
Seasonal Molecular Difference in Fibrillar Collagen Extracts Derived from the Marine Sponge Chondrosia reniformis (Nardo, 1847) and Their Impact on Its Derived Biomaterials

 ,
,  ,
,  ,
,  ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
2.1. Biochemical Characterization of Sponge Fibrillar Collagen Extracts
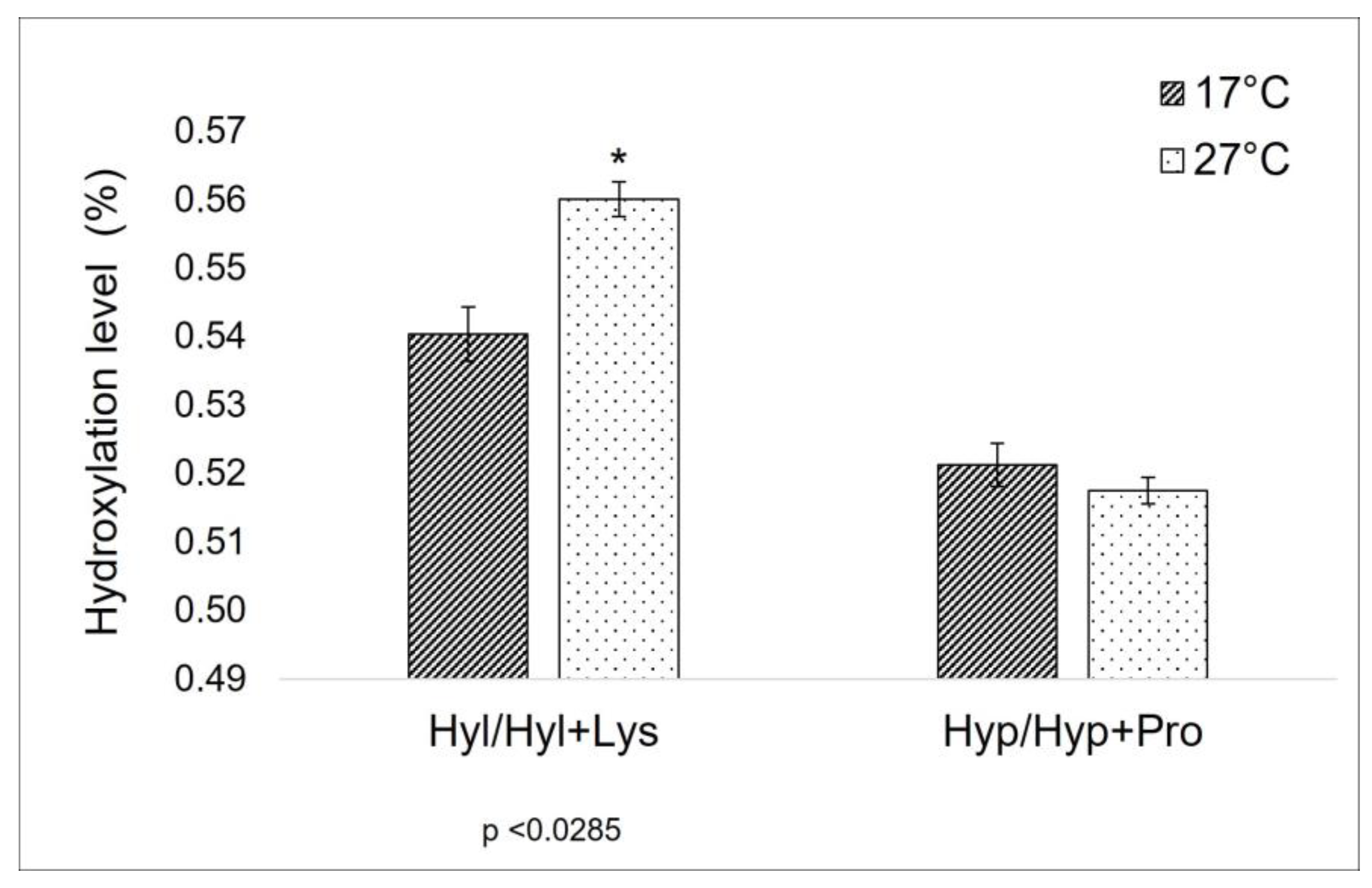
2.1.1. Evaluation of Amino-Acids Percentage Composition
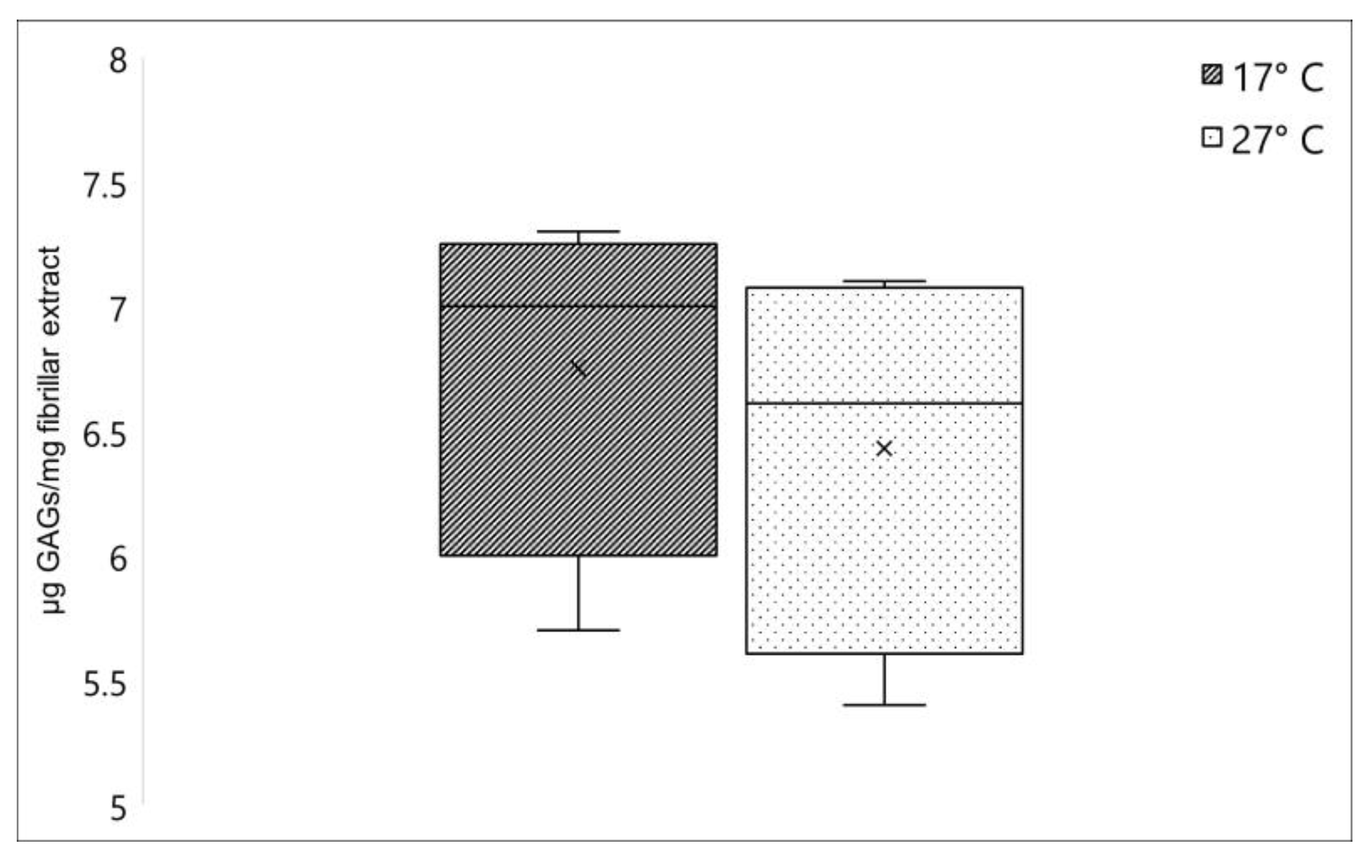
2.1.2. Glycosaminoglycans Quantification
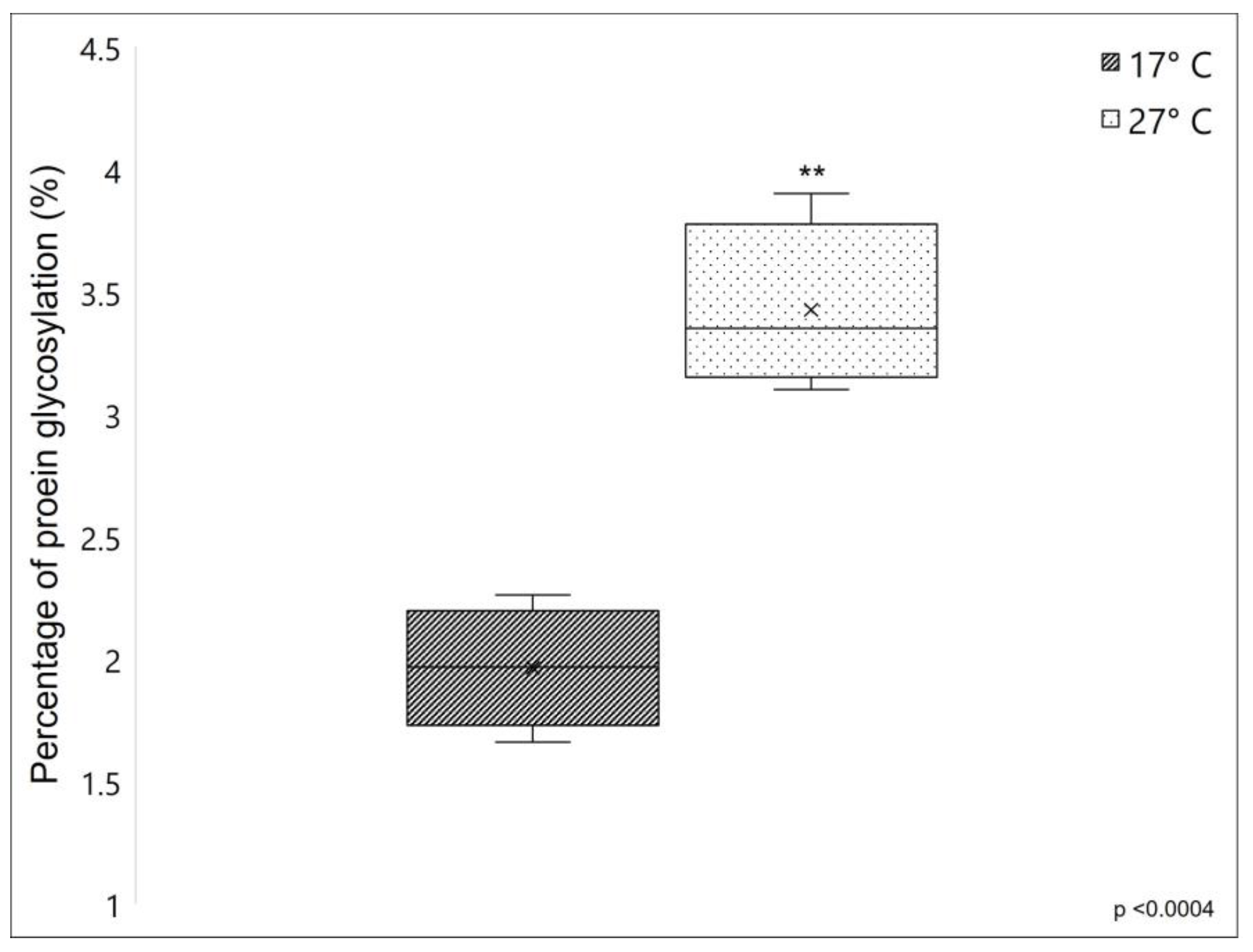
2.1.3. Proteins Glycosylation
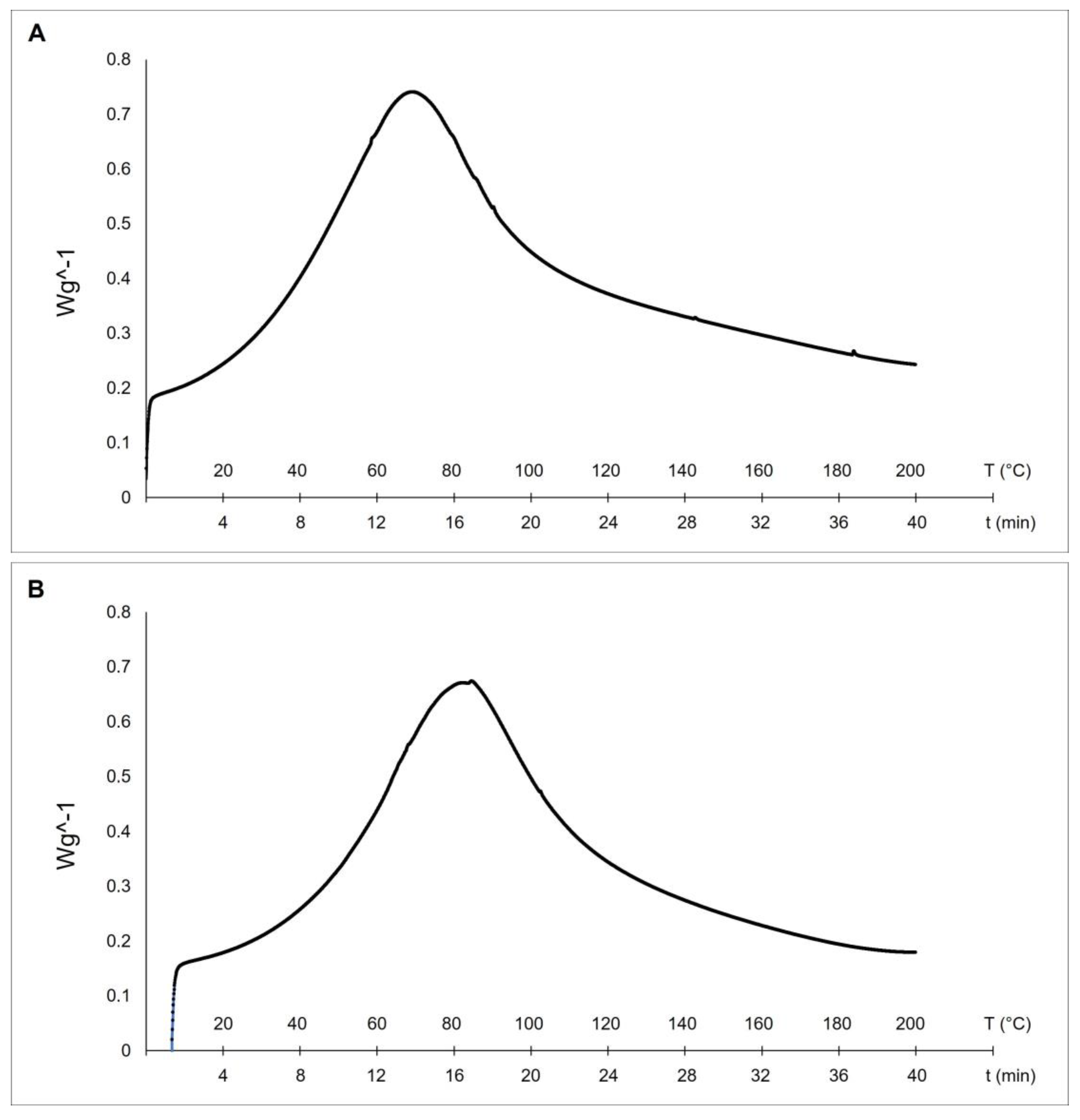
2.2. Thermal Stability Analysis
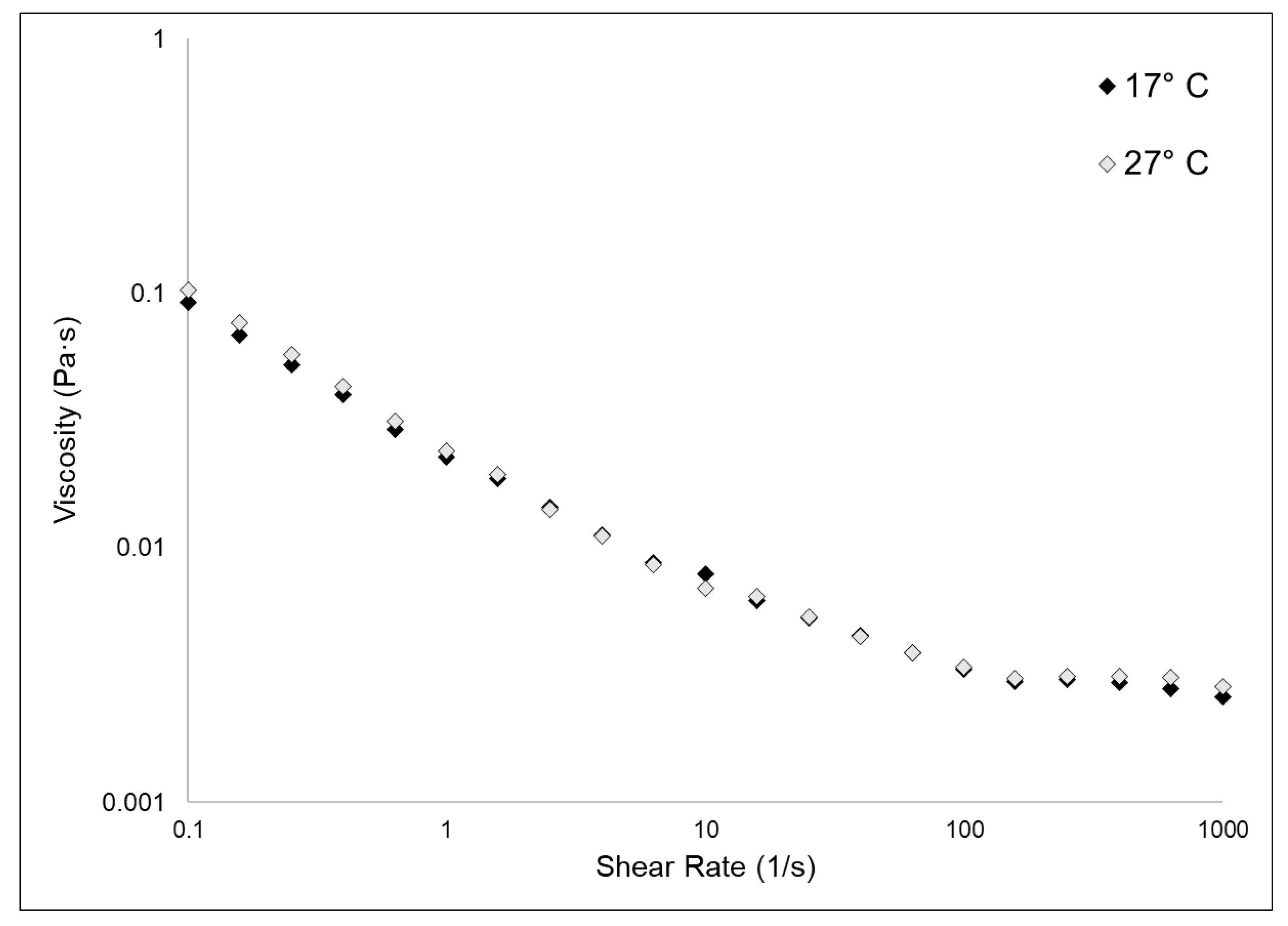
2.3. Viscosity Evaluation
2.4. Collagen Membranes Characterization
2.4.1. Membranes Surface Morphologies
2.4.2. Dynamic Mechanical and Thermal Analysis
2.4.3. Membranes’ Antioxidant Properties
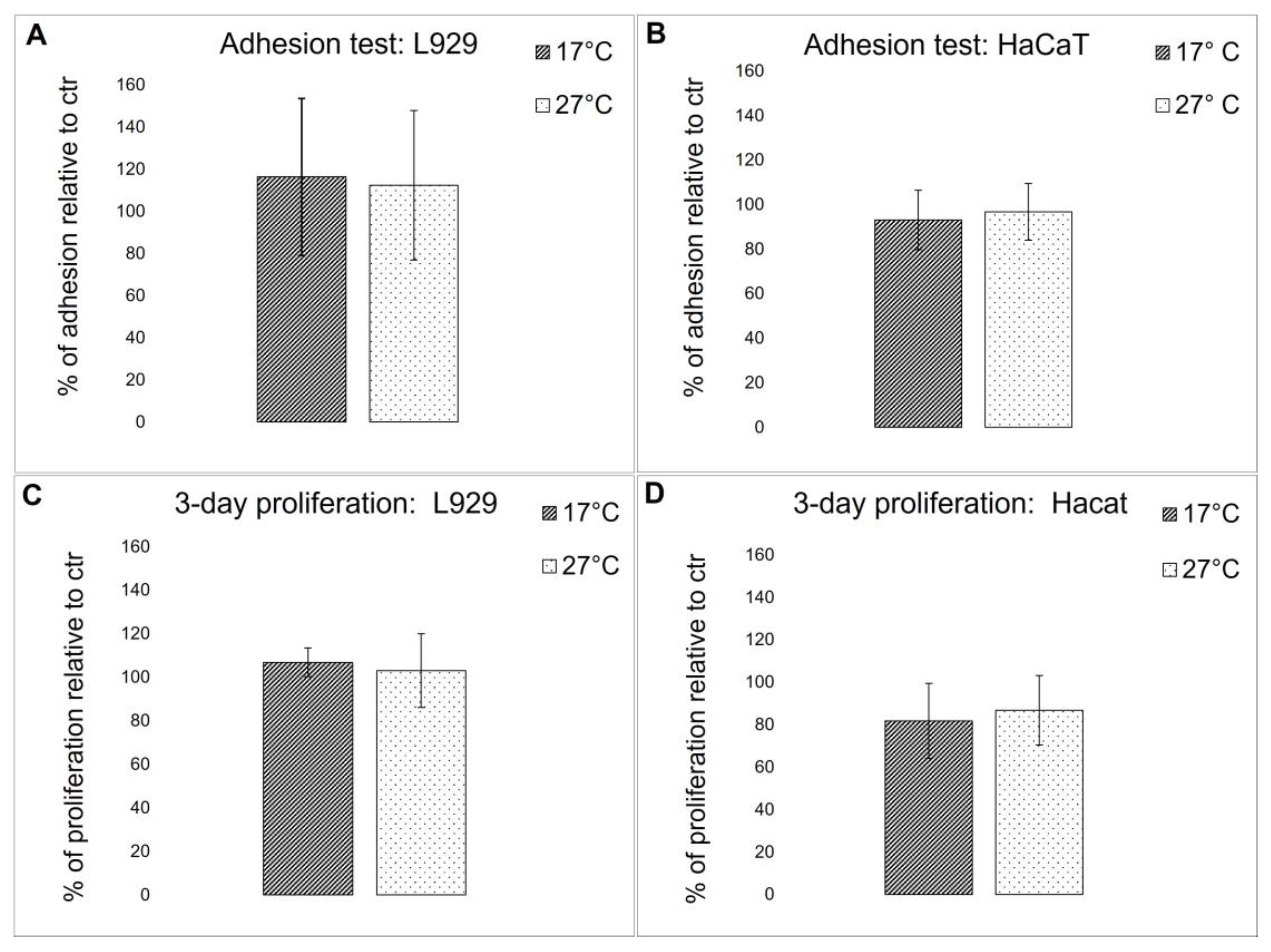
2.4.4. Biocompatibility Evaluation
3. Materials and Methods
3.1. Chemicals
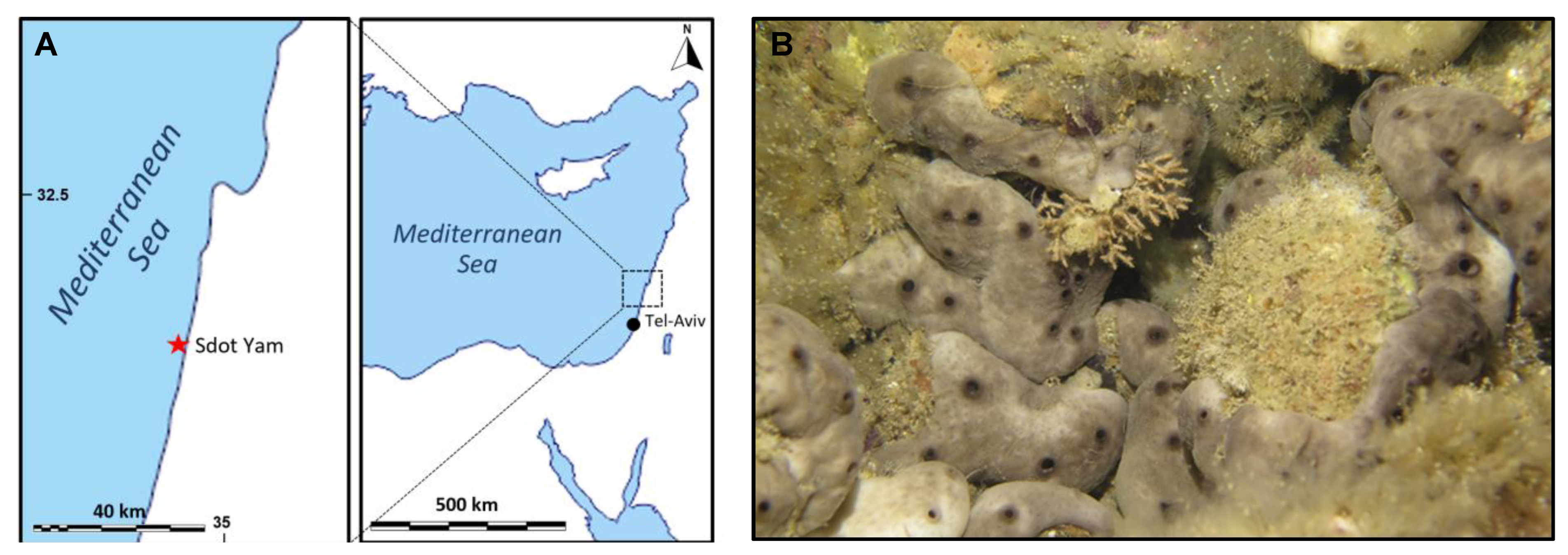
3.2. Sponge Sampling
3.3. Fibrillar Collagen Extraction
3.4. Biochemical Characterization of Collagen Extracts
3.4.1. Evaluation of Amino-Acids Percentage Composition
3.4.2. Glycosaminoglycans (GAGs) Quantification
3.4.3. Evaluation of the Percentage of Protein Glycosylation
3.4.4. Differential Scanning Calorimetry (DSC) Analysis
3.4.5. Viscosity Evaluation
3.5. 2D Sponge Collagen Membranes Production
3.6. Sponge Collagen Membranes Characterization
3.6.1. Field-Emission Scanning Electron Microscope (FE-SEM) Observation
3.6.2. DPPH Radical Scavenging Activity
3.6.3. Dynamic Mechanical Analysis (DMA) and Dynamic Mechanical-Thermal Analysis (DMTA)
3.7. Adhesion and Viability Tests on Cells
3.7.1. Cell Cultures
3.7.2. Cell Adhesion and Cell Proliferation
3.8. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lim, Y.-S.; Ok, Y.-J.; Hwang, S.-Y.; Kwak, J.-Y.; Yoon, S. Marine Collagen as A Promising Biomaterial for Biomedical Applications. Mar. Drugs 2019, 17, 467. [Google Scholar] [CrossRef] [PubMed]
