
A Tale of Toxin Promiscuity: The Versatile Pharmacological Effects of Hcr 1b-2 Sea Anemone Peptide on Voltage-Gated Ion Channels

 ,
,  ,
, 
Abstract
1. Introduction
2. Results
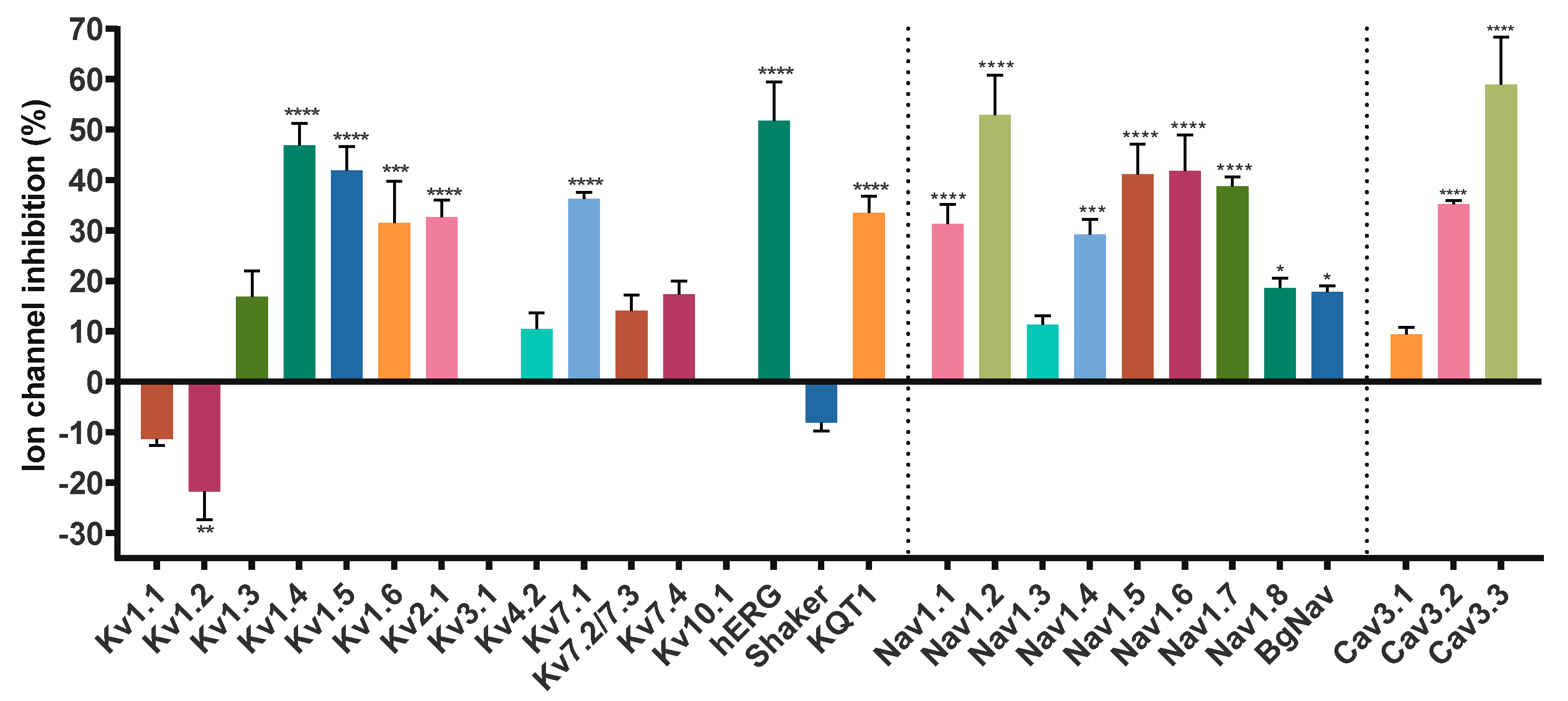
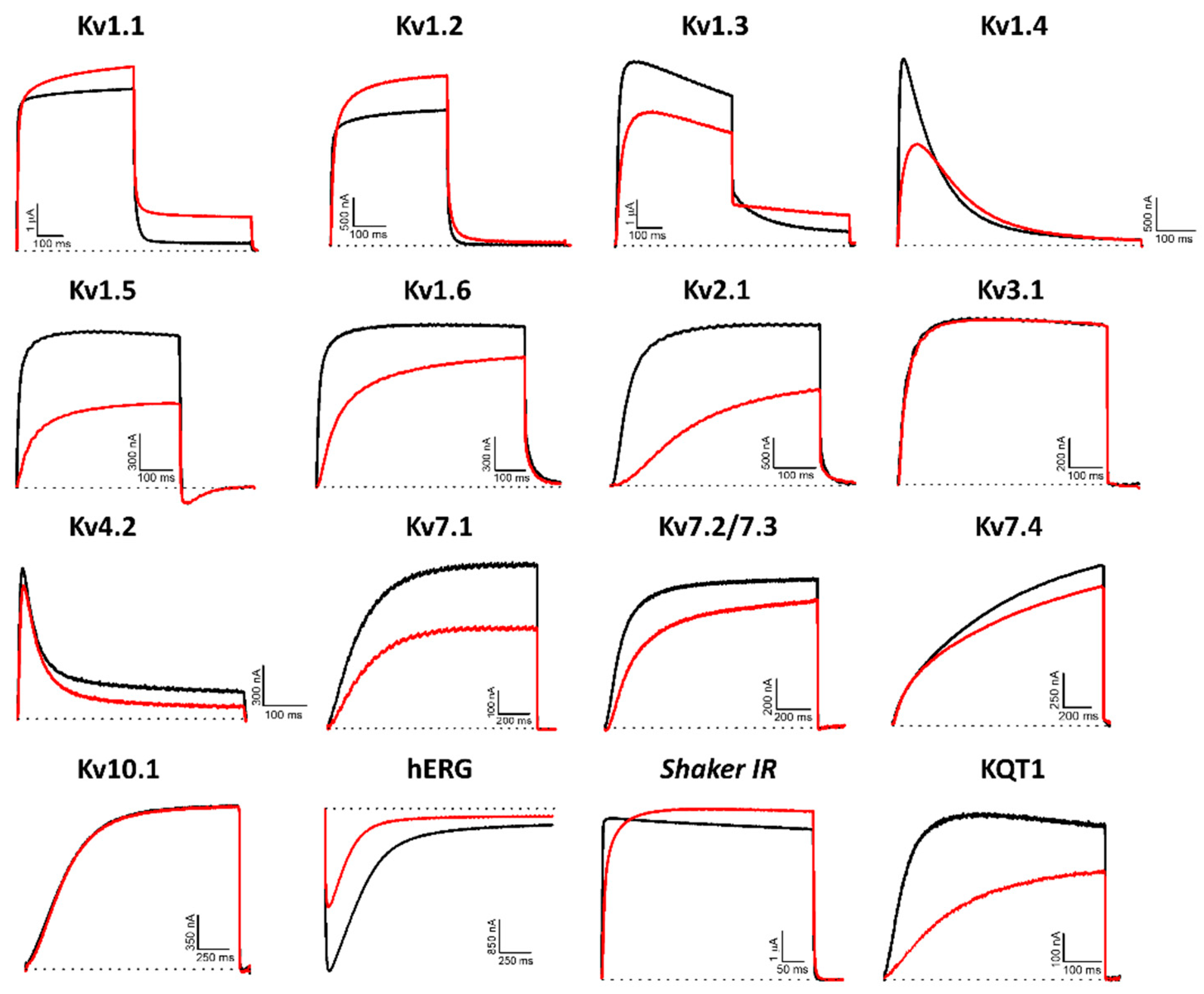
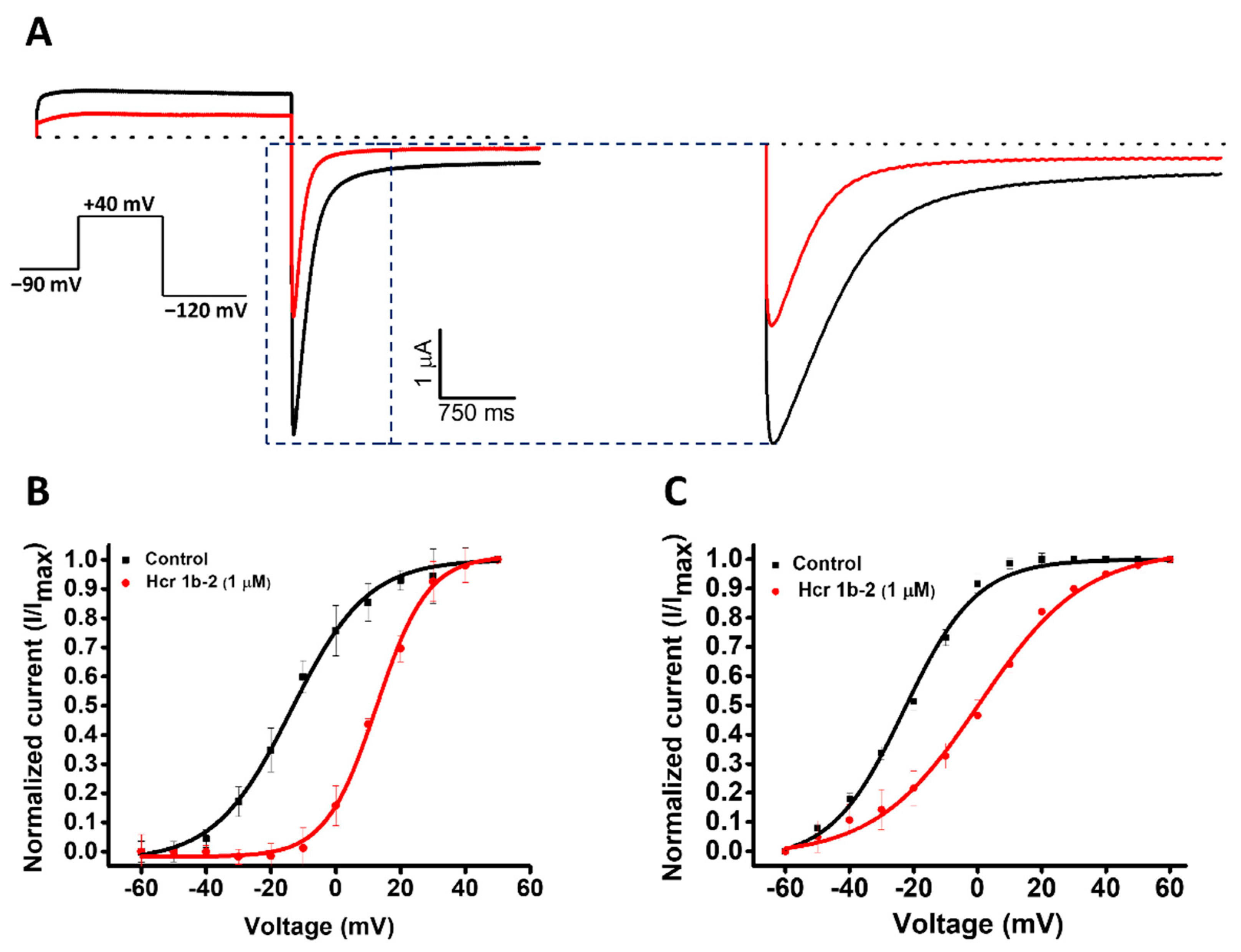
2.1. Modulation of Kv Channels
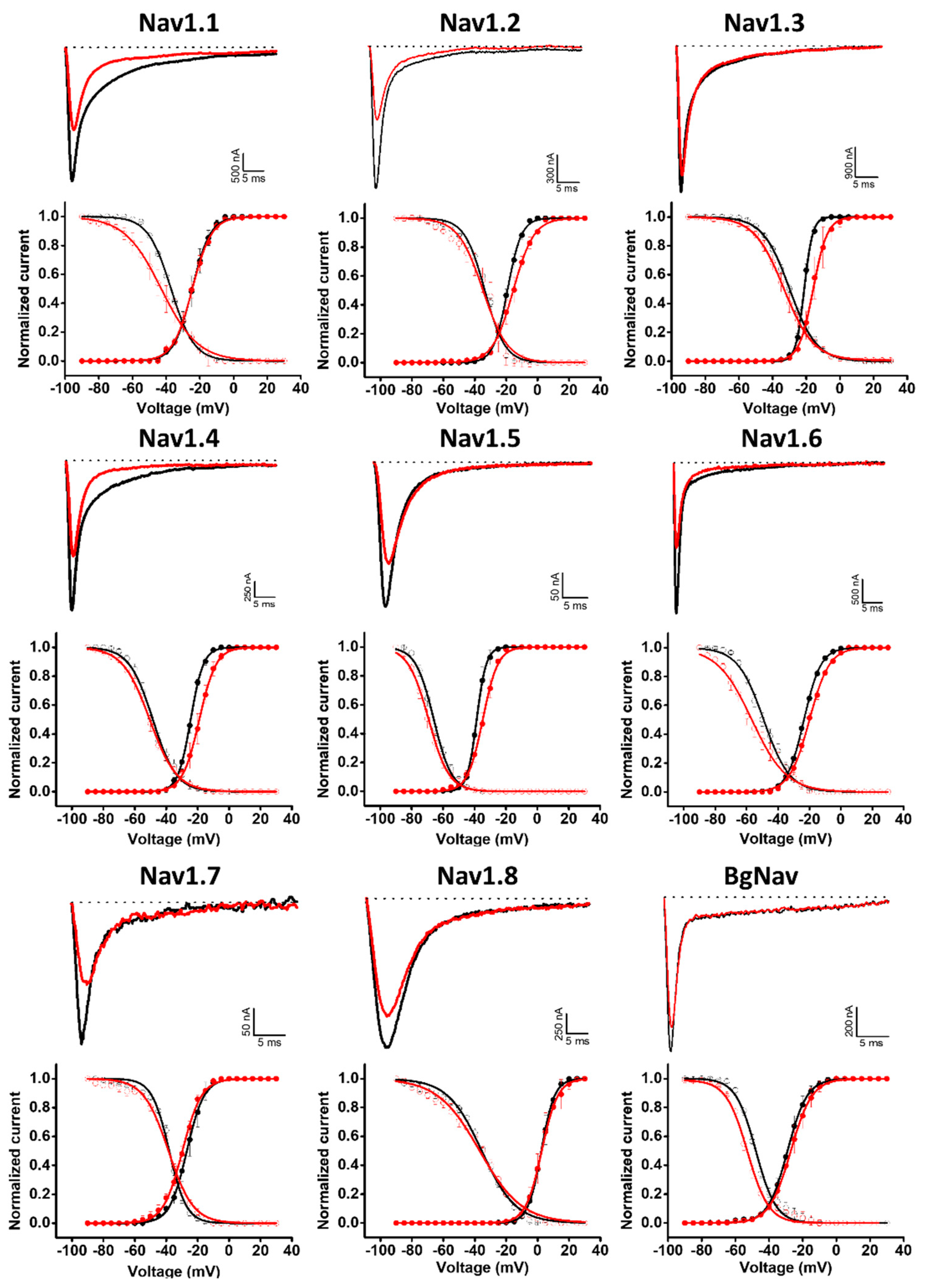
2.2. Modulation of Nav Channels
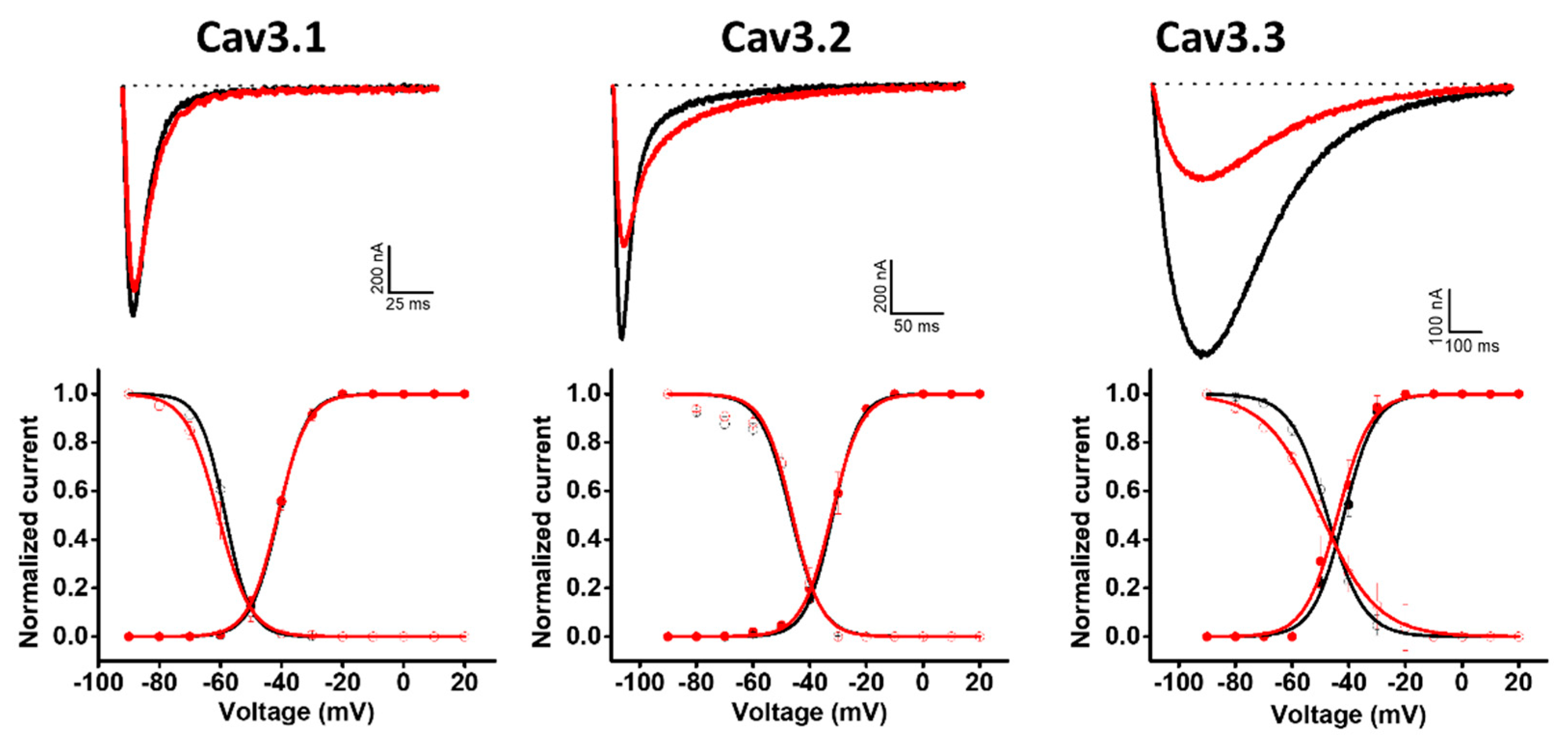
2.3. Modulation of Cav Channels
3. Discussion
4. Materials and Methods
4.1. Peptide Isolation and Primary Structure Determination
4.2. Electrophysiological Assays
4.2.1. Expression of Ion Channels in Xenopus Laevis Oocytes
4.2.2. Electrophysiological Recordings
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Junior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liao, Y.; Jin, A.H.; Gao, B. Discovery of novel peptide neurotoxins from sea anemone species. Front. Biosci. 2021, 26, 1256–1273. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Leychenko, E.; Isaeva, M.; Tkacheva, E.; Zelepuga, E.; Kvetkina, A.; Guzev, K.; Monastyrnaya, M.; Kozlovskaya, E. Multigene Family of Pore-Forming Toxins from Sea Anemone Heteractis crispa. Mar. Drugs 2018, 16, 183. [Google Scholar] [CrossRef] [PubMed]
- Gladkikh, I.; Peigneur, S.; Sintsova, O.; Lopes Pinheiro-Junior, E.; Klimovich, A.; Menshov, A.; Kalinovsky, A.; Isaeva, M.; Monastyrnaya, M.; Kozlovskaya, E.; et al. Kunitz-Type Peptides from the Sea Anemone Heteractis crispa Demonstrate Potassium Channel Blocking and Anti-Inflammatory Activities. Biomedicines 2020, 8, 473. [Google Scholar] [CrossRef] [PubMed]
