
Historical Perspective of the Characterization of Conotoxins Targeting Voltage-Gated Sodium Channels

Abstract
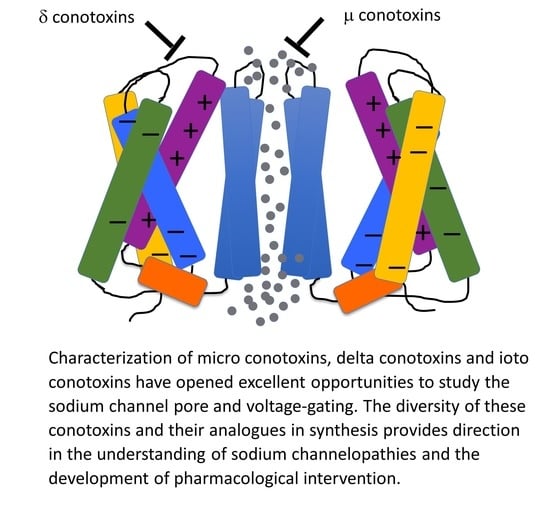
1. An Overview of Bioactive Marine Toxins
2. Toxins Targeting Voltage-Gated Sodium Channels
3. Conotoxins Targeting Voltage-Gated Sodium Channels
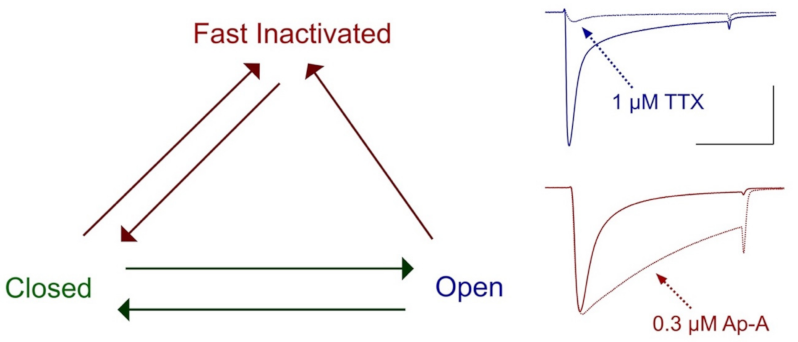
4. Isolation and Initial Characterization of Actions of μ-Conotoxins on Nerve and Muscle
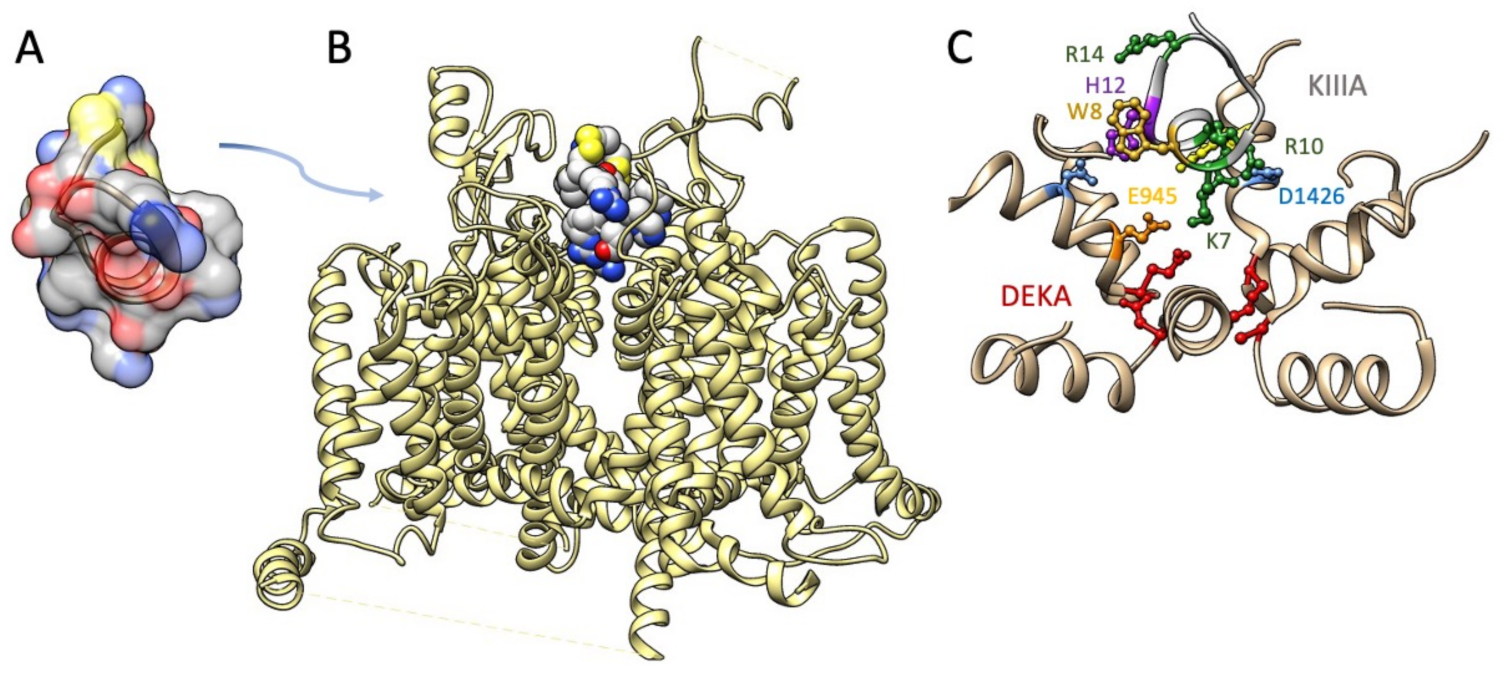
5. Isolation and Characterization of μ-Conotoxins Targeting Neuronal Sodium Channels
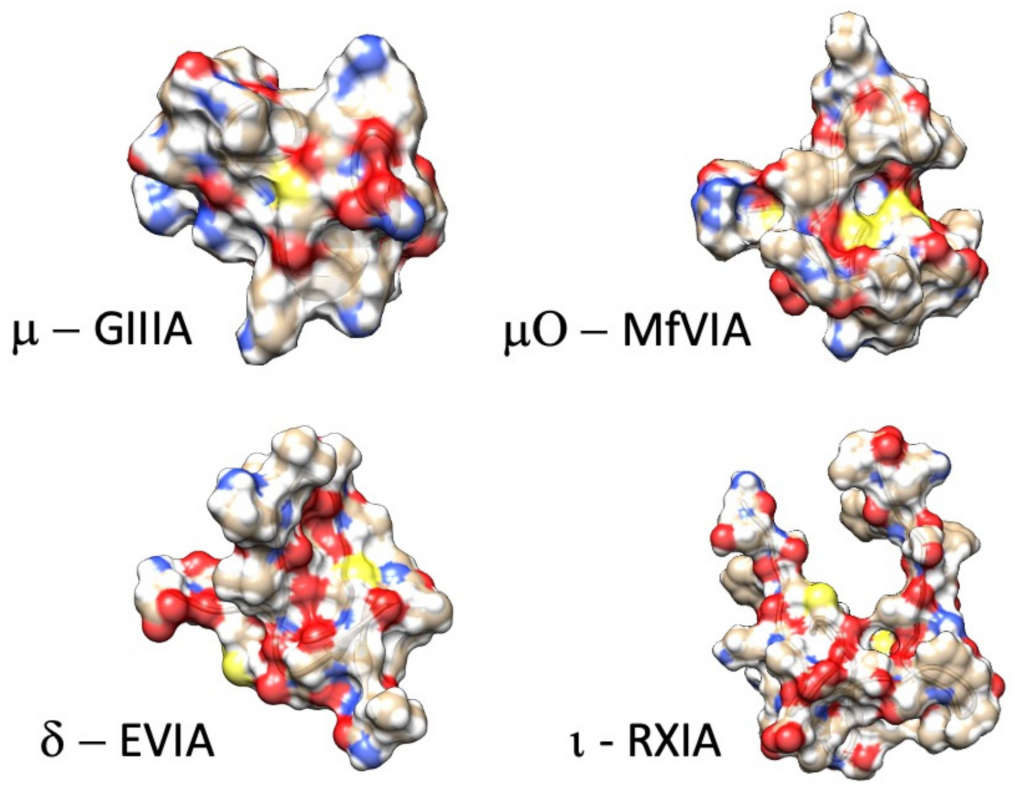
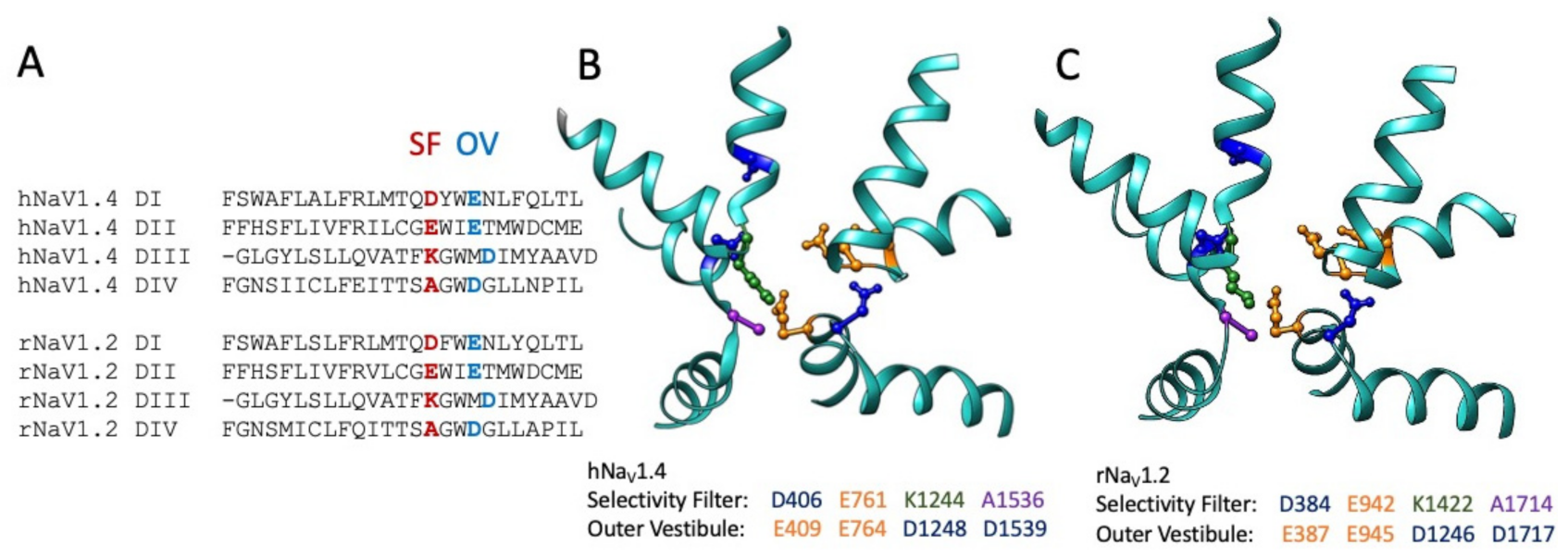
6. Structural Divergence in M4 and M5 Conotoxins Affecting Channel Selectivity
7. Isolation and Characterization of μO-Conotoxins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Conus Species | Sequence | References |
|---|---|---|---|
| μ-O | ---C------C-------------CC----C----C--- | ||
| MrVIA | C. marmoreus | --ACRKKWEYC----IVPIIGFIYCCPGLICGPFVCV---- | [159] |
| MrVIB | C. marmoreus | --ACSKKWEYC----IVPILGFVYCCPGLICGPFVCV---- | [159] |
| MfVIA | C. magnificus | -RDCQEKWEYC----IVPILGFVYCCPGLICGPFVCV---- | [196] |
| δ | |||
| Tx(V)IA | C. textile | --WCKQSGEMCN---LLD--QN--CCDGY-CIVLVCT---- | [197,198,199] |
| TxIB | C. textile | --WCKQSGEMCN---VLD--QN--CCDGY-CIVFVCT---- | [198] |
| GmVIA | C. gloriamus | VKPCRKEGQLCD-----PIFQN--CCRGWNCV-LFCV---- | [200] |
| NgVIA | C. nigropunctatus | -SKCFSOGTFCG---IKO--GL--CCSVR-CFSLFCISFE- | [201] |
| PVIA | C. purpurascens | -EACYAOGTFCG---IKO--GL--CCSEF-CLPGVCFG--- | [202] |
| SVIE | C. striatus | -DGCSSGGTFCG---IHO--GL--CCSEF-CF-LWCITFID | [203] |
| CnVIA | C. consors | -YECYSTGTFCG---ING--GL--CCSNL-CLFFVCLTFS | [203] |
| Am2766 | C. amadis | ---CKQAGESCD---IFS--QN--CCVGT-CA-FICIE--- | [204] |
| EVIA | C. ermineus | -DDCIKOYGFCSLPILKN--GL--CCSGA-CV-GVCADL-- | [205] |
| CnVIB | C. consors | -DECFSOGTFCG---TKO--GL--CCSAR-CFSFFCISLEF | [206] |
| CnVIC | C. consors | -DECFSOGTFCG---IKO--GL--CCSAR-CLSFFCISLEF | [206] |
| CnVID | C. consors | -DECFSOGTFCG---FKO--GL--CCSAR-CFSLFCISLEF | [206] |
| TsVIA | C. tessulatus | ---CAAFGSFCG---L-P--GLVDCCSGR-CF-IVCLL--- | [207] |
| SuVIA | C. suturatus | ---CAGIGSFCG---L-P--GLVDCCSDR-CF-IVCLP--- | [2] |
8. Delta (δ-)Conotoxins
9. Isolation and Characterization of Iota (ι)-Conotoxins
10. Summary
Funding
Data Availability Statement
Conflicts of Interest
References
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex Cocktails: The Evolutionary Novelty of Venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.-H.; Israel, M.R.; Inserra, M.C.; Smith, J.J.; Lewis, R.J.; Alewood, P.F.; Vetter, I.; Dutertre, S. δ-Conotoxin SuVIA Suggests an Evolutionary Link between Ancestral Predator Defence and the Origin of Fish-Hunting Behaviour in Carnivorous Cone Snails. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150817. [Google Scholar] [CrossRef] [PubMed]
- Jost, M.C.; Hillis, D.M.; Lu, Y.; Kyle, J.W.; Fozzard, H.A.; Zakon, H.H. Toxin-Resistant Sodium Channels: Parallel Adaptive Evolution across a Complete Gene Family. Mol. Biol. Evol. 2008, 25, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Brodie, E.D., III; Brodie, E.D., Jr. Predictably Convergent Evolution of Sodium Channels in the Arms Race between Predators and Prey. Brain. Behav. Evol. 2015, 86, 48–57. [Google Scholar] [CrossRef]
- Feldman, C.R.; Durso, A.M.; Hanifin, C.T.; Pfrender, M.E.; Ducey, P.K.; Stokes, A.N.; Barnett, K.E.; Brodie, E.D.; Brodie, E.D. Is There More than One Way to Skin a Newt? Convergent Toxin Resistance in Snakes Is Not Due to a Common Genetic Mechanism. Heredity 2017, 119, 468. [Google Scholar] [CrossRef]
- Geffeney, S.L.; Williams, B.L.; Rosenthal, J.J.C.; Birk, M.A.; Felkins, J.; Wisell, C.M.; Curry, E.R.; Hanifin, C.T. Convergent and Parallel Evolution in a Voltage-Gated Sodium Channel Underlies TTX-Resistance in the Greater Blue-Ringed Octopus: Hapalochlaena Lunulata. Toxicon 2019, 170, 77–84. [Google Scholar] [CrossRef]
- Van Thiel, J.; Khan, M.A.; Wouters, R.M.; Harris, R.J.; Casewell, N.R.; Fry, B.G.; Kini, R.M.; Mackessy, S.P.; Vonk, F.J.; Wüster, W.; et al. Convergent Evolution of Toxin Resistance in Animals. Biol. Rev. 2022, 97, 1823–1843. [Google Scholar] [CrossRef]
- Olivera, B.M.; Seger, J.; Horvath, M.P.; Fedosov, A.E. Prey-Capture Strategies of Fish-Hunting Cone Snails: Behavior, Neurobiology and Evolution. Brain. Behav. Evol. 2015, 86, 58–74. [Google Scholar] [CrossRef]
- Olivera, B.M.; Showers Corneli, P.; Watkins, M.; Fedosov, A. Biodiversity of Cone Snails and Other Venomous Marine Gastropods: Evolutionary Success Through Neuropharmacology. Annu. Rev. Anim. Biosci. 2014, 2, 487–513. [Google Scholar] [CrossRef]
- Olivera, B.M.; Raghuraman, S.; Schmidt, E.W.; Safavi-Hemami, H. Linking Neuroethology to the Chemical Biology of Natural Products: Interactions between Cone Snails and Their Fish Prey, a Case Study. J. Comp. Physiol. A 2017, 203, 717–735. [Google Scholar] [CrossRef]
- Olivera, B.M.; Walker, C.; Cartier, G.E.; Hooper, D.; Santos, A.D.; Schoenfeld, R.; Shetty, R.; Watkins, M.; Bandyopadhyay, P.; Hillyard, D.R. Speciation of Cone Snails and Interspecific Hyperdivergence of Their Venom Peptides: Potential Evolutionary Significance of Intronsa. Ann. N. Y. Acad. Sci. 1999, 870, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Kapil, S.; Hendriksen, S.; Cooper, J.S. Cone Snail Toxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Yu, C.; Yu, H.; Li, P. Highlights of Animal Venom Research on the Geographical Variations of Toxin Components, Toxicities and Envenomation Therapy. Int. J. Biol. Macromol. 2020, 165, 2994–3006. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.-H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and Biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Norton, R. Conotoxin Gene Superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef]
- Phuong, M.A.; Mahardika, G.N. Targeted Sequencing of Venom Genes from Cone Snail Genomes Improves Understanding of Conotoxin Molecular Evolution. Mol. Biol. Evol. 2018, 35, 1210–1224. [Google Scholar] [CrossRef]
- Benoit, E. Mechanism of action of neurotoxins acting on the inactivation of voltage-gated sodium channels. Comptes Rendus Seances Soc. Biol. Fil. 1998, 192, 409–436. [Google Scholar]
- Terlau, H.; Olivera, B.M. Conus Venoms: A Rich Source of Novel Ion Channel-Targeted Peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus Venom Peptide Pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef]
- Green, B.R.; Olivera, B.M. Venom Peptides From Cone Snails. In Current Topics in Membranes; Elsevier: Amsterdam, The Netherlands, 2016; Volume 78, pp. 65–86. ISBN 978-0-12-805386-7. [Google Scholar]
- Duque, H.M.; Campos Dias, S.; Franco, O. Structural and Functional Analyses of Cone Snail Toxins. Mar. Drugs 2019, 17, 370. [Google Scholar] [CrossRef]
- Mackieh, R.; Abou-Nader, R.; Wehbe, R.; Mattei, C.; Legros, C.; Fajloun, Z.; Sabatier, J.M. Voltage-Gated Sodium Channels: A Prominent Target of Marine Toxins. Mar. Drugs 2021, 19, 562. [Google Scholar] [CrossRef]
- Daly, N.L.; Craik, D.J. Structural Studies of Conotoxins. IUBMB Life 2009, 61, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Zhorov, B.S.; Tikhonov, D.B. Computational Structural Pharmacology and Toxicology of Voltage-Gated Sodium Channels. In Current Topics in Membranes; Elsevier: Amsterdam, The Netherlands, 2016; Volume 78, pp. 117–144. ISBN 978-0-12-805386-7. [Google Scholar]
- Tikhonov, D.B.; Zhorov, B.S. Predicting Structural Details of the Sodium Channel Pore Basing on Animal Toxin Studies. Front. Pharmacol. 2018, 9, 880. [Google Scholar] [CrossRef] [PubMed]
- Knapp, O.; McArthur, J.R.; Adams, D.J. Conotoxins Targeting Neuronal Voltage-Gated Sodium Channel Subtypes: Potential Analgesics? Toxins 2012, 4, 1236–1260. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Lewis, R.J. Therapeutic Potential of Cone Snail Venom Peptides (Conopeptides). Curr. Top. Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef]
- Munasinghe, N.; Christie, M. Conotoxins That Could Provide Analgesia through Voltage Gated Sodium Channel Inhibition. Toxins 2015, 7, 5386–5407. [Google Scholar] [CrossRef]
- Tosti, E.; Boni, R.; Gallo, A. Μ-Conotoxins Modulating Sodium Currents in Pain Perception and Transmission: A Therapeutic Potential. Mar. Drugs 2017, 15, 295. [Google Scholar] [CrossRef]
