
Anthraquinone Derivatives and Other Aromatic Compounds from Marine Fungus Asteromyces cruciatus KMM 4696 and Their Effects against Staphylococcus aureus

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
2.1. Isolated Compounds from Asteromyces cruciatus
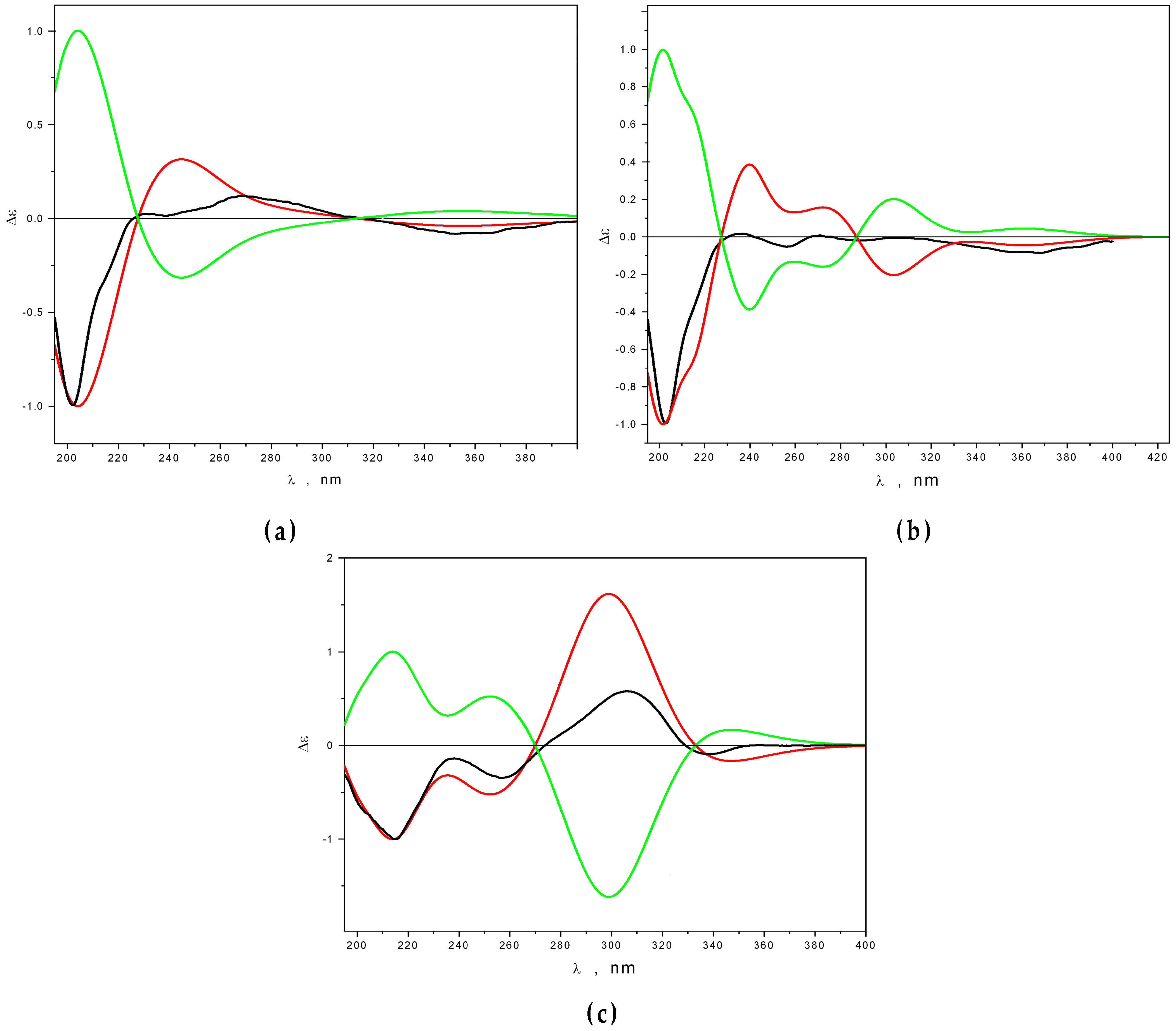
2.2. Structural Characterization of New Compounds
2.3. Bioactivity of Isolated Compounds
2.3.1. Antimicrobial Activity
2.3.2. Influence of Some Isolated Compounds on Urease Activity
2.3.3. Influence of Some Isolated Compounds on Sortase A Activity
2.3.4. Effects of Compounds on HaCaT Keratinocytes Infected with Staphylococcus aureus
3. Discussion
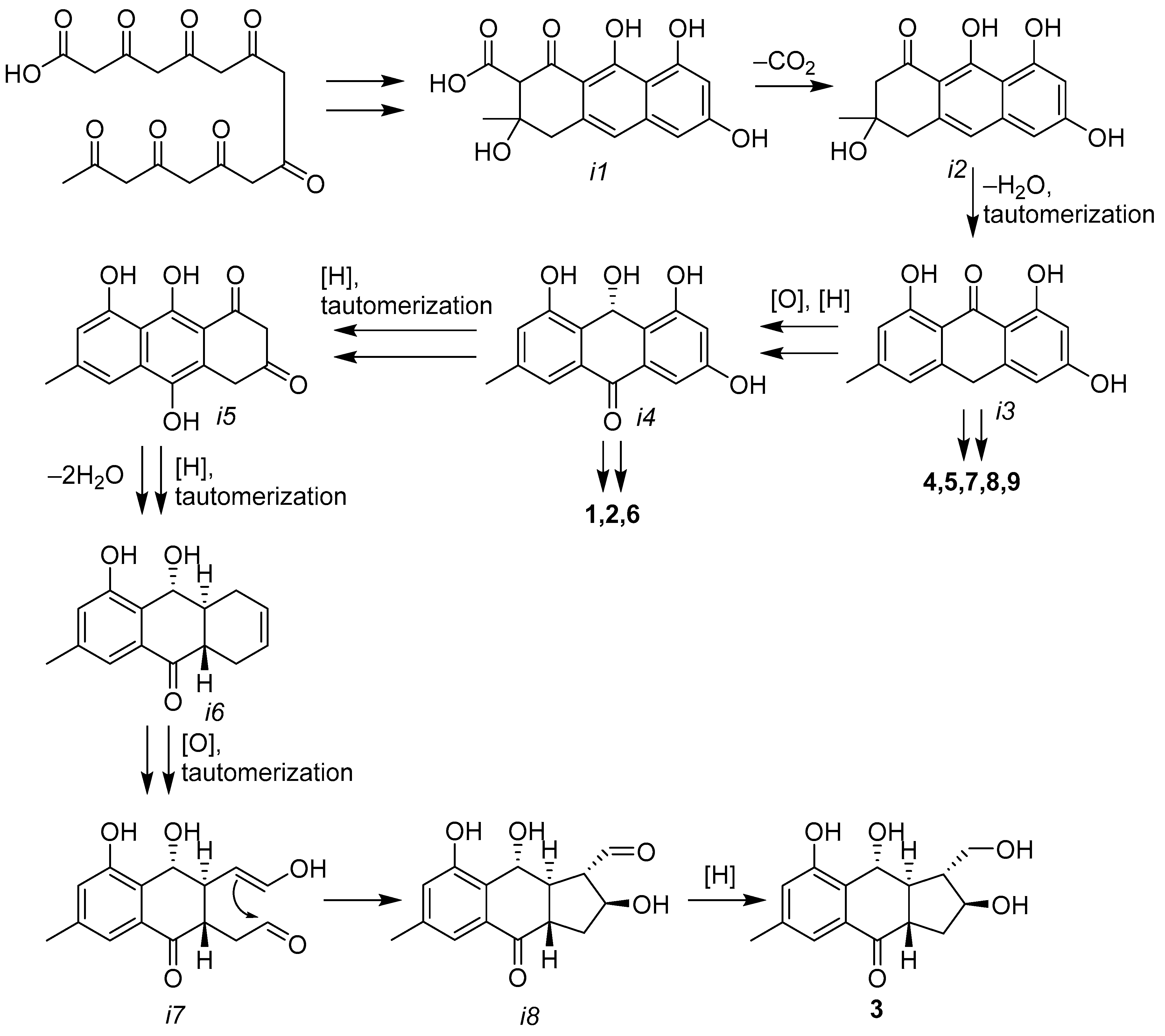
3.1. Secondary Metabolites of Asteromyces cruciatus KMM4696
3.2. Biological Activity of Isolated Anthraquinone Derivatives
3.3. Perspectives of Isolated Anthraquinones for the Treatment of Skin Infections
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Fungal Strain
4.3. Cultivation of Fungus
4.4. Extraction and Isolation
4.5. Spectral Data
4.6. Quantum Chemical Calculations
4.7. Sortase A Activity Inhibition Assay
4.8. Urease Inhibition Assay
4.9. Antimicrobial Activity
4.10. Cell Line and Culture Conditions
4.11. Cocultivation of HaCaT Cells with S. aureus and Lactate Dehydrigenase Release Test
4.12. Migration of HaCaT Cells Cocultivated with S. aureus
4.13. Proliferation of HaCaT Cells Cocultivated with S. aureus
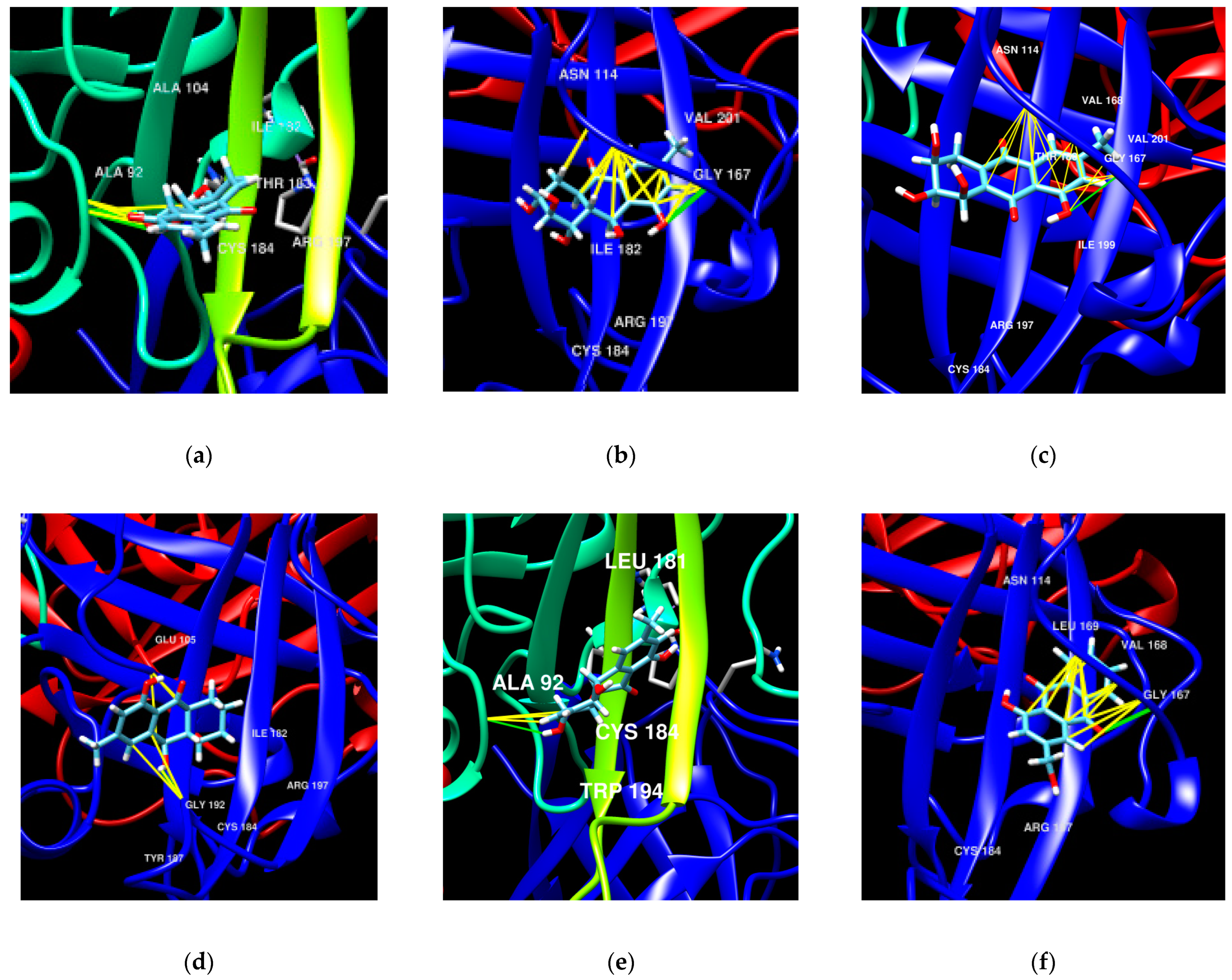
4.14. Molecular Docking
4.15. Statistical Data Evaluation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghoran, S.H.; Taktaz, F.; Ayatollahi, S.A.; Kijjoa, A. Anthraquinones and Their Analogues from Marine-Derived Fungi: Chemistry and Biological Activities. Mar. Drugs 2022, 20, 474. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Evidente, A. Fungal Bioactive Anthraquinones and Analogues. Toxins 2020, 12, 714. [Google Scholar] [CrossRef] [PubMed]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed]
