
Valorization of Side Stream Products from Sea Cage Fattened Bluefin Tuna (Thunnus thynnus): Production and In Vitro Bioactivity Evaluation of Enriched ω-3 Polyunsaturated Fatty Acids

 , , , , , and
, , , , , and
Abstract
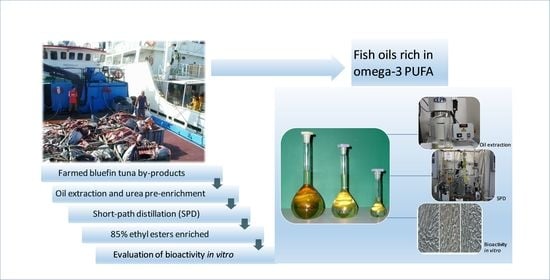
1. Introduction
2. Results and Discussion
2.1. Recovery of Processing Side Stream from Sea Cage Fattened Bluefin Tuna
2.2. Extraction, Yield and Quality of Tuna Oil (TO)
2.3. Pre-Enrichment by Urea Complexation
2.4. PUFA Enrichment by SPD
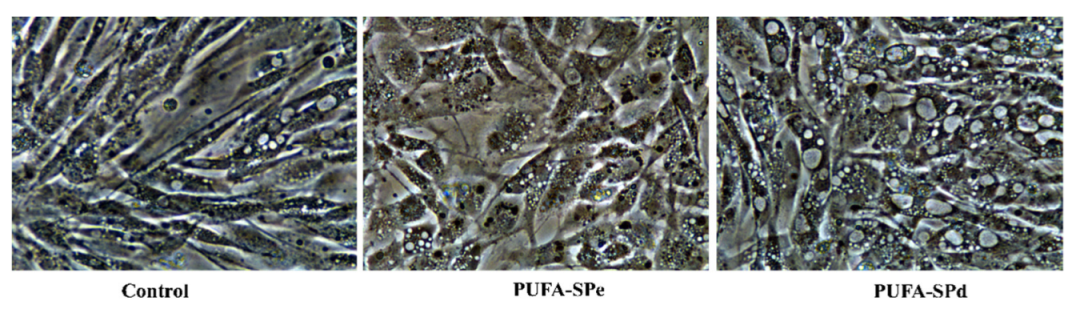
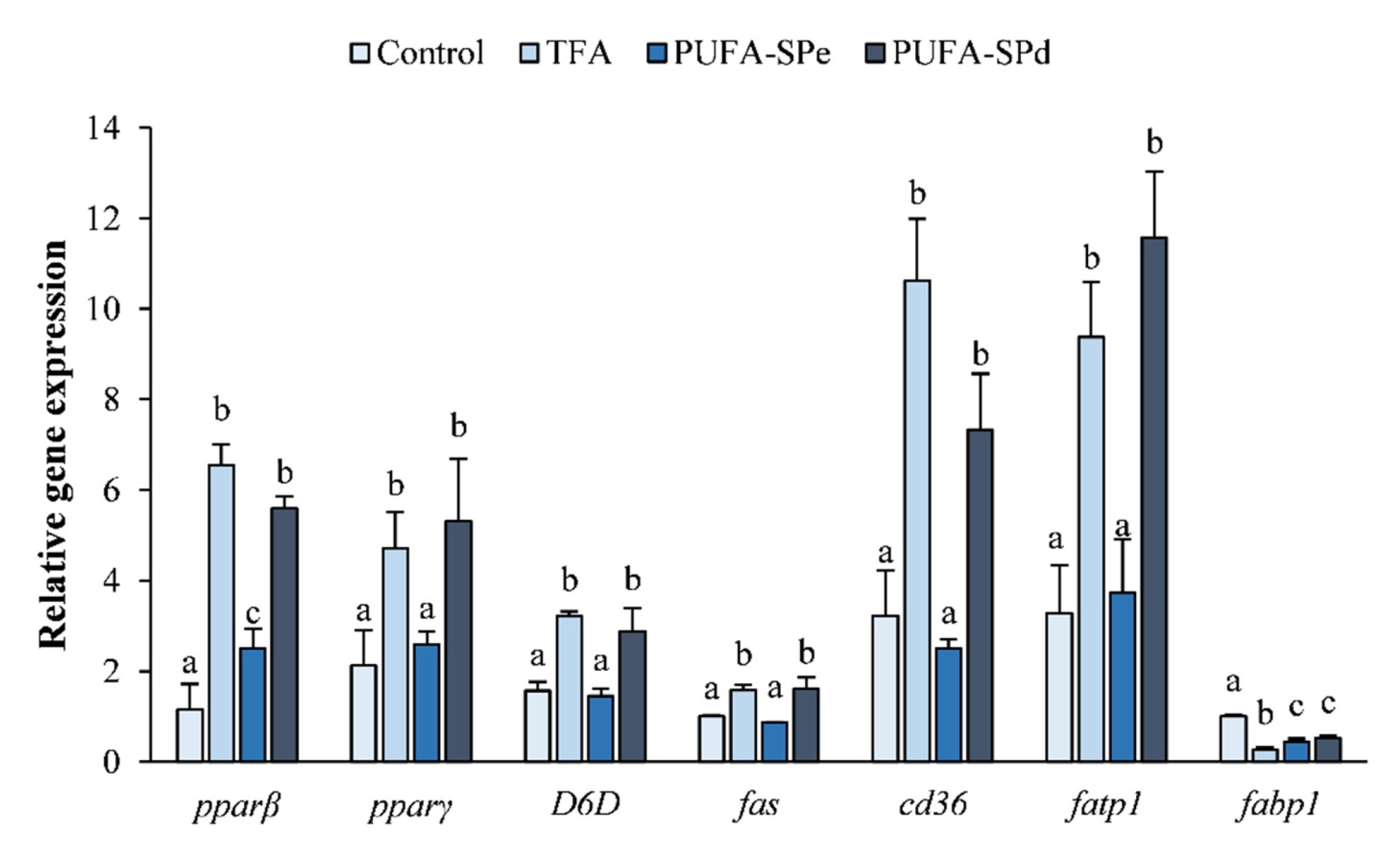
2.5. Effects of Fatty Acids Ethyl Esters on Lipid Accumulation and Related Genes, in SAF-1 Cell Lines
3. Materials and Methods
3.1. Sampling
3.2. Proximate Composition and FA Profile
- the polyene index (PI) (Equation (1)), used as a measure of PUFA damage [100]:where C20:5ω-3 represents EPA, C22:6ω-3 DHA and C16:0 palmitic acid.PI = ((C20:5ω-3 + C22:6ω-3)/C16:0)
- the atherogenicity index (AI) (Equation (2)), is a nutritional quality index for assessing the risk of platelet aggregation [101]:where C12:0 represents lauric acid, C14:0 miristyc acid, C16:0 palmitic acid, MUFA monounsaturated fatty acid and PUFA polyunsaturated fatty acid.AI = (C12:0 + 4 × C14:0 + C16:0)/(MUFA + PUFA)
- the thrombogenicity index (TI) (Equation (3)) [101], is a nutritional quality index representing the potential to form clots in the blood vesselswhere C14:0 represents miristyc acid, C16:0 palmitic acid, C18:0 stearic acid, MUFA is monounsaturated fatty acid, Totω-6 the total amount of omega 6 fatty acid and Totω-3 the total amount of omega 3 fatty acids.TI = [(C14:0 + C16:0 + C18:0)/(0.5 × MUFA + 0.5 × Totω-6 + 3 × Totω-3 + (Totω-3/Totω-6))]
3.3. Extraction of Crude Oil
3.4. Assessment of Oil Quality
3.5. Urea PUFA Enrichment
3.6. Shorth Path Distillation PUFA Enrichment
- EPA and DHA%.
