
Bioactive Marine Xanthones: A Review


 ,
, 
 ,
,  and
and 
Abstract
:
1. Introduction
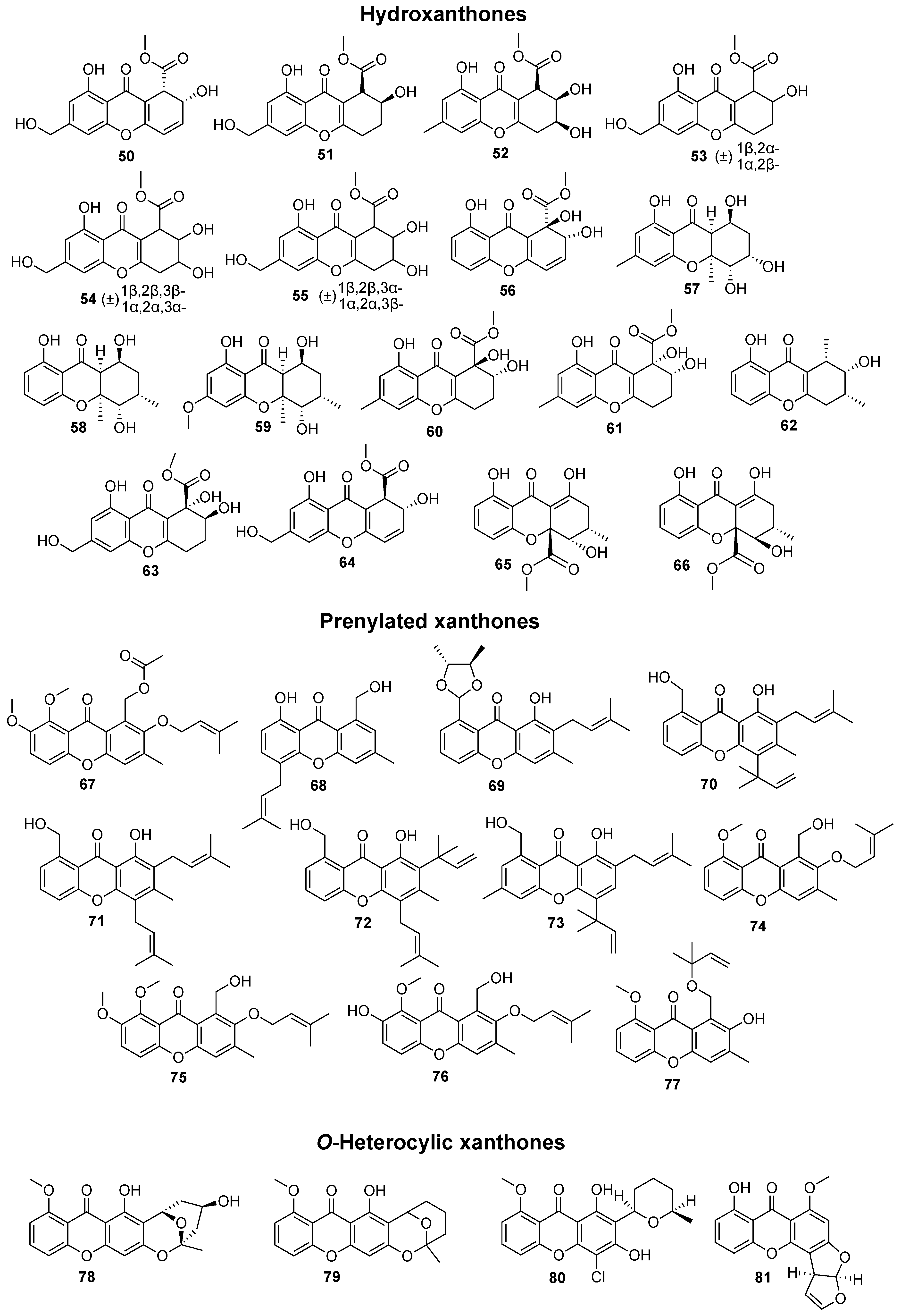