- Razpotnik, A.; Krizaj, I.; Sribar, J.; Kordis, D.; Macek, P.; Frangez, R.; Kem, W.R.; Turk, T. A new phospholipase A2 isolated from the sea anemone Urticina crassicornis—Its primary structure and phylogenetic classification. FEBS J. 2010, 277, 2641–2653. [Google Scholar] [CrossRef]
- Cooper, R.A.; de Freitas, J.C.; Porreca, F.; Eisenhour, C.M.; Lukas, R.; Huxtable, R.J. The sea anemone purine, caissarone: Adenosine receptor antagonism. Toxicon 1995, 33, 1025–1031. [Google Scholar] [CrossRef]
- Ferreira Junior, W.A.; Zaharenko, A.J.; Kazuma, K.; Picolo, G.; Gutierrez, V.P.; de Freitas, J.C.; Konno, K.; Cury, Y. Peripheral 5-HT3 Receptors Are Involved in the Antinociceptive Effect of Bunodosine 391. Toxins 2017, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B. Channelopathies. Korean J. Pediatr. 2014, 57, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Imbrici, P.; Liantonio, A.; Camerino, G.M.; De Bellis, M.; Camerino, C.; Mele, A.; Giustino, A.; Pierno, S.; De Luca, A.; Tricarico, D.; et al. Therapeutic Approaches to Genetic Ion Channelopathies and Perspectives in Drug Discovery. Front. Pharmacol. 2016, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Wulff, H.; Castle, N.A.; Pardo, L.A. Voltage-gated potassium channels as therapeutic targets. Nat. Rev. Drug Discov. 2009, 8, 982–1001. [Google Scholar] [CrossRef] [PubMed]