- Abril, A.G.; Villa, T.G.; Barros-Velázquez, J.; Cañas, B.; Sánchez-Pérez, A.; Calo-Mata, P.; Carrera, M. Staphylococcus aureus Exotoxins and Their Detection in the Dairy Industry and Mastitis. Toxins 2020, 12, 537. [Google Scholar] [CrossRef] [PubMed]
- Abbafati, C.; Machado, D.B.; Cislaghi, B.; Salman, O.M.; Karanikolos, M.; McKee, M.; Abbas, K.M.; Brady, O.J.; Larson, H.J.; Trias-Llimós, S.; et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Zhuravleva, O.I.; Oleinikova, G.K.; Antonov, A.S.; Kirichuk, N.N.; Pelageev, D.N.; Rasin, A.B.; Menshov, A.S.; Popov, R.S.; Kim, N.Y.; Chingizova, E.A.; et al. New Antibacterial Chloro-Containing Polyketides from the Alga-Derived Fungus Asteromyces cruciatus KMM 4696. J. Fungi 2022, 8, 454. [Google Scholar] [CrossRef]
- Tanaka, M.; Ohra, J.; Tsujtno, Y.; Fujimori, T.; Ago, H.; Tsuge, H. Dendryol A, B, C, and D, Four New Compounds Produced by a Weed Pathogenic Fungus Dendryphiella sp. Z. Naturforschung C 1995, 50, 751–756. [Google Scholar] [CrossRef]
- Zhang, C.; Ondeyka, J.G.; Zink, D.L.; Basilio, A.; Vicente, F.; Collado, J.; Platas, G.; Huber, J.; Dorso, K.; Motyl, M.; et al. Isolation, structure and antibacterial activity of pleosporone from a pleosporalean ascomycete discovered by using antisense strategy. Bioorg. Med. Chem. 2009, 17, 2162–2166. [Google Scholar] [CrossRef]
- Sun, P.; Huo, J.; Kurtan, T.; Mandi, A.; Antus, S.; Tang, H.; Draeger, S.; Schulz, B.; Hussain, H.; Krohn, K.; et al. Structural and stereochemical studies of hydroxyanthraquinone derivatives from the endophytic fungus Coniothyrium sp. Chirality 2013, 25, 141–148. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, A.; Chen, H.; Wang, M.; Ding, G.; Sun, L.; Li, L.; Dai, M. Anthraquinones from the saline-alkali plant endophytic fungus Eurotium rubrum. J. Antibiot. 2017, 70, 1138–1141. [Google Scholar] [CrossRef]