- FA ratio (R) (Equation (5)) [69]:where C20:5ω-3 represents EPA, C22:6ω-3 DHA, 16:0 palmitic acid and 18:1ω9 oleic acid.R = (C20:5ω-3 + C22:6ω-3)/(16:0 + 18:1ω9)
- Enrichment factor for EPA, DHA, and PUFA.
- Ratio of total PUFA to total SFA (PUFA/SFA).
3.7. SAF-1 Cell Culture
3.8. Image Acquisition
3.9. RNA isolation and Rt-qPCR
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020; ISBN 9789251326923. [Google Scholar]
- de la Fuente, B.; Pallarés, N.; Barba, F.J.; Berrada, H. An integrated approach for the valorization of sea bass (Dicentrarchus labrax) side streams: Evaluation of contaminants and development of antioxidant protein extracts by pressurized liquid extraction. Foods 2021, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Thirukumaran, R.; Anu Priya, V.K.; Krishnamoorthy, S.; Ramakrishnan, P.; Moses, J.A.; Anandharamakrishnan, C. Resource recovery from fish waste: Prospects and the usage of intensified extraction technologies. Chemosphere 2022, 299, 134361. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef] [PubMed]
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C.; De Pascale, D.; Santulli, A. Fish Waste: From Problem to Valuable Resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef]
- Messina, C.M.; Arena, R.; Manuguerra, S.; Renda, G.; Laudicella, V.A.; Ficano, G.; Fazio, G.; La Barbera, L.; Santulli, A. Farmed Gilthead Sea Bream (Sparus aurata) by-Products Valorization: Viscera Oil ω-3 Enrichment by Short-Path Distillation and In Vitro Bioactivity Evaluation. Mar. Drugs 2021, 19, 160. [Google Scholar] [CrossRef]
- Messina, C.M.; Manuguerra, S.; Arena, R.; Renda, G.; Ficano, G.; Randazzo, M.; Fricano, S.; Sadok, S.; Santulli, A. In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolisates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “From Waste to Profit”. Mar. Drugs 2021, 19, 216. [Google Scholar] [CrossRef]
- Kim, S.K.; Mendis, E. Bioactive compounds from marine processing byproducts—A review. Food Res. Int. 2006, 39, 383–393. [Google Scholar] [CrossRef]
- Rustad, T.; Storrø, I.; Slizyte, R. Possibilities for the utilisation of marine by-products. Int. J. Food Sci. Technol. 2011, 46, 2001–2014. [Google Scholar] [CrossRef]
- Galanakis, C.M. Recovery of high added-value components from food wastes: Conventional, emerging technologies and commercialized applications. Trends Food Sci. Technol. 2012, 26, 68–87. [Google Scholar] [CrossRef]
- Pateiro, M.; Munekata, P.E.S.; Wang, M.; Barba, F.J.; Berm, R.; Lorenzo, J.M. Nutritional Profiling and the Value of Processing By-Products from Gilthead Sea Bream (Sparus aurata). Mar. Drugs 2020, 18, 101. [Google Scholar] [CrossRef]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F.J. Functional and Bioactive Properties of Peptides Derived from Marine Side Streams. Mar. Drugs 2021, 19, 71. [Google Scholar] [CrossRef]
- García-de-Vinuesa, A.; Demestre, M.; Carreño, A.; Lloret, J. The Bioactive Potential of Trawl Discard: Case Study from A Crinoid Bed off Blanes (North-Western Mediterranean). Mar. Drugs 2021, 19, 83. [Google Scholar] [CrossRef]
- Messina, C.; Renda, G.; La Barbera, L.; Santulli, A. By-products of farmed European sea bass (Dicentrarchus labrax L.) as a potential source of n-3 PUFA. Biologia 2013, 68, 288–293. [Google Scholar] [CrossRef][Green Version]
- Gnanavel, V.; Roopan, S.M.; Rajeshkumar, S. Aquaculture: An overview of chemical ecology of seaweeds (food species) in natural products. Aquaculture 2019, 507, 1–6. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Gustafson, K.R. Marine pharmacology in 2003–2004: Anti-tumour and cytotoxic compounds. Eur. J. Cancer 2006, 42, 2241–2270. [Google Scholar] [CrossRef]
- Aspevik, T.; Thoresen, L.; Steinsholm, S.; Carlehög, M.; Kousoulaki, K. Sensory and Chemical Properties of Protein Hydrolysates Based on Mackerel (Scomber scombrus) and Salmon (Salmo salar) Side Stream Materials. J. Aquat. Food Prod. Technol. 2021, 30, 176–187. [Google Scholar] [CrossRef]
- Lapi, I.; Kolliniati, O.; Aspevik, T.; Deiktakis, E.E.; Axarlis, K.; Daskalaki, M.G.; Dermitzaki, E.; Tzardi, M.; Kampranis, S.C.; El Marsni, Z.; et al. Collagen-containing fish sidestream-derived protein hydrolysates support skin repair via chemokine induction. Mar. Drugs 2021, 19, 396. [Google Scholar] [CrossRef]
- Alfio, V.G.; Manzo, C.; Micillo, R. From Fish Waste to Value: An Overview of the Sustainable Recovery of Omega-3 for Food Supplements. Molecules 2021, 26, 1002. [Google Scholar] [CrossRef]