- Coppola, D.; Oliviero, M.; Vitale, G.A.; Lauritano, C.; D’Ambra, I.; Iannace, S.; De Pascale, D. Marine Collagen from Alternative and Sustainable Sources: Extraction, Processing and Applications. Mar. Drugs 2020, 18, 214. [Google Scholar] [CrossRef] [PubMed]
- Pozzolini, M.; Scarfì, S.; Gallus, L.; Castellano, M.; Vicini, S.; Cortese, K.; Gagliani, M.C.; Bertolino, M.; Costa, G.; Giovine, M. Production, Characterization and Biocompatibility Evaluation of Collagen Membranes Derived from Marine Sponge Chondrosia reniformis Nardo, 1847. Mar. Drugs 2018, 16, 111. [Google Scholar] [CrossRef]
- Pozzolini, M.; Tassara, E.; Dodero, A.; Castellano, M.; Vicini, S.; Ferrando, S.; Aicardi, S.; Cavallo, D.; Bertolino, M.; Petrenko, I.; et al. Potential Biomedical Applications of Collagen Filaments derived from the Marine Demosponges Ircinia oros (Schmidt, 1864) and Sarcotragus foetidus (Schmidt, 1862). Mar. Drugs 2021, 19, 563. [Google Scholar] [CrossRef]
- Ferrario, C.; Rusconi, F.; Pulaj, A.; Macchi, R.; Landini, P.; Paroni, M.; Colombo, G.; Martinello, T.; Melotti, L.; Gomiero, C.; et al. From Food Waste to Innovative Biomaterial: Sea Urchin-Derived Collagen for Applications in Skin Regenerative Medicine. Mar. Drugs 2020, 18, 414. [Google Scholar] [CrossRef]
- Subhan, F.; Ikram, M.; Shehzad, A.; Ghafoor, A. Marine Collagen: An Emerging Player in Biomedical applications. J. Food Sci. Technol. 2015, 52, 4703–4707. [Google Scholar] [CrossRef]
- Bielajew, B.J.; Hu, J.C.; Athanasiou, K.A. Collagen: Quantification, biomechanics and role of minor subtypes in cartilage. Nat. Rev. Mater. 2020, 5, 730–747. [Google Scholar] [CrossRef]
- Exposito, J.-Y.; Cluzel, C.; Garrone, R.; Lethias, C. Evolution of collagens. Anat. Rec. 2002, 268, 302–316. [Google Scholar] [CrossRef]
