
Biomedical Potential of the Neglected Molluscivorous and Vermivorous Conus Species



Abstract
:1. Introduction
2. Results and Discussion
2.1. Feeding Habits and Evolution Path
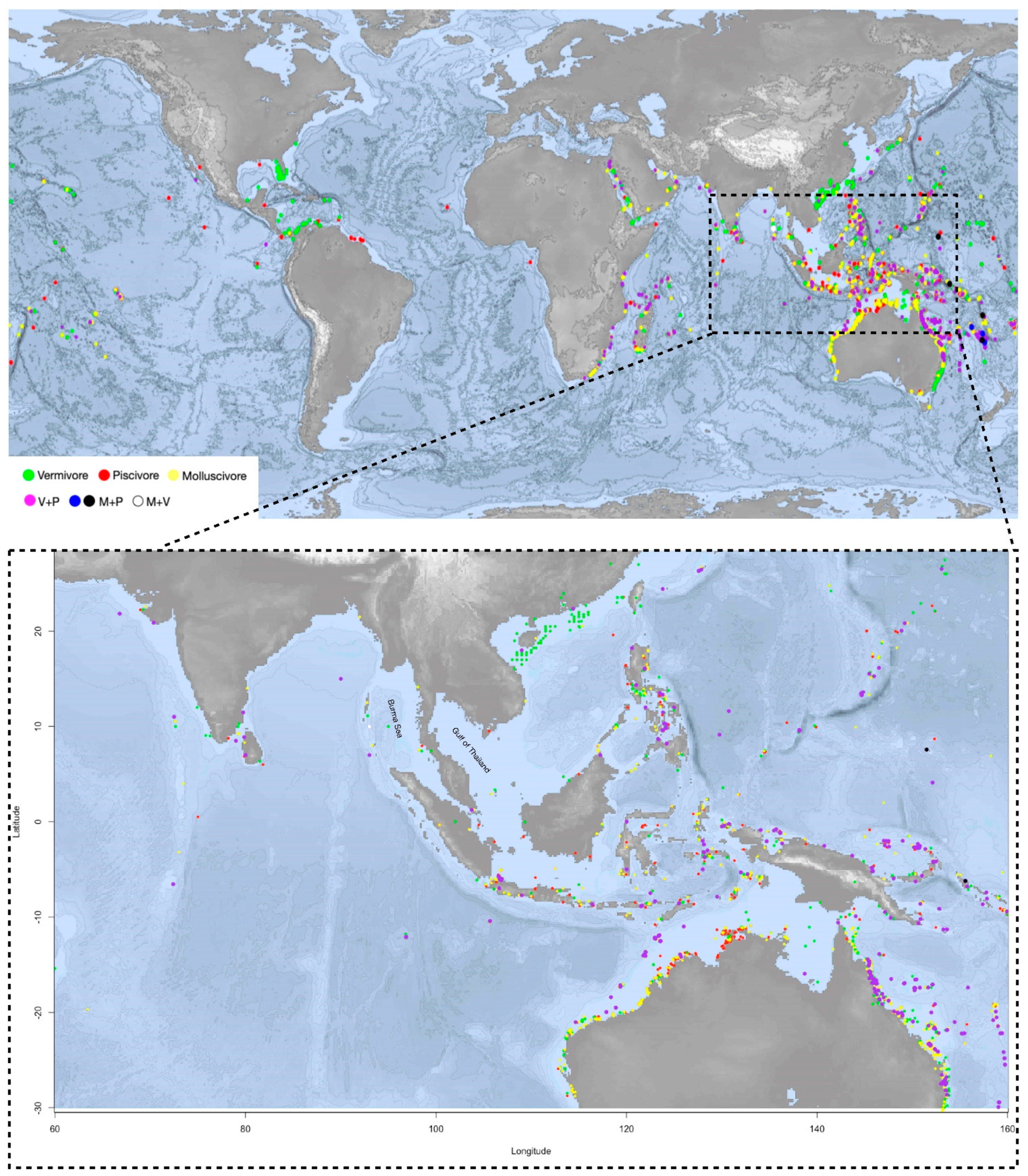
2.2. Distribution
2.3. Conservation Status
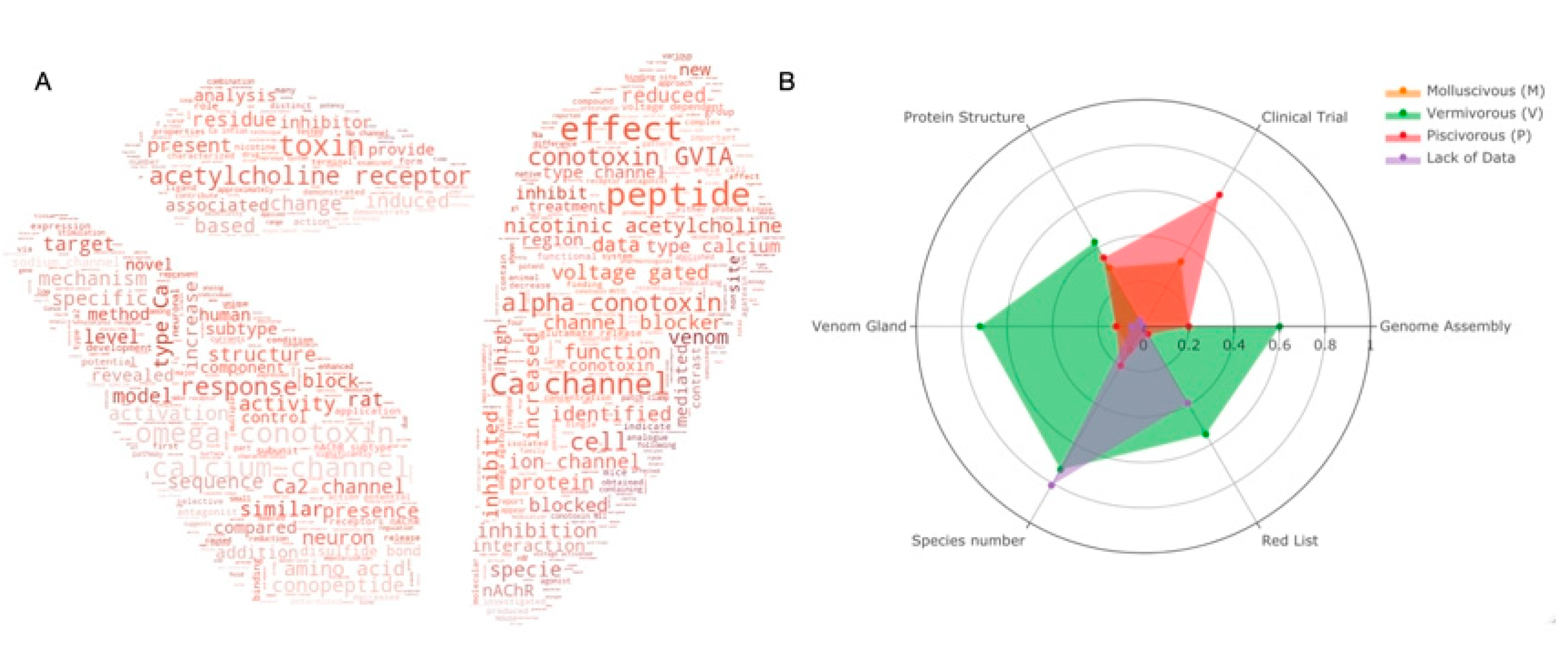
2.4. Drug Discovery
2.5. Structural and Functional Studies of Conotoxins with Their Receptors
2.6. Potential Value in COVID-19 and Other Diseases
3. Materials and Methods
3.1. Species and Feeding Habits
3.2. Geographic Coordinates Information
3.3. Sequence Alignment and Phylogenetic Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duda, T.F.; Kohn, A.J.; Palumbi, S.R. Origins of diverse feeding ecologies within Conus, a genus of venomous marine gastropods. Biol. J. Linn. Soc. 2001, 73, 391–409. [Google Scholar] [CrossRef]
- Puillandre, N.; Duda, T.F.; Meyer, C.P.; Olivera, B.M.; Bouchet, P. One, four or 100 genera? A new classification of the cone snails. J. Molluscan Stud. 2015, 81, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Peng, C.; Yang, J.; Yi, Y.; Zhang, J.; Shi, Q. Cone snails: A big store of conotoxins for novel drug discovery. Toxins 2017, 9, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, G.; Rochelle, L. Panorama sur La Diversite des Conidae 110 Espèces Prédatrices des Plus Efficaces. 2021. Available online: https://www.researchgate.net/publication/353337774_PANORAMA_SUR_LA_DIVERSITE_DES_CONIDAE_110_especes_predatrices_des_plus_efficaces (accessed on 7 January 2022).