- Cologna, C.T.; Peigneur, S.; Rustiguel, J.K.; Nonato, M.C.; Tytgat, J.; Arantes, E.C. Investigation of the relationship between the structure and function of Ts2, a neurotoxin from Tityus serrulatus venom. FEBS J. 2012, 279, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Peng, C.; Yi, Y.; Gao, B.; Shi, Q. A Transcriptomic Survey of Ion Channel-Based Conotoxins in the Chinese Tubular Cone Snail (Conus betulinus). Mar. Drugs 2017, 15, 228. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Paiva, A.L.B.; Cordeiro, M.N.; Borges, M.H.; Diniz, M.R.V.; de Lima, M.E.; Tytgat, J. Phoneutria nigriventer Spider Toxin PnTx2-1 (delta-Ctenitoxin-Pn1a) Is a Modulator of Sodium Channel Gating. Toxins 2018, 10, 337. [Google Scholar] [CrossRef]
- Kalina, R.; Gladkikh, I.; Dmitrenok, P.; Chernikov, O.; Koshelev, S.; Kvetkina, A.; Kozlov, S.; Kozlovskaya, E.; Monastyrnaya, M. New APETx-like peptides from sea anemone Heteractis crispa modulate ASIC1a channels. Peptides 2018, 104, 41–49. [Google Scholar] [CrossRef]
- Kozlov, S.; Grishin, E. Convenient nomenclature of cysteine-rich polypeptide toxins from sea anemones. Peptides 2012, 33, 240–244. [Google Scholar] [CrossRef]
- Kvetkina, A.; Kostina, E.; Gladkikh, I.; Chausova, V.; Yurchenko, E.; Bakunina, I.; Pivkin, M.; Anastyuk, S.; Popov, R.; Monastyrnaya, M. Deep-Sea Anemones Are Prospective Source of New Antimicrobial and Cytotoxic Compounds. Mar. Drugs 2021, 19, 654. [Google Scholar] [CrossRef]
- Peigneur, S.; Beress, L.; Moller, C.; Mari, F.; Forssmann, W.G.; Tytgat, J. A natural point mutation changes both target selectivity and mechanism of action of sea anemone toxins. FASEB J. 2012, 26, 5141–5151. [Google Scholar] [CrossRef]