- Geahchan, S.; Baharlouei, P.; Rahman, A. Marine Collagen: A Promising Biomaterial for Wound Healing, Skin Anti-Aging, and Bone Regeneration. Mar. Drugs 2022, 20, 61. [Google Scholar] [CrossRef]
- Wilkie, I.C. Mutable Collagenous Tissue: Overview and Biotechnological Perspective. In Echinodermata; Matranga, V., Ed.; Progress in Molecular and Subcellular Biology; Springer: Berlin/Heidelberg, Germany, 2005; pp. 221–250. ISBN 978-3-540-27683-8. [Google Scholar]
- Haq, F.; Ahmed, N.; Qasim, M. Comparative genomic analysis of collagen gene diversity. 3 Biotech 2019, 9, 83. [Google Scholar] [CrossRef]
- Birch, H.L. Extracellular Matrix and Ageing. In Biochemistry and Cell Biology of Ageing: Part I Biomedical Science; Harris, J.R., Korolchuk, V.I., Eds.; Subcellular Biochemistry; Springer: Singapore, 2018; pp. 169–190. ISBN 9789811328350. [Google Scholar]
- Fujii, K.K.; Taga, Y.; Takagi, Y.K.; Masuda, R.; Hattori, S.; Koide, T. The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature. Int. J. Mol. Sci. 2022, 23, 2040. [Google Scholar] [CrossRef]
- Tonelotto, V.; Trapani, V.; Bretaud, S.; Heumüller, S.E.; Wagener, R.; Ruggiero, F.; Bonaldo, P. Spatio-temporal expression and distribution of collagen VI during zebrafish development. Sci. Rep. 2019, 9, 19851. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Bächinger, H.P. A molecular ensemble in the rER for procollagen maturation. Biochim. Biophys. Acta BBA Mol. Cell Res. 2013, 1833, 2479–2491. [Google Scholar] [CrossRef]
- Basak, T.; Vega-Montoto, L.; Zimmerman, L.J.; Tabb, D.L.; Hudson, B.G.; Vanacore, R.M. Comprehensive Characterization of Glycosylation and Hydroxylation of Basement Membrane Collagen IV by High-Resolution Mass Spectrometry. J. Proteome Res. 2015, 15, 245–258. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Taga, Y.; Tanaka, K.; Hattori, S.; Mizuno, K. In-depth correlation analysis demonstrates that 4-hydroxyproline at the Yaa position of Gly-Xaa-Yaa repeats dominantly stabilizes collagen triple helix. Matrix Biol. Plus 2021, 10, 100067. [Google Scholar] [CrossRef]