- Twede, V.D.; Miljanich, G.; Olivera, B.M.; Bulaj, G. Neuroprotective and Cardioprotective Conopeptides: An Emerging Class of Drug Leads. Curr. Opin. Drug Discov. Devel. 2009, 12, 231–239. [Google Scholar]
- Coulter-Parkhill, A.; McClean, S.; Gault, V.A.; Irwin, N. Therapeutic Potential of Peptides Derived from Animal Venoms: Current Views and Emerging Drugs for Diabetes. Clin. Med. Insights Endocrinol. Diabetes 2021, 14, 117955142110060. [Google Scholar] [CrossRef]
- De Castro Figueiredo Bordon, K.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef]
- Livett, B.G.; Gayler, K.R.; Khalil, Z. Drugs from the Sea: Conopeptides as Potential Therapeutics. Curr. Med. Chem. 2004, 11, 1715–1723. [Google Scholar] [CrossRef]
- Miljanich, G.P. Ziconotide: Neuronal Calcium Channel Blocker for Treating Severe Chronic Pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.; Clark, R. G-Protein Coupled Receptors Targeted by Analgesic Venom Peptides. Toxins 2017, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Pérez de Vega, M.J.; Ferrer-Montiel, A.; González-Muñiz, R. Recent Progress in Non-Opioid Analgesic Peptides. Arch. Biochem. Biophys. 2018, 660, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Brogan, S.E.; Olivera, B.M. Pain Therapeutics from Cone Snail Venoms: From Ziconotide to Novel Non-Opioid Pathways. J. Proteom. 2019, 190, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Jergova, S.; Perez, C.; Imperial, J.S.; Gajavelli, S.; Jain, A.; Abin, A.; Olivera, B.M.; Sagen, J. Cannabinoid Receptor Agonists from Conus Venoms Alleviate Pain-Related Behavior in Rats. Pharmacol. Biochem. Behav. 2021, 205, 173182. [Google Scholar] [CrossRef]
- Han, T.S.; Zhang, M.-M.; Gowd, K.H.; Walewska, A.; Yoshikami, D.; Olivera, B.M.; Bulaj, G. Disulfide-Depleted Selenoconopeptides: Simplified Oxidative Folding of Cysteine-Rich Peptides. ACS Med. Chem. Lett. 2010, 1, 140–144. [Google Scholar] [CrossRef]
- McIntosh, J.M.; Santos, A.D.; Olivera, B.M. Conus Peptides Targeted to Specific Nicotinic Acetylcholine Receptor Subtypes. Annu. Rev. Biochem. 1999, 68, 59–88. [Google Scholar] [CrossRef]
- Nicke, A.; Wonnacott, S.; Lewis, R.J. Alpha-Conotoxins as Tools for the Elucidation of Structure and Function of Neuronal Nicotinic Acetylcholine Receptor Subtypes. Eur. J. Biochem. 2004, 271, 2305–2319. [Google Scholar] [CrossRef]
- Janes, R.W. α-Conotoxins as Selective Probes for Nicotinic Acetylcholine Receptor Subclasses. Curr. Opin. Pharmacol. 2005, 5, 280–292. [Google Scholar] [CrossRef]
- Dutertre, S.; Nicke, A.; Tsetlin, V.I. Nicotinic Acetylcholine Receptor Inhibitors Derived from Snake and Snail Venoms. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef]
- Abraham, N.; Lewis, R. Neuronal Nicotinic Acetylcholine Receptor Modulators from Cone Snails. Mar. Drugs 2018, 16, 208. [Google Scholar] [CrossRef]
- Bekbossynova, A.; Zharylgap, A.; Filchakova, O. Venom-Derived Neurotoxins Targeting Nicotinic Acetylcholine Receptors. Molecules 2021, 26, 3373. [Google Scholar] [CrossRef] [PubMed]
- Kasheverov, I.; Kudryavtsev, D.; Shelukhina, I.; Nikolaev, G.; Utkin, Y.; Tsetlin, V. Marine Origin Ligands of Nicotinic Receptors: Low Molecular Compounds, Peptides and Proteins for Fundamental Research and Practical Applications. Biomolecules 2022, 12, 189. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.; Peigneur, S.; Wijesekara, I.; Tytgat, J. Conotoxins Targeting Nicotinic Acetylcholine Receptors: An Overview. Mar. Drugs 2014, 12, 2970–3004. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.W.; Marquart, L.A.; Phillips, P.D.; McDougal, O.M. Mutagenesis of α-Conotoxins for Enhancing Activity and Selectivity for Nicotinic Acetylcholine Receptors. Toxins 2019, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Adams, D.J.; Hung, A. Interactions of the A3β2 Nicotinic Acetylcholine Receptor Interfaces with α-Conotoxin LsIA and Its Carboxylated C-Terminus Analogue: Molecular Dynamics Simulations. Mar. Drugs 2020, 18, 349. [Google Scholar] [CrossRef]
- Xu, Q.; Tae, H.-S.; Wang, Z.; Jiang, T.; Adams, D.J.; Yu, R. Rational Design of α-Conotoxin RegIIA Analogues Selectively Inhibiting the Human A3β2 Nicotinic Acetylcholine Receptor through Computational Scanning. ACS Chem. Neurosci. 2020, 11, 2804–2811. [Google Scholar] [CrossRef] [PubMed]
- Hone, A.J.; Servent, D.; McIntosh, J.M. A9-containing Nicotinic Acetylcholine Receptors and the Modulation of Pain. Br. J. Pharmacol. 2018, 175, 1915–1927. [Google Scholar] [CrossRef]
- Hone, A.J.; Kaas, Q.; Kearns, I.; Hararah, F.; Gajewiak, J.; Christensen, S.; Craik, D.J.; McIntosh, J.M. Computational and Functional Mapping of Human and Rat A6β4 Nicotinic Acetylcholine Receptors Reveals Species-Specific Ligand-Binding Motifs. J. Med. Chem. 2021, 64, 1685–1700. [Google Scholar] [CrossRef]
- Wu, X.; Craik, D.J.; Kaas, Q. Interactions of Globular and Ribbon [Γ4E]GID with A4β2 Neuronal Nicotinic Acetylcholine Receptor. Mar. Drugs 2021, 19, 482. [Google Scholar] [CrossRef]
- Liang, J.; Tae, H.-S.; Zhao, Z.; Li, X.; Zhang, J.; Chen, S.; Jiang, T.; Adams, D.J.; Yu, R. Mechanism of Action and Structure–Activity Relationship of α-Conotoxin Mr1.1 at the Human A9α10 Nicotinic Acetylcholine Receptor. J. Med. Chem. 2022, 65, 16204–16217. [Google Scholar] [CrossRef] [PubMed]
- Quik, M.; Polonskaya, Y.; Kulak, J.M.; McIntosh, J.M. Vulnerability of 125I-α-Conotoxin MII Binding Sites to Nigrostriatal Damage in Monkey. J. Neurosci. 2001, 21, 5494–5500. [Google Scholar] [CrossRef]
- Quik, M.; Bordia, T.; Forno, L.; McIntosh, J.M. Loss of Alpha-ConotoxinMII- and A85380-Sensitive Nicotinic Receptors in Parkinson’s Disease Striatum. J. Neurochem. 2004, 88, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Quik, M.; McIntosh, J.M. Striatal A6 * Nicotinic Acetylcholine Receptors: Potential Targets for Parkinson’s Disease Therapy. J. Pharmacol. Exp. Ther. 2006, 316, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Bordia, T.; Grady, S.R.; McIntosh, J.M.; Quik, M. Nigrostriatal Damage Preferentially Decreases a Subpopulation of A6β2 * NAChRs in Mouse, Monkey, and Parkinson’s Disease Striatum. Mol. Pharmacol. 2007, 72, 52–61. [Google Scholar] [CrossRef]
- Azam, L.; McIntosh, J.M. Alpha-Conotoxins as Pharmacological Probes of Nicotinic Acetylcholine Receptors. Acta Pharmacol. Sin. 2009, 30, 771–783. [Google Scholar] [CrossRef]
- Turner, M.; Eidemiller, S.; Martin, B.; Narver, A.; Marshall, J.; Zemp, L.; Cornell, K.A.; McIntosh, J.M.; McDougal, O.M. Structural Basis for α-Conotoxin Potency and Selectivity. Bioorg. Med. Chem. 2009, 17, 5894–5899. [Google Scholar] [CrossRef]
- Satkunanathan, N.; Livett, B.; Gayler, K.; Sandall, D.; Down, J.; Khalil, Z. Alpha-Conotoxin Vc1.1 Alleviates Neuropathic Pain and Accelerates Functional Recovery of Injured Neurones. Brain Res. 2005, 1059, 149–158. [Google Scholar] [CrossRef]
- Livett, B.G.; Sandall, D.W.; Keays, D.; Down, J.; Gayler, K.R.; Satkunanathan, N.; Khalil, Z. Therapeutic Applications of Conotoxins That Target the Neuronal Nicotinic Acetylcholine Receptor. Toxicon 2006, 48, 810–829. [Google Scholar] [CrossRef]
- Pacini, A.; Micheli, L.; Maresca, M.; Branca, J.J.V.; McIntosh, J.M.; Ghelardini, C.; Di Cesare Mannelli, L. The A9α10 Nicotinic Receptor Antagonist α-Conotoxin RgIA Prevents Neuropathic Pain Induced by Oxaliplatin Treatment. Exp. Neurol. 2016, 282, 37–48. [Google Scholar] [CrossRef]
- Liu, C.; Wu, P.; Zhu, H.; Grieco, P.; Yu, R.; Gao, X.; Wu, G.; Wang, D.; Xu, H.; Qi, W. Rationally Designed α-Conotoxin Analogues Maintained Analgesia Activity and Weakened Side Effects. Molecules 2019, 24, 337. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, X.; Zhangsun, D.; Yu, G.; Su, R.; Luo, S. The A9α10 Nicotinic Acetylcholine Receptor Antagonist AO-Conotoxin GeXIVA [1,2] Alleviates and Reverses Chemotherapy-Induced Neuropathic Pain. Mar. Drugs 2019, 17, 265. [Google Scholar] [CrossRef] [PubMed]
- Dyachenko, I.A.; Palikova, Y.A.; Palikov, V.A.; Korolkova, Y.V.; Kazakov, V.A.; Egorova, N.S.; Garifulina, A.I.; Utkin, Y.N.; Tsetlin, V.I.; Kryukova, E.V. α-Conotoxin RgIA and Oligoarginine R8 in the Mice Model Alleviate Long-Term Oxaliplatin Induced Neuropathy. Biochimie 2022, 194, 127–136. [Google Scholar] [CrossRef]
- Stevens, M.; Peigneur, S.; Tytgat, J. Neurotoxins and Their Binding Areas on Voltage-Gated Sodium Channels. Front. Pharmacol. 2011, 2, 71. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-Y.; Nau, C.; Wang, G.K. Residues in Na+ Channel D3-S6 Segment Modulate Both Batrachotoxin and Local Anesthetic Affinities. Biophys. J. 2000, 79, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-Y.; Barile, M.; Wang, G.K. Disparate Role of Na+ Channel D2-S6 Residues in Batrachotoxin and Local Anesthetic Action. Mol. Pharmacol. 2001, 59, 1100–1107. [Google Scholar] [CrossRef]