- Regueiro, L.; Newton, R.; Soula, M.; Méndez, D.; Kok, B.; Little, D.C.; Pastres, R.; Johansen, J.; Ferreira, M. Opportunities and limitations for the introduction of circular economy principles in EU aquaculture based on the regulatory framework. J. Ind. Ecol. 2021, 1–12. [Google Scholar] [CrossRef]
- Stübler, A.S.; Heinz, V.; Aganovic, K. Development of food products. Curr. Opin. Green Sustain. Chem. 2020, 25, 100356. [Google Scholar] [CrossRef]
- Šimat, V.; Vlahovic, J.; Soldo, B.; Skroza, D.; Ljubenkov, I.; Mekinic, I.G. Production and refinement of omega-3 rich oils from processing by-products of farmed fish species. Foods 2019, 8, 125. [Google Scholar] [CrossRef]
- Shepherd, C.J.; Jackson, A.J. Global fishmeal and fish-oil supply: Inputs, outputs and markets. J. Fish Biol. 2013, 83, 1046–1066. [Google Scholar] [CrossRef]
- Ferraro, V.; Cruz, I.B.; Jorge, R.F.; Malcata, F.X.; Pintado, M.E.; Castro, P.M.L. Valorisation of natural extracts from marine source focused on marine by-products: A review. Food Res. Int. 2010, 43, 2221–2233. [Google Scholar] [CrossRef]
- Zárate, R.; el Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef]
- Mentoor, I.; Engelbrecht, A.M.; Nell, T. Fatty acids: Adiposity and breast cancer chemotherapy, a bad synergy? Prostaglandins Leukot. Essent. Fat. Acids 2019, 140, 18–33. [Google Scholar] [CrossRef]
- Omabe, M.; Ezeani, M.; Omabe, K.N. Lipid metabolism and cancer progression: The missing target in metastatic cancer treatment. J. Appl. Biomed. 2015, 13, 47–59. [Google Scholar] [CrossRef]
- Schley, P.D.; Field, C.J.; Brindley, D.N. Omega-3 polyunsaturated fatty acids alter raft lipid composition and decrease epidermal growth factor receptor levels in lipid rafts of human breast cancer cells. FASEB J. 2007, 21, 3–8. [Google Scholar]
- Siddiqui, R.A.; Harvey, K.; Stillwell, W. Anticancer properties of oxidation products of docosahexaenoic acid. Chem. Phys. Lipids 2008, 153, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Tarasiuk, A.; Mosińska, P.; Fichna, J. The mechanisms linking obesity to colon cancer: An overview. Obes. Res. Clin. Pract. 2018, 12, 251–259. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 fatty acids and cardiovascular disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [PubMed]
- Rimm, E.B.; Appel, L.J.; Chiuve, S.E.; Djoussé, L.; Engler, M.B.; Kris-Etherton, P.M.; Mozaffarian, D.; Siscovick, D.S.; Lichtenstein, A.H. Seafood Long-Chain n-3 Polyunsaturated Fatty Acids and Cardiovascular Disease: A Science Advisory from the American Heart Association. Circulation 2018, 138, e35–e47. [Google Scholar] [CrossRef]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C.; Vázquez, J.A.; Meduíña, A.; Durán, A.I.; Nogueira, M.; Fernández-Compás, A.; Pérez-Martín, R.I.; Rodríguez-Amado, I.; et al. Impact of omega-3 polyunsaturated fatty acids on vascular function and blood pressure: Relevance for cardiovascular outcomes. Circulation 2019, 138, 191–200. [Google Scholar]
- Ofori-Mensah, S.; Yıldız, M.; Arslan, M.; Eldem, V. Fish oil replacement with different vegetable oils in gilthead seabream, Sparus aurata diets: Effects on fatty acid metabolism based on whole-body fatty acid balance method and genes expression. Aquaculture 2020, 529, 735609. [Google Scholar] [CrossRef]
- Antraco, V.J.; Hirata, B.K.S.; de Jesus Simão, J.; Cruz, M.M.; da Silva, V.S.; da Cunha de Sá, R.D.C.; Abdala, F.M.; Armelin-Correa, L.M.; Alonso-Vale, M.I.C. Omega-3 Polyunsaturated Fatty Acids Prevent Nonalcoholic Steatohepatitis (NASH) and Stimulate Adipogenesis. Nutrients 2021, 13, 622. [Google Scholar] [CrossRef]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 Long-Chain Polyunsaturated Fatty Acids, EPA and DHA: Bridging the Gap between Supply and Demand. Nutrients 2019, 11, 89. [Google Scholar]
- Salmerón, C. Adipogenesis in fish. J. Exp. Biol. 2018, 221, jeb161588. [Google Scholar] [CrossRef]
- Todorčević, M.; Vegusdal, A.; Gjøen, T.; Sundvold, H.; Torstensen, B.E.; Kjær, M.A.; Ruyter, B. Changes in fatty acids metabolism during differentiation of Atlantic salmon preadipocytes; Effects of n-3 and n-9 fatty acids. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2008, 1781, 326–335. [Google Scholar] [CrossRef]
- Murali, G.; Desouza, C.V.; Clevenger, M.E.; Ramalingam, R.; Saraswathi, V. Differential effects of eicosapentaenoic acid and docosahexaenoic acid in promoting the differentiation of 3T3-L1 preadipocytes. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 13–21. [Google Scholar] [CrossRef]
