1. Introduction
Chlorpropham (CIPC), isopropyl 3-chlorocarbanilate (IUPAC), is a carbamate herbicide and is very widely used worldwide as a general plant growth regulator to control sprouting and as a herbicide against target weeds [
1,
2]. Recently, Xu and Harvey [
3] showed that the addition of micromolar quantities of chlorpropham to cultures of the microalga,
Dunaliella salina, inhibited cell division and resulted in the massive overaccumulation of the colourless carotenoids, phytoene and phytofluene.
D. salina is a halotolerant marine microalga and well known for producing high quantities of β-carotene [
4]. Phytoene and phytofluene are precursors of β-carotene and are naturally found in a limited amount in the alga. Chlorpropham was proposed to disrupt synchronised control between nuclear and chloroplast events in cell division, which would normally be associated with carotenogenesis and β-carotene accumulation [
4].
Phytoene and phytofluene are similar to β-carotene in being comprised of a C40 backbone of isoprenoid units, which confer antioxidant and anti-inflammatory properties [
5]. Unlike β-carotene, however, the absorbance maxima for phytoene and phytoene lie within the UV wavelength range (280–350 nm). Phytoene and phytoene are consequently sought after as UV-protective ingredients in skin protection products [
6,
7]. Both compounds are found naturally in various stereoisomeric forms, which differ in their physicochemical properties and shape. In simulated gastro-intestinal studies,
cis-isomers of phytoene and phytofluene from different fruit juices presented with a higher bioaccessibility than their
trans-isomer counterparts, suggesting that they may have a higher therapeutical value [
8]. The isomers from
D. salina were recently comprehensively characterised and quantified after treatment of
D. salina cultures with chlorpropham [
7].
Cis- forms of the isomers are the predominant form in
D. salina. However, phytoene and phytofluene normally are only found in a limited amount in
D. salina. This has, until now, limited the potential for industrial-scale developments using this alga [
9].
To date, chlorpropham is known to inhibit the process of mitosis in plants and algae by interfering with the spindle microtubule organising centre, causing abnormal or complete suppression of microtubule synthesis and organisation. However, its specific site of action is not yet known [
10]. Sterling [
11] showed that lipophilic and neutral herbicides, including chlorpropham, could penetrate the cell membrane of some lower and several higher plants by passive diffusion; the kinetics studies of the uptake and efflux of different neutral pesticides have been cited. Little is known about the cellular uptake and accumulation of chlorpropham by microalgae and specifically
D. salina. Likewise, the biotransformation of chlorpropham has been investigated in higher plants and mainly in potatoes due to its extensive use on this crop. In studies performed with potatoes, chlorpropham was shown to be decarboxylated to 3-chloroaniline (3-CA) [
12], which is an organochlorine compound listed on the European Community priority pollutant Circular No 90–55 (1990) [
13], and classified as highly toxic for the environment and humans. Studies on the metabolism of chlorpropham in algae are limited. To our knowledge, only John et al. [
14] investigated its degradation by a selection of green algae (such as
Chlorella pyrenoidosa and
Chlamydomonas, Ulothrix fimbriata) and blue green algae (
Anacistis nidulans). Using colorimetric methods for detection of 3-CA, they observed that only
A. nidulans transformed chlorpropham to 3-CA.
The aim of the present work was to gain insights into the interaction between the herbicide chlorpropham and D. salina cells by studying its uptake and possible metabolism to 3-CA in cell cultures of D. salina. The objectives involved the detection of chlorpropham and its metabolites in the algal biomass, the study of its concentration over time and its removal from the biomass.