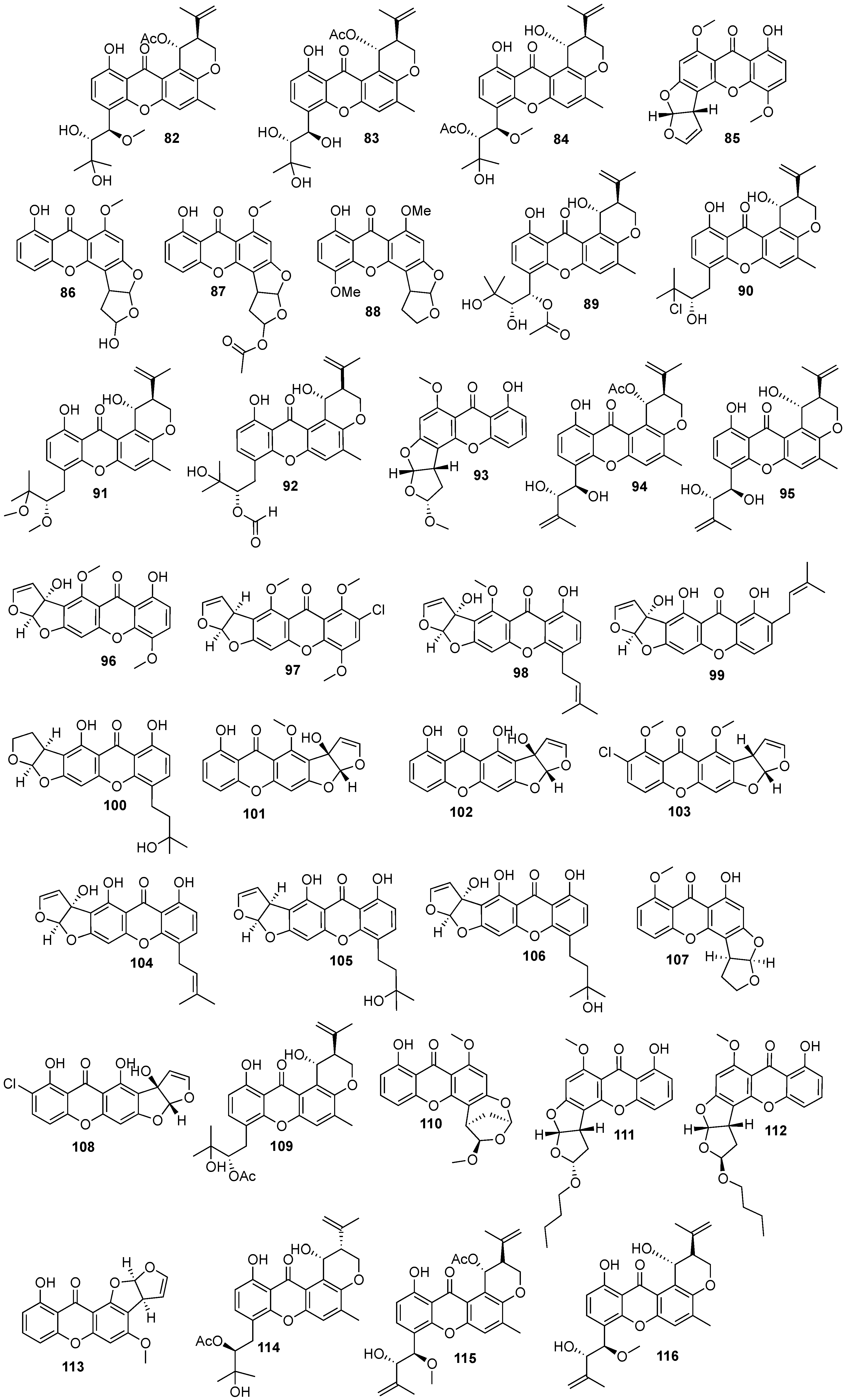
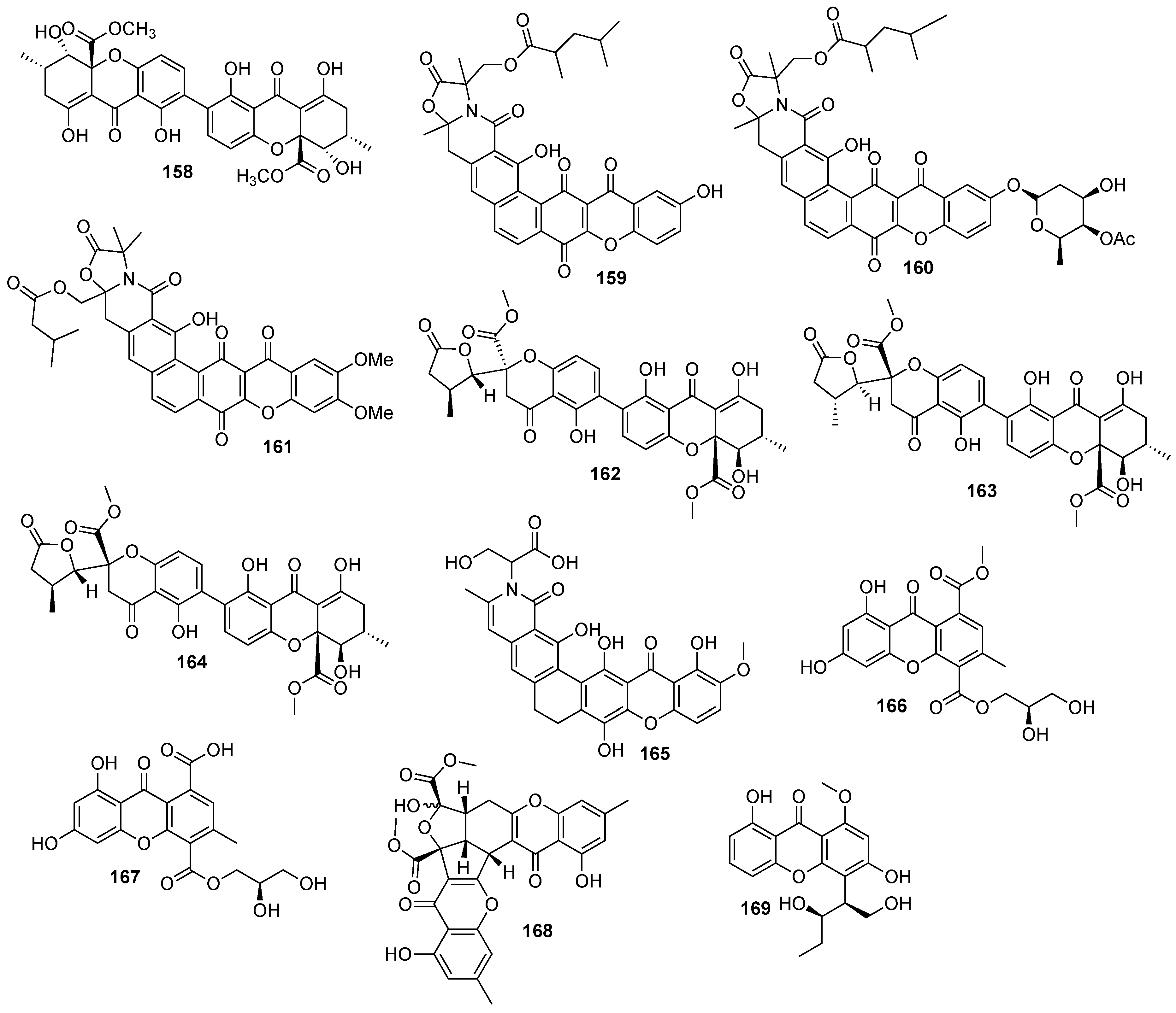