- Martins, F.F.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Eicosapentaenoic and docosapentaenoic acids lessen the expression of PPARγ/Cidec affecting adipogenesis in cultured 3T3-L1 adipocytes. Acta Histochem. 2020, 122, 151504. [Google Scholar] [CrossRef]
- Wayan Suriani, N.; Komansilan, A. Enrichment of omega-3 fatty acids, waste oil by-products canning tuna (Thunnus sp.) with urea crystallization. J. Phys. Conf. Ser. 2019, 1317, 012056. [Google Scholar] [CrossRef]
- Tang, S.; Qin, C.; Wang, H.; Li, S.; Tian, S. Study on supercritical extraction of lipids and enrichment of DHA from oil-rich microalgae. J. Supercrit. Fluids 2011, 57, 44–49. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Chen, G.; Zhang, J.; Wang, C.; Liu, B. Extraction and purification of eicosapentaenoic acid and docosahexaenoic acid from microalgae: A critical review. Algal Res. 2019, 43, 101619. [Google Scholar] [CrossRef]
- Kahveci, D.; Xu, X. Repeated hydrolysis process is effective for enrichment of omega 3 polyunsaturated fatty acids in salmon oil by Candida rugosa lipase. Food Chem. 2011, 129, 1552–1558. [Google Scholar]
- Chakraborty, K.; Vijayagopal, P.; Chakraborty, R.D.; Vijayan, K.K. Preparation of eicosapentaenoic acid concentrates from sardine oil by Bacillus circulans lipase. Food Chem. 2010, 120, 433–442. [Google Scholar] [CrossRef][Green Version]
- Gámez-Meza, N.; Noriega-Rodríguez, J.A.; Medina-Juárez, L.A.; Ortega-García, J.; Monroy-Rivera, J.; Toro-Vázquez, F.J.; García, H.S.; Angulo-Guerrero, O. Concentration of eicosapentaenoic acid and docosahexaenoic acid from fish oil by hydrolysis and urea complexation. Food Res. Int. 2003, 36, 721–727. [Google Scholar] [CrossRef]
- Xu, X. Short-Path Distillation for Lipid Processing. In Healthful Lipids; AOCS Publishing: New York, NY, USA, 2019; pp. 127–144. ISBN 9780429104497. [Google Scholar]
- Solaesa, Á.G.; Sanz, M.T.; Falkeborg, M.; Beltrán, S.; Guo, Z. Production and concentration of monoacylglycerols rich in omega-3 polyunsaturated fatty acids by enzymatic glycerolysis and molecular distillation. Food Chem. 2016, 190, 960–967. [Google Scholar] [CrossRef]
- Mylonas, C.C.; de la Gándara, F.; Corriero, A.; Ríos, A.B. Atlantic bluefin tuna (Thunnus thynnus) farming and fattening in the mediterranean sea. Rev. Fish. Sci. 2010, 18, 266–280. [Google Scholar] [CrossRef]
- Šimat, V.; Vlahović, J.; Soldo, B.; Generalić Mekinić, I.; Čagalj, M.; Hamed, I.; Skroza, D. Production and characterization of crude oils from seafood processing by-products. Food Biosci. 2020, 33, 100484. [Google Scholar] [CrossRef]
- Topic Popovic, N.; Kozacinski, L.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Jadan, M.; Cvrtila-Fleck, Z.; Barisic, J. Fatty acid and proximate composition of bluefin tuna (Thunnus thynnus) muscle with regard to plasma lipids. Aquac. Res. 2012, 43, 722–729. [Google Scholar] [CrossRef]
- Schneider, M. Marine phospholipids and their applications: Next-generation omega-3 lipids. In Omega-6/3 Fatty Acids: Functions, Sustainability Strategies and Perspectives; Humana Press: Totowa, NJ, USA, 2013; pp. 297–308. ISBN 9781627032155. [Google Scholar]
- Ferdosh, S.; Sarker, M.Z.I.; Rahman, N.N.N.A.; Akand, M.J.H.; Ghafoor, K.; Awang, M.B.; Kadir, M.O.A. Supercritical carbon dioxide extraction of oil from Thunnus tonggol head by optimization of process parameters using response surface methodology. Korean J. Chem. Eng. 2013, 30, 1466–1472. [Google Scholar] [CrossRef]
- de la Fuente, B.; Pinela, J.; Mandim, F.; Heleno, S.A.; Ferreira, I.C.F.R.; Barba, F.J.; Berrada, H.; Caleja, C.; Barros, L. Nutritional and bioactive oils from salmon (Salmo salar) side streams obtained by Soxhlet and optimized microwave-assisted extraction. Food Chem. 2022, 386, 132778. [Google Scholar] [CrossRef] [PubMed]
- Othman, O.C.; Ngassapa, F.N. Physicochemical characteristics of some imported edible vegetable oils and fat marketed in Dares Salaam. Tanzan. J. Nat. Appl. Sci. 2010, 1, 138–147. [Google Scholar]
- Bako, T.; Umogbai, V.I.; Awulu, J.O. Criteria for the extraction of fish oil. Agric. Eng. Int. CIGR J. 2017, 19, 120–132. [Google Scholar]
- Codex Alimentarius Commission. Standards for Fish Oils; Codex Alimentarius Commission: Rome, Italy, 2017. [Google Scholar]
- Franklin, E.C.; Haq, M.; Roy, V.C.; Park, J.S.; Chun, B.S. Supercritical CO2 extraction and quality comparison of lipids from Yellowtail fish (Seriola quinqueradiata) waste in different conditions. J. Food Process. Preserv. 2020, 44, e14892. [Google Scholar] [CrossRef]