- Ahmed, R.; Haq, M.; Chun, B.-S. Characterization of marine derived collagen extracted from the by-products of bigeye tuna (Thunnus obesus). Int. J. Biol. Macromol. 2019, 135, 668–676. [Google Scholar] [CrossRef]
- Terajima, M.; Taga, Y.; Nakamura, T.; Guo, H.-F.; Kayashima, Y.; Maeda-Smithies, N.; Parag-Sharma, K.; Kim, J.S.; Amelio, A.L.; Mizuno, K.; et al. Lysyl Hydroxylase 2 Mediated Collagen Post-Translational Modifications and Functional Outcomes. Sci. Rep. 2022, 12, 14256. [Google Scholar] [CrossRef]
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef]
- Nicklas, M.; Schatton, W.; Heinemann, S.; Hanke, T.; Kreuter, J. Preparation and characterization of marine sponge collagen nanoparticles and employment for the transdermal delivery of 17β-estradiol-hemihydrate. Drug Dev. Ind. Pharm. 2009, 35, 1035–1042. [Google Scholar] [CrossRef]
- Pozzolini, M.; Millo, E.; Oliveri, C.; Mirata, S.; Salis, A.; Damonte, G.; Arkel, M.; Scarfì, S. Elicited ROS Scavenging Activity, Photoprotective, and Wound-Healing Properties of Collagen-Derived Peptides from the Marine Sponge Chondrosia reniformis. Mar. Drugs 2018, 16, 465. [Google Scholar] [CrossRef] [PubMed]
- Fassini, D.; Duarte, A.R.C.; Reis, R.L.; Silva, T.H. Bioinspiring Chondrosia reniformis (Nardo, 1847) Collagen-Based Hydrogel: A New Extraction Method to Obtain a Sticky and Self-Healing Collagenous Material. Mar. Drugs 2017, 15, 380. [Google Scholar] [CrossRef]
- Gökalp, M.; Wijgerde, T.; Sarà, A.; De Goeij, J.M.; Osinga, R. Development of an Integrated Mariculture for the Collagen-Rich Sponge Chondrosia reniformis. Mar. Drugs 2019, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- Gökalp, M.; Kooistra, T.; Rocha, M.S.; Silva, T.H.; Osinga, R.; Murk, A.J.; Wijgerde, T. The Effect of Depth on the Morphology, Bacterial Clearance, and Respiration of the Mediterranean Sponge Chondrosia reniformis (Nardo, 1847). Mar. Drugs 2020, 18, 358. [Google Scholar] [CrossRef] [PubMed]