3. Discussion
Chlorpropham is a plant herbicide and a mitotic inhibitor which has been used extensively as a potato sprout suppressant. When
D. salina cultures were treated with a micromolar concentration of chlorpropham, the two colourless carotenoid precursors, phytoene and phytofluene, massively accumulated [
3]. The present work was undertaken to explore the interaction between chlorpropham and
D. salina cells and the accumulation and removal of chlorpropham in the algal biomass.
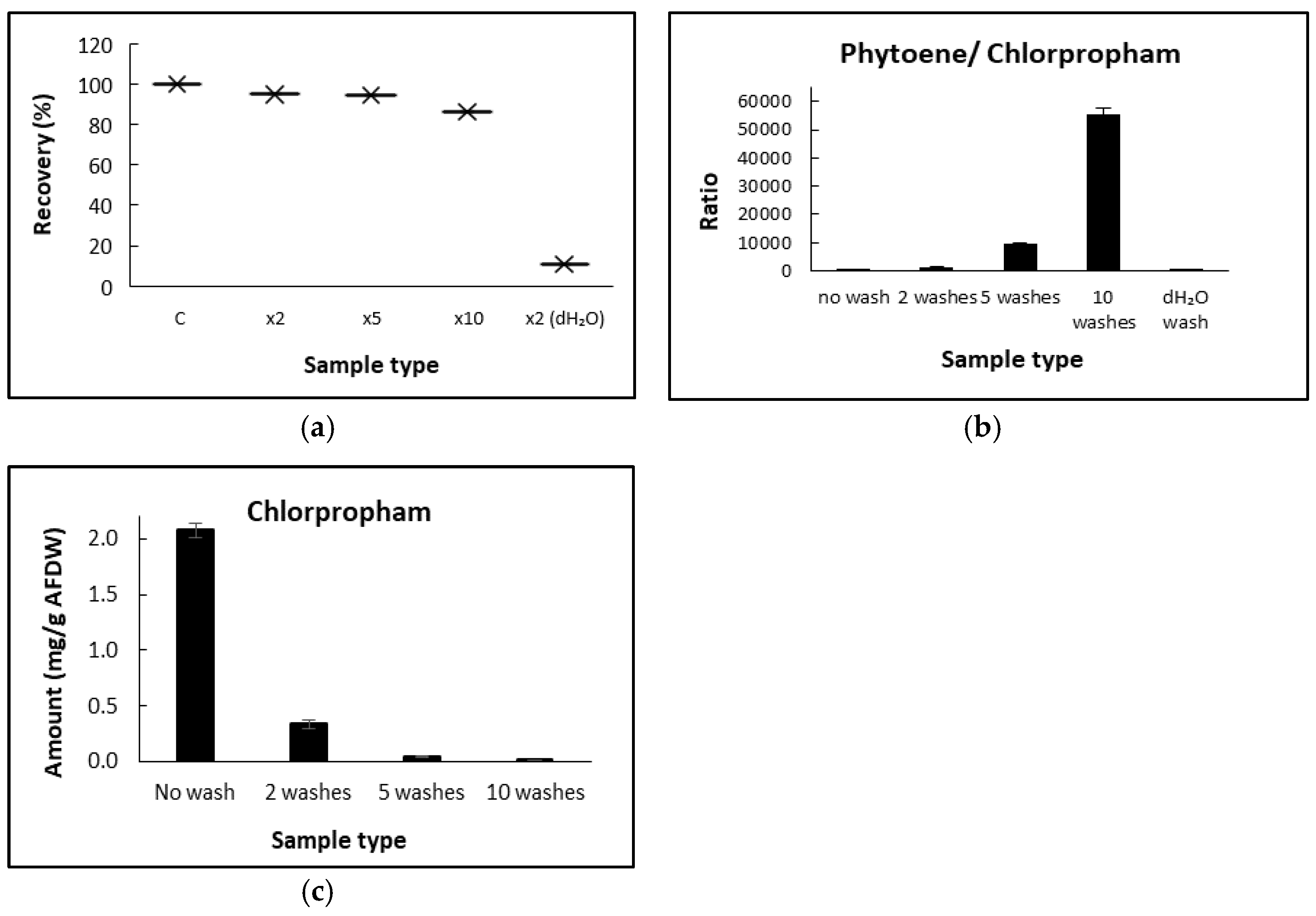
D. salina biomass concentrated chlorpropham from the culture medium, but most could be removed by washing the biomass. A similar phenomenon has been previously reported using higher plant tissues [
15]: in the study of the absorption of C-labelled herbicides washed for different cycles and time periods, herbicides such as fluorodifen (log K ow of 1.84 at 25 °C), which were concentrated by higher plant tissues, were removed after 10 min washing period. In the present study, the fresh culture medium was chosen as the washing solution to maintain the osmotic pressure and reduce the likelihood of cell rupture during washing since
D. salina cells have no rigid cell wall [
16]. Under these circumstances, the intracellular phytoene content remained within the cells.