- Diochot, S.; Loret, E.; Bruhn, T.; Beress, L.; Lazdunski, M. APETx1, a new toxin from the sea anemone Anthopleura elegantissima, blocks voltage-gated human ether-a-go-go-related gene potassium channels. Mol. Pharmacol. 2003, 64, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.E.; Cristofori-Armstrong, B.; Anangi, R.; Rosengren, K.J.; Lau, C.H.; Mobli, M.; Brust, A.; Alewood, P.F.; King, G.F.; Rash, L.D. Understanding the molecular basis of toxin promiscuity: The analgesic sea anemone peptide APETx2 interacts with acid-sensing ion channel 3 and hERG channels via overlapping pharmacophores. J. Med. Chem. 2014, 57, 9195–9203. [Google Scholar] [CrossRef] [PubMed]
- Moreels, L.; Peigneur, S.; Galan, D.T.; De Pauw, E.; Beress, L.; Waelkens, E.; Pardo, L.A.; Quinton, L.; Tytgat, J. APETx4, a Novel Sea Anemone Toxin and a Modulator of the Cancer-Relevant Potassium Channel KV10.1. Mar. Drugs 2017, 15, 287. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K.; Shimomura, T.; Kubo, Y.; Oka, T.; Kobayashi, N.; Imai, S.; Yanase, N.; Akimoto, M.; Fukuda, M.; Yokogawa, M.; et al. Mechanism of hERG inhibition by gating-modifier toxin, APETx1, deduced by functional characterization. BMC Mol. Cell Biol. 2021, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Torres, I.O.; Jin, T.B.; Cadene, M.; Chait, B.T.; Poget, S.F. Discovery and characterisation of a novel toxin from Dendroaspis angusticeps, named Tx7335, that activates the potassium channel KcsA. Sci. Rep. 2016, 6, 23904. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.S.; Syeda, R.; Montal, M. Stabilization of the conductive conformation of a voltage-gated K+ (Kv) channel: The lid mechanism. J. Biol. Chem. 2013, 288, 16619–16628. [Google Scholar] [CrossRef]