- Catterall, W.A.; Trainer, V.; Baden, D.G. Molecular Properties of the Sodium Channel: A Receptor for Multiple Neurotoxins. Bull. Soc. Pathol. Exot. 1990 1992, 85, 481–485. [Google Scholar]
- Trainer, V.L.; Baden, D.G.; Catterall, W.A. Identification of Peptide Components of the Brevetoxin Receptor Site of Rat Brain Sodium Channels. J. Biol. Chem. 1994, 269, 19904–19909. [Google Scholar] [CrossRef]
- Lewis, R.J. Ion Channel Toxins and Therapeutics: From Cone Snail Venoms to Ciguatera. Ther. Drug Monit. 2000, 22, 61–64. [Google Scholar] [CrossRef]
- Wang, S. Voltage-Gated Sodium Channels as Primary Targets of Diverse Lipid-Soluble Neurotoxins. Cell. Signal. 2003, 15, 151–159. [Google Scholar] [CrossRef]
- Norton, R.S. Structure and Structure-Function Relationships of Sea Anemone Proteins That Interact with the Sodium Channel. Toxicon 1991, 29, 1051–1084. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Martin-Eauclaire, M.-F.; Cestèle, S.; Kopeyan, C.; Carlier, E.; Khalifa, R.B.; Pelhate, M.; Rochat, H. Scorpion Toxins Affecting Sodium Current Inactivation Bind to Distinct Homologous Receptor Sites on Rat Brain and Insect Sodium Channels. J. Biol. Chem. 1996, 271, 8034–8045. [Google Scholar] [CrossRef] [PubMed]
- Cestèle, S. Molecular Mechanisms of Neurotoxin Action on Voltage-Gated Sodium Channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Cestèle, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-Gated Ion Channels and Gating Modifier Toxins. Toxicon 2007, 49, 124–141. [Google Scholar] [CrossRef]
- Hanck, D.A.; Sheets, M.F. Site-3 Toxins and Cardiac Sodium Channels. Toxicon 2007, 49, 181–193. [Google Scholar] [CrossRef]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea Anemone Toxins Affecting Voltage-Gated Sodium Channels—Molecular and Evolutionary Features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef]
- Leipold, E.; Hansel, A.; Olivera, B.M.; Terlau, H.; Heinemann, S.H. Molecular Interaction of δ-Conotoxins with Voltage-Gated Sodium Channels. FEBS Lett. 2005, 579, 3881–3884. [Google Scholar] [CrossRef]
- Heinemann, S.H.; Leipold, E. Conotoxins of the O-Superfamily Affecting Voltage-Gated Sodium Channels. Cell. Mol. Life Sci. 2007, 64, 1329–1340. [Google Scholar] [CrossRef]
- Shen, H.; Li, Z.; Jiang, Y.; Pan, X.; Wu, J.; Cristofori-Armstrong, B.; Smith, J.J.; Chin, Y.K.Y.; Lei, J.; Zhou, Q.; et al. Structural Basis for the Modulation of Voltage-Gated Sodium Channels by Animal Toxins. Science 2018, 362, eaau2596. [Google Scholar] [CrossRef]
- Wisedchaisri, G.; Tonggu, L.; Gamal El-Din, T.M.; McCord, E.; Zheng, N.; Catterall, W.A. Structural Basis for High-Affinity Trapping of the NaV1.7 Channel in Its Resting State by Tarantula Toxin. Mol. Cell 2021, 81, 38–48.e4. [Google Scholar] [CrossRef]
- Clairfeuille, T.; Cloake, A.; Infield, D.T.; Llongueras, J.P.; Arthur, C.P.; Li, Z.R.; Jian, Y.; Martin-Eauclaire, M.-F.; Bougis, P.E.; Ciferri, C.; et al. Structural Basis of α-Scorpion Toxin Action on Nav Channels. Science 2019, 363, eaav8573. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Yu, R.; Jin, A.-H.; Dutertre, S.; Craik, D.J. ConoServer: Updated Content, Knowledge, and Discovery Tools in the Conopeptide Database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef] [PubMed]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, Synthesis, and Structure–Activity Relationships of Conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Fuller, E.; Green, B.R.; Catlin, P.; Buczek, O.; Nielsen, J.S.; Olivera, B.M.; Bulaj, G. Oxidative Folding of Conotoxins Sharing an Identical Disulfide Bridging Framework: Oxidative Folding of Conotoxins. FEBS J. 2005, 272, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Corpuz, G.P.; Jacobsen, R.B.; Jimenez, E.C.; Watkins, M.; Walker, C.; Colledge, C.; Garrett, J.E.; McDougal, O.; Li, W.; Gray, W.R.; et al. Definition of the M-Conotoxin Superfamily: Characterization of Novel Peptides from Molluscivorous Conus Venoms. Biochemistry 2005, 44, 8176–8186. [Google Scholar] [CrossRef]
- Jacob, R.B.; McDougal, O.M. The M-Superfamily of Conotoxins: A Review. Cell. Mol. Life Sci. 2010, 67, 17–27. [Google Scholar] [CrossRef]
- Green, B.R.; Bulaj, G.; Norton, R.S. Structure and Function of μ-Conotoxins, Peptide-Based Sodium Channel Blockers with Analgesic Activity. Future Med. Chem. 2014, 6, 1677–1698. [Google Scholar] [CrossRef]
- Ekberg, J.; Craik, D.J.; Adams, D.J. Conotoxin Modulation of Voltage-Gated Sodium Channels. Int. J. Biochem. Cell Biol. 2008, 40, 2363–2368. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Kohda, D.; Hatanaka, H.; Lancelin, J.M.; Ishida, Y.; Oya, M.; Nakamura, H.; Inagaki, F.; Sato, K. Structure-Activity Relationships of.Mu.-Conotoxin GIIIA: Structure Determination of Active and Inactive Sodium Channel Blocker Peptides by NMR and Simulated Annealing Calculations. Biochemistry 1992, 31, 12577–12584. [Google Scholar] [CrossRef]
- Deuis, J.R.; Dekan, Z.; Inserra, M.C.; Lee, T.-H.; Aguilar, M.-I.; Craik, D.J.; Lewis, R.J.; Alewood, P.F.; Mobli, M.; Schroeder, C.I.; et al. Development of a ΜO-Conotoxin Analogue with Improved Lipid Membrane Interactions and Potency for the Analgesic Sodium Channel NaV1.8. J. Biol. Chem. 2016, 291, 11829–11842. [Google Scholar] [CrossRef]
- Volpon, L.; Lamthanh, H.; Barbier, J.; Gilles, N.; Molgó, J.; Ménez, A.; Lancelin, J.-M. NMR Solution Structures of δ-Conotoxin EVIA from Conus Ermineus That Selectively Acts on Vertebrate Neuronal Na+ Channels. J. Biol. Chem. 2004, 279, 21356–21366. [Google Scholar] [CrossRef] [PubMed]
- Buczek, O.; Wei, D.; Babon, J.J.; Yang, X.; Fiedler, B.; Chen, P.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure and Sodium Channel Activity of an Excitatory I1 -Superfamily Conotoxin. Biochemistry 2007, 46, 9929–9940. [Google Scholar] [CrossRef]
- Endean, R.; Parish, G.; Gyr, P. Pharmacology of the Venom of Conus Geographus. Toxicon 1974, 12, 131–138. [Google Scholar] [CrossRef]
- Nakamura, H.; Kobayashi, J.; Ohizumi, Y.; Hirata, Y. Isolation and Amino Acid Compositions of Geographutoxin I and II from the Marine SnailConus Geographus Linné. Experientia 1983, 39, 590–591. [Google Scholar] [CrossRef]
- Sato, S.; Nakamura, H.; Ohizumi, Y.; Kobayashi, J.; Hirata, Y. The Amino Acid Sequences of Homologous Hydroxyproline-Containing Myotoxins from the Marine Snal Conus Geographus Venom. FEBS Lett. 1983, 155, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Gray, W.R.; Olivera, B.M.; Zeikus, R.D.; Kerr, L.; Yoshikami, D.; Moczydlowski, E. Conus Geographus Toxins That Discriminate between Neuronal and Muscle Sodium Channels. J. Biol. Chem. 1985, 260, 9280–9288. [Google Scholar] [CrossRef] [PubMed]
- Gonoi, T.; Ohizumi, Y.; Nakamura, H.; Kobayashi, J.; Catterall, W. The Conus Toxin Geographutoxin IL Distinguishes Two Functional Sodium Channel Subtypes in Rat Muscle Cells Developing in Vitro. J. Neurosci. 1987, 7, 1728–1731. [Google Scholar] [CrossRef]
- Gonoi, T.; Hagihara, Y.; Kobayashi, J.; Nakamura, H.; Ohizumi, Y. Geographutoxin-Sensitive and Insensitive Sodium Currents in Mouse Skeletal Muscle Developing in Situ. J. Physiol. 1989, 414, 159–177. [Google Scholar] [CrossRef]
- Kobayashi, M.; Wu, C.H.; Yoshii, M.; Narahashi, T.; Nakamura, H.; Kobayashi, J.; Ohizumi, Y. Preferential Block of Skeletal Muscle Sodium Channels by Geographutoxin II, a New Peptide Toxin FromConus Geographus. Pflügers Archiv. Eur. J. Physiol. 1986, 407, 241–243. [Google Scholar] [CrossRef]
- Noda, M.; Shimizu, S.; Tanabe, T.; Takai, T.; Kayano, T.; Ikeda, T.; Takahashi, H.; Nakayama, H.; Kanaoka, Y.; Minamino, N.; et al. Primary Structure of Electrophorus Electricus Sodium Channel Deduced from CDNA Sequence. Nature 1984, 312, 121–127. [Google Scholar] [CrossRef]
- Noda, M.; Ikeda, T.; Kayano, T.; Suzuki, H.; Takeshima, H.; Kurasaki, M.; Takahashi, H.; Numa, S. Existence of Distinct Sodium Channel Messenger RNAs in Rat Brain. Nature 1986, 320, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Kayano, T.; Noda, M.; Flockerzi, V.; Takahashi, H.; Numa, S. Primary Structure of Rat Brain Sodium Channel III Deduced from the CDNA Sequence. FEBS Lett. 1988, 228, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Rogart, R.B.; Cribbs, L.L.; Muglia, L.K.; Kephart, D.D.; Kaiser, M.W. Molecular Cloning of a Putative Tetrodotoxin-Resistant Rat Heart Na+ Channel Isoform. Proc. Natl. Acad. Sci. USA 1989, 86, 8170–8174. [Google Scholar] [CrossRef] [PubMed]