Monitoring of the chlorpropham uptake by
D. salina over time showed that the association of chlorpropham with the harvested biomass occurred within the first few minutes of treatment. Lipophilic herbicides have been shown to freely transfer across the cell membrane of cells via passive diffusion until the chemical equilibrium between the internal and external concentration is reached [
11] and at a rate dependent on their lipophilicity. Chlorpropham is relatively lipophilic with a partition coefficient of octanol/water = 5.75 × 10
3.
D. salina cells, moreover, are bounded from the extracellular medium by a lipophilic membrane comprising a glycocalyx-like cell layer of varying thickness [
16]. Although the precise site of action of chlorpropham is not clear, it is likely to be internal, as chlorpropham is known to disrupt mitosis by interfering with the spindle microtubule organising centres in lower and higher plants species [
10,
17] or by interacting with the microtubules directly [
18]. In
D. salina cultures, chlorpropham may be taken up by cells by internal passive diffusion through the cell membrane; this needs further exploration. In the present study,
D. salina cells treated with chlorpropham for either 10 min or 24 h continued to accumulate phytoene even after their transfer into herbicide-free fresh medium and at the same rate as those cells that remained in chlorpropham-containing culture medium.
Chlorpropham accumulation during cell growth may parallel the increase in cell volume; increases in cell volume in cells treated with chlorpropham have been shown previously [
19]. Our data show that the content of phytoene significantly accumulated over the period tested (21 days), and cell volumes of all treated cultures were observed to increase over time after the treatment while phytoene and phytofluene were accumulated, in accord also with [
3]. Additionally, the sharp increase in volume in cultures of low cell number paralleled higher carotenoid accumulation, suggesting a correlation between
D. salina cell number and the amount of chlorpropham added. Therefore, the physiological changes observed on day 21 of treatment in cultures with lower cell density may be linked to a higher toxicity effect of intracellular herbicide. It was shown previously that cell swelling occurred in cells grown once a specific level of chlorpropham was reached [
20]. On the other hand, when cultures were treated for only 10 min or 24 h and resuspended in fresh medium, the effect of chlorpropham on the overaccumulation of phytoene was shown even after 24 days, proving the irreversibility of the herbicide after short exposure without showing cell toxicity caused by prolonged exposures to the herbicide.
The formation of the metabolite, 3-chloroaniline, has been a concern for chlorpropham applications. In this study, no 3-chloroaniline was detected in cultures treated with up to 1 mM herbicide with the method used, suggesting that
D. salina might not be capable of breaking down chlorpropham. There is little information available regarding the degradation of chlorpropham in microalgal species. John et al. [
14] investigated the presence of 3-chloroaniline in different microalgae and a cyanobacterium and showed that
Ulothrix fimbriata, which belongs to the same phylum
Chlorophyta as
D. salina, did not produce 3-chloroaniline, in contrast to the blue-green alga
Anacystis nidulans, which possessed the enzyme acylamidase. On the other hand, studies on potato tissues attributed the presence of 3-chloroaniline after chlorpropham treatment to the thermal degradation of chlorpropham during the fogging application [
21], when extremely high temperatures (>300 °C) are used; high temperatures have been reported to trigger the degradation to 3-chloroaniline [
22]. Additionally, 3-chloroanline may form during potato storage due to the activity of microorganisms capable of degrading the parent compound to this metabolite [
23].
The present work aimed to elucidate details of the use of chlorpropham in D. salina cultivation for phytoene production. Chlorpropham added to cultures was concentrated in the biomass. The effects caused by chlorpropham on cell metabolism are irreversible. The constant increase in the chlorpropham content associated with the biomass over time could be linked to increased cell volume during the accumulation of the colourless carotenoids, phytoene and phytofluene. Washing repeatedly with fresh medium will remove most chlorpropham associated with harvested biomass, up to more than 99% of its initial amount. Phytoene and phytofluene are high valued compounds which are sought to have high beneficial health properties; however, their availability in nature is low. The use of chlorpropham with D. salina cultivation may represent a facile, low-cost method for producing large quantities of phytoene.