- Lange, A.; Giller, K.; Hornig, S.; Martin-Eauclaire, M.F.; Pongs, O.; Becker, S.; Baldus, M. Toxin-induced conformational changes in a potassium channel revealed by solid-state NMR. Nature 2006, 440, 959–962. [Google Scholar] [CrossRef]
- Peroz, D.; Rodriguez, N.; Choveau, F.; Baro, I.; Merot, J.; Loussouarn, G. Kv7.1 (KCNQ1) properties and channelopathies. J. Physiol. 2008, 586, 1785–1789. [Google Scholar] [CrossRef]
- Ferrer, T.; Arechiga-Figueroa, I.A.; Shapiro, M.S.; Tristani-Firouzi, M.; Sanchez-Chapula, J.A. Tamoxifen inhibition of kv7.2/kv7.3 channels. PLoS ONE 2013, 8, e76085. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Zeng, D.Y.; Hu, Y.T.; He, Y.W.; Pan, N.; Ding, J.P.; Cao, Z.J.; Liu, M.L.; Li, W.X.; Yi, H.; et al. Structural and functional diversity of acidic scorpion potassium channel toxins. PLoS ONE 2012, 7, e35154. [Google Scholar] [CrossRef] [PubMed]
- Landoulsi, Z.; Miceli, F.; Palmese, A.; Amoresano, A.; Marino, G.; El Ayeb, M.; Taglialatela, M.; Benkhalifa, R. Subtype-selective activation of K(v)7 channels by AaTXKAaTXKβ(2–64), a novel toxin variant from the Androctonus australis scorpion venom. Mol. Pharmacol. 2013, 84, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.Y.; Thompson, D.; Wang, Z.; Fedida, D.; Robertson, B. Modulation of Kv3 subfamily potassium currents by the sea anemone toxin BDS: Significance for CNS and biophysical studies. J. Neurosci. 2005, 25, 8735–8745. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Jo, S.; Bean, B.P. Modulation of neuronal sodium channels by the sea anemone peptide BDS-I. J. Neurophysiol. 2012, 107, 3155–3167. [Google Scholar] [CrossRef] [PubMed]