- Sills, M.N.; Xu, Y.C.; Baracchini, E.; Goodman, R.H.; Cooperman, S.S.; Mandel, G.; Chien, K.R. Expression of Diverse Na+ Channel Messenger RNAs in Rat Myocardium. Evidence for a Cardiac-Specific Na+ Channel. J. Clin. Investig. 1989, 84, 331–336. [Google Scholar] [CrossRef]
- Gellens, M.E.; George, A.L.; Chen, L.Q.; Chahine, M.; Horn, R.; Barchi, R.L.; Kallen, R.G. Primary Structure and Functional Expression of the Human Cardiac Tetrodotoxin-Insensitive Voltage-Dependent Sodium Channel. Proc. Natl. Acad. Sci. USA 1992, 89, 554–558. [Google Scholar] [CrossRef]
- White, M.M.; Chen, L.Q.; Kleinfield, R.; Kallen, R.G.; Barchi, R.L. SkM2, a Na+ Channel CDNA Clone from Denervated Skeletal Muscle, Encodes a Tetrodotoxin-Insensitive Na+ Channel. Mol. Pharmacol. 1991, 39, 604–608. [Google Scholar]
- Trimmer, J.S.; Cooperman, S.S.; Tomiko, S.A.; Zhou, J.; Crean, S.M.; Boyle, M.B.; Kalen, R.G.; Sheng, Z.; Barchi, R.L.; Sigworth, F.J.; et al. Primary Structure and Functional Expression of a Mammalian Skeletal Muscle Sodium Channel. Neuron 1989, 3, 33–49. [Google Scholar] [CrossRef]
- George, A.L.; Komisarof, J.; Kallen, R.G.; Barchi, R.L. Primary Structure of the Adult Human Skeletal Muscle Voltage-Dependent Sodium Channel. Ann. Neurol. 1992, 31, 131–137. [Google Scholar] [CrossRef]
- Goldin, A.L.; Snutch, T.; Lübbert, H.; Dowsett, A.; Marshall, J.; Auld, V.; Downey, W.; Fritz, L.C.; Lester, H.A.; Dunn, R. Messenger RNA Coding for Only the Alpha Subunit of the Rat Brain Na Channel Is Sufficient for Expression of Functional Channels in Xenopus Oocytes. Proc. Natl. Acad. Sci. USA 1986, 83, 7503–7507. [Google Scholar] [CrossRef]
- Stühmer, W.; Methfessel, C.; Sakmann, B.; Noda, M.; Numa, S. Patch Clamp Characterization of Sodium Channels Expressed from Rat Brain CDNA. Eur. Biophys. J. 1987, 14, 131–138. [Google Scholar] [CrossRef]
- Suzuki, H.; Beckh, S.; Kubo, H.; Yahagi, N.; Ishida, H.; Kayano, T.; Noda, M.; Numa, S. Functional Expression of Cloned CDNA Encoding Sodium Channel III. FEBS Lett. 1988, 228, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Chahine, M.; Bennett, P.B.; George, A.L.; Horn, R. Functional Expression and Properties of the Human Skeletal Muscle Sodium Channel. Pflügers Arch. 1994, 427, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Ikeda, T.; Suzuki, H.; Takeshima, H.; Takahashi, T.; Kuno, M.; Numa, S. Expression of Functional Sodium Channels from Cloned CDNA. Nature 1986, 322, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.J.; Watson, M.; Adams, D.J.; Hammarström, A.K.; Gage, P.W.; Hill, J.M.; Craik, D.J.; Thomas, L.; Adams, D.; Alewood, P.F.; et al. Solution Structure of μ-Conotoxin PIIIA, a Preferential Inhibitor of Persistent Tetrodotoxin-Sensitive Sodium Channels. J. Biol. Chem. 2002, 277, 27247–27255. [Google Scholar] [CrossRef]
- Harvey, P.; Kurniawan, N.; Finol-Urdaneta, R.; McArthur, J.; Van Lysebetten, D.; Dash, T.; Hill, J.; Adams, D.; Durek, T.; Craik, D. NMR Structure of μ-Conotoxin GIIIC: Leucine 18 Induces Local Repacking of the N-Terminus Resulting in Reduced NaV Channel Potency. Molecules 2018, 23, 2715. [Google Scholar] [CrossRef]
- Sheets, M.F.; Hanck, D.A. Voltage-Dependent Open-State Inactivation of Cardiac Sodium Channels: Gating Current Studies with Anthopleurin—A Toxin. J. Gen. Physiol. 1995, 106, 617–640. [Google Scholar] [CrossRef]
- Sheets, M.F.; Hanck, D.A. Gating of Skeletal and Cardiac Muscle Sodium Channels in Mammalian Cells. J. Physiol. 1999, 514, 425–436. [Google Scholar] [CrossRef]
- Ohizumi, Y.; Nakamura, H.; Kobayashi, J.; Catterall, W.A. Specific Inhibition of [3H] Saxitoxin Binding to Skeletal Muscle Sodium Channels by Geographutoxin II, a Polypeptide Channel Blocker. J. Biol. Chem. 1986, 261, 6149–6152. [Google Scholar] [CrossRef]
- Moczydlowski, E.; Olivera, B.M.; Gray, W.R.; Strichartz, G.R. Discrimination of Muscle and Neuronal Na-Channel Subtypes by Binding Competition between [3H]Saxitoxin and Mu-Conotoxins. Proc. Natl. Acad. Sci. USA 1986, 83, 5321–5325. [Google Scholar] [CrossRef]
- Yanagawa, Y.; Abe, T.; Satake, M. Blockade of [3H]Lysine-Tetrodotoxin Binding to Sodium Channel Proteins by Conotoxin GIII. Neurosci. Lett. 1986, 64, 7–12. [Google Scholar] [CrossRef]
- Cruz, L.J.; Kupryszewski, G.; LeCheminant, G.W.; Gray, W.R.; Olivera, B.M.; Rivier, J.M. Conotoxin GIIIA, a Peptide Ligand for Muscle Sodium Channels: Chemical Synthesis, Radiolabeling and Receptor Characterization. Biochemistry 1989, 28, 3437–3442. [Google Scholar] [CrossRef] [PubMed]
- French, R.J.; Horn, R. Shifts of Macroscopic Current Activation in Partially Blocked Sodium Channels. Interaction between the Voltage Sensor and a μ-Conotoxin. In From Ion Channels to Cell-to-Cell Conversations; Latorre, R., Sáez, J.C., Eds.; Springer: Boston, MA, USA, 1997; pp. 67–89. ISBN 978-1-4899-1797-3. [Google Scholar]
- French, R.J.; Yoshikami, D.; Sheets, M.F.; Olivera, B.M. The Tetrodotoxin Receptor of Voltage-Gated Sodium Channels—Perspectives from Interactions with μ-Conotoxins. Mar. Drugs 2010, 8, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- Capes, D.L.; Arcisio-Miranda, M.; Jarecki, B.W.; French, R.J.; Chanda, B. Gating Transitions in the Selectivity Filter Region of a Sodium Channel Are Coupled to the Domain IV Voltage Sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 2648–2653. [Google Scholar] [CrossRef] [PubMed]
- Lancelin, J.M.; Kohda, D.; Tate, S.; Yanagawa, Y.; Abe, T.; Satake, M.; Inagaki, F. Tertiary Structure of Conotoxin GIIIA in Aqueous Solution. Biochemistry 1991, 30, 6908–6916. [Google Scholar] [CrossRef]
- Hill, J.M.; Alewood, P.F.; Craik, D.J. Three-Dimensional Solution Structure of μ-Conotoxin GIIIB, a Specific Blocker of Skeletal Muscle Sodium Channels. Biochemistry 1996, 35, 8824–8835. [Google Scholar] [CrossRef]
- Sato, K.; Ishida, Y.; Wakamatsu, K.; Kato, R.; Honda, H.; Ohizumi, Y.; Nakamura, H.; Ohya, M.; Lancelin, J.M.; Kohda, D. Active Site of Mu-Conotoxin GIIIA, a Peptide Blocker of Muscle Sodium Channels. J. Biol. Chem. 1991, 266, 16989–16991. [Google Scholar] [CrossRef]
- Becker, S.; Prusak-Sochaczewski, E.; Zamponi, G.; Beck-Sickinger, A.G.; Gordon, R.D.; French, R.J. Action of Derivatives of.Mu.-Conotoxin GIIIA on Sodium Channels. Single Amino Acid Substitutions in the Toxin Separately Affect Association and Dissociation Rates. Biochemistry 1992, 31, 8229–8238. [Google Scholar] [CrossRef]
- Chahine, M.; Chen, L.Q.; Fotouhi, N.; Walsky, R.; Fry, D.; Santarelli, V.; Horn, R.; Kallen, R.G. Characterizing the Mu-Conotoxin Binding Site on Voltage-Sensitive Sodium Channels with Toxin Analogs and Channel Mutations. Recept. Channels 1995, 3, 161–174. [Google Scholar]
- Han, P.; Wang, K.; Dai, X.; Cao, Y.; Liu, S.; Jiang, H.; Fan, C.; Wu, W.; Chen, J. The Role of Individual Disulfide Bonds of μ-Conotoxin GIIIA in the Inhibition of NaV1.4. Mar. Drugs 2016, 14, 213. [Google Scholar] [CrossRef]
- Khan, A.; Romantseva, L.; Lam, A.; Lipkind, G.; Fozzard, H.A. Role of Outer Ring Carboxylates of the Rat Skeletal Muscle Sodium Channel Pore in Proton Block. J. Physiol. 2002, 543, 71–84. [Google Scholar] [CrossRef]
- Lipkind, G.M.; Fozzard, H.A. Voltage-Gated Na Channel Selectivity: The Role of the Conserved Domain III Lysine Residue. J. Gen. Physiol. 2008, 131, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Dudev, T.; Lim, C. Ion Selectivity Strategies of Sodium Channel Selectivity Filters. Acc. Chem. Res. 2014, 47, 3580–3587. [Google Scholar] [CrossRef]
- Flood, E.; Boiteux, C.; Allen, T.W. Selective Ion Permeation Involves Complexation with Carboxylates and Lysine in a Model Human Sodium Channel. PLoS Comput. Biol. 2018, 14, e1006398. [Google Scholar] [CrossRef] [PubMed]
- Penzotti, J.L.; Fozzard, H.A.; Lipkind, G.M.; Dudley, S.C. Differences in Saxitoxin and Tetrodotoxin Binding Revealed by Mutagenesis of the Na+ Channel Outer Vestibule. Biophys. J. 1998, 75, 2647–2657. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Heinemann, S.H.; Stühmer, W.; Pusch, M.; Conti, F.; Imoto, K.; Numa, S. Mapping the Site of Block by Tetrodotoxin and Saxitoxin of Sodium Channel II. FEBS Lett. 1991, 293, 93–96. [Google Scholar] [CrossRef]
