
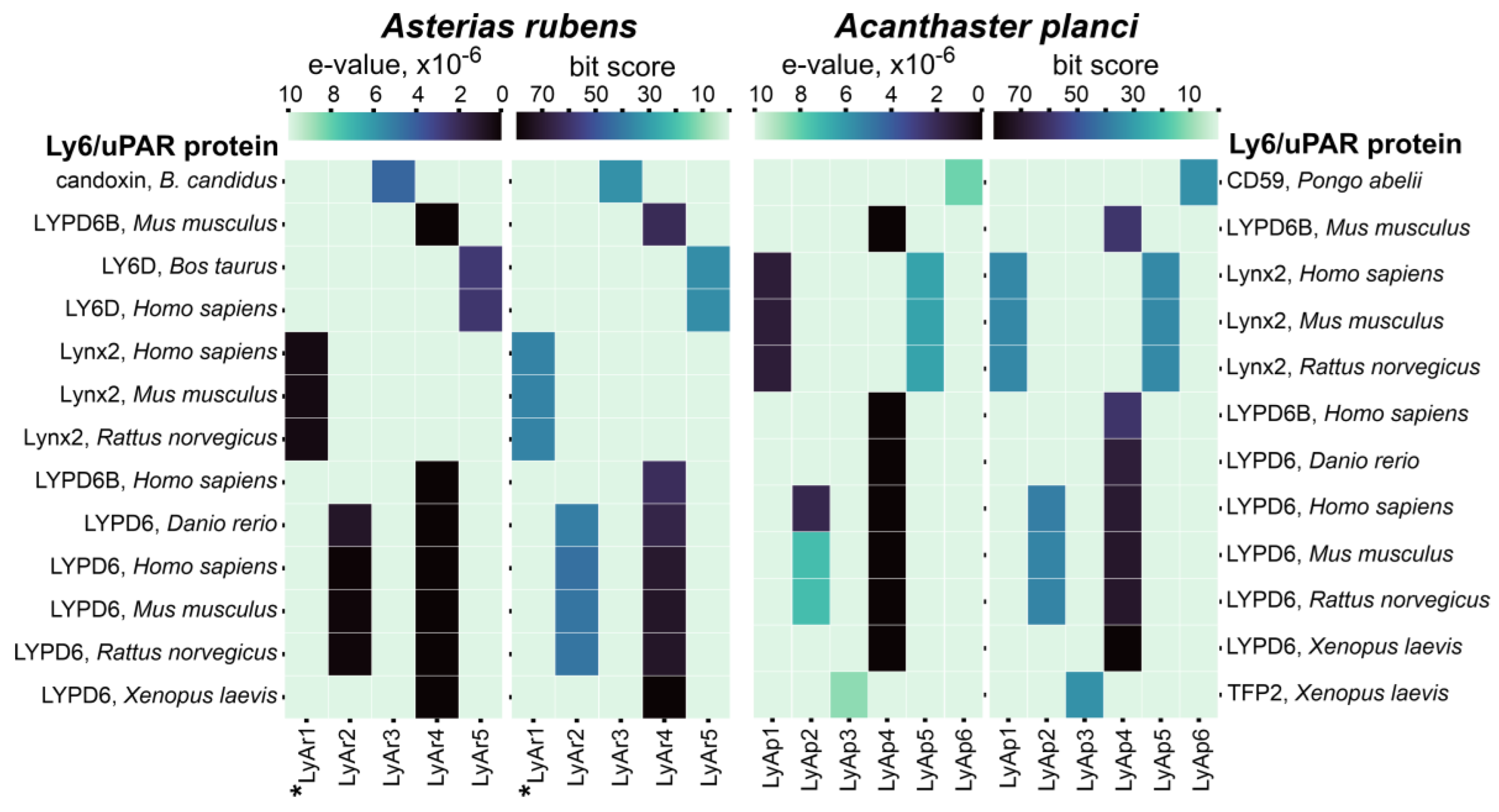
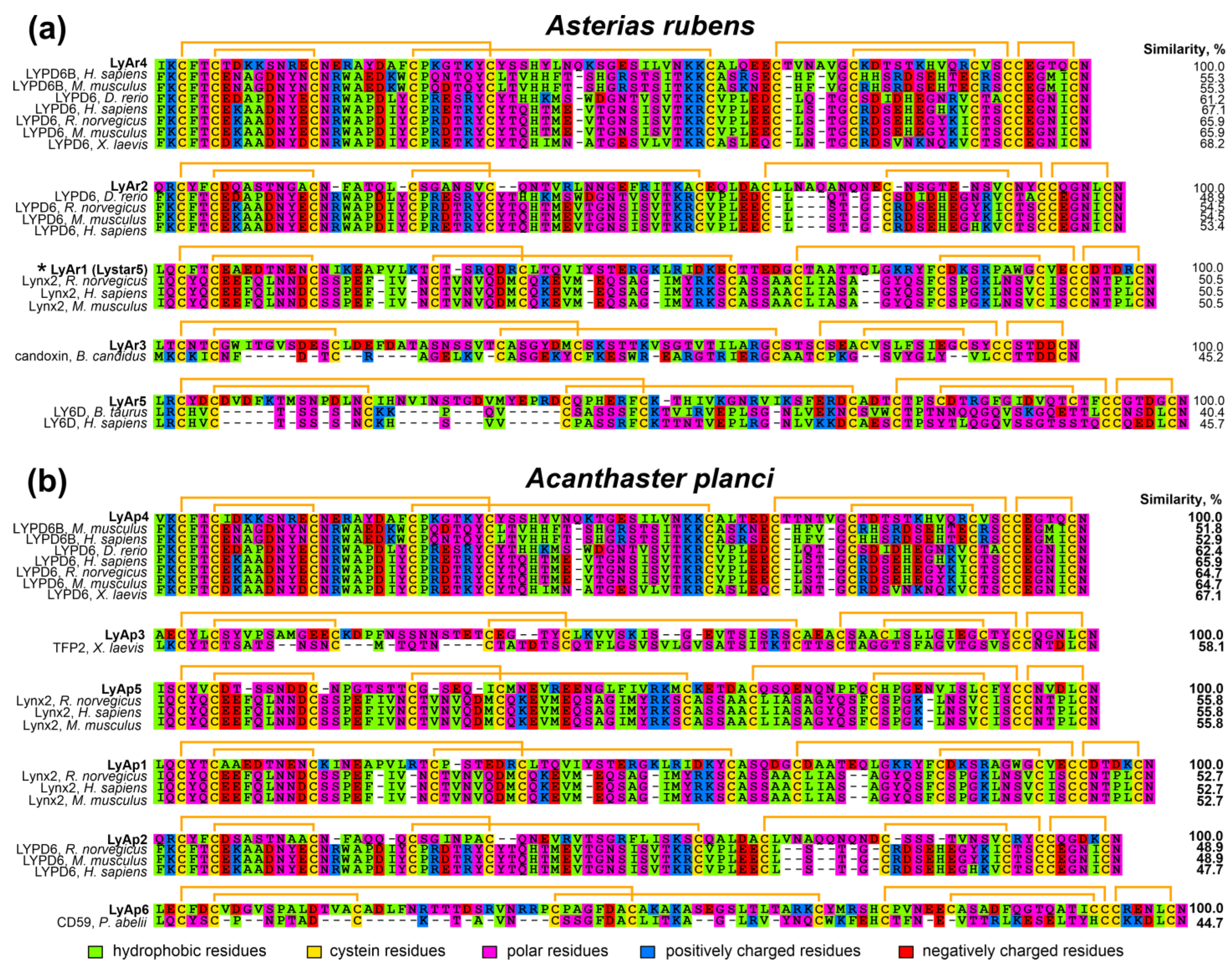
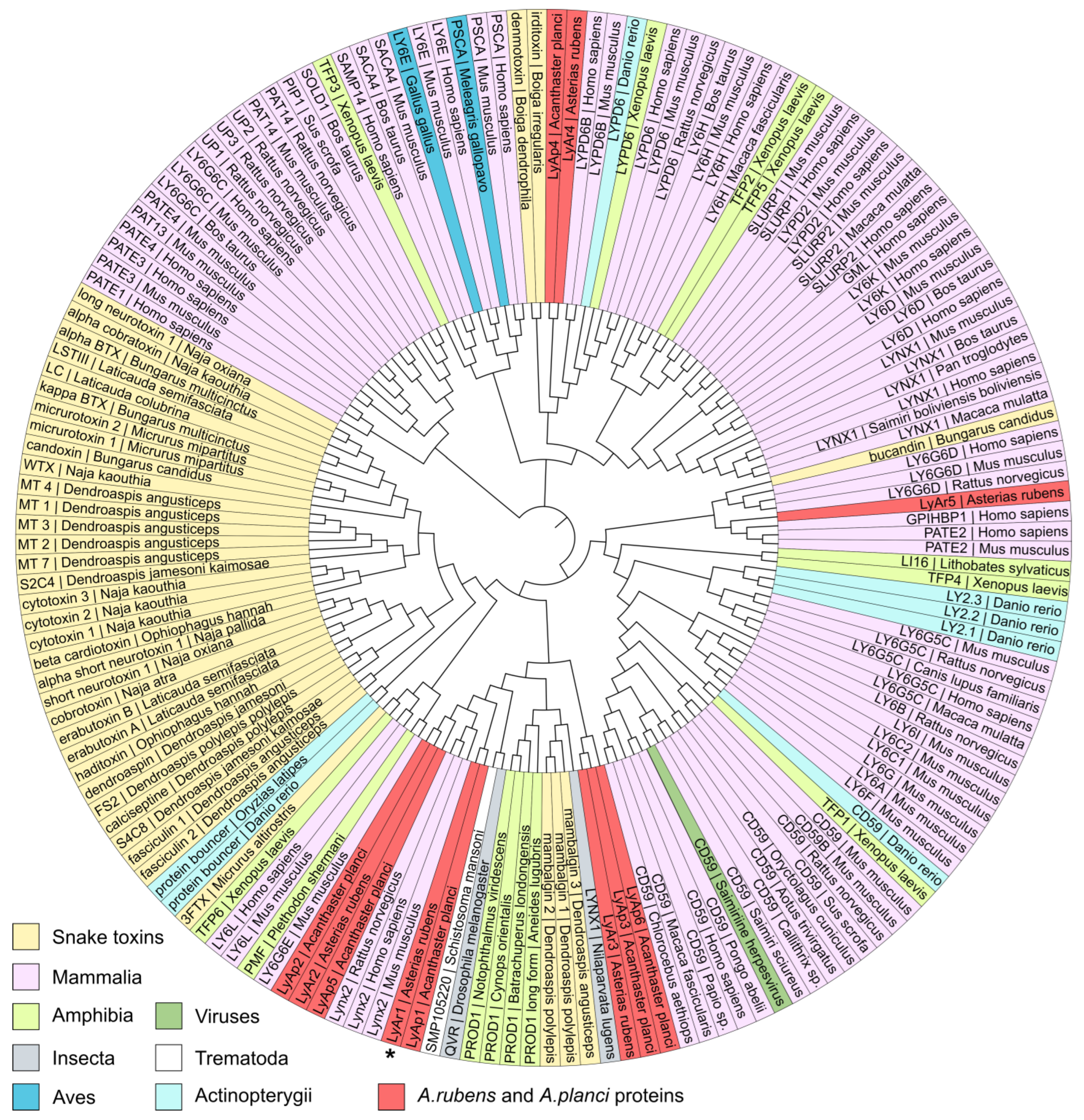
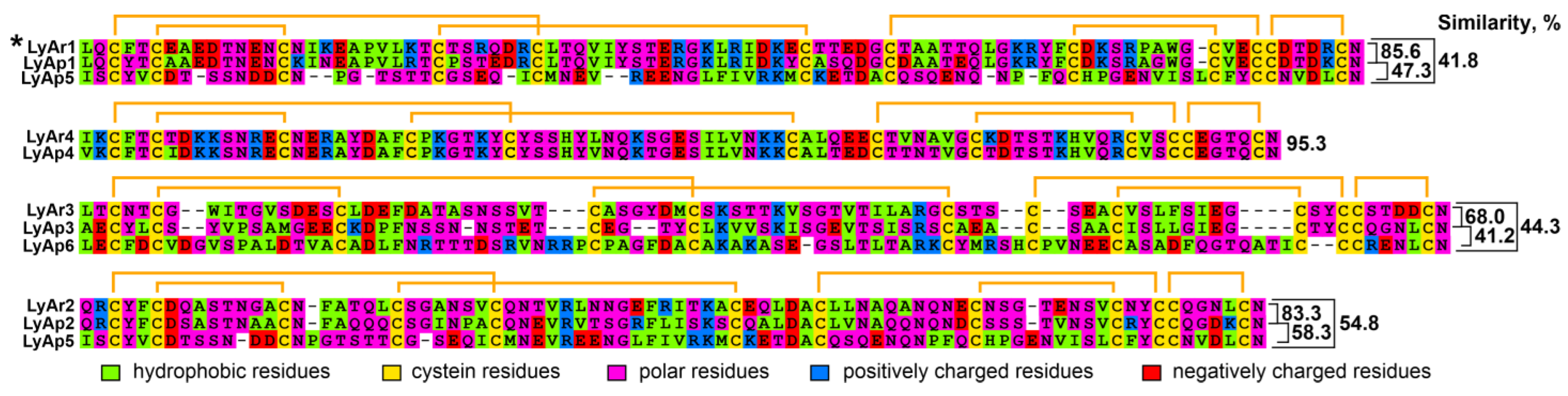
New Three-Finger Protein from Starfish Asteria rubens Shares Structure and Pharmacology with Human Brain Neuromodulator Lynx2



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
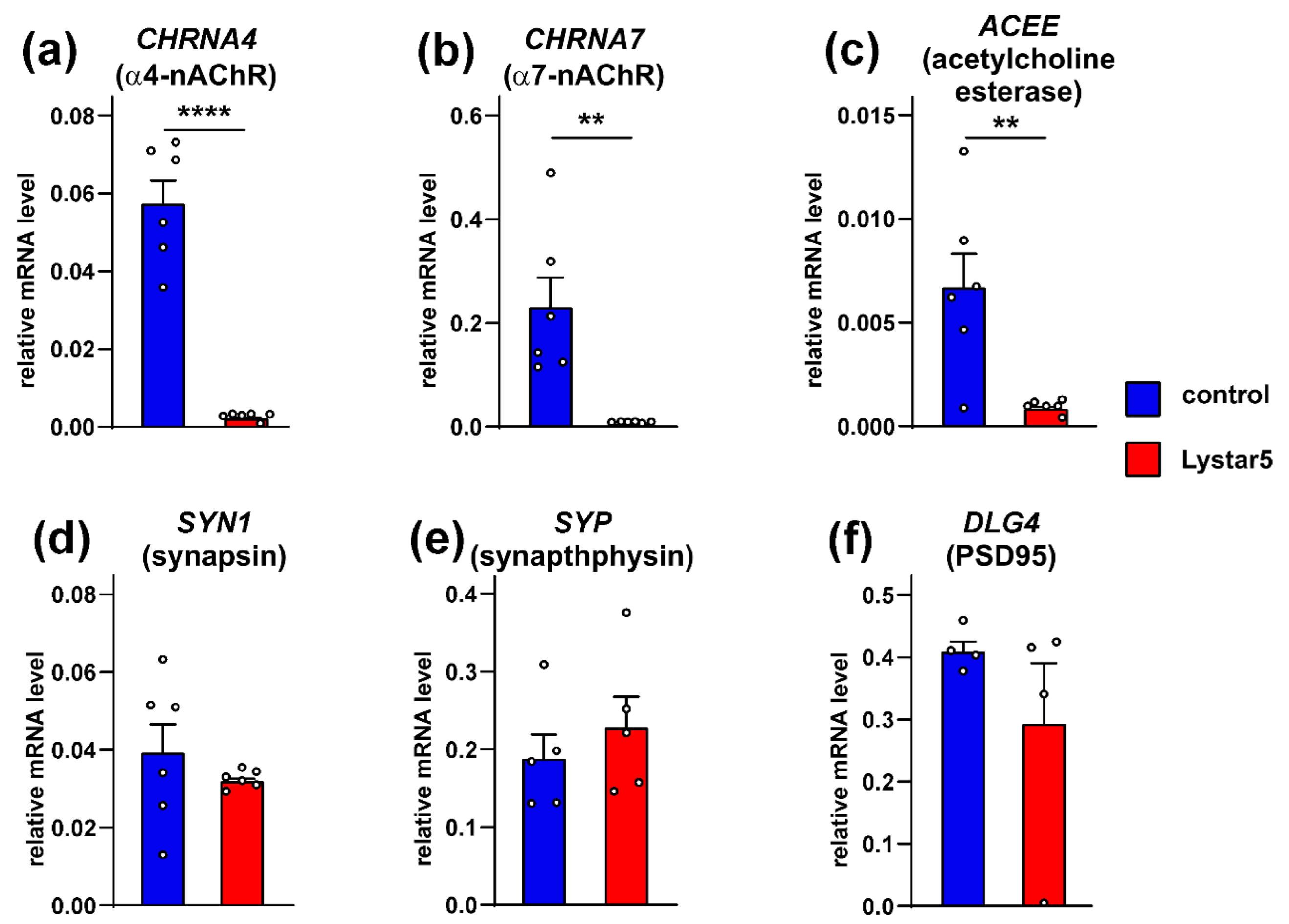
Abstract