- Orel, B.; Giovine, M.; Ilan, M. On the Path to Thermo-Stable Collagen: Culturing the Versatile Sponge Chondrosia reniformis. Mar. Drugs 2021, 19, 669. [Google Scholar] [CrossRef]
- Rodríguez, F.; Morán, L.; González, G.; Troncoso, E.; Zúñiga, R.N. Collagen Extraction from Mussel Byssus: A New Marine Collagen Source with Physicochemical Properties of Industrial Interest. J. Food Sci. Technol. 2017, 54, 1228–1238. [Google Scholar] [CrossRef]
- Minh Thuy, L.T.; Okazaki, E.; Osako, K. Isolation and Characterization of Acid-Soluble Collagen from the Scales of Marine Fishes from Japan and Vietnam. Food Chem. 2014, 149, 264–270. [Google Scholar] [CrossRef]
- Heinemann, S.; Ehrlich, H.; Douglas, T.; Heinemann, C.; Worch, H.; Schatton, W.; Hanke, T. Ultrastructural Studies on the Collagen of the Marine Sponge Chondrosia reniformis Nardo. Biomacromolecules 2007, 8, 3452–3457. [Google Scholar] [CrossRef]
- Bretscher, L.E.; Jenkins, C.L.; Taylor, K.M.; DeRider, M.L.; Raines, R.T. Conformational Stability of Collagen Relies on a Stereoelectronic Effect. J. Am. Chem. Soc. 2001, 123, 777–778. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E.H.; et al. A Guide to the Composition and Functions of the Extracellular Matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Fassini, D.; Parma, L.; Wilkie, I.C.; Bavestrello, G.; Bonasoro, F.; Candia Carnevali, M.D. Ecophysiology of Mesohyl Creep in the Demosponge Chondrosia reniformis (Porifera: Chondrosida). J. Exp. Mar. Biol. Ecol. 2012, 428, 24–31. [Google Scholar] [CrossRef]
- Wilkie, I.C.; Parma, L.; Bonasoro, F.; Bavestrello, G.; Cerrano, C.; Candia Carnevali, M.D. Mechanical Adaptability of a Sponge Extracellular Matrix: Evidence for Cellular Control of Mesohyl Stiffness in Chondrosia reniformis Nardo. J. Exp. Biol. 2006, 209, 4436–4443. [Google Scholar] [CrossRef]