- Dudley, S.C.; Todt, H.; Lipkind, G.; Fozzard, H.A. A Mu-Conotoxin-Insensitive Na+ Channel Mutant: Possible Localization of a Binding Site at the Outer Vestibule. Biophys. J. 1995, 69, 1657–1665. [Google Scholar] [CrossRef]
- Li, R.A.; Tsushima, R.G.; Kallen, R.G.; Backx, P.H. Pore Residues Critical for Mu-CTX Binding to Rat Skeletal Muscle Na+ Channels Revealed by Cysteine Mutagenesis. Biophys. J. 1997, 73, 1874–1884. [Google Scholar] [CrossRef]
- Chahine, M.; Sirois, J.; Marcotte, P.; Chen, L.-Q.; Kallen, R.G. Extrapore Residues of the S5-S6 Loop of Domain 2 of the Voltage-Gated Skeletal Muscle Sodium Channel (RSkM1) Contribute to the μ-Conotoxin GIIIA Binding Site. Biophys. J. 1998, 75, 236–246. [Google Scholar] [CrossRef]
- Carbonneau, E.; Vijayaragavan, K.; Chahine, M. A Tryptophan Residue (W736) in the Amino-Terminus of the P-Segment of Domain II Is Involved in Pore Formation in Nav1.4 Voltage-Gated Sodium Channels. Pflügers Arch. Eur. J. Physiol. 2002, 445, 18–24. [Google Scholar] [CrossRef]
- Li, R.A.; Hui, K.; French, R.J.; Sato, K.; Henrikson, C.A.; Tomaselli, G.F.; Marbán, E. Dependence of μ-Conotoxin Block of Sodium Channels on Ionic Strength but Not on the Permeating [Na+]. J. Biol. Chem. 2003, 278, 30912–30919. [Google Scholar] [CrossRef]
- Nakamura, M.; Ishida, Y.; Kohno, T.; Sato, K.; Oba, Y.; Nakamura, H. Effects of Modification at the Fifth Residue of Mu-Conotoxin GIIIA with Bulky Tags on the Electrically Stimulated Contraction of the Rat Diaphragm. J. Pept. Res. Off. J. Am. Pept. Soc. 2004, 64, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, G.; Aliste, M.P.; Tieleman, D.P.; French, R.J.; Dudley, S.C., Jr. Docking of μ-Conotoxin GIIIA in the Sodium Channel Outer Vestibule. Channels 2007, 1, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.; Lipkind, G.; Fozzard, H.A.; French, R.J. Electrostatic and Steric Contributions to Block of the Skeletal Muscle Sodium Channel by μ-Conotoxin. J. Gen. Physiol. 2002, 119, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.S.; French, R.J.; Lipkind, G.M.; Fozzard, H.A.; Dudley, S. Predominant Interactions between μ-Conotoxin Arg-13 and the Skeletal Muscle Na+ Channel Localized by Mutant Cycle Analysis. Biochemistry 1998, 37, 4407–4419. [Google Scholar] [CrossRef]
- Li, R.A.; Sato, K.; Kodama, K.; Kohno, T.; Xue, T.; Tomaselli, G.F.; Marbán, E. Charge Conversion Enables Quantification of the Proximity between a Normally-Neutral μ-Conotoxin (GIIIA) Site and the Na+ Channel Pore. FEBS Lett. 2002, 511, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Li, R.A.; Ennis, I.L.; Tomaselli, G.F.; French, R.J.; Marbán, E. Latent Specificity of Molecular Recognition in Sodium Channels Engineered To Discriminate between Two “Indistinguishable” μ-Conotoxins. Biochemistry 2001, 40, 6002–6008. [Google Scholar] [CrossRef] [PubMed]
- Cummins, T.R.; Aglieco, F.; Dib-Hajj, S.D. Critical Molecular Determinants of Voltage-Gated Sodium Channel Sensitivity to μ-Conotoxins GIIIA/B. Mol. Pharmacol. 2002, 61, 1192–1201. [Google Scholar] [CrossRef]
- Pan, X.; Li, Z.; Huang, X.; Huang, G.; Gao, S.; Shen, H.; Liu, L.; Lei, J.; Yan, N. Molecular Basis for Pore Blockade of Human Na+ Channel Nav 1.2 by the μ-Conotoxin KIIIA. Science 2019, 363, 1309–1313. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Li, R.A.; Ennis, I.L.; Vélez, P.; Tomaselli, G.F.; Marbán, E. Novel Structural Determinants of Mu-Conotoxin (GIIIB) Block in Rat Skeletal Muscle (Mu1) Na+ Channels. J. Biol. Chem. 2000, 275, 27551–27558. [Google Scholar] [CrossRef]
- Li, R.A.; Vélez, P.; Chiamvimonvat, N.; Tomaselli, G.F.; Marbán, E. Charged Residues between the Selectivity Filter and S6 Segments Contribute to the Permeation Phenotype of the Sodium Channel. J. Gen. Physiol. 2000, 115, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Ennis, I.L.; Sato, K.; French, R.J.; Li, R.A. Novel Interactions Identified between μ-Conotoxin and the Na+ Channel Domain I P-Loop: Implications for Toxin-Pore Binding Geometry. Biophys. J. 2003, 85, 2299–2310. [Google Scholar] [CrossRef] [PubMed]
- Todt, H.; Dudley, S.C., Jr.; Kyle, J.W.; French, R.J.; Fozzard, H.A. Ultra-Slow Inactivation in Μ1 Na+ Channels Is Produced by a Structural Rearrangement of the Outer Vestibule. Biophys. J. 1999, 76, 1335–1345. [Google Scholar] [CrossRef]
- Li, R.A.; Ennis, I.L.; French, R.J.; Dudley, S.C.; Tomaselli, G.F.; Marbán, E. Clockwise Domain Arrangement of the Sodium Channel Revealed by ¼-Conotoxin (GIIIA) Docking Orientation. J. Biol. Chem. 2001, 276, 11072–11077. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Hasson, A.; Spira, M.E.; Gray, W.R.; Li, W.; Marsh, M.; Hillyard, D.R.; Olivera, B.M. A New Family of Conotoxins That Blocks Voltage-Gated Sodium Channels. J. Biol. Chem. 1995, 270, 16796–16802. [Google Scholar] [CrossRef] [PubMed]
- Shon, K.J.; Olivera, B.M.; Watkins, M.; Jacobsen, R.B.; Gray, W.R.; Floresca, C.Z.; Cruz, L.J.; Hillyard, D.R.; Brink, A.; Terlau, H.; et al. Mu-Conotoxin PIIIA, a New Peptide for Discriminating among Tetrodotoxin-Sensitive Na Channel Subtypes. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 4473–4481. [Google Scholar] [CrossRef]
- Lewis, R.J.; Schroeder, C.I.; Ekberg, J.; Nielsen, K.J.; Loughnan, M.; Thomas, L.; Adams, D.A.; Drinkwater, R.; Adams, D.J.; Alewood, P.F. Isolation and Structure-Activity of μ-Conotoxin TIIIA, A Potent Inhibitor of Tetrodotoxin-Sensitive Voltage-Gated Sodium Channels. Mol. Pharmacol. 2007, 71, 676–685. [Google Scholar] [CrossRef]
- Walewska, A.; Skalicky, J.J.; Davis, D.R.; Zhang, M.-M.; Lopez-Vera, E.; Watkins, M.; Han, T.S.; Yoshikami, D.; Olivera, B.M.; Bulaj, G. NMR-Based Mapping of Disulfide Bridges in Cysteine-Rich Peptides: Application to the μ-Conotoxin SxIIIA. J. Am. Chem. Soc. 2008, 130, 14280–14286. [Google Scholar] [CrossRef]
- Yang, M.; Zhao, S.; Min, X.; Shao, M.; Chen, Y.; Chen, Z.; Zhou, M. A Novel μ-Conotoxin from Worm-Hunting Conus Tessulatus That Selectively Inhibit Rat TTX-Resistant Sodium Currents. Toxicon 2017, 130, 11–18. [Google Scholar] [CrossRef]
- West, P.J.; Bulaj, G.; Garrett, J.E.; Olivera, B.M.; Yoshikami, D. μ-Conotoxin SmIIIA, a Potent Inhibitor of Tetrodotoxin-Resistant Sodium Channels in Amphibian Sympathetic and Sensory Neurons. Biochemistry 2002, 41, 15388–15393. [Google Scholar] [CrossRef]
- Bulaj, G.; West, P.J.; Garrett, J.E.; Watkins, M.; Zhang, M.-M.; Norton, R.S.; Smith, B.J.; Yoshikami, D.; Olivera, B.M. Novel Conotoxins from Conus Striatus and Conus Kinoshitai Selectively Block TTX-Resistant Sodium Channels. Biochemistry 2005, 44, 7259–7265. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Z.; Zhang, H.; Jiang, H.; Lu, W.; Zhao, Z.-Q.; Chi, C.-W. A Novel Conotoxin from Conus Striatus, μ-SIIIA, Selectively Blocking Rat Tetrodotoxin-Resistant Sodium Channels. Toxicon 2006, 47, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-M.; Fiedler, B.; Green, B.R.; Catlin, P.; Watkins, M.; Garrett, J.E.; Smith, B.J.; Yoshikami, D.; Olivera, B.M.; Bulaj, G. Structural and Functional Diversities among μ-Conotoxins Targeting TTX-Resistant Sodium Channels. Biochemistry 2006, 45, 3723–3732. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, C.I.; Ekberg, J.; Nielsen, K.J.; Adams, D.; Loughnan, M.L.; Thomas, L.; Adams, D.J.; Alewood, P.F.; Lewis, R.J. Neuronally Selective μ-Conotoxins from Conus Striatus Utilize an α-Helical Motif to Target Mammalian Sodium Channels. J. Biol. Chem. 2008, 283, 21621–21628. [Google Scholar] [CrossRef]
- Holford, M.; Zhang, M.-M.; Gowd, K.H.; Azam, L.; Green, B.R.; Watkins, M.; Ownby, J.-P.; Yoshikami, D.; Bulaj, G.; Olivera, B.M. Pruning Nature: Biodiversity-Derived Discovery of Novel Sodium Channel Blocking Conotoxins from Conus Bullatus. Toxicon 2009, 53, 90–98. [Google Scholar] [CrossRef]
- Khoo, K.K.; Gupta, K.; Green, B.R.; Zhang, M.-M.; Watkins, M.; Olivera, B.M.; Balaram, P.; Yoshikami, D.; Bulaj, G.; Norton, R.S. Distinct Disulfide Isomers of μ-Conotoxins KIIIA and KIIIB Block Voltage-Gated Sodium Channels. Biochemistry 2012, 51, 9826–9835. [Google Scholar] [CrossRef]
- Favreau, P.; Benoit, E.; Hocking, H.G.; Carlier, L.; D’hoedt, D.; Leipold, E.; Markgraf, R.; Schlumberger, S.; Córdova, M.A.; Gaertner, H.; et al. A Novel Μ-Conopeptide, CnIIIC, Exerts Potent and Preferential Inhibition of NaV1.2/1.4 Channels and Blocks Neuronal Nicotinic Acetylcholine Receptors: A µ-Conopeptide with Novel Neuropharmacology. Br. J. Pharmacol. 2012, 166, 1654–1668. [Google Scholar] [CrossRef]