- Chakraborty, K.; Joseph, D. Production and characterization of refined oils obtained from Indian oil sardine (Sardinella longiceps). J. Agric. Food Chem. 2015, 63, 998–1009. [Google Scholar] [CrossRef]
- Crexi, V.T.; Monte, M.L.; Monte, M.L.; Pinto, L.A.A. Polyunsaturated fatty acid concentrates of carp oil: Chemical hydrolysis and urea complexation. JAOCS J. Am. Oil Chem. Soc. 2012, 89, 329–334. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards Scientific Opinion on Fish Oil for Human Consumption Food Hygiene, including Rancidity. EFSA J. 2010, 8, 1874. [CrossRef]
- Magallanes, L.M.; Tarditto, L.V.; Grosso, N.R.; Pramparo, M.C.; Gayol, M.F. Highly concentrated omega-3 fatty acid ethyl esters by urea complexation and molecular distillation. J. Sci. Food Agric. 2019, 99, 877–884. [Google Scholar] [CrossRef]
- González-Fernández, M.J.; Fabrikov, D.; Lyashenko, S.; Ferrón-Carrillo, F.; Guil-Guerrero, J.L. Highly concentrated very long-chain PUFA obtainment by Urea complexation methodology. Environ. Technol. Innov. 2020, 18, 100736. [Google Scholar] [CrossRef]
- Lopes da Silva, T.; Santos, A.R.; Gomes, R.; Reis, A. Valorizing fish canning industry by-products to produce ω-3 compounds and biodiesel. Environ. Technol. Innov. 2018, 9, 74–81. [Google Scholar] [CrossRef]
- Hayes, D.G. Effect of temperature programming on the performance of urea inclusion compound-based free fatty acid fractionation. JAOCS J. Am. Oil Chem. Soc. 2006, 83, 253–259. [Google Scholar] [CrossRef]
- Görgün, S.; Zengin, G. Determination of fatty acid profiles and esterase activities in the gills and gonads of Vimba vimba (L., 1758). JAOCS J. Am. Oil Chem. Soc. 2015, 92, 353–360. [Google Scholar] [CrossRef]
- Jankowska, B.; Zakeś, Z.; Zmijewski, T.; Szczepkowski, M. Fatty acid profile of muscles, liver and mesenteric fat in wild and reared perch (Perca fluviatilis L.). Food Chem. 2010, 118, 764–768. [Google Scholar] [CrossRef]
- Oliveira, A.C.M.; Miller, M.R. Purification of Alaskan walleye pollock (Gadus chalcogrammus) and New Zealand hoki (Macruronus novaezelandiae) liver oil using short path distillation. Nutrients 2014, 6, 2059–2076. [Google Scholar] [CrossRef]
- Liang, J.H.; Hwang, L.S. Fractionation of squid visceral oil ethyl esters by short-path distillation. JAOCS J. Am. Oil Chem. Soc. 2000, 77, 773–777. [Google Scholar] [CrossRef]
- Marsol-Vall, A.; Aitta, E.; Guo, Z.; Yang, B. Green technologies for production of oils rich in n-3 polyunsaturated fatty acids from aquatic sources. Crit. Rev. Food Sci. Nutr. 2020, 62, 2942–2962. [Google Scholar] [CrossRef]
- Wang, W.; Li, T.; Ning, Z.; Wang, Y.; Yang, B.; Ma, Y.; Yang, X. A process for the synthesis of PUFA-enriched triglycerides from high-acid crude fish oil. J. Food Eng. 2012, 109, 366–371. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Xie, D.; Zou, S.; Jin, Q.; Wang, X. Synthesis and concentration of 2-monoacylglycerols rich in polyunsaturated fatty acids. Food Chem. 2018, 250, 60–66. [Google Scholar] [CrossRef]
- Valverde, L.M.; Moreno, P.A.G.; Callejón, M.J.J.; Cerdán, L.E.; Medina, A.R. Concentration of eicosapentaenoic acid (EPA) by selective alcoholysis catalyzed by lipases. Eur. J. Lipid Sci. Technol. 2013, 115, 990–1004. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Sánchez-Moya, A.; Balbuena-Pecino, S.; Fontanillas, R.; Gutiérrez, J.; Capilla, E.; Navarro, I. The combination of palm and rapeseed oils emerges as a good dietary alternative for optimal growth and balanced lipid accumulation in juvenile gilthead sea bream reared at an elevated temperature. Aquaculture 2020, 526, 735396. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Chen, H.Y. Effects of different fatty acids on cell differentiation and lipid accumulation in preadipocytes of warm water fish cobia (Rachycentron canadum Linnaeus, 1766). Aquac. Res. 2015, 46, 590–601. [Google Scholar] [CrossRef]
- Huang, T.S.; Todorčević, M.; Ruyter, B.; Torstensen, B.E. Altered expression of CCAAT/enhancer binding protein and FABP11 genes during adipogenesis in vitro in Atlantic salmon (Salmo salar). Aquac. Nutr. 2010, 16, 72–80. [Google Scholar] [CrossRef]
- Vegusdal, A.; Sundvold, H.; Gjøen, T.; Ruyter, B. An in vitro method for studying the proliferation and differentiation of Atlantic salmon preadipocytes. Lipids 2003, 38, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Riera-heredia, N.; Lutfi, E.; Sánchez-moya, A.; Gutiérrez, J.; Capilla, E.; Navarro, I. Short-term responses to fatty acids on lipid metabolism and adipogenesis in rainbow trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2020, 21, 1623. [Google Scholar] [CrossRef] [PubMed]