- Misevic, G.; Garbarino, E. Glycan-to-Glycan Binding: Molecular Recognition through Polyvalent Interactions Mediates Specific Cell Adhesion. Molecules 2021, 26, 397. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in Health and Disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Mann, K.; Mechling, D.E.; Bächinger, H.P.; Eckerskorn, C.; Gaill, F.; Timpl, R. Glycosylated Threonine but Not 4-Hydroxyproline Dominates the Triple Helix Stabilizing Positions in the Sequence of a Hydrothermal Vent Worm Cuticle Collagen. J. Mol. Biol. 1996, 261, 255–266. [Google Scholar] [CrossRef]
- Bann, J.G.; Peyton, D.H.; Bächinger, H.P. Sweet Is Stable: Glycosylation Stabilizes Collagen. FEBS Lett. 2000, 473, 237–240. [Google Scholar] [CrossRef]
- Kotch, F.W.; Guzei, I.A.; Raines, R.T. Stabilization of the Collagen Triple Helix by O-Methylation of Hydroxyproline Residues. J. Am. Chem. Soc. 2008, 130, 2952–2953. [Google Scholar] [CrossRef]
- Mezger, T.G. The Rheology Handbook: For Users of Rotational and Oscillatory Rheometers, 2nd ed.; Vincentz Network: Hannover, Germany, 2006; ISBN 978-3-86-630842-8. [Google Scholar]
- Depalle, B.; Qin, Z.; Shefelbine, S.J.; Buehler, M.J. Influence of cross-link structure, density and mechanical properties in the mesoscale deformation mechanisms of collagen fibrils. J. Mech. Behav. Biomed. Mater. 2015, 52, 1–13. [Google Scholar] [CrossRef]
- Yamauchi, M.; Woodley, D.T.; Mechanic, G.L. Aging and Cross-Linking of Skin Collagen. Biochem. Biophys. Res. Commun. 1988, 152, 898–903. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Coppari, M.; Bartolucci, I.; Bo, M.; Betti, F.; Bertolino, M.; Calcinai, B.; Cerrano, C.; De Grandis, G.; Bavestrello, G. Temporal Variations in Growth and Reproduction of Tedania anhelans and Chondrosia reniformis in the North Adriatic Sea. In Ancient Animals, New Challenges: Developments in Sponge Research; Maldonado, M., Turon, X., Becerro, M., Jesús Uriz, M., Eds.; Developments in Hydrobiology; Springer: Dordrecht, The Netherlands, 2012; pp. 299–313. ISBN 978-94-007-4688-6. [Google Scholar]