4. Materials and Methods
4.1. Alga Strain and Cultivation
D. salina strain DF15 (CCAP 19/41) was obtained from the Marine Biological Association, UK (MBA). Cultures were cultivated in 500 mL Modified Johnsons Medium [
24] containing 1.5 M NaCl and 10 mM NaHCO
3 in an illuminated incubator (Varicon Aqua, Worcester, UK) under white light of 500 µmol m
−2 s
−1 at 25 ± 2 °C. For the different kinetic studies of treatments with herbicides, triplicate sets of cultures were grown to mid-late log phase, and each set was treated with chlorpropham at different concentrations from 20 µM up to 1 mM. Flasks containing only fresh culture media added with the same amount of herbicide served as blank controls to monitor the natural degradation rate of chlorpropham over time without algal cells.
4.2. Standards and Solvents
Phytoene standard (LGC Limited, Teddington, UK), chlorpropham (PESTANAL, analytical standard) and 3-chloroaniline (99% purity) were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Methanol (MeOH) and Methyl Tert Butyl Ether (MTBE), both HPLC grade, were purchased from Fischer Scientific UK Ltd. (Loughborough, Leicestershire, UK).
4.3. Extraction and Analysis of Phytoene
To determine the yield of phytoene and other carotenoids in chlorpropham-treated and control cultures of
D. salina at different time periods, 5 to 10 mL of
D. salina cultures were harvested at 3000×
g for 5 min at 20 °C with an Eppendorf centrifuge 5810R (Eppendorf, UK). Carotenoids were extracted and analysed by HPLC as described in our previous work [
7]: 10 mL of MeOH: MTBE (80:20) was added to the samples, which were first sonicated for 60 s and then vortexed for 20 s. Samples were clarified at the centrifuge, and the top solvent phase was collected. The extracts were filtered (0.20 µm filter) into amber HPLC vials before analysis. The carotenoid extracts were routinely flushed with nitrogen after extraction. To determine the kinetics for phytoene accumulation,
D. salina cultures (cell density = 85.5 × 10