Share and Cite
Paramonov, A.S.; Shulepko, M.A.; Makhonin, A.M.; Bychkov, M.L.; Kulbatskii, D.S.; Chernikov, A.M.; Myshkin, M.Y.; Shabelnikov, S.V.; Shenkarev, Z.O.; Kirpichnikov, M.P.; et al. New Three-Finger Protein from Starfish Asteria rubens Shares Structure and Pharmacology with Human Brain Neuromodulator Lynx2. Mar. Drugs 2022, 20, 503. https://doi.org/10.3390/md20080503
Paramonov AS, Shulepko MA, Makhonin AM, Bychkov ML, Kulbatskii DS, Chernikov AM, Myshkin MY, Shabelnikov SV, Shenkarev ZO, Kirpichnikov MP, et al. New Three-Finger Protein from Starfish Asteria rubens Shares Structure and Pharmacology with Human Brain Neuromodulator Lynx2. Marine Drugs. 2022; 20(8):503. https://doi.org/10.3390/md20080503
Chicago/Turabian StyleParamonov, Alexander S., Mikhail A. Shulepko, Alexey M. Makhonin, Maxim L. Bychkov, Dmitrii S. Kulbatskii, Andrey M. Chernikov, Mikhail Yu. Myshkin, Sergey V. Shabelnikov, Zakhar O. Shenkarev, Mikhail P. Kirpichnikov, and et al. 2022. "New Three-Finger Protein from Starfish Asteria rubens Shares Structure and Pharmacology with Human Brain Neuromodulator Lynx2" Marine Drugs 20, no. 8: 503. https://doi.org/10.3390/md20080503
APA StyleParamonov, A. S., Shulepko, M. A., Makhonin, A. M., Bychkov, M. L., Kulbatskii, D. S., Chernikov, A. M., Myshkin, M. Y., Shabelnikov, S. V., Shenkarev, Z. O., Kirpichnikov, M. P., & Lyukmanova, E. N. (2022). New Three-Finger Protein from Starfish Asteria rubens Shares Structure and Pharmacology with Human Brain Neuromodulator Lynx2. Marine Drugs, 20(8), 503. https://doi.org/10.3390/md20080503