- Bou, M.; Montfort, J.; Le Cam, A.; Rallière, C.; Lebret, V.; Gabillard, J.C.; Weil, C.; Gutiérrez, J.; Rescan, P.Y.; Capilla, E.; et al. Gene expression profile during proliferation and differentiation of rainbow trout adipocyte precursor cells. BMC Genom. 2017, 18, 347. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Rong, Y.; Bao, L.; Nie, B.; Ren, G.; Zheng, C.; Amin, R.; Arnold, R.D.; Jeganathan, R.B.; Huggins, K.W. Suppression of adipocyte differentiation and lipid accumulation by stearidonic acid (SDA) in 3T3-L1 cells. Lipids Health Dis. 2017, 16, 181. [Google Scholar] [CrossRef] [PubMed]
- Torstensen, B.E.; Nanton, D.A.; Olsvik, P.A.; Sundvold, H.; Stubhaug, I. Gene expression of fatty acid-binding proteins, fatty acid transport proteins (cd36 and FATP) and β-oxidation-related genes in Atlantic salmon (Salmo salar L.) fed fish oil or vegetable oil. Aquac. Nutr. 2009, 15, 440–451. [Google Scholar] [CrossRef]
- Lei, C.; Ji, H.; Zhang, J.; Li, J. Effects of Dietary DHA/EPA Ratios on Fatty Acid Composition, Lipid Metabolism-related Enzyme Activity, and Gene Expression of Juvenile Grass Carp, Ctenopharyngodon idellus. J. World Aquac. Soc. 2016, 47, 287–296. [Google Scholar] [CrossRef]
- Li, C.; Liu, P.; Ji, H.; Huang, J.; Zhang, W. Dietary n-3 highly unsaturated fatty acids affect the biological and serum biochemical parameters, tissue fatty acid profile, antioxidation status and expression of lipid-metabolism-related genes in grass carp, Ctenopharyngodon idellus. Aquac. Nutr. 2015, 21, 373–383. [Google Scholar] [CrossRef]
- Barber, E.; Sinclair, A.J.; Cameron-Smith, D. Comparative actions of omega-3 fatty acids on in-vitro lipid droplet formation. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 359–366. [Google Scholar] [CrossRef]
- Prostek, A.; Gajewska, M.; Kamola, D.; Bałasińska, B. The influence of EPA and DHA on markers of inflammation in 3T3-L1 cells at different stages of cellular maturation. Lipids Health Dis. 2014, 13, 3. [Google Scholar] [CrossRef]
- Wójcik, C.; Lohe, K.; Kuang, C.; Xiao, Y.; Jouni, Z.; Poels, E. Modulation of adipocyte differentiation by omega-3 polyunsaturated fatty acids involves the ubiquitin-proteasome system. J. Cell. Mol. Med. 2014, 18, 590–599. [Google Scholar] [CrossRef]
- Vagner, M.; Santigosa, E. Characterization and modulation of gene expression and enzymatic activity of delta-6 desaturase in teleosts: A review. Aquaculture 2011, 315, 131–143. [Google Scholar] [CrossRef]
- Huang, C.W.; Chien, Y.S.; Chen, Y.J.; Ajuwon, K.M.; Mersmann, H.M.; Ding, S.T. Role of n-3 polyunsaturated fatty acids in ameliorating the obesity-induced metabolic syndrome in animal models and humans. Int. J. Mol. Sci. 2016, 17, 1689. [Google Scholar] [CrossRef]
- Seiliez, I.; Panserat, S.; Corraze, G.; Kaushik, S.; Bergot, P. Cloning and nutritional regulation of a Δ6-desaturase-like enzyme in the marine teleost gilthead seabream (Sparus aurata). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 135, 449–460. [Google Scholar] [CrossRef]
- Gregory, M.K.; Collins, R.O.; Tocher, D.R.; James, M.J.; Turchini, G.M. Nutritional regulation of long-chain PUFA biosynthetic genes in rainbow trout (Oncorhynchus mykiss). Br. J. Nutr. 2016, 115, 1721–1729. [Google Scholar] [CrossRef]
- Silva-Brito, F.; Magnoni, L.J.; Fonseca, S.B.; Peixoto, M.J.; Castro, L.F.C.; Cunha, I.; De Almeida Ozório, R.O.A.; Magalhães, F.; Gonçalves, J.F.M. Dietary Oil Source and Selenium Supplementation Modulate Fads2 and Elovl5 Transcriptional Levels in Liver and Brain of Meagre (Argyrosomus regius); Dietary Oil Source and Selenium Supplementation Modulate Fads2 and Elovl5. Lipids 2016, 51, 729–741. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, Z.; Wang, Y.; Wu, Q.; Sun, Y.; Gao, Y. Impact of dietary docosahexaenoic to eicosapentaenoic acid ratio (DHA/EPA) on growth, fatty acid composition, lipid metabolism and antioxidant status of juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). N. Am. J. Aquacul. 2021. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Lutfi, E.; Gutiérrez, J.; Navarro, I.; Capilla, E. Fatty acids from fish or vegetable oils promote the adipogenic fate of mesenchymal stem cells derived from gilthead sea bream bone potentially through different pathways. PLoS ONE 2019, 14, e0215926. [Google Scholar] [CrossRef]