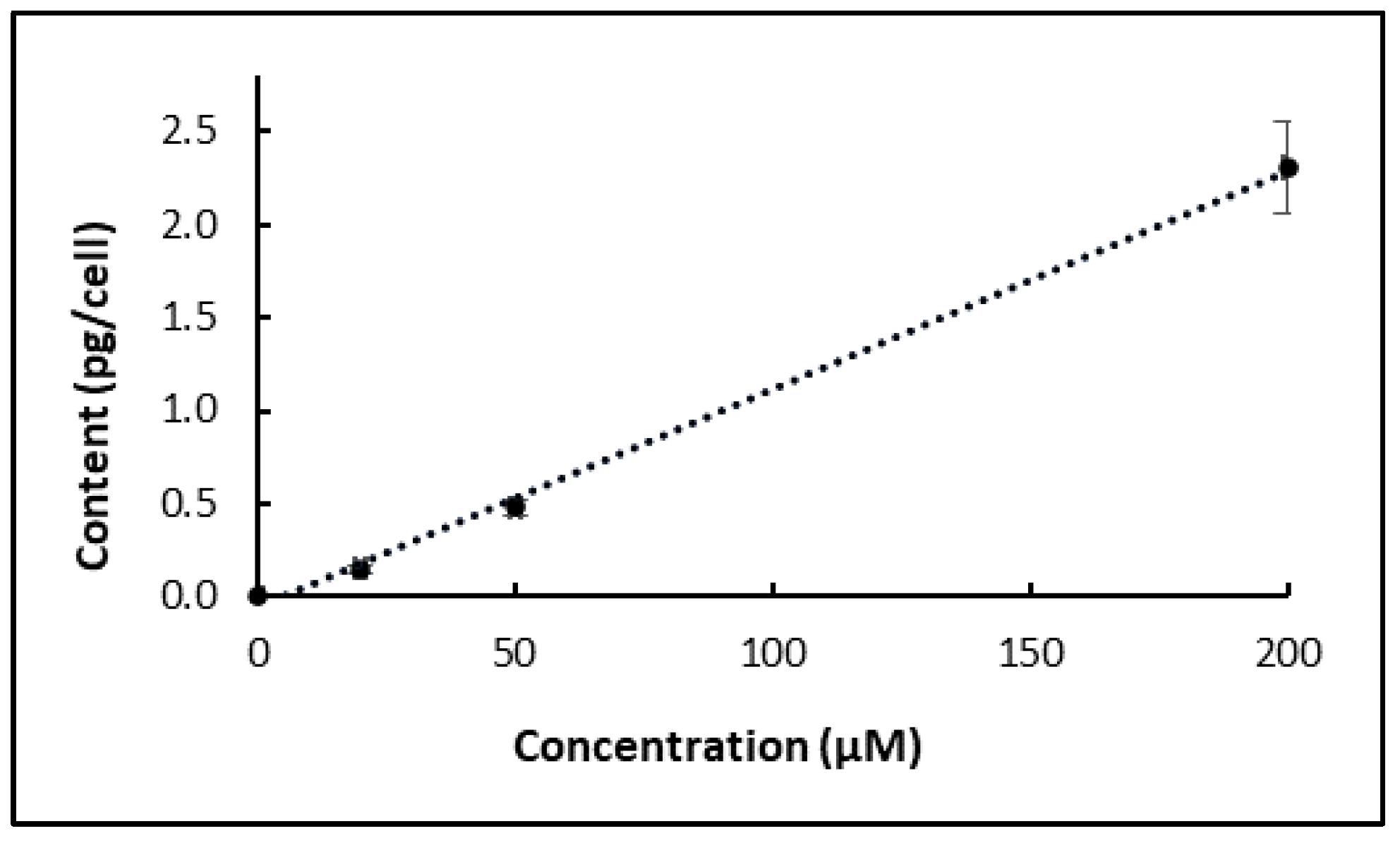
4/mL) were treated with chlorpropham for either 10 min or for 24 h, and then aliquots of the cultures (30 mL) were transferred to fresh, herbicide-free, culture medium (60 mL), and the rate of phytoene accumulation (pg/cell) determined for 24 days. The phytoene yield in those cultures was compared to cultures not treated and cultures treated for all the 24 days analysis period. To determine the yield of phytoene, a calibration curve was generated for its quantification.
4.4. Extraction and Analysis of Chlorpropham and Its Potential Metabolites
4.4.1. Sample Preparation
Analytical methods for chlorpropham and its metabolite 3-chloroaniline include high-performance liquid chromatography or gas chromatography. However, gas chromatography often requires the derivatisation of the compounds before analysis and, therefore, HPLC was selected as the main tool for chlorpropham and 3-CA analysis in this study. To monitor the amount of chlorpropham in D. salina cells on different days of treatment (day 0 to day 28), 10 to 15 mL cultures were harvested at 3000× g for 5 min at 20 °C. The extraction of chlorpropham from D. salina biomass was evaluated with the following solvent extracts: MeOH (100%), MTBE (100%), Ethanol (100%), MTBE/MeOH (20/80). 2.5 mL MeOH (100%) was added to the biomass, and the solvent suspension was sonicated for 20 s, vortexed for 20 s and centrifuged at 3,000× g for 5 min. The upper phase with the solvent was collected, filtered with 0.2 µm size filters into amber HPLC vials and analysed by HPLC.
To enhance the compatibility with the HPLC mobile phase, as a stronger sample phase solvent than the mobile phase may lead to inaccuracies in the results, a final sample phase consisting of 90% sample (in MeOH) and 10% of HPLC grade water was prepared. To analyse samples deriving from the algal supernatant, 5 mL of each collected supernatant was diluted with 15 mL of MeOH (HPLC grade) to reach a ratio (25:75, H2O/MeOH).