- Napp, J.; Monje, F.; Stuhmer, W.; Pardo, L.A. Glycosylation of Eag1 (Kv10.1) potassium channels: Intracellular trafficking and functional consequences. J. Biol. Chem. 2005, 280, 29506–29512. [Google Scholar] [CrossRef]
- Whicher, J.R.; MacKinnon, R. Structure of the voltage-gated K(+) channel Eag1 reveals an alternative voltage sensing mechanism. Science 2016, 353, 664–669. [Google Scholar] [CrossRef]
- Boldrini-Franca, J.; Pinheiro-Junior, E.L.; Peigneur, S.; Pucca, M.B.; Cerni, F.A.; Borges, R.J.; Costa, T.R.; Carone, S.E.I.; Fontes, M.R.M.; Sampaio, S.V.; et al. Beyond hemostasis: A snake venom serine protease with potassium channel blocking and potential antitumor activities. Sci. Rep. 2020, 10, 4476. [Google Scholar] [CrossRef]
- Pinheiro-Junior, E.L.; Boldrini-França, J.; Takeda, A.A.S.; Costa, T.R.; Peigneur, S.; Cardoso, I.A.; de Oliveira, I.S.; Sampaio, S.V.; de Mattos Fontes, M.R.; Tytgat, J.; et al. Towards toxin PEGylation: The example of rCollinein-1, a snake venom thrombin-like enzyme, as a PEGylated biopharmaceutical prototype. Int. J. Biol. Macromol. 2021, 190, 564–573. [Google Scholar] [CrossRef]
- Koren, G.; Liman, E.R.; Logothetis, D.E.; Nadal-Ginard, B.; Hess, P. Gating mechanism of a cloned potassium channel expressed in frog oocytes and mammalian cells. Neuron 1990, 4, 39–51. [Google Scholar] [CrossRef]







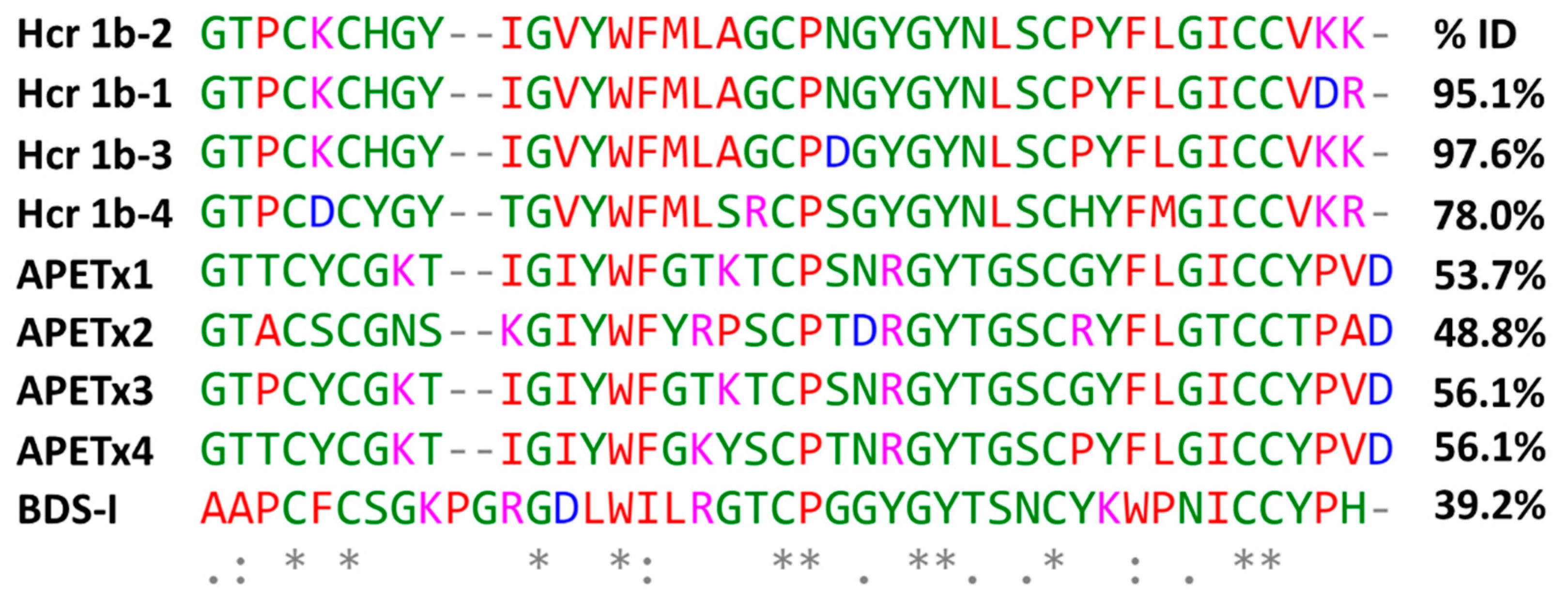{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vhalf Activation (mV ± SEM) | |||