- Franklin, J.B.; Apte, D.A. Two New Records of Conidae (Mollusca: Caenogastropoda) from the Andaman Islands, India. J. Bombay Nat. Hist. Soc. 2020, 117, 152546. [Google Scholar]
- Franklin, J.B.; Fernando, A.A.; Chalke, B.A.; Krishnan, K.S. Radular morphology of Conus (Gastropoda: Caenogastropoda: Conidae) from India. Molluscan Res. 2007, 27, 111–122. [Google Scholar]
- Sanpanich, K.; Duangdee, T. A survey of marine mollusc diversity in the Southern Mergui Archipelago, Myanmar. Phuket Mar. Biol. Cent. Res. Bull. 2018, 75, 45–60. [Google Scholar]
- Lluisma, A.O.; Milash, B.A.; Moore, B.; Olivera, B.M.; Bandyopadhyay, P.K. Novel venom peptides from the cone snail Conus pulicarius discovered through next-generation sequencing of its venom duct transcriptome. Mar Genomics. Mar. Genom. 2012, 5, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Olivera, B.M. Conus venom peptides: Reflections from the biology of clades and species. Annu. Rev. Ecol. Syst. 2002, 33, 25–47. [Google Scholar] [CrossRef]
- Jiménez-Tenorio, M. Cone radular anatomy as a proxy for phylogeny and for conotoxin diversity. In Proceedings of the CONODAYS—Satellite event to Natural Peptide to Drugs International, Zermatt, Switzerland, 7–9 December 2011. [Google Scholar] [CrossRef]
- Leal, J.H.; Kohn, A.J.; Mensch, R. A Veliconcha Unveiled: Observations on the Larva and Radula of Conus spurius, with Implications for the Origin of Molluscivory in Conus. Am. Malacol. Bull. 2017, 35, 111–118. [Google Scholar] [CrossRef]
- Dutertre, S.; Lewis, R.J. Cone Snail Biology, Bioprospecting and Conservation. In Snails: Biology, Ecology and Conservation; Nova Science Publishers: New York, NY, USA, 2013. [Google Scholar]
- Buczek, O.; Bulaj, G.; Olivera, B.M. Conotoxins and the posttranslational modification of secreted gene products. Cell. Mol. Life Sci. 2005, 62, 3067–3079. [Google Scholar] [CrossRef]
- Neves, J.L.B.; Lin, Z.; Imperial, J.S.; Antunes, A.; Vasconcelos, V.; Olivera, B.M.; Schmidt, E.W. Small Molecules in the Cone Snail Arsenal. Org. Lett. 2015, 17, 4933–4935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, J.R.; Jan, I.; Sangha, G.; Azizi, E. The high speed radular prey strike of a fish-hunting cone snail. Curr. Biol. 2019, 29, R788–R789. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.; Gilly, W.F. Piscivorous behavior of a temperate cone snail, Conus californicus. Biol. Bull. 2005, 209, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation-and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abalde, S.; Jimenez-Tenorio, M.; Afonso, C.M.L.; Zardoya, R. Conotoxin diversity in Chelyconus ermineus (Born, 1778) and the convergent origin of piscivory in the atlantic and indo-pacific cones. Genome Biol. Evol. 2018, 10, 2643–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prator, C.; Murayama, K.M.; Schulz, J.R. Venom variation during prey capture by the cone snail, Conus textile. PLoS ONE 2014, 9, e98991. [Google Scholar] [CrossRef] [Green Version]
- Duda, T.F.; Chang, D.; Lewis, B.D.; Lee, T. Geographic variation in venom allelic composition and diets of the widespread predatory marine gastropod Conus ebraeus. PLoS ONE 2009, 4, e6245. [Google Scholar] [CrossRef]
- Neves, J.; Campos, A.; Osório, H.; Antunes, A.; Vasconcelos, V. Conopeptides from Cape Verde Conus crotchii. Mar. Drugs 2013, 11, 2203–2215. [Google Scholar] [CrossRef] [Green Version]