- McMahon, K.L.; Tran, H.N.T.; Deuis, J.R.; Lewis, R.J.; Vetter, I.; Schroeder, C.I. Discovery, Pharmacological Characterisation and NMR Structure of the Novel µ-Conotoxin SxIIIC, a Potent and Irreversible NaV Channel Inhibitor. Biomedicines 2020, 8, 391. [Google Scholar] [CrossRef]
- Floresca, C.Z. A Comparison of the μ-Conotoxins by [3H]Saxitoxin Binding Assays in Neuronal and Skeletal Muscle Sodium Channel. Toxicol. Appl. Pharmacol. 2003, 190, 95–101. [Google Scholar] [CrossRef]
- McArthur, J.R.; Singh, G.; O’Mara, M.L.; McMaster, D.; Ostroumov, V.; Tieleman, D.P.; French, R.J. Orientation of μ-Conotoxin PIIIA in a Sodium Channel Vestibule, Based on Voltage Dependence of Its Binding. Mol. Pharmacol. 2011, 80, 219–227. [Google Scholar] [CrossRef]
- McArthur, J.R.; Ostroumov, V.; Al-Sabi, A.; McMaster, D.; French, R.J. Multiple, Distributed Interactions of μ-Conotoxin PIIIA Associated with Broad Targeting among Voltage-Gated Sodium Channels. Biochemistry 2011, 50, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Safo, P.; Rosenbaum, T.; Shcherbatko, A.; Choi, D.-Y.; Han, E.; Toledo-Aral, J.J.; Olivera, B.M.; Brehm, P.; Mandel, G. Distinction among Neuronal Subtypes of Voltage-Activated Sodium Channels by μ-Conotoxin PIIIA. J. Neurosci. 2000, 20, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Keizer, D.W.; West, P.J.; Lee, E.F.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structural Basis for Tetrodotoxin-Resistant Sodium Channel Binding by μ-Conotoxin SmIIIA. J. Biol. Chem. 2003, 278, 46805–46813. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Gruszczynski, P.; Walewska, A.; Bulaj, G.; Olivera, B.M.; Yoshikami, D. Cooccupancy of the Outer Vestibule of Voltage-Gated Sodium Channels by μ-Conotoxin KIIIA and Saxitoxin or Tetrodotoxin. J. Neurophysiol. 2010, 104, 88–97. [Google Scholar] [CrossRef]
- Tran, H.N.T.; McMahon, K.L.; Deuis, J.R.; Vetter, I.; Schroeder, C.I. Structural and Functional Insights into the Inhibition of Human Voltage-Gated Sodium Channels by μ-Conotoxin KIIIA Disulfide Isomers. J. Biol. Chem. 2022, 298, 101728. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and Structure-Function Relationships of Voltage-Gated Sodium Channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Vetter, I.; Mozar, C.A.; Durek, T.; Wingerd, J.S.; Alewood, P.F.; Christie, M.J.; Lewis, R.J. Characterisation of Nav Types Endogenously Expressed in Human SH-SY5Y Neuroblastoma Cells. Biochem. Pharmacol. 2012, 83, 1562–1571. [Google Scholar] [CrossRef]
- Wilson, M.J.; Yoshikami, D.; Azam, L.; Gajewiak, J.; Olivera, B.M.; Bulaj, G.; Zhang, M.-M. μ-Conotoxins That Differentially Block Sodium Channels NaV1.1 through 1.8 Identify Those Responsible for Action Potentials in Sciatic Nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 10302–10307. [Google Scholar] [CrossRef]
- Yang, M.; Zhou, M. μ-Conotoxin TsIIIA, a Peptide Inhibitor of Human Voltage-Gated Sodium Channel HNav1.8. Toxicon 2020, 186, 29–34. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Green, B.R.; Catlin, P.; Fiedler, B.; Azam, L.; Chadwick, A.; Terlau, H.; McArthur, J.R.; French, R.J.; Gulyas, J.; et al. Structure/Function Characterization of μ-Conotoxin KIIIA, an Analgesic, Nearly Irreversible Blocker of Mammalian Neuronal Sodium Channels. J. Biol. Chem. 2007, 282, 30699–30706. [Google Scholar] [CrossRef]
- Khoo, K.K.; Feng, Z.-P.; Smith, B.J.; Zhang, M.-M.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure of the Analgesic μ-Conotoxin KIIIA and Effects on the Structure and Function of Disulfide Deletion. Biochemistry 2009, 48, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Zhang, M.-M.; Yoshikami, D.; Azam, L.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure, Dynamics, and Selectivity of the Sodium Channel Blocker μ-Conotoxin SIIIA. Biochemistry 2008, 47, 10940–10949. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, C.I.; Adams, D.; Thomas, L.; Alewood, P.F.; Lewis, R.J. N- and c-Terminal Extensions of μ-Conotoxins Increase Potency and Selectivity for Neuronal Sodium Channels. Biopolymers 2012, 98, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Leipold, E.; Markgraf, R.; Miloslavina, A.; Kijas, M.; Schirmeyer, J.; Imhof, D.; Heinemann, S.H. Molecular Determinants for the Subtype Specificity of μ-Conotoxin SIIIA Targeting Neuronal Voltage-Gated Sodium Channels. Neuropharmacology 2011, 61, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Markgraf, R.; Leipold, E.; Schirmeyer, J.; Paolini-Bertrand, M.; Hartley, O.; Heinemann, S.H. Mechanism and Molecular Basis for the Sodium Channel Subtype Specificity of Μ-Conopeptide CnIIIC: Subtype Specificity of µ-CnIIIC. Br. J. Pharmacol. 2012, 167, 576–586. [Google Scholar] [CrossRef]
- Stevens, M.; Peigneur, S.; Dyubankova, N.; Lescrinier, E.; Herdewijn, P.; Tytgat, J. Design of Bioactive Peptides from Naturally Occurring μ-Conotoxin Structures. J. Biol. Chem. 2012, 287, 31382–31392. [Google Scholar] [CrossRef]
- Van Der Haegen, A.; Peigneur, S.; Tytgat, J. Importance of Position 8 in μ-Conotoxin KIIIA for Voltage-Gated Sodium Channel Selectivity: Residue 8 in KIIIA as Key Residue for Selectivity. FEBS J. 2011, 278, 3408–3418. [Google Scholar] [CrossRef]
- McMahon, K.L.; Tran, H.N.T.; Deuis, J.R.; Craik, D.J.; Vetter, I.; Schroeder, C.I. Μ-Conotoxins Targeting the Human Voltage-Gated Sodium Channel Subtype NaV1.7. Toxins 2022, 14, 600. [Google Scholar] [CrossRef]
- Pallaghy, P.K.; Norton, R.S.; Nielsen, K.J.; Craik, D.J. A Common Structural Motif Incorporating a Cystine Knot and a Triple-Stranded β-Sheet in Toxic and Inhibitory Polypeptides. Protein Sci. 1994, 3, 1833–1839. [Google Scholar] [CrossRef]
- Norton, R.S.; Pallaghy, P.K. The Cystine Knot Structure of Ion Channel Toxins and Related Polypeptides. Toxicon 1998, 36, 1573–1583. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Waine, C. The Cystine Knot Motif in Toxins and Implications for Drug Design. Toxicon 2001, 39, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Dekan, Z.; Knapp, O.; Adams, D.J.; Alewood, P.F.; Lewis, R.J. Isolation, Characterization and Total Regioselective Synthesis of the Novel ΜO-Conotoxin MfVIA from Conus Magnificus That Targets Voltage-Gated Sodium Channels. Biochem. Pharmacol. 2012, 84, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Hillyard, D.R.; Olivera, B.M.; Woodward, S.; Corpuz, G.P.; Gray, W.R.; Ramilo, C.A.; Cruz, L.J. A Molluskivorous Conus Toxin: Conserved Frameworks in Conotoxins. Biochemistry 1989, 28, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Fainzilber, M.; Gordon, D.; Hasson, A.; Spira, M.E.; Zlotkin, E. Mollusc-Specific Toxins from the Venom of Conus Textile Neovicarius. Eur. J. Biochem. 1991, 202, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Kohno, T.; Sasaki, T.; Kobayashi, K.; Fainzilber, M.; Sato, K. Three-Dimensional Solution Structure of the Sodium Channel Agonist/Antagonist δ-Conotoxin TxVIA. J. Biol. Chem. 2002, 277, 36387–36391. [Google Scholar] [CrossRef]
- Shon, K.-J.; Hasson, A.; Spira, M.E.; Cruz, L.J.; Gray, W.R.; Olivera, B.M. Delta.-Conotoxin GmVIA, a Novel Peptide from the Venom of Conus Gloriamaris. Biochemistry 1994, 33, 11420–11425. [Google Scholar] [CrossRef]
- Fainzilber, M.; van der Schors, R.; Lodder, J.C.; Li, K.W.; Geraerts, W.P.M.; Kits, K.S. New Sodium Channel-Blocking Conotoxins Also Affect Calcium Currents in Lymnaea Neurons. Biochemistry 1995, 34, 5364–5371. [Google Scholar] [CrossRef]
- Shon, K.-J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, Characterization, Synthesis, and Cloning of the Lockjaw Peptide from Conus Purpurascens Venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar] [CrossRef]
- Bulaj, G.; DeLaCruz, R.; Azimi-Zonooz, A.; West, P.; Watkins, M.; Yoshikami, D.; Olivera, B.M. δ-Conotoxin Structure/Function through a Cladistic Analysis. Biochemistry 2001, 40, 13201–13208. [Google Scholar] [CrossRef]
- Sudarslal, S.; Majumdar, S.; Ramasamy, P.; Dhawan, R.; Pal, P.P.; Ramaswami, M.; Lala, A.K.; Sikdar, S.K.; Sarma, S.P.; Krishnan, K.S.; et al. Sodium Channel Modulating Activity in a δ-Conotoxin from an Indian Marine Snail. FEBS Lett. 2003, 553, 209–212. [Google Scholar] [CrossRef]
