. | Response to organic substance | 2.615 × 10−49 | |
| 3. | Response to lipid | 1.942 × 10−32 | |
| GO CC | 1. | Cytoplasm | 7.661 × 10−21 |
| 2. | Cytosol | 8.938 × 10−14 | |
| 3. | Somatodendritic compartment | 2.459 × 10−13 | |
| KEGG | 1. | Serotonergic synapse | 9.785 × 10−14 |
| 2. | Steroid hormone biosynthesis | 1.024 × 10−11 | |
| 3. | Apoptosis | 5.850 × 10−11 |
| Compounds | CDC25B (4WH7) | PARP1 (4RV6) | PTGS2 (5KIR) |
|---|---|---|---|
| 1-acetyl-β-carboline (1) | 81.89 | 77.49 | 89.21 |
| 1H-indole-3-carbaldehyde (2) | 67.34 | 59.19 | 73.36 |
| Anthranilic acid (3) | 60.44 | 55.44 | 70.56 |
| Indole-3-carboxylic acid (4) | 74.56 | 65.38 | 79.47 |
| Tamiflu * | 84.34 | 86.13 | 91.29 |
| CDC25B (4WH7) | PARP1 (4RV6) | PTGS2 (5KIR) | |
|---|---|---|---|
| Conventional Hydrogen Bond | TYR428, SER549 | ASP770, ARG878 | TYR385, SER530 |
| van der Waals | PRO444, ARG447, ARG479, MET483, PHE543, ARG544, ARG548, TRP550 | VAL773, GLN875, ILE879, ALA880, PRO881, TYR889 | LEU352, TYR355, LEU359, LEU384, TRP387, SER353, LEU531, PHE518 |
| Amide-Pi Stacking | N.A. | N.A. | GLY526 |
| Pi-Alkyl | LEU445 | ARG878 | VAL349, VAL523, ALA527 |
| Pi-anion | GLU446, ARG482 | ASP766, ARG878 | N.A. |
| Pi-Donor Hydrogen Bond | ARG548 | N.A. | N.A. |
| Pi-Sigma | THR547 | LEU769 | N.A. |
| Pi-Sulfur | N.A. | N.A. | MET522 |
| Pi-cation | GLU446, ARG482 | ASP766, ARG878 | N.A. |
| CDC25B (4WH7) | PARP1 (4RV6) | PTGS2 (5KIR) | |
|---|---|---|---|
| Conventional Hydrogen Bond | LYS399, MET505 | ILE879, ARG878 | SER119 |
| Carbon Hydrogen Bond | ASP397 | ARG878 | PRO86, ARG120 |
| van der Waals | GLU377, LEU378, ILE379, GLY380, PHE386, CYS484, ARG492, PRO503, GLU504, TYR506, ILE507 | LYS703, GLN707, ASP766, ASP770, GLN875, ALA880, GLU883 | LYS83, PRO84, VAL89, LEU93, TYR115, VAL116, TYR355, PHE470, ARG513, GLU524 |
| Alkyl | TYR382, LEU398, ARG485, ARG488 | ILE706, TYR710, LEU769, VAL773, PRO881. | HIS90, LEU123, MET471, VAL523. |
| Pi-Alkyl | TYR382, LEU398, ARG485, ARG488 | ILE706, TYR710, LEU769, VAL773, PRO881 | HIS90, LEU123, MET471, VAL523 |
| Compound | *CID | Structure |
|---|---|---|
| 1-acetyl-beta-carboline | 638667 |  |
| 1H-indole-3-carbaldehyde | 10256 |  |
| Anthranilic acid | 227 | 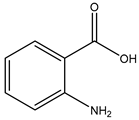 |
| Indole-3-carboxylic acid | 69867 | 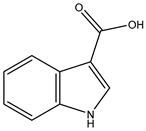 |
| Tamiflu | 78000 | 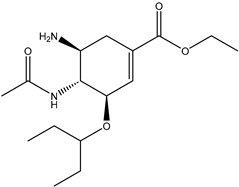 |
| Name | Binding Spheres (X, Y, Z) | Radius | ||
|---|---|---|---|---|
| X | Y | Z | ||
| CDC25B (4WH7) | 9.64 | −8.13 | −10.89 | 14 |
| PARP1 (4RV6) | 91.90 | −16.98 | 134.36 | 14 |
| PTGS2 (5KIR) | 18.43 | 10.98 | 33.81 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Hsieh, C.-Y.; Chiou, C.-T.; Caro, E.J.G.V.; Tayo, L.L.; Tsai, P.-W. In Vitro and In Silico Studies on the Anti-H1N1 Activity of Bioactive Compounds from Marine-Derived Streptomyces ardesiacus. Mar. Drugs 2025, 23, 149. https://doi.org/10.3390/md23040149
Chen Y-H, Hsieh C-Y, Chiou C-T, Caro EJGV, Tayo LL, Tsai P-W. In Vitro and In Silico Studies on the Anti-H1N1 Activity of Bioactive Compounds from Marine-Derived Streptomyces ardesiacus. Marine Drugs. 2025; 23(4):149. https://doi.org/10.3390/md23040149
Chicago/Turabian StyleChen, Yung-Husan, Cheng-Yang Hsieh, Chun-Tang Chiou, Engelo John Gabriel V. Caro, Lemmuel L. Tayo, and Po-Wei Tsai. 2025. "In Vitro and In Silico Studies on the Anti-H1N1 Activity of Bioactive Compounds from Marine-Derived Streptomyces ardesiacus" Marine Drugs 23, no. 4: 149. https://doi.org/10.3390/md23040149
APA StyleChen, Y.-H., Hsieh, C.-Y., Chiou, C.-T., Caro, E. J. G. V., Tayo, L. L., & Tsai, P.-W. (2025). In Vitro and In Silico Studies on the Anti-H1N1 Activity of Bioactive Compounds from Marine-Derived Streptomyces ardesiacus. Marine Drugs, 23(4), 149. https://doi.org/10.3390/md23040149