- Imre, S.; Sar, S.; Thomson, R.H. Anthraquinones in Digitalis species. Phytochemistry 1976, 15, 317. [Google Scholar] [CrossRef]
- Arrebola, M.L.; Ringbom, T.; Verpoorte, R. Anthraquinones from Isoplexis isabelliana cell suspension cultures. Phytochemistry 1999, 52, 1283–1286. [Google Scholar] [CrossRef]
- Abraham, W.R.; Arfmann, H.A. Hydroxy-(methylbutenynyl)-benzoic acid and derivatives from Curvularia fallax. Phytochemistry 1990, 29, 2641. [Google Scholar] [CrossRef]
- Prompanya, C.; Dethoup, T.; Gales, L.; Lee, M.; Pereira, J.A.C.; Silva, A.M.S.; Pinto, M.M.M.; Kijjoa, A. New Polyketides and New Benzoic Acid Derivatives from the Marine Sponge-Associated Fungus Neosartorya quadricincta KUFA 0081. Mar. Drugs 2016, 14, 134. [Google Scholar] [CrossRef] [PubMed]
- Golyshin, P.N.; Golyshina, O.V.; Timmis, K.N.; Chernikova, T.; Waliczek, A.; Ferrer, M.; Beloqui, A.; Guazzaroni, M.E.; Vieites, J.M.; Pazos, F.; et al. Preparation of fluorescent probe-based reactome array for detection, immobilization and isolation of enzymes. Patent EP 2230312A1, 22 September 2010. [Google Scholar]
- Shin, J.; Fenical, W. Isolation of gliovictin from the marine deuteromycete Asteromyces cruciatus. Phytochemistry 1987, 26, 3347. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16 Rev. A.03; Scientific Research Publishing: Wallingford, CT, USA, 2016. [Google Scholar]
- Zhou, C.; Bhinderwala, F.; Lehman, M.K.; Thomas, V.C.; Chaudhari, S.S.; Yamada, K.J.; Foster, K.W.; Powers, R.; Kielian, T.; Fey, P.D. Urease is an essential component of the acid response network of Staphylococcus aureus and is required for a persistent murine kidney infection. PLOS Pathog. 2019, 15, e1007538. [Google Scholar] [CrossRef]