- Barbier, J.; Lamthanh, H.; Le Gall, F.; Favreau, P.; Benoit, E.; Chen, H.; Gilles, N.; Ilan, N.; Heinemann, S.H.; Gordon, D.; et al. A δ-Conotoxin from Conus Ermineus Venom Inhibits Inactivation in Vertebrate Neuronal Na+ Channels but Not in Skeletal and Cardiac Muscles. J. Biol. Chem. 2004, 279, 4680–4685. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Paolini-Bertrand, M.; Gaertner, H.; Biass, D.; Violette, A.; Stöcklin, R.; Favreau, P.; Tytgat, J.; Hartley, O. δ-Conotoxins Synthesized Using an Acid-Cleavable Solubility Tag Approach Reveal Key Structural Determinants for NaV Subtype Selectivity. J. Biol. Chem. 2014, 289, 35341–35350. [Google Scholar] [CrossRef] [PubMed]
- Aman, J.W.; Imperial, J.S.; Ueberheide, B.; Zhang, M.-M.; Aguilar, M.; Taylor, D.; Watkins, M.; Yoshikami, D.; Showers-Corneli, P.; Safavi-Hemami, H.; et al. Insights into the Origins of Fish Hunting in Venomous Cone Snails from Studies of Conus tessulatus. Proc. Natl. Acad. Sci. USA 2015, 112, 5087–5092. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Stocker, M.; Shon, K.J.; McIntosh, J.M.; Olivera, B.M. MicroO-Conotoxin MrVIA Inhibits Mammalian Sodium Channels, but Not through Site I. J. Neurophysiol. 1996, 76, 1423–1429. [Google Scholar] [CrossRef]
- Leipold, E.; Hansel, A.; Borges, A.; Heinemann, S.H. Subtype Specificity of Scorpion β-Toxin Tz1 Interaction with Voltage-Gated Sodium Channels Is Determined by the Pore Loop of Domain 3. Mol. Pharmacol. 2006, 70, 340–347. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Yarov-Yarovoy, V.; Scheuer, T.; Karbat, I.; Cohen, L.; Gordon, D.; Gurevitz, M.; Catterall, W.A. Mapping the Interaction Site for a β-Scorpion Toxin in the Pore Module of Domain III of Voltage-Gated Na+ Channels. J. Biol. Chem. 2012, 287, 30719–30728. [Google Scholar] [CrossRef]
- Sokolov, S.; Kraus, R.L.; Scheuer, T.; Catterall, W.A. Inhibition of Sodium Channel Gating by Trapping the Domain II Voltage Sensor with Protoxin II. Mol. Pharmacol. 2008, 73, 1020–1028. [Google Scholar] [CrossRef]
- Cestèle, S.; Qu, Y.; Rogers, J.C.; Rochat, H.; Scheuer, T.; Catterall, W.A. Voltage Sensor–Trapping. Neuron 1998, 21, 919–931. [Google Scholar] [CrossRef]
- Cestèle, S.; Yarov-Yarovoy, V.; Qu, Y.; Sampieri, F.; Scheuer, T.; Catterall, W.A. Structure and Function of the Voltage Sensor of Sodium Channels Probed by a β-Scorpion Toxin. J. Biol. Chem. 2006, 281, 21332–21344. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Yarov-Yarovoy, V.; Scheuer, T.; Karbat, I.; Cohen, L.; Gordon, D.; Gurevitz, M.; Catterall, W.A. Structure-Function Map of the Receptor Site for β-Scorpion Toxins in Domain II of Voltage-Gated Sodium Channels. J. Biol. Chem. 2011, 286, 33641–33651. [Google Scholar] [CrossRef]
- Leipold, E.; DeBie, H.; Zorn, S.; Adolfo, B.; Olivera, B.M.; Terlau, H.; Heinemann, S.H. ΜO-Conotoxins Inhibit NaV Channels by Interfering with Their Voltage Sensors in Domain-2. Channels 2007, 1, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Zorn, S.; Leipold, E.; Hansel, A.; Bulaj, G.; Olivera, B.M.; Terlau, H.; Heinemann, S.H. The ΜO-Conotoxin MrVIA Inhibits Voltage-Gated Sodium Channels by Associating with Domain-3. FEBS Lett. 2006, 580, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Stürzebecher, A.S.; Hu, J.; Smith, E.S.J.; Frahm, S.; Santos-Torres, J.; Kampfrath, B.; Auer, S.; Lewin, G.R.; Ibañez-Tallon, I. RAPID REPORT: An in Vivo Tethered Toxin Approach for the Cell-Autonomous Inactivation of Voltage-Gated Sodium Channel Currents in Nociceptors: Pain Modulation by Tethered Toxins. J. Physiol. 2010, 588, 1695–1707. [Google Scholar] [CrossRef]
- Bulaj, G.; Zhang, M.-M.; Green, B.R.; Fiedler, B.; Layer, R.T.; Wei, S.; Nielsen, J.S.; Low, S.J.; Klein, B.D.; Wagstaff, J.D.; et al. Synthetic ΜO-Conotoxin MrVIB Blocks TTX-Resistant Sodium Channel NaV1.8 and Has a Long-Lasting Analgesic Activity. Biochemistry 2006, 45, 7404–7414. [Google Scholar] [CrossRef]
- Daly, N.L.; Ekberg, J.A.; Thomas, L.; Adams, D.J.; Lewis, R.J.; Craik, D.J. Structures of ΜO-Conotoxins from Conus Marmoreus. J. Biol. Chem. 2004, 279, 25774–25782. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.J.; Zhang, M.-M.; Azam, L.; Olivera, B.M.; Bulaj, G.; Yoshikami, D. NaV β Subunits Modulate the Inhibition of NaV 1.8 by the Analgesic Gating Modifier ΜO-Conotoxin MrVIB. J. Pharmacol. Exp. Ther. 2011, 338, 687–693. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Wilson, M.J.; Azam, L.; Gajewiak, J.; Rivier, J.E.; Bulaj, G.; Olivera, B.M.; Yoshikami, D. Co-Expression of NaV β Subunits Alters the Kinetics of Inhibition of Voltage-Gated Sodium Channels by Pore-Blocking μ-Conotoxins: NaV β Co-Expression Alters μ-Conotoxin Kinetics. Br. J. Pharmacol. 2013, 168, 1597–1610. [Google Scholar] [CrossRef]
- Pi, C.; Liu, J.; Wang, L.; Jiang, X.; Liu, Y.; Peng, C.; Chen, S.; Xu, A. Soluble Expression, Purification and Functional Identification of a Disulfide-Rich Conotoxin Derived from Conus Litteratus. J. Biotechnol. 2007, 128, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pi, C.; Liu, J.; Chen, S.; Peng, C.; Sun, D.; Zhou, M.; Xiang, H.; Ren, Z.; Xu, A. Identification and Characterization of a Novel O-Superfamily Conotoxin from Conus Litteratus: Discovery of a novel o-Superfamily Conotoxin. J. Pept. Sci. 2008, 14, 1077–1083. [Google Scholar] [CrossRef]
- Spira, M.E.; Hasson, A.; Fainzilber, M.; Gordon, D.; Zlotkin, E. Chemical and Electrophysiological Characterization of New Peptide Neurotoxins from the Venom of the Molluscivorous Snail Conus Textile Neovicarius: A Review. Isr. J. Med. Sci. 1993, 29, 530–543. [Google Scholar]
- Possani, L.D.; Becerril, B.; Delepierre, M.; Tytgat, J. Scorpion Toxins Specific for Na+-Channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Capes, D.L.; Goldschen-Ohm, M.P.; Arcisio-Miranda, M.; Bezanilla, F.; Chanda, B. Domain IV Voltage-Sensor Movement Is Both Sufficient and Rate Limiting for Fast Inactivation in Sodium Channels. J. Gen. Physiol. 2013, 142, 101–112. [Google Scholar] [CrossRef]
- Campos, F.V.; Chanda, B.; Beirão, P.S.L.; Bezanilla, F. α-Scorpion Toxin Impairs a Conformational Change That Leads to Fast Inactivation of Muscle Sodium Channels. J. Gen. Physiol. 2008, 132, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Groome, J.; Lehmann-Horn, F.; Holzherr, B. Open- and Closed-State Fast Inactivation in Sodium Channels: Differential Effects of a Site-3 Anemone Toxin. Channels 2011, 5, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Hasson, A.; Fainzilber, M.; Zlotkin, E.; Spira, M. Electrophysiological Characterization of a Novel Conotoxin That Blocks Molluscan Sodium Channels. Eur. J. Neurosci. 1995, 7, 815–818. [Google Scholar] [CrossRef]
- Hasson, A.; Shon, K.J.; Olivera, B.M.; Spira, M.E. Alterations of Voltage-Activated Sodium Current by a Novel Conotoxin from the Venom of Conus Gloriamaris. J. Neurophysiol. 1995, 73, 1295–1301. [Google Scholar] [CrossRef]
- Sarma, S.; Kumar, G.S.; Sudarslal, S.; Iengar, P.; Ramasamy, P.; Sikdar, S.; Krishnan, K.S.; Balaram, P. Solution Structure of Am2766: A Highly Hydrophobic d-Conotoxin from Conus Amadis That Inhibits Inactivation of Neuronal Voltage-Gated Sodium Channels. Chem. Biodivers. 2005, 2, 535–556. [Google Scholar] [CrossRef]
- West, P.J.; Bulaj, G.; Yoshikami, D. Effects of δ-Conotoxins PVIA and SVIE on Sodium Channels in the Amphibian Sympathetic Nervous System. J. Neurophysiol. 2005, 94, 3916–3924. [Google Scholar] [CrossRef]
- Leipold, E.; Lu, S.; Gordon, D.; Hansel, A.; Heinemann, S.H. Combinatorial Interaction of Scorpion Toxins Lqh-2, Lqh-3, and LqhαIT with Sodium Channel Receptor Sites-3. Mol. Pharmacol. 2004, 65, 685–691. [Google Scholar] [CrossRef]
- Jimenez, E.C.; Shetty, R.P.; Lirazan, M.; Rivier, J.; Walker, C.; Abogadie, F.C.; Yoshikami, D.; Cruz, L.J.; Olivera, B.M. Novel Excitatory Conus Peptides Define a New Conotoxin Superfamily: Peripherally Active Excitatory Conotoxins. J. Neurochem. 2003, 85, 610–621. [Google Scholar] [CrossRef]
- Buczek, O.; Yoshikami, D.; Watkins, M.; Bulaj, G.; Jimenez, E.C.; Olivera, B.M. Characterization of D-Amino-Acid-Containing Excitatory Conotoxins and Redefinition of the I-Conotoxin Superfamily: Defining the I-Conotoxin Superfamily. FEBS J. 2005, 272, 4178–4188. [Google Scholar] [CrossRef] [PubMed]