- Neves, J.L.B.; Imperial, J.S.; Morgenstern, D.; Ueberheide, B.; Gajewiak, J.; Antunes, A.; Robinson, S.D.; Espino, S.; Watkins, M.; Vasconcelos, V.; et al. Characterization of the First Conotoxin from Conus ateralbus, a Vermivorous Cone Snail from the Cabo Verde Archipelago. Mar. Drugs 2019, 17, 432. [Google Scholar] [CrossRef] [Green Version]
- Mir, R.; Karim, S.; Kamal, M.A.; Wilson, C.; Mirza, Z. Conotoxins: Structure, Therapeutic Potential and Pharmacological Applications. Curr. Pharm. Des. 2016, 22, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.D.; Han, Y.H.; Wang, C.G.; Chi, C.W. From the identification of gene organization of α conotoxins to the cloning of novel toxins. Toxicon 2007, 49, 1135–1149. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Westermann, J.C.; Craik, D.J. Conopeptide characterization and classifications: An analysis using ConoServer. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.; Kaas, Q.; Craik, D.J.; Lewis, R.; Alewood, P.F. Conotoxins: Chemistry and Biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef] [PubMed]
- Ekberg, J.; Craik, D.J.; Adams, D.J. Conotoxin modulation of voltage-gated sodium channels. Int. J. Biochem. Cell Biol. 2008, 40, 2363–2368. [Google Scholar] [CrossRef]
- Kaas, Q.; Yu, R.; Jin, A.-H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2011, 40, D325–D330. [Google Scholar] [CrossRef]
- Schroeder, C.I.; Craik, D.J. Therapeutic potential of conopeptides. Future Med. Chem. 2012, 4, 1243–1255. [Google Scholar] [CrossRef]
- Miljanich, G. Ziconotide: Neuronal Calcium Channel Blocker for Treating Severe Chronic Pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef]
- Herrick, C.J. The cranial and first spinal nerves of menidia: A contribution upon the nerve components of the bony fishes. Section 13. Conclusions. J. Comp. Neurol. 1899, 9, 419–455. [Google Scholar] [CrossRef]
- Olivera, B.M. Conus peptides: Biodiversity-based discovery and exogenomics. J. Biol. Chem. 2006, 281, 31173–31177. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.-H.; Kaas, Q.; Jones, A.; Alewood, P.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [Green Version]
- Duda, T.F.; Palumbi, S.R. Evolutionary diversification of multigene families: Allelic selection of toxins in predatory cone snails. Mol. Biol. Evol. 2000, 17, 1286–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Fu, Y.; Wang, L.; Liang, A.; Chen, S.; Xu, A. Identifying novel conopepetides from the venom ducts of Conus litteratus through integrating transcriptomics and proteomics. J. Proteom. 2019, 192, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Peng, C.; Zhu, Y.; Fan, C.; Jiang, H.; Chen, J.; Cao, Y.; Shi, Q. High-throughput identification and analysis of novel conotoxins from three vermivorous cone snails by transcriptome sequencing. Mar. Drugs 2019, 17, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Watkins, M.; Robinson, S.D.; Safavi-Hemami, H.; Yandell, M. Discovery of novel conotoxin candidates using machine learning. Toxins 2018, 10, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Characterization of the Conus bullatus genome and its venom-duct transcriptome. BMC Genom. 2011, 12, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreson, R.; Roosaare, M.; Kaplinski, L.; Laht, S.; Kõressaar, T.; Lepamets, M.; Brauer, A.; Kukuškina, V.; Remm, M. Gene content of the fish-hunting cone snail Conus consors. bioRxiv 2019, bioRxiv:590695. [Google Scholar] [CrossRef]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. Structural features of conopeptide genes inferred from partial sequences of the Conus tribblei genome. Mol. Genet. Genom. 2015, 291, 411–422. [Google Scholar] [CrossRef]