- Sapra, R.; Rajora, A.K.; Kumar, P.; Maurya, G.P.; Pant, N.; Haridas, V. Chemical Biology of Sortase A Inhibition: A Gateway to Anti-infective Therapeutic Agents. J. Med. Chem. 2021, 64, 13097–13130. [Google Scholar] [CrossRef]
- Zong, Y.; Bice, T.W.; Ton-That, H.; Schneewind, O.; Narayana, S.V.L. Crystal Structures of Staphylococcus aureus Sortase A and Its Substrate Complex. J. Biol. Chem. 2004, 279, 31383–31389. [Google Scholar] [CrossRef]
- Thappeta, K.R.; Zhao, L.N.; Nge, C.E.; Crasta, S.; Leong, C.Y.; Ng, V.; Kanagasundaram, Y.; Fan, H.; Ng, S.B. In-Silico Identified New Natural Sortase A Inhibitors Disrupt S. aureus Biofilm Formation. Int. J. Mol. Sci. 2020, 21, 8601. [Google Scholar] [CrossRef]
- Wiegand, C.; Abel, M.; Ruth, P.; Hipler, U.C. HaCaT keratinocytes in co-culture with Staphylococcus aureus can be protected from bacterial damage by polihexanide. Wound Repair Regen. 2009, 17, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Girich, E.V.; Trinh, P.T.; Nesterenko, L.E.; Popov, R.S.; Kim, N.Y.; Rasin, A.B.; Menchinskaya, E.S.; Kuzmich, A.S.; Chingizova, E.A.; Minin, A.S.; et al. Absolute Stereochemistry and Cytotoxic Effects of Vismione E from Marine Sponge-Derived Fungus Aspergillus sp. 1901NT-1.2.2. Int. J. Mol. Sci. 2023, 24, 8150. [Google Scholar] [CrossRef] [PubMed]
- Sebak, M.; Molham, F.; Greco, C.; Tammam, M.A.; Sobeh, M.; El-Demerdash, A. Chemical diversity, medicinal potentialities, biosynthesis, and pharmacokinetics of anthraquinones and their congeners derived from marine fungi: A comprehensive update. RSC Adv. 2022, 12, 24887–24921. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, S.; Mesarich, C.H.; Saccomanno, B.; Vaisberg, A.; De Wit, P.J.; Cox, R.; Collemare, J. Elucidation of cladofulvin biosynthesis reveals a cytochrome P450 monooxygenase required for anthraquinone dimerization. Proc. Natl. Acad. Sci. USA 2016, 113, 6851–6856. [Google Scholar] [CrossRef]