- Dong, Y.; Dai, Z. Physicochemical, Structural and Antioxidant Properties of Collagens from the Swim Bladder of Four Fish Species. Mar. Drugs 2022, 20, 550. [Google Scholar] [CrossRef]
- Chen, K.; Yang, X.; Huang, Z.; Jia, S.; Zhang, Y.; Shi, J.; Hong, H.; Feng, L.; Luo, Y. Modification of Gelatin Hydrolysates from Grass Carp (Ctenopharyngodon idellus) Scales by Maillard Reaction: Antioxidant Activity and Volatile Compounds. Food Chem. 2019, 295, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Parma, L.; Fassini, D.; Bavestrello, G.; Wilkie, I.C.; Bonasoro, F.; Candia Carnevali, M.D. Ecology and Physiology of Mesohyl Creep in Chondrosia reniformis; 2007 Porifera Research: Biodiversity, Innovation & Sustainability: Série Livros, 28; Museu Nacional: Rio de Janeiro, Brazil, 2007; pp. 503–508. [Google Scholar]
- Gross, J.; Sokal, Z.; Rougvie, M. Structural and Chemical Studies on the Connective Tissue of Marine Sponges. J. Histochem. Cytochem. 1956, 4, 227–246. [Google Scholar] [CrossRef]
- Frazier, S.B.; Roodhouse, K.A.; Hourcade, D.E.; Zhang, L. The Quantification of Glycosaminoglycans: A Comparison of HPLC, Carbazole, and Alcian Blue Methods. Open Glycosci. 2008, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AA | 17 °C | S.D. | 27 °C | S.D. |
|---|---|---|---|---|
| Mean (%) | Mean (%) | |||
| Asp | 7.56 | 0.09 | 7.47 | 0.24 |
| Glu | 7.34 | 0.15 | 7.24 | 0.16 |
| Ser | 4.76 | 0.58 | 4.71 | 0.36 |
| Gly | 46.59 | 2.33 | 49.48 | 2.43 |
| Thr | 2.43 | 0.17 | 2.08 | 0.18 |
| Ala + Arg | 7.68 | 0.15 | 7.91 | 0.19 |
| Tyr | 0.97 | 0.20 | 0.80 | 0.08 |
| Val | 1.27 | 0.17 | 1.04 | 0.12 |
| Met | 1.04 | 0.35 | 0.78 | 0.07 |
| Hyl | 2.40 | 0.31 | 2.13 | 0.15 |
| Phe | 1.41 | 0.17 | 1.26 | 0.09 |
| Ile | 0.99 | 0.17 | 0.82 | 0.08 |
| Leu | 2.05 | 0.14 | 1.90 | 0.11 |
| Lys | 2.04 | 0.28 | 1.71 | 0.14 |
| Hyp | 5.98 | 0.06 | 5.51 | 0.19 |