|---|---|---|---|
| Channel Isoform | Control (ND96) | Hcr 1b-2 (1 µM) | Shift (mV) |
| Kv1.1 | −13.8 ± 0.3 | −18.6 ± 0.3 | −4.8 |
| Kv1.2 | −24.7 ± 0.7 | −29.6 ± 0.89 | −4.9 |
| Kv1.4 | 7.5 ± 0.7 | 10.0 ± 0.5 | 2.5 |
| Kv2.1 | 5.3 ± 0.3 | 8.0 ± 0.2 | 2.6 |
| Kv7.1 | 5.8 ± 0.4 | 12.2 ± 0.6 | 6.4 |
| Kv7.2/7.3 | 14.6 ± 0.5 | 15.9 ± 0.7 | 1.3 |
| Kv7.4 | 3.8 ± 0.3 | 10.9 ± 0.3 | 7.1 |
| KQT1 | 14.6 ± 0.4 | 22.4 ± 0.5 | 7.9 |
| hERG (peak) | −13.7 ± 0.5 | 12.6 ± 0.4 | 25.7 |
| hERG (tail) | −22.9 ± 0.6 | 0.6 ± 0.7 | 24.8 |
| Vhalf Activation (mV ± SEM) | Vhalf Inactivation (mV ± SEM) | |||||
|---|---|---|---|---|---|---|
| Channel Isoform | Control (ND96) | Hcr 1b-2 (1 µM) | Shift (mV) | Control (ND96) | Hcr 1b-2 (1 µM) | Shift (mV) |
| Nav1.1 | −24.6 ± 0.1 | −24.5 ± 0.1 | 0.1 | −37.2 ± 0.2 | −43 ± 0.4 | −5.8 |
| Nav1.2 | −19.2 ± 0.1 | −15.5 ± 0.2 | 3.7 | −33.6 ± 0.4 | −34.7 ± 0.6 | −1.1 |
| Nav1.3 | −21.2 ± 0.1 | −15.9 ± 0.1 | 5.3 | −29.9 ± 0.1 | −33.3 ± 0.1 | −3.4 |
| Nav1.4 | −24.4 ± 0.1 | −18.9 ± 0.1 | 5.5 | −48.5 ± 0.3 | −50.2 ± 0.2 | −1.7 |
| Nav1.5 | −38.5 ± 0.1 | −34.7 ± 0.1 | 3.8 | −66.0 ± 0.2 | −69.2 ± 0.2 | −3.2 |
| Nav1.6 | −23.5 ± 0.1 | −20.0 ± 0.1 | 3.5 | −49.1 ± 0.3 | −56.7 ± 0.4 | −7.6 |
| Nav1.7 | −26.9 ± 0.1 | −29.7 ± 0.1 | −2.8 | −38.1 ± 0.3 | −38.6 ± 0.4 | −0.5 |
| Nav1.8 | −2.3 ± 0.1 | −2.6 ± 0.1 | −0.3 | −35.6 ± 0.4 | −36.9 ± 0.5 | −1.3 |
| BgNav | −29.0 ± 0.1 | −27.1 ± 0.1 | 1.9 | −47.9 ± 0.3 | −52.9 ± 0.3 | −5.0 |
| Vhalf Activation (mV) | Vhalf Inactivation (mV) | |||||
|---|---|---|---|---|---|---|