- Hinegardner, R. Cellular DNA content of the Mollusca. Comp. Biochem. Physiol. Part A Physiol. 1974, 47, 447–460. [Google Scholar] [CrossRef]
- Peng, C.; Huang, Y.; Bian, C.; Li, J.; Liu, J.; Zhang, K.; You, X.; Lin, Z.; He, Y.; Chen, J.; et al. The first Conus genome assembly reveals a primary genetic central dogma of conopeptides in C. betulinus. Cell Discov. 2021, 7, 11. [Google Scholar] [CrossRef]
- Pardos-Blas, J.R.; Irisarri, I.; Abalde, S.; Afonso, C.M.L.; Tenorio, M.J.; Zardoya, R. The genome of the venomous snail Lautoconus ventricosus sheds light on the origin of conotoxin diversity. Gigascience 2021, 10, giab037. [Google Scholar] [CrossRef]
- Thiriot-Quiévreux, C. Advances in chromosomal studies of gastropod molluscs. J. Molluscan Stud. 2003, 69, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Dalet, J.T.; Saloma, C.P.; Olivera, B.M.; Heralde, F.M. Karyological analysis and FISH physical mapping of 18S rDNA genes, (GATA)n centromeric and (TTAGGG)n telomeric sequences in Conus magus Linnaeus, 1758. J. Molluscan Stud. 2014, 81, 274–289. [Google Scholar] [CrossRef] [Green Version]
- Ebied, A.M.; Hassan, H.A.; Abu-Almaaty, A.H.; Yasen, A.E. Cytogenetic studies on metaphase chromosomes of eight gastropod species of orders Mesogastropoda and Neogastropoda from the Red Sea (Prosobranchia-Mollusca). J. Egypt. Ger. Soc. Zool. 2000, 33, 317–336. [Google Scholar]
- Vitturi, R.; Catalano, E. Spermatocyte chromosomes in 7 species of the sub-class Prosobranchia (Mollusca, Gasteropoda). Biol. Zentralbl. 1984, 103, 69–76. [Google Scholar]
- Abalde, S.; Tenorio, M.J.; Uribe, J.E.; Zardoya, R. Conidae phylogenomics and evolution. Zool. Scr. 2019, 48, 194–214. [Google Scholar] [CrossRef]
- Puillandre, N.; Bouchet, P.; Duda, T.; Kauferstein, S.; Kohn, A.; Olivera, B.; Watkins, M.; Meyer, C. Molecular phylogeny and evolution of the cone snails (Gastropoda, Conoidea). Mol. Phylogenet. Evol. 2014, 78, 290–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, P.J. Patterns of speciation in marine gastropods: A review of the phylogenetic evidence for localized radiations in the sea. Am. Malacol. Bull. 2011, 29, 169–186. [Google Scholar] [CrossRef]
- Uribe, J.E.; Puillandre, N.; Zardoya, R. Beyond Conus: Phylogenetic relationships of Conidae based on complete mitochondrial genomes. Mol. Phylogenet. Evol. 2017, 107, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Betancur, R.R.; Wiley, E.O.; Arratia, G.; Acero, A.; Bailly, N.; Miya, M.; Lecointre, G.; Ortí, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17, 162. [Google Scholar] [CrossRef] [Green Version]
- Kocot, K.M.; Cannon, J.; Todt, C.; Citarella, M.R.; Kohn, A.B.; Meyer, A.; Santos, S.R.; Schander, C.; Moroz, L.; Lieb, B.; et al. Phylogenomics reveals deep molluscan relationships. Nature 2011, 477, 452–456. [Google Scholar] [CrossRef] [Green Version]