- Chapla, V.M.; Honório, A.E.; Gubiani, J.R.; Vilela, A.F.L.; Young, M.C.M.; Cardoso, C.L.; Pavan, F.R.; Cicarelli, R.M.; Ferreira, P.M.P.; Bolzani, V.d.S.; et al. Acetylcholinesterase inhibition and antifungal activity of cyclohexanoids from the endophytic fungus Saccharicola sp. Phytochem. Lett. 2020, 39, 116–123. [Google Scholar] [CrossRef]
- Gulder, T.A.M.; Hong, H.; Correa, J.; Egereva, E.; Wiese, J.; Imhoff, J.F.; Gross, H. Isolation, Structure Elucidation and Total Synthesis of Lajollamide A from the Marine Fungus Asteromyces cruciatus. Mar. Drugs 2012, 10, 2912–2935. [Google Scholar] [CrossRef]
- Li, H.L.; Li, X.M.; Li, X.; Wang, C.Y.; Liu, H.; Kassack, M.U.; Meng, L.H.; Wang, B.G. Antioxidant Hydroanthraquinones from the Marine Algal-Derived Endophytic Fungus Talaromyces islandicus EN-501. J. Nat. Prod. 2017, 80, 162–168. [Google Scholar] [CrossRef]
- Mohanlall, V.; Odhav, B. Antibacterial, anti-inflammatory and antioxidant activities of anthraquinones from Ceratotheca triloba (Bernh) Hook F. J. Med. Plant Res. 2013, 7, 877–886. [Google Scholar]
- Chingizova, E.A.; Menchinskaya, E.S.; Chingizov, A.R.; Pislyagin, E.A.; Girich, E.V.; Yurchenko, A.N.; Guzhova, I.V.; Mikhailov, V.V.; Aminin, D.L.; Yurchenko, E.A. Marine Fungal Cerebroside Flavuside B Protects HaCaT Keratinocytes against Staphylococcus aureus Induced Damage. Mar. Drugs 2021, 19, 553. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef]
- WHO. Ten Threats to Global Health in 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Cascioferro, S.; Totsika, M.; Schillaci, D. Sortase A: An ideal target for anti-virulence drug development. Microb. Pathog. 2014, 77, 105–112. [Google Scholar] [CrossRef]