- Basto-Silva, C.; Balbuena-Pecino, S.; Oliva-Teles, A.; Riera-Heredia, N.; Navarro, I.; Guerreiro, I.; Capilla, E. Gilthead seabream (Sparus aurata) in vitro adipogenesis and its endocrine regulation by leptin, ghrelin, and insulin. Comp. Biochem. Physiol.-Part A Mol. Integr. Physiol. 2020, 249, 110772. [Google Scholar] [CrossRef]
- Sun, L.; Nicholson, A.C.; Hajjar, D.P.; Gotto, A.M.; Han, J. Adipogenic differentiating agents regulate expression of fatty acid binding protein and CD36 in the J744 macrophage cell line. J. Lipid Res. 2003, 44, 1877–1886. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis; Association of Official Analytical Chemist: Washington, DC, USA, 1990. [Google Scholar]
- AOAC Official Method 981.10 Crude Protein in Meat Block Digestion Method; Association of Official Analytical Chemist: Washington, DC, USA, 1992.
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Lubis, Z.; Buckle, K.A. Rancidity and lipid oxidation of dried-salted sardines. Int. J. Food Sci. Technol. 1990, 25, 295–303. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Sathivel, S.; Prinyawiwatkul, W.; King, J.M.; Grimm, C.C.; Lloyd, S. Oil production from catfish viscera. JAOCS J. Am. Oil Chem. Soc. 2003, 80, 377–382. [Google Scholar] [CrossRef]
- Vázquez, L.; Akoh, C.C. Fractionation of short and medium chain fatty acid ethyl esters from a blend of oils via ethanolysis and short-path distillation. JAOCS J. Am. Oil Chem. Soc. 2010, 87, 917–928. [Google Scholar] [CrossRef]
- Hsieh, C.W.; Chang, C.J.; Ko, W.C. Supercritical CO2 extraction and concentration of n-3 polyunsaturated fatty acid ethyl esters from tuna cooking juice. Fish. Sci. 2005, 71, 441–447. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Moisture | Ash | Total Lipid | Protein | |
|---|---|---|---|---|
| BFT-W | 62.1 ± 2.0 c | 4.8 ± 0.4 a | 19.6 ± 1.5 b | 13.3 ± 3.4 a |
| BFT-F > 200 kg | 46.8 ± 1.8 a | 5.52 ± 2.1 a | 32.1 ± 2.1 c | 14.2 ± 1.8 a |
| BFT-F < 60 kg | 57.1 ± 1.1 b | 4.25 ± 0.5 a | 22.5 ± 1.2 b | 19.2 ± 2.4 b |
| YFT-W | 61.0 ± 1.5 c | 6.4 ± 1.1 b | 10.8 ± 0.8 a | 20.5 ± 3.2 c |
| FA | BFT-F |
|---|---|
| 14:0 | 5.79 ± 0.18 |
| 16:0 | 20.16 ± 0.96 |
| 16:1ω-7 | 6.84 ± 0.45 |
| 16:2ω-4 | 1.03 ± 0.02 |
| 16:3ω-4 | 0.56 ± 0.19 |
| 18:0 | 4.43 ± 0.13 |
| 18:1ω-9 | 15.71 ± 0.61 |
| 18:1ω-7 | 6.78 ± 0.27 |
| 18:2ω-6 | 1.22 ± 0.03 |
| 18:3ω-3 | 6.03 ± 0.59 |
| 20:1ω-9 | 2.77 ± 0.09 |
| 20:4ω-6 | 0.54 ± 0.01 |
| 20:4ω-3 | 0.29 ± 0.01 |
| 20:5ω-3 (EPA) | 9.93 ± 0.36 |
| 22:1ω-11 | 4.30 ± 0.57 |
| 22:6ω-3 (DHA) | 13.64 ± 0.56 |
| SFA | 30.38 ± 0.71 |
| MUFA | 36.40 ± 1.36 |
| PUFA | 33.22 ± 1.72 |
| Totω-3 | 29.89 ± 1.49 |
| Totω-6 | 1.75 ± 0.03 |
| EPA + DHA | 23.57 ± 0.92 |
| PI | 1.17 ± 0.09 |
| AI | 0.62 ± 0.01 |
| AT | 0.24 ± 0.01 |
| TO | CO | |
|---|---|---|
| PV (meq O2/kg) | 2.96 ± 0.55 b | 2.10 ± 0.53 a |
| ρ-AV | 12.93 ± 1.53 b | 4.96 ± 0.89 a |
| TBARS (MDA µg/g) | 11.81 ± 2.45 b | 5.51 ± 0.81 a |
| TOTOX | 20.80 ± 0.88 b | 10.83 ± 0.24 a |
| Acid value (FFA%) | 2.25 ± 0.32 b | 0.50 ± 0.04 a |
| FA | TO | TFA | PUFA-Ue |
|---|---|---|---|
| 14:0 | 6.82 ± 0.11 b | 6.99 ± 0.10 b | 4.37 ± 0.18 a |
| 16:0 | 21.15 ± 0.31 c | 19.38 ± 0.22 b | 9.21 ± 0.23 a |
| 16:1ω-7 | 7.27 ± 0.09 c | 6.24 ± 0.09 b | 4.09 ± 0.06 a |
| 16:2ω-4 | 1.53 ± 0.19 b | 1.34 ± 0.11 b | 0.27 ± 0.03 a |
| 16:3ω-4 | 0.84 ± 0.13 b | 0.79 ± 0.03 b | 0.41 ± 0.08 a |
| 18:0 | 3.75 ± 0.22 b | 2.85 ± 0.12 a | 3.73 ± 0.05 b |
| 18:1ω-9 | 15.42 ± 0.4 c | 13.80 ± 0.3 b | 8.52 ± 0.35 a |
| 18:1ω-7 | 6.66 ± 0.13 b | 6.96 ± 0.31 b | 3.67 ± 0.28 a |
| 18:2ω-6 | 1.21 ± 0.02 a | 3.25 ± 0.08 b | 5.29 ± 0.09 c |
| 18:3ω-6 | 0.44 ± 0.29 a | 0.99 ± 0.25 a | 2.50 ± 0.12 b |
| 18:3ω-3 | 0.92 ± 0.09 a | 0.89 ± 0.15 a | 8.47 ± 0.1 b |
| 20:1ω-9 | 3.00 ± 0.42 a | 3.04 ± 0.18 a | 3.66 ± 0.34 a |
| 20:4ω-6 | 0.59 ± 0.04 a | 0.34 ± 0.21 a | 2.38 ± 0.22 b |
| 20:4ω-3 | 0.44 ± 0.08 a | 0.20 ± 0.13 a | 1.78 ± 0.07 b |
| 20:5ω-3 | 9.73 ± 0.35 a | 10.26 ± 0.05 a | 10.96 ± 0.03 b |
| 22:1ω-11 | 4.84 ± 0.35 a | 5.02 ± 0.32 a | 5.06 ± 0.18 a |
| 22:1ω-9 | 0.16 ± 0.05 a | 0.14 ± 0.03 a | 0.16 ± 0.04 a |
| 22:4ω-6 | 0.41 ± 0.10 a | 1.37 ± 0.29 b | 1.53 ± 0.09 c |
| 22:5ω-3 | 1.21 ± 0.05 a | 1.25 ± 0.09 a | 1.43 ± 0.012 b |
| 22:6ω-3 | 13.60 ± 0.08 a | 14.90 ± 0.16 b | 22.50 ± 0.45 c |
| SFA | 31.72 ± 0.90 a | 29.23 ± 1.21 a | 17.31 ± 1.51 b |
| MUFA | 37.36 ± 1.88 c | 35.19 ± 1.18 b | 25.16 ± 1.18 a |
| PUFA | 30.92 ± 1.08 a | 35.58 ± 1.19 b | 57.53 ± 1.35 c |
| Totω-3 | 25.90 ± 0.65 a | 27.50 ± 0.95 b | 45.14 ± 1.75 c |