- Peters, H.; O’Leary, B.C.; Hawkins, J.P.; Carpenter, K.E.; Roberts, C.M. Conus: First comprehensive conservation red list assessment of a marine gastropod mollusc genus. PLoS ONE 2013, 8, e83353. [Google Scholar] [CrossRef] [PubMed]
- Peters, H.; O’Leary, B.C.; Hawkins, J.P.; Roberts, C.M. The cone snails of Cape Verde: Marine endemism at a terrestrial scale. Glob. Ecol. Conserv. 2016, 7, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F. Have marine natural product drug discovery efforts been productive and how can we improve their efficiency? Expert Opin. Drug Discov. 2019, 14, 717–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, D.C.I.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. 2006, 63, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.C.W.; Teoh, H. Benefits of Modest Weight Loss on the Management of Type 2 Diabetes Mellitus. Can. J. Diabetes 2013, 37, 128–134. [Google Scholar] [CrossRef]
- Wehbe, R.; Frangieh, J.; Rima, M.; El Obeid, D.; Sabatier, J.-M.; Fajloun, Z. Bee venom: Overview of main compounds and bioactivities for therapeutic interests. Molecules 2019, 24, 2997. [Google Scholar] [CrossRef] [Green Version]
- Sunagar, K.; Fry, B.G.; Jackson, T.N.W.; Casewell, N.R.; Undheim, E.A.B.; Vidal, N.; Ali, S.A.; King, G.F.; Vasudevan, K.; Vasconcelos, V.; et al. Molecular evolution of vertebrate neurotrophins: Co-option of the highly conserved nerve growth factor gene into the advanced snake venom arsenal. PLoS ONE 2013, 8, e81827. [Google Scholar] [CrossRef]
- Fry, B.G.; Undheim, E.; Ali, S.A.; Jackson, T.; Debono, J.; Scheib, H.; Ruder, T.; Morgenstern, D.; Cadwallader, L.; Whitehead, D.; et al. Squeezers and leaf-cutters: Differential diversification and degeneration of the venom system in toxicoferan reptiles. Mol. Cell. Proteom. 2013, 12, 1881–1899. [Google Scholar] [CrossRef] [Green Version]
- Pineda, S.S.; Sollod, B.L.; Wilson, D.; Darling, A.; Sunagar, K.; Undheim, E.A.B.; Kely, L.; Antunes, A.; Fry, B.G.; King, G.F. Diversification of a single ancestral gene into a successful toxin superfamily in highly venomous Australian funnel-web spiders. BMC Genom. 2014, 15, 177. [Google Scholar] [CrossRef] [Green Version]
- Ali, M. Studies on Bee Venom and Its Medical Uses. Int. J. Adv. Res. Technol. 2012, 1, 69–83. [Google Scholar]
- Han, T.S.; Teichert, R.W.; Olivera, B.M.; Bulaj, G. Conus Venoms—A Rich Source of Peptide-Based Therapeutics. Curr. Pharm. Des. 2008, 14, 2462–2479. [Google Scholar] [CrossRef] [PubMed]
- Alonso, D.; Khalil, Z.; Satkunanthan, N.; Livett, B. Drugs From the Sea: Conotoxins as Drug Leads for Neuropathic Pain and Other Neurological Conditions. Mini Rev. Med. Chem. 2003, 3, 785–787. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Yu, J.; Zhu, X.; Zhangsun, D.; Luo, S. Characterization of an α 4/7-Conotoxin LvIF from Conus lividus That Selectively Blocks α3β2 Nicotinic Acetylcholine Receptor. Mar. Drugs 2021, 19, 398. [Google Scholar] [CrossRef]