- Nitulescu, G.; Margina, D.; Zanfirescu, A.; Olaru, O.T.; Nitulescu, G.M. Targeting bacterial sortases in search of anti-virulence therapies with low risk of resistance development. Pharmaceuticals 2021, 14, 415. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.W.; Yi, S.W.; Paek, S.-M. Design and Synthesis of Small Molecules as Potent Staphylococcus aureus Sortase A Inhibitors. Antibiotics 2020, 9, 706. [Google Scholar] [CrossRef]
- Hameed, A.; Al-Rashida, M.; Uroos, M.; Qazi, S.U.; Naz, S.; Ishtiaq, M.; Khan, K.M. A patent update on therapeutic applications of urease inhibitors (2012–2018). Expert Opin. Ther. Pat. 2019, 29, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Kafarski, P.; Talma, M. Recent advances in design of new urease inhibitors: A review. J. Adv. Res. 2018, 13, 101–112. [Google Scholar] [CrossRef]
- Gottlieb, H.E.; Kotlyar, V.; Nudelman, A. NMR chemical shifts of common laboratory solvents as trace impurities. J. Org. Chem. 1997, 62, 7512–7515. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Brooks, C.L., III; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [PubMed]
- Haberthür, U.; Caflisch, A. FACTS: Fast analytical continuum treatment of solvation. J. Comput. Chem. 2008, 29, 701–715. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. Fast docking using the CHARMM force field with EADock DSS. J. Comput. Chem. 2011, 32, 2149–2159. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | 2 | ||
|---|---|---|---|---|
| δC, mult | δH (J in Hz) a | δC, mult | δH (J in Hz) b | |
| 1 | 78.9, CH | 3.99, t (9.0) | 77.9, CH | 4.09, t (9.5) |
| 2 | 70.1, CH | 3.81, m | 77.8, CH | 3.61, dd (9.6, 8.4) |
| 3 | 34.7, CH2 | α: 2.76, m β: 2.30, m | 72.8, CH | 4.30, m |