| Pro | 5.50 | 0.13 | 5.14 | 0.26 |
| Sample | η0 (mPa·s) | η∞ (mPa·s) |
|---|---|---|
| 17 °C | 92.23 | 2.58 |
| 27 °C | 103.01 | 2.84 |
| 17 °C | S.D. | 27 °C | S.D. | |
|---|---|---|---|---|
| E’ @ 1 Hz, 25 °C (MPa) | 2368.84 | 73.05 | 2105.30 | 45.78 |
| E” @ 1 Hz, 25 °C (MPa) | 181.64 | 33.61 | 138.59 | 34.22 |
| E’ @ 1 Hz, 37 °C (MPa) | 2156.03 | 53.25 | 1864.24 | 28.99 |
| E” @ 1 Hz, 37 °C (MPa) | 90.88 | 14.22 | 77.98 | 4.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tassara, E.; Orel, B.; Ilan, M.; Cavallo, D.; Dodero, A.; Castellano, M.; Vicini, S.; Giovine, M.; Pozzolini, M. Seasonal Molecular Difference in Fibrillar Collagen Extracts Derived from the Marine Sponge Chondrosia reniformis (Nardo, 1847) and Their Impact on Its Derived Biomaterials. Mar. Drugs 2023, 21, 210. https://doi.org/10.3390/md21040210
Tassara E, Orel B, Ilan M, Cavallo D, Dodero A, Castellano M, Vicini S, Giovine M, Pozzolini M. Seasonal Molecular Difference in Fibrillar Collagen Extracts Derived from the Marine Sponge Chondrosia reniformis (Nardo, 1847) and Their Impact on Its Derived Biomaterials. Marine Drugs. 2023; 21(4):210. https://doi.org/10.3390/md21040210
Chicago/Turabian StyleTassara, Eleonora, Boaz Orel, Micha Ilan, Dario Cavallo, Andrea Dodero, Maila Castellano, Silvia Vicini, Marco Giovine, and Marina Pozzolini. 2023. "Seasonal Molecular Difference in Fibrillar Collagen Extracts Derived from the Marine Sponge Chondrosia reniformis (Nardo, 1847) and Their Impact on Its Derived Biomaterials" Marine Drugs 21, no. 4: 210. https://doi.org/10.3390/md21040210
APA StyleTassara, E., Orel, B., Ilan, M., Cavallo, D., Dodero, A., Castellano, M., Vicini, S., Giovine, M., & Pozzolini, M. (2023). Seasonal Molecular Difference in Fibrillar Collagen Extracts Derived from the Marine Sponge Chondrosia reniformis (Nardo, 1847) and Their Impact on Its Derived Biomaterials. Marine Drugs, 21(4), 210. https://doi.org/10.3390/md21040210