- Vincler, M.; Wittenauer, S.; Parker, R.; Ellison, M.; Olivera, B.M.; McIntosh, J.M. Molecular mechanism for analgesia involving specific antagonism of α9α10 nicotinic acetylcholine receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 17880–17884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, J.; Harrington, A.M.; Garcia-Caraballo, S.; Maddern, J.; Grundy, L.; Zhang, J.; Page, G.; Miller, P.E.; Craik, D.J.; Adams, D.J.; et al. α-Conotoxin Vc1.1 inhibits human dorsal root ganglion neuroexcitability and mouse colonic nociception via GABA B receptors. Gut 2016, 66, 1083–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2018, 679, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Durek, T.; Craik, D.J. Therapeutic conotoxins: A US patent literature survey. Expert Opin. Ther. Patients 2015, 25, 1159–1173. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Zhangsun, D.; Yu, G.; Su, R.; Luo, S. The α9α10 nicotinic acetylcholine receptor antagonist αo-conotoxin GeXIVA[1,2] alleviates and reverses chemotherapy-induced neuropathic pain. Mar. Drugs 2019, 17, 265. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Pan, S.; Xu, M.; Zhang, L.; Yu, J.; Yu, J.; Wu, Y.; Fan, Y.; Li, H.; Kasheverov, I.E.; et al. High Selectivity of an α-Conotoxin LvIA Analogue for α3β2 Nicotinic Acetylcholine Receptors Is Mediated by β2 Functionally Important Residues. J. Med. Chem. 2020, 63, 13656–13668. [Google Scholar] [CrossRef]
- Dyachenko, I.A.; Palikova, Y.A.; Palikov, V.A.; Korolkova, Y.V.; Kazakov, V.A.; Egorova, N.S.; Garifulina, A.I.; Utkin, Y.N.; Tsetlin, V.I.; Kryukova, E.V. α-Conotoxin RgIA and oligoarginine R8 in the mice model alleviate long-term oxaliplatin induced neuropathy. Biochimie 2021, in press. [Google Scholar] [CrossRef]
- Kryukova, E.V.; Ivanov, I.A.; Lebedev, D.S.; Spirova, E.N.; Egorova, N.S.; Zouridakis, M.; Kasheverov, I.E.; Tzartos, S.J.; Tsetlin, V.I. Orthosteric and/or allosteric binding of a-conotoxins to nicotinic acetylcholine receptors and their models. Mar. Drugs 2018, 16, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, A.; Dovell, S.; Chesnokov, O.; Hoggard, M.; Oleinikov, A.V.; Marí, F. Conus venom fractions inhibit the adhesion of Plasmodium falciparum erythrocyte membrane protein 1 domains to the host vascular receptors. J. Proteom. 2021, 234, 104083. [Google Scholar] [CrossRef] [PubMed]
- Brogna, C.; Cristoni, S.; Petrillo, M.; Querci, M.; Piazza, O.; Van den Eede, G. Toxin-like peptides in plasma, urine and faecal samples from COVID-19 patients. F1000Research 2021, 10, 550. [Google Scholar] [CrossRef]
- Oliveira, A.S.F.; Ibarra, A.A.; Bermudez, I.; Casalino, L.; Gaieb, Z.; Shoemark, D.K.; Gallagher, T.; Sessions, R.B.; Amaro, R.E.; Mulholland, A.J.; et al. A potential interaction between the SARS-CoV-2 spike protein and nicotinic acetylcholine receptors. Biophys. J. 2021, 120, 983–993. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Tenorio, M.J.; Abalde, S.; Pardos-Blas, J.R.; Zardoya, R. Taxonomic revision of West African cone snails (Gastropoda: Conidae) based upon mitogenomic studies: Implications for conservation. Eur. J. Taxon. 2020, 2020, 663. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Conotoxin Family | Definition | Gene Superfamily | Cysteine Framework |
|---|---|---|---|
| α (ALPHA) | Nicotinic acetylcholine receptor (nAChR) | A, B3, D, J, L, M, S | I, II, III, IV, VIII, XIV, XX, XXV |
| γ (GAMMA) | Neuronal pacemaker cation currents (inward cation current) | O1, O2 | VI/VII |
| δ (DELTA) | Voltage-gated Na Channel (agonist, delay inactivation) | O1 | VI/VII |
| ε (EPISILON) | Presynaptic calcium channels or G protein-coupled presynaptic receptor | T | V |