| 4 | 137.1, CH | 6.98, m | 139.6, CH | 6.85, t (2.7) |
| 5 | 120.1, CH | 7.35, d (1.3) | 120.1, CH | 7.36, d (1.2) |
| 6 | 140.0, C | - | 140.1, C | - |
| 7 | 123.3, CH | 6.87, d (1.3) | 123.6, CH | 6.89, d (1.2) |
| 8 | 157.4, C | 157.6, C | ||
| 9 | 75.0, CH | 5.37, d (9.8) | 74.9, CH | 5.39, d (9.9) |
| 10 | 184.1, C | - | 184.3, C | - |
| 11 | 132.9, C | - | 132.8, C | - |
| 12 | 125.5, C | - | 125.5, C | - |
| 13 | 48.5, CH | 2.96, m | 48.6, CH | 3.02, m |
| 14 | 132.2, C | - | 130.9, C | - |
| 15 | 21.0, CH3 | 2.30, s | 21.0, CH3 | 2.31, s |
| Pos. | δC, Mult | δH (J in Hz) |
|---|---|---|
| 1 | 54.4, CH | 2.14, m |
| 2 | 62.9, CH2 | 3.76, t (10.3) 4.13, dd (10.7, 2.8) |
| 3 | 73.1, CH | 4.47, brs |
| 4 | 35.0, CH2 | α: 1.98, ddd (14.3, 9.5, 2.9) β: 2.35, m |
| 5 | 118.9, CH | 7.25, s |
| 6 | 139.7, C | - |
| 7 | 122.7, CH | 6.86, s |
| 8 | 158.3, C | - |
| 9 | 74.5, CH | 5.07, d (9.4) |
| 10 | 197.8, C | - |
| 11 | 134.7, C | - |
| 12 | 127.8, C | - |
| 13 | 54.7, CH | 2.37, m |
| 14 | 51.4, CH | 2.69, dt (13.6, 9.2) |
| 15 | 21.0, CH3 | 2.31, s |
| 2-OH | - | 5.31, brs |
| 3-OH | - | 3.82, d (4.7) |
| 8-OH | - | 9.36, brs |
| 9-OH | - | 7.31, brs |
| Compound | 3 | 8 | 9 | 10 | 12 | 13 |
|---|---|---|---|---|---|---|
| IC50, µM | ≥100 | 35.4 ± 1.5 | 45.3 ± 3.1 | 49.7 ± 2.4 | 52.1 ± 1.3 | 58.2 ± 1.7 |
| Sample 1 | Division 0, % of Total Amount | Division 1, % of Total Amount | Division 2, % of Total Amount | Division 3, % of Total Amount |
|---|---|---|---|---|
| HaCaT cells | 3.0 ± 0.8 | 37.7 ± 3.0 | 43.2 ± 1.4 | 15.5 ± 3.7 |
| S. aureus | 8.7 ± 1.2 | 58.2 ± 2.0 | 16.8 ± 6.0 | 14.9 ± 1.5 |
| 1 | 3.6 ± 0.5 | 34.0 ± 3.3 | 25.9 ± 0.4 | 35.5 ± 4.2 |
| 4 | 3.1 ± 0.4 | 63.2 ± 2.3 | 15.4 ± 1.9 | 17.6 ± 1.7 |
| 6 | 8.6 ± 4.7 | 60.5 ± 3.4 | 12.1 ± 1.2 | 18.1 ± 2.0 |
| 7 | 7.2 ± 2.1 | 65.5 ± 6.4 | 11.6 ± 1.3 | 15.4 ± 5.6 |