| Totω-6 | 2.20 ± 0.38 a | 4.96 ± 0.34 b | 9.20 ± 0.86 c |
| EPA + DHA | 23.33 ± 0.98 a | 25.15 ± 0.75 b | 33.46 ± 1.02 c |
| PI | 1.10 ± 0.02 a | 1.30 ± 0.03 b | 3.63 ± 0.25 c |
| AI | 0.71 ± 0.09 b | 0.67 ± 0.09 b | 0.32 ± 0.02 a |
| TI | 0.29 ± 0.04 b | 0.27 ± 0.02 b | 0.11 ± 0.01 a |
| FA | PUFA-Ue | PUFA-SPe | PUFA-SPd |
|---|---|---|---|
| 14:0 | 4.37 ± 0.18 b | 0.86 ± 0.03 a | 6.39 ± 0.49 c |
| 16:0 | 9.21 ± 0.23 b | 2.23 ± 0.06 a | 14.38 ± 0.53 c |
| 16:1ω-7 | 4.09 ± 0.06 b | 0.99 ± 0.01 a | 5.99 ± 0.12 c |
| 16:2ω-4 | 0.27 ± 0.03 b | 0.22 ± 0.01 b | 0.07 ± 0.03 a |
| 16:3ω-4 | 0.41 ± 0.08 b | 0.26 ± 0.00 a | 0.80 ± 0.12 c |
| 18:0 | 3.73 ± 0.05 b | 1.64 ± 0.06 a | 4.37 ± 0.24 c |
| 18:1ω-9 | 8.52 ± 0.35 b | 2.39 ± 0.19 a | 14.58 ± 0.11 c |
| 18:1ω-7 | 3.67 ± 0.28 b | 0.95 ± 0.03 a | 4.52 ± 0.14 c |
| 18:2ω-6 | 5.29 ± 0.09 b | 3.14 ± 0.06 a | 5.29 ± 0.67 a |
| 18:3ω-6 | 2.50 ± 0.12 b | 1.48 ± 0.16 a | 2.86 ± 0.23 b |
| 18:3ω-3 | 8.47 ± 0.10 b | 8.33 ± 0.10 b | 4.99 ± 0.28 a |
| 20:1ω-9 | 3.66 ± 0.34 b | 1.29 ± 0.06 a | 5.70 ± 0.27 a |
| 20:4ω-6 | 2.38 ± 0.22 b | 2.63 ± 0.13 b | 0.48 ± 0.08 a |
| 20:4ω-3 | 1.78 ± 0.07 b | 1.77 ± 0.00 b | 0.85 ± 0.06 a |
| 20:5ω-3 | 10.96 ± 0.03 b | 17.59 ± 0.59 c | 4.85 ± 0.45 a |
| 22:1ω-11 | 5.06 ± 0.18 b | 2.10 ± 0.06 a | 5.20 ± 0.17 b |
| 22:1ω-9 | 0.16 ± 0.04 a | 0.79 ± 0.00 b | 0.26 ± 0.05 a |
| 22:4ω-6 | 1.53 ± 0.09 b | 1.86 ± 0.03 c | 0.68 ± 0.13 a |
| 22:5ω-3 | 1.43 ± 0.12 a | 2.97 ± 0.20 b | 1.49 ± 0.01 a |
| 22:6ω-3 | 22.50 ± 0.45 b | 46.51 ± 0.34 c | 16.25 ± 0.14 a |
| SFA | 17.31 ± 1.51 b | 4.73 ± 0.03 a | 25.13 ± 0.20 c |
| MUFA | 25.16 ± 1.18 b | 8.51 ± 0.10 a | 36.26 ± 0.42 c |
| PUFA | 57.53 ± 1.35 b | 86.76 ± 0.07 c | 38.61 ± 0.22 a |
| Totω-3 | 45.14 ± 1.75 b | 77.17 ± 0.24 c | 28.44 ± 0.52 a |
| Totω-6 | 9.20 ± 0.86 c | 7.63 ± 0.20 b | 6.44 ± 0.87 a |
| EPA + DHA | 33.46 ± 1.02 b | 64.10 ± 0.25 c | 21.09 ± 0.31 a |
| PI | 3.63 ± 0.25 b | 28.81 ± 0.95 c | 1.47 ± 0.03 a |
| IA | 0.32 ± 0.02 b | 0.06 ± 0.00 a | 0.53 ± 0.02 c |
| IT | 0.11 ± 0.01 b | 0.02 ± 0.00 a | 0.23 ± 0.01 c |
| R | 1.89 ± 0.23 b | 13.95 ± 0.83 c | 0.73 ± 0.01 a |
| EPA Enrichment Factor | 1.00 ± 0.07 b | 1.54 ± 0.05 c | 0.90 ± 0.08 a |
| DHA Enrichment Factor | 1.00 ± 0.07 b | 1.99 ± 0.01 c | 0.69 ± 0.01 a |
| PUFA Enrichment Factor | 1.00 ± 0.07 b | 1.45 ± 0.02 c | 0.70 ± 0.01 a |
| PUFA/SFA | 3.32 ± 0.25 b | 18.35 ± 0.10 c | 1.54 ± 0.01 a |
| Gene | F/R Primer Sequence (5′–3′) |
|---|---|
| pparβ | Fw CAGGTGACTTTGCTGAAGTA |
| pparγ | Rv TAAAGGGCTTTCTTAAGCTG |
| Fw CTCAAGAGTCTCAGGAAACC | |
| D6D | Rv GATAATGACAGCCAGAAACA |
| Fw TACCTTCGCTACCTGTGCTG | |
| fas | Rv TTGCAGGTGGTTGTAACTG |
| Fw TGCCATTGCCATAGCACTCA | |
| cd36 | Rv ACCTTTGCCCTTTGTGTGGA |
| Fw GTCGTGGCTCAAGTCTTCCA | |
| fatp1 | Rv TTTCCCGTGGCCTGTATTCC |
| Fw CAACAGAGGTGGAGGGCATT | |
| fabp1 | Rv GGGGAGATACGCAGGAACAC |
| Fw CATGAAGGCGATTGGTCTCC | |
| Rv GTCTCCAAGTCTGCCTCCTT | |
| 18s | Fw TGTGCCGCTAGAGGTGAAATT |
| Rv GCAAATGCTTTCGCTTTCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, C.M.; Arena, R.; Manuguerra, S.; La Barbera, L.; Curcuraci, E.; Renda, G.; Santulli, A. Valorization of Side Stream Products from Sea Cage Fattened Bluefin Tuna (Thunnus thynnus): Production and In Vitro Bioactivity Evaluation of Enriched ω-3 Polyunsaturated Fatty Acids. Mar. Drugs 2022, 20, 309. https://doi.org/10.3390/md20050309
Messina CM, Arena R, Manuguerra S, La Barbera L, Curcuraci E, Renda G, Santulli A. Valorization of Side Stream Products from Sea Cage Fattened Bluefin Tuna (Thunnus thynnus): Production and In Vitro Bioactivity Evaluation of Enriched ω-3 Polyunsaturated Fatty Acids. Marine Drugs. 2022; 20(5):309. https://doi.org/10.3390/md20050309
Chicago/Turabian StyleMessina, Concetta Maria, Rosaria Arena, Simona Manuguerra, Laura La Barbera, Eleonora Curcuraci, Giuseppe Renda, and Andrea Santulli. 2022. "Valorization of Side Stream Products from Sea Cage Fattened Bluefin Tuna (Thunnus thynnus): Production and In Vitro Bioactivity Evaluation of Enriched ω-3 Polyunsaturated Fatty Acids" Marine Drugs 20, no. 5: 309. https://doi.org/10.3390/md20050309
APA StyleMessina, C. M., Arena, R., Manuguerra, S., La Barbera, L., Curcuraci, E., Renda, G., & Santulli, A. (2022). Valorization of Side Stream Products from Sea Cage Fattened Bluefin Tuna (Thunnus thynnus): Production and In Vitro Bioactivity Evaluation of Enriched ω-3 Polyunsaturated Fatty Acids. Marine Drugs, 20(5), 309. https://doi.org/10.3390/md20050309