| Channel Isoform | Control (ND96 Ca2+ Free) | Hcr 1b-2 (1 µM) | Shift | Control (ND96 Ca2+ Free) | Hcr 1b-2 (1 µM) | Shift |
| Cav3.1 | −41.0 ± 0.1 | −41.2 ± 0.1 | −0.2 | −58.4 ± 0.2 | −60.7 ± 0.2 | −2.3 |
| Cav3.2 | −31.9 ± 0.1 | −32.3 ± 0.2 | −0.4 | −46.6 ± 0.8 | −46.1 ± 0.6 | 0.5 |
| Cav3.3 | −41.7 ± 0.4 | −43.9 ± 0.5 | −2.2 | −47.6 ± 0.2 | −49.7 ± 0.3 | −2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro-Junior, E.L.; Kalina, R.; Gladkikh, I.; Leychenko, E.; Tytgat, J.; Peigneur, S. A Tale of Toxin Promiscuity: The Versatile Pharmacological Effects of Hcr 1b-2 Sea Anemone Peptide on Voltage-Gated Ion Channels. Mar. Drugs 2022, 20, 147. https://doi.org/10.3390/md20020147
Pinheiro-Junior EL, Kalina R, Gladkikh I, Leychenko E, Tytgat J, Peigneur S. A Tale of Toxin Promiscuity: The Versatile Pharmacological Effects of Hcr 1b-2 Sea Anemone Peptide on Voltage-Gated Ion Channels. Marine Drugs. 2022; 20(2):147. https://doi.org/10.3390/md20020147
Chicago/Turabian StylePinheiro-Junior, Ernesto Lopes, Rimma Kalina, Irina Gladkikh, Elena Leychenko, Jan Tytgat, and Steve Peigneur. 2022. "A Tale of Toxin Promiscuity: The Versatile Pharmacological Effects of Hcr 1b-2 Sea Anemone Peptide on Voltage-Gated Ion Channels" Marine Drugs 20, no. 2: 147. https://doi.org/10.3390/md20020147
APA StylePinheiro-Junior, E. L., Kalina, R., Gladkikh, I., Leychenko, E., Tytgat, J., & Peigneur, S. (2022). A Tale of Toxin Promiscuity: The Versatile Pharmacological Effects of Hcr 1b-2 Sea Anemone Peptide on Voltage-Gated Ion Channels. Marine Drugs, 20(2), 147. https://doi.org/10.3390/md20020147