| 9 | 3.4 ± 1.2 | 46.3 ± 7.2 | 18.3 ± 6.5 | 30.2 ± 4.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuravleva, O.I.; Chingizova, E.A.; Oleinikova, G.K.; Starnovskaya, S.S.; Antonov, A.S.; Kirichuk, N.N.; Menshov, A.S.; Popov, R.S.; Kim, N.Y.; Berdyshev, D.V.; et al. Anthraquinone Derivatives and Other Aromatic Compounds from Marine Fungus Asteromyces cruciatus KMM 4696 and Their Effects against Staphylococcus aureus. Mar. Drugs 2023, 21, 431. https://doi.org/10.3390/md21080431
Zhuravleva OI, Chingizova EA, Oleinikova GK, Starnovskaya SS, Antonov AS, Kirichuk NN, Menshov AS, Popov RS, Kim NY, Berdyshev DV, et al. Anthraquinone Derivatives and Other Aromatic Compounds from Marine Fungus Asteromyces cruciatus KMM 4696 and Their Effects against Staphylococcus aureus. Marine Drugs. 2023; 21(8):431. https://doi.org/10.3390/md21080431
Chicago/Turabian StyleZhuravleva, Olesya I., Ekaterina A. Chingizova, Galina K. Oleinikova, Sofya S. Starnovskaya, Alexandr S. Antonov, Natalia N. Kirichuk, Alexander S. Menshov, Roman S. Popov, Natalya Yu. Kim, Dmitrii V. Berdyshev, and et al. 2023. "Anthraquinone Derivatives and Other Aromatic Compounds from Marine Fungus Asteromyces cruciatus KMM 4696 and Their Effects against Staphylococcus aureus" Marine Drugs 21, no. 8: 431. https://doi.org/10.3390/md21080431
APA StyleZhuravleva, O. I., Chingizova, E. A., Oleinikova, G. K., Starnovskaya, S. S., Antonov, A. S., Kirichuk, N. N., Menshov, A. S., Popov, R. S., Kim, N. Y., Berdyshev, D. V., Chingizov, A. R., Kuzmich, A. S., Guzhova, I. V., Yurchenko, A. N., & Yurchenko, E. A. (2023). Anthraquinone Derivatives and Other Aromatic Compounds from Marine Fungus Asteromyces cruciatus KMM 4696 and Their Effects against Staphylococcus aureus. Marine Drugs, 21(8), 431. https://doi.org/10.3390/md21080431