- Buczek, O.; Yoshikami, D.; Bulaj, G.; Jimenez, E.C.; Olivera, B.M. Post-Translational Amino Acid Isomerization. J. Biol. Chem. 2005, 280, 4247–4253. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, B.; Zhang, M.-M.; Buczek, O.; Azam, L.; Bulaj, G.; Norton, R.S.; Olivera, B.M.; Yoshikami, D. Specificity, Affinity and Efficacy of Iota-Conotoxin RXIA, an Agonist of Voltage-Gated Sodium Channels NaV1.2, 1.6 and 1.7. Biochem. Pharmacol. 2008, 75, 2334–2344. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, Y.; Abe, T.; Satake, M.; Odani, S.; Suzuki, J.; Ishikawa, K. A Novel Sodium Channel Inhibitor from Conus Geographus: Purification, Structure, and Pharmacological Properties. Biochemistry 1988, 27, 6256–6262. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.M.; Atkins, A.R.; Loughnan, M.L.; Jones, A.; Adams, D.A.; Martin, R.C.; Lewis, R.J.; Craik, D.J.; Alewood, P.F. Conotoxin TVIIA, a Novel Peptide from the Venom of Conus Tulipa: 1. Isolation, Characterization and Chemical Synthesis. Eur. J. Biochem. 2000, 267, 4642–4648. [Google Scholar] [CrossRef]
- Aguilar, M.B.; Lezama-Monfil, L.; Maillo, M.; Pedraza-Lara, H.; López-Vera, E.; Heimer de la Cotera, E.P. A Biologically Active Hydrophobic T-1-Conotoxin from the Venom of Conus Spurius. Peptides 2006, 27, 500–505. [Google Scholar] [CrossRef]
- Ruelas-Callejas, A.; Aguilar, M.B.; Arteaga-Tlecuitl, R.; Gomora, J.C.; López-Vera, E. The T-1 Conotoxin μ-SrVA from the Worm Hunting Marine Snail Conus Spurius Preferentially Blocks the Human NaV1.5 Channel. Peptides 2022, 156, 170859. [Google Scholar] [CrossRef]
- Mahdavi, S.; Kuyucak, S. Molecular Dynamics Study of Binding of Μ-Conotoxin GIIIA to the Voltage-Gated Sodium Channel Nav1.4. PLoS ONE 2014, 9, e105300. [Google Scholar] [CrossRef]
- Zhao, L.; Barber, L.M.; Hung, A. Structural and Dynamical Effects of Targeted Mutations on ΜO-Conotoxin MfVIA: Molecular Simulation Studies. J. Mol. Graph. Model. 2021, 102, 107777. [Google Scholar] [CrossRef]
- Rashid, M.; Mahdavi, S.; Kuyucak, S. Computational Studies of Marine Toxins Targeting Ion Channels. Mar. Drugs 2013, 11, 848–869. [Google Scholar] [CrossRef]
- Deplazes, E. Molecular Simulations of Disulfide-Rich Venom Peptides with Ion Channels and Membranes. Molecules 2017, 22, 362. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Chung, S.-H. Binding Modes of μ-Conotoxin to the Bacterial Sodium Channel (NaVAb). Biophys. J. 2012, 102, 483–488. [Google Scholar] [CrossRef]
- Korkosh, V.S.; Zhorov, B.S.; Tikhonov, D.B. Folding Similarity of the Outer Pore Region in Prokaryotic and Eukaryotic Sodium Channels Revealed by Docking of Conotoxins GIIIA, PIIIA, and KIIIA in a NavAb-Based Model of Nav1.4. J. Gen. Physiol. 2014, 144, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Mahdavi, S.; Kuyucak, S. Computational Study of Binding of μ-Conotoxin GIIIA to Bacterial Sodium Channels NaV Ab and NaV Rh. Biochemistry 2016, 55, 1929–1938. [Google Scholar] [CrossRef]
- Finol-Urdaneta, R.K.; McArthur, J.R.; Korkosh, V.S.; Huang, S.; McMaster, D.; Glavica, R.; Tikhonov, D.B.; Zhorov, B.S.; French, R.J. Extremely Potent Block of Bacterial Voltage-Gated Sodium Channels by µ-Conotoxin PIIIA. Mar. Drugs 2019, 17, 510. [Google Scholar] [CrossRef]
- Chen, R.; Robinson, A.; Chung, S.-H. Mechanism of μ-Conotoxin PIIIA Binding to the Voltage-Gated Na+ Channel NaV1.4. PLoS ONE 2014, 9, e93267. [Google Scholar] [CrossRef]
- Tietze, A.A.; Tietze, D.; Ohlenschläger, O.; Leipold, E.; Ullrich, F.; Kühl, T.; Mischo, A.; Buntkowsky, G.; Görlach, M.; Heinemann, S.H.; et al. Structurally Diverse μ-Conotoxin PIIIA Isomers Block Sodium Channel NaV 1.4. Angew. Chem. Int. Ed. 2012, 51, 4058–4061. [Google Scholar] [CrossRef] [PubMed]
- Paul George, A.A.; Heimer, P.; Leipold, E.; Schmitz, T.; Kaufmann, D.; Tietze, D.; Heinemann, S.H.; Imhof, D. Effect of Conformational Diversity on the Bioactivity of Μ-Conotoxin PIIIA Disulfide Isomers. Mar. Drugs 2019, 17, 390. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep Venomics Reveals the Mechanism for Expanded Peptide Diversity in Cone Snail Venom. Mol. Cell. Proteomics 2013, 12, 312–329. [Google Scholar] [CrossRef]
- Gorson, J.; Holford, M. Small Packages, Big Returns: Uncovering the Venom Diversity of Small Invertebrate Conoidean Snails. Integr. Comp. Biol. 2016, 56, 962–972. [Google Scholar] [CrossRef]
- Gao, B.; Peng, C.; Yang, J.; Yi, Y.; Zhang, J.; Shi, Q. Cone Snails: A Big Store of Conotoxins for Novel Drug Discovery. Toxins 2017, 9, 397. [Google Scholar] [CrossRef]
- Himaya, S.; Lewis, R. Venomics-Accelerated Cone Snail Venom Peptide Discovery. Int. J. Mol. Sci. 2018, 19, 788. [Google Scholar] [CrossRef] [PubMed]
- Koua, D.; Ebou, A.; Dutertre, S. Improved Prediction of Conopeptide Superfamilies with ConoDictor 2.0. Bioinform. Adv. 2021, 1, vbab011. [Google Scholar] [CrossRef] [PubMed]
- Espiritu, M.J.; Cabalteja, C.C.; Sugai, C.K.; Bingham, J.-P. Incorporation of Post-Translational Modified Amino Acids as an Approach to Increase Both Chemical and Biological Diversity of Conotoxins and Conopeptides. Amino Acids 2014, 46, 125–151. [Google Scholar] [CrossRef]
- Fu, Y.; Li, C.; Dong, S.; Wu, Y.; Zhangsun, D.; Luo, S. Discovery Methodology of Novel Conotoxins from Conus Species. Mar. Drugs 2018, 16, 417. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S. Enhancing the Therapeutic Potential of Peptide Toxins. Expert Opin. Drug Discov. 2017, 12, 611–623. [Google Scholar] [CrossRef]
- Bernáldez-Sarabia, J.; Figueroa-Montiel, A.; Dueñas, S.; Cervantes-Luévano, K.; Beltrán, J.; Ortiz, E.; Jiménez, S.; Possani, L.; Paniagua-Solís, J.; Gonzalez-Canudas, J.; et al. The Diversified O-Superfamily in Californiconus Californicus Presents a Conotoxin with Antimycobacterial Activity. Toxins 2019, 11, 128. [Google Scholar] [CrossRef]
- Ebou, A.; Koua, D.; Addablah, A.; Kakou-Ngazoa, S.; Dutertre, S. Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails. Biomedicines 2021, 9, 344. [Google Scholar] [CrossRef]
- Primon-Barros, M.; José Macedo, A. Animal Venom Peptides: Potential for New Antimicrobial Agents. Curr. Top. Med. Chem. 2017, 17, 1119–1156. [Google Scholar] [CrossRef]





| Superfamily | Conotoxins | Cysteine Pattern | Loops | Connectivity | Fold Pattern | Motif |
|---|---|---|---|---|---|---|
| M | μ-conotoxin | CC---C---C---CC | 3 | 1–4, 2–5, 3–6 | B | - |
| O | μO-conotoxin | C---C---CC---C---C | 4 | 1–4, 2–5, 3–6 | C | ICK |
| O | δ-conotoxin | C---C---CC---C---C | 4 | 1–4, 2–5, 3–6 | C | ICK |
| I | ι-conotoxin | C---C---CC---CC---C---C | 5 | 1–4, 2–5, 3–6, 5–8 | C | ICK |
| Toxin | Conus Species | Sequence | Reference |
|---|---|---|---|
| M4 branch | ---CC------C----C----CC--- | ||
| GIIIA | C. geographus | -RDCCTOOKK-CKDRQCKOQRCCA- | [97,98,99] |
| GIIIB | C. geographus | -RDCCTOORK-CKDRRCKOMKCCA- | [97,98,99] |
| GIIIC | C. geographus | -RDCCTOOKK-CKDRRCKOLKCCA- | [99] |
| PIIIA | C. purpurascens | ZRLCCGFOKS-CRSRQCKOHRCC- | [160] |
| TIIIA | C. tulipa | RHGCCKGOKG-CSSRECROQHCC- | [161] |
| SxIIIA | C. striolatus | --RCCTGKKGSCSGRACKNLKCCA- | [162] |
| SxIIIB | C. striolatus | -QKCCTGKKGSCSGRACKNLRCCA- | [162] |
| TsIIIA | C. tessulatus | --GCCRWP---CPSR-CGMARCCSS | [163] |
| M5 branch | ----CC--------C----C-----CC--- | ||
| SmIIIA | C. stercusmuscarum | --ZRCCN---GRRGCSSRWCRDHSRCC--- | [164] |
| KIIIA | C. kinoshitai | ----CCN-------CSSKWCRDHSRCC--- | [165] |
| SIIIA | C. striatus | --ZNCCN---G--GCSSKWCRDHARCC--- | [165,166] |
| CIIIA | C. catus | --GRCCE---GPNGCSSRWCKDHARCC--- | [167] |
| CnIIIA | C. consors | --GRCCD---VPNACSGRWCRDHAQCC--- | [167] |
| CnIIIB | C. consors | --ZGCCG---EPNLCFTRWCRNNARCCRQQ | [167] |
| MIIIA | C. magus | --ZGCCN---VPNGCSGRWCRDHAQCC--- | [167] |
| SIIIB | C. striatus | --ZNCCN-----GGCSSKWCKGHARCC--- | [168] |
| BuIIIA | C. bullatus | VTDRCCK---GKREC-GRWCRDHSRCC--- | [169] |
| BuIIIB | C. bullatus | VGERCCK--NGKRGC-GRWCRDHSRCC--- | [169] |
| BuIIIC | C. bullatus | IVDRCCNKGNGKRGC-SRWCRDHSRCC--- | [169] |
| KIIIB | C. kinoshitai | --NGCCN-------CSSKWCRDHSRCC--- | [170] |
| CnIIIC | C. consors | --ZGCCN---GPKGCSSKWCRDHARCC--- | [171] |
| SxIIIC | C. striolatus | --RGCCN---GRGGCSSRWCRDHARCC--- | [172] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groome, J.R. Historical Perspective of the Characterization of Conotoxins Targeting Voltage-Gated Sodium Channels. Mar. Drugs 2023, 21, 209. https://doi.org/10.3390/md21040209
Groome JR. Historical Perspective of the Characterization of Conotoxins Targeting Voltage-Gated Sodium Channels. Marine Drugs. 2023; 21(4):209. https://doi.org/10.3390/md21040209
Chicago/Turabian StyleGroome, James R. 2023. "Historical Perspective of the Characterization of Conotoxins Targeting Voltage-Gated Sodium Channels" Marine Drugs 21, no. 4: 209. https://doi.org/10.3390/md21040209
APA StyleGroome, J. R. (2023). Historical Perspective of the Characterization of Conotoxins Targeting Voltage-Gated Sodium Channels. Marine Drugs, 21(4), 209. https://doi.org/10.3390/md21040209