4.4.2. HPLC Analysis of Chlorpropham and 3-Chloroaniline
The presence of chlorpropham and its metabolite, 3-chloroaniline, was evaluated with HPLC by matching the retention time and the UV-vis spectra features of authentic standards; these values are shown in
Table 3. The instrument was equipped with Diode-Array Detection (HPLC-DAD; Agilent Technologies 1200 series, Agilent, Santa Clara, CA, USA), an online degasser and a quaternary pump system. Samples were separated using a C18 250 × 4.6 mm Waters Spherisorb 5 µm ODS2. MeOH/ dH
2O mixtures with different ratios of water (from 0 to 10%) were tested as mobile phase solvents for analysis. The method parameters chosen were as follows: the column temperature was set at 20 °C, and the gradient solvent system was MeOH/ dH
2O (90/10) (A) running at 100% A for the first 8 min at a flow rate of 0.6 mL/min, then running at 100% MeOH (B) for one minute, before adjusting to 80% B and 20% C (C = 100% MTBE) for the next 20 min at a flow rate of 1.0 mL/min (to elute the carotenoids) before going back to the initial conditions at 30 min. The total run time was 45 min. The absorbance at different wavelengths (210, 240, 250, 282, 355, 450, 480 nm) was monitored. All data were acquired and analysed with Chemstation for LC System software. The method was validated for precision linearity, selectivity, limit of detection and quantification as in [
25]. Calibration curves were obtained for chlorpropham and chloroaniline quantification by spiking known concentrations of chlorpropham and chloroaniline standards into crude algal extracts and supernatant and preparing serial dilution (at least 7 points) within the range needed for the experiment. At least three fresh solutions for each concentration were used, and the mean and standard deviation was obtained. Each area value was plotted against each corresponding concentration to obtain the best-fitted line. To detect the presence of 3-chloroaniline in the biomass of
D. salina, cultures were treated with an increasing concentration of chlorpropham (up to 1 mM) and aliquots of cultures (from 10 to 100 mL) were harvested and extracted. Then, the solvent was evaporated with the vacuum evaporator Genevac and resuspended to the compatible mobile phase.
4.5. Confocal Microscopy Analysis
The cell volume of D. salina in treated cultures was measured with an LSM880 Bruker confocal microscope. Two triplicate sets of cultures with different cell densities were set up, with the culture of lower density diluted from the one of higher density. Both cultures were treated with 20 µM chlorpropham. The laser channel was set at 488 nm, and 2D images were acquired as follows: the speed was set at 5, and the averaging number was 4. Images size was: 425.1 µM × 425.1 µM, and the pixel size was 0.42 µm. D. salina cells were fixed with formalin (2%) before analysis. Different images for each culture flask were acquired to have at least 50 to 100 cells per replicate flask to measure, and the volume of the cells was calculated by measuring their diameter and assuming that D. salina has a spherical shape.
4.6. Chlorpropham Removal by Washing Process
To eliminate the traces of chlorpropham surrounding the biomass of D. salina, samples were washed with water or the culture medium. Furthermore, the amounts of chlorpropham derived from the biomass washed with different washing cycles (0, 2, 5 and 10) or different washing solutions/harvest ratios were determined. Samples of each washing experiment were collected in triplicate from the same culture flask. The washing experiments conducted with different washing cycles were performed as follows: 10 mL culture was harvested via centrifugation at 3000× g for 5 min, the supernatant discharged, and the biomass pellet was resuspended completely in 10 mL washing solution (either culture medium or dH2O) by vortexing for 5 s; the suspension was centrifuged at 3,000× g for 5 min. For the experiment conducted with different harvest cultures/ washing solution volumes, the ratios were as follows: (1) 10 mL culture/10 mL washing solution; (2) 10 mL culture/20 mL washing solution; (3) 10 mL culture/40 mL washing solution.
4.7. Statistical Analysis
Data are presented as the mean and standard deviation of triplicates. IBM SPSS statistics 64-bit was used to perform ANOVA analysis with a significant level of p < 0.05 to compare the significance between data at different treatment times and between control and treatment cultures. Microsoft Excel for Office MSO 64-bit was used for graphs representations.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}