2. Bioactive Xanthones Isolated from the Marine Environment
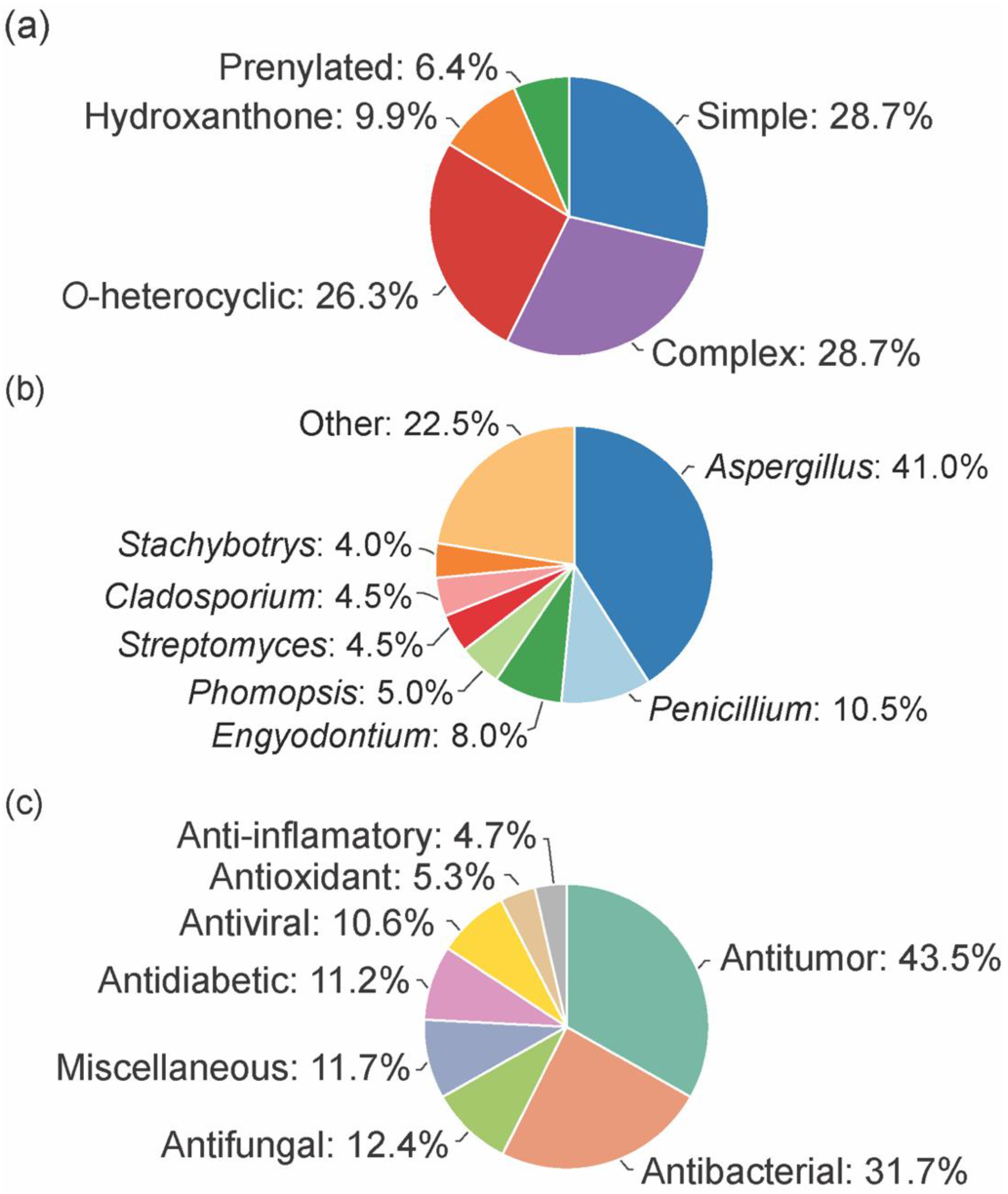
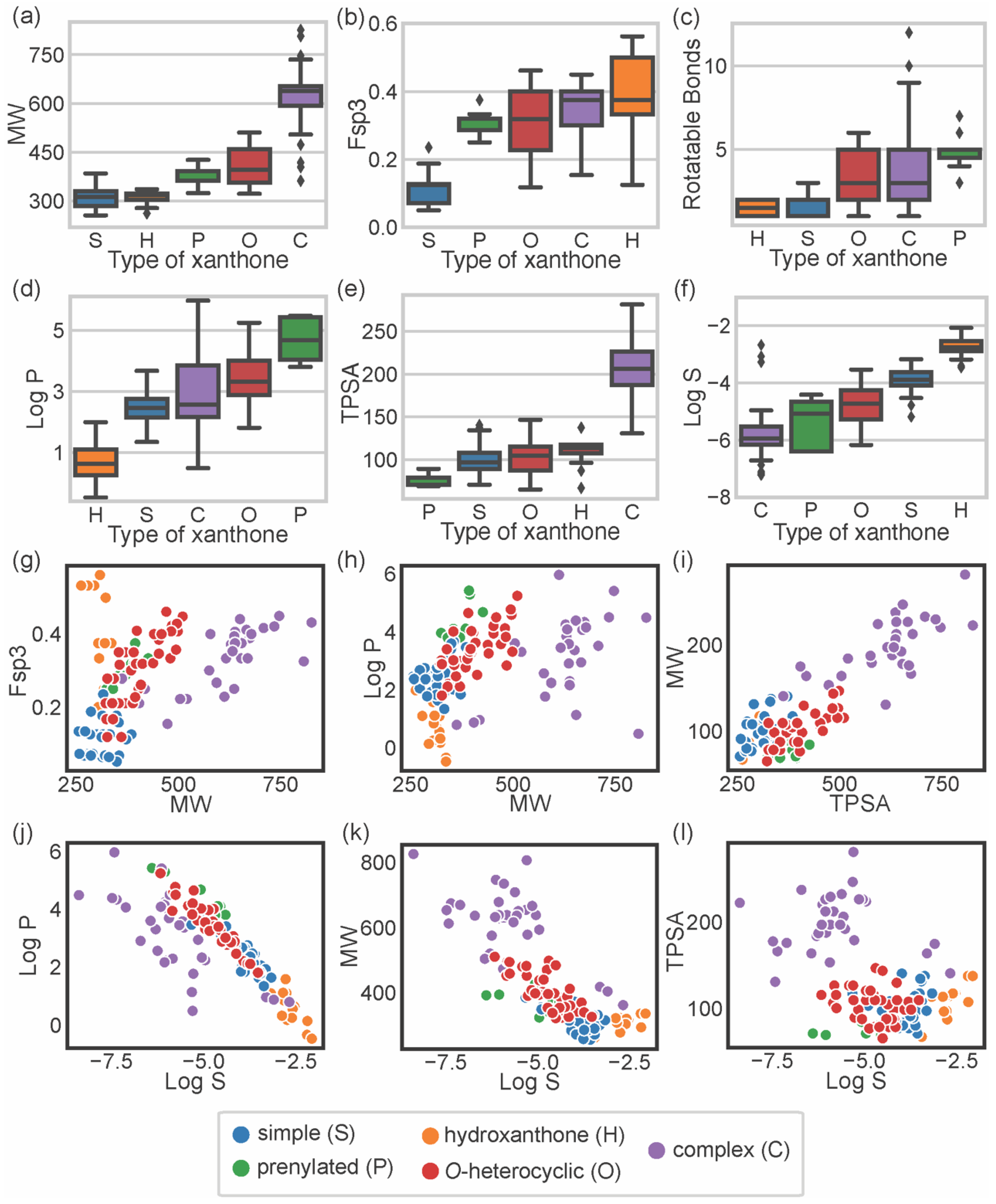