| I (IOTA) | Voltage-gated Na Channel (agonist, no delayed inactivation) | I1, M | III, XI |
| κ (KAPPA) | Voltage-gated K Channel (blocker) | A, I2, J, M, O1 | III, IV, VI/VII, XI, XIV |
| μ (MU) | Voltage-gated Na Channel (antagonist, blocker) | M, O1,T | III, IV, V, VI, VII |
| ρ (RHO) | Alpha 1 adrenoreceptor (GPCR) | A | I |
| σ (SIGMA) | Serotonin-gated ion channels (GPCR) | S | VIII |
| τ (TAU) | Somatostatin receptor | T | V |
| χ (CHI) | Neuronal noradrenaline transporter | T | X |
| ω (OMEGA) | Voltage-gated calcium channel | O1, O2 | VI/VII, XVI, XXVI |
| Conopeptide | Commercial Name | Comment | Target | Stage | Company | Conus Species (**) | Reference | |
|---|---|---|---|---|---|---|---|---|
| 1 | α-Vc1.1 | ACV1 | Neuropathic pain | nAChR (α9α10) | Phase II * | Metabolic Pharmaceuticals, Melbourne, Australia | Conus victoriae (m) | [3] |
| 2 | ω-CVID | AM336 | Neuropathic pain | Ca2+ channel (CaV2.2) N-type calcium channels/blocker | Phase IIa * | Relevare Pharmaceuticals LTD., Australia | Conus catus (p) | [3] |
| 3 | μO-MrVIB | CGX-1002 | Neuropathic pain | Sodium channels/subtype selective blocker | Preclinical * | Cognetix Inc, Salt Lake City, USA | Conus tulipa (p) | [3] |
| 4 | Conantokin-G | CGX-1007 | Intractable epilepsy / pain | NMDA receptor (NR2B) | Preclinical * | Cognetix Inc, Salt Lake City, USA | Conus geographus (p) | [3] |
| 5 | Contulakin-G | CGX-1160 | Neuropathic pain | Neurotensin receptor | Phase Ib * | Cognetix Inc, Salt Lake City, USA | Conus geographus (p) | [3] |
| 6 | ω-MVIIA | SNX-III, C1002, Ziconotide, Prialt | Intractable pain | Ca2+ channel (CaV2.2) N-type calcium channels/blocker | FDA-approved | Elan Corporation (Elan Pharmaceuticals), CA, USA | Conus magus (p) | [3] |
| 7 | χ-MrIA | Xen2174 | Neuropathic pain | Norepinephrine transporter/inhibitor | Phase IIa * | Xenome, Ltd., Brisbane, Qld., Australia | n.a. | [3] |
| 8 | κ-PVIIA | CGX-1051 | Acute Myocardial Infarct, Cardioprotection | K+ channel (KV1)/blocker | Preclinical | n.a. | n.a. | [64] |
| 9 | n.a. | CGX-1204 | Muscle relaxer / pain | Nicotinic acetylcholine receptors/antagonist | Preclinical | n.a. | n.a. | [64] |
| 10 | μ-SIIIA | PEG-SIIIA | Inflammatory pain | Sodium channels/blocker | Preclinical | n.a. | n.a. | [64] |
| 11 | ρ-Conotoxin TIA | n.a. | n.a. | α-1 adrenergic receptors | Preclinical | Xenome, Ltd., Brisbane, Qld., Australia | Conus tulipa (p) | [65] |
| 12 | χ-conopeptides (χ-CTX MrIA/B) | n.a. | Neuropathic pain | Neurotransmitter transporters | Preclinical | Xenome, Ltd., Brisbane, Qld., Australia | Conus marmoreus (m) | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Antunes, A. Biomedical Potential of the Neglected Molluscivorous and Vermivorous Conus Species. Mar. Drugs 2022, 20, 105. https://doi.org/10.3390/md20020105
Zhao Y, Antunes A. Biomedical Potential of the Neglected Molluscivorous and Vermivorous Conus Species. Marine Drugs. 2022; 20(2):105. https://doi.org/10.3390/md20020105
Chicago/Turabian StyleZhao, Yihe, and Agostinho Antunes. 2022. "Biomedical Potential of the Neglected Molluscivorous and Vermivorous Conus Species" Marine Drugs 20, no. 2: 105. https://doi.org/10.3390/md20020105
APA StyleZhao, Y., & Antunes, A. (2022). Biomedical Potential of the Neglected Molluscivorous and Vermivorous Conus Species. Marine Drugs, 20(2), 105. https://doi.org/10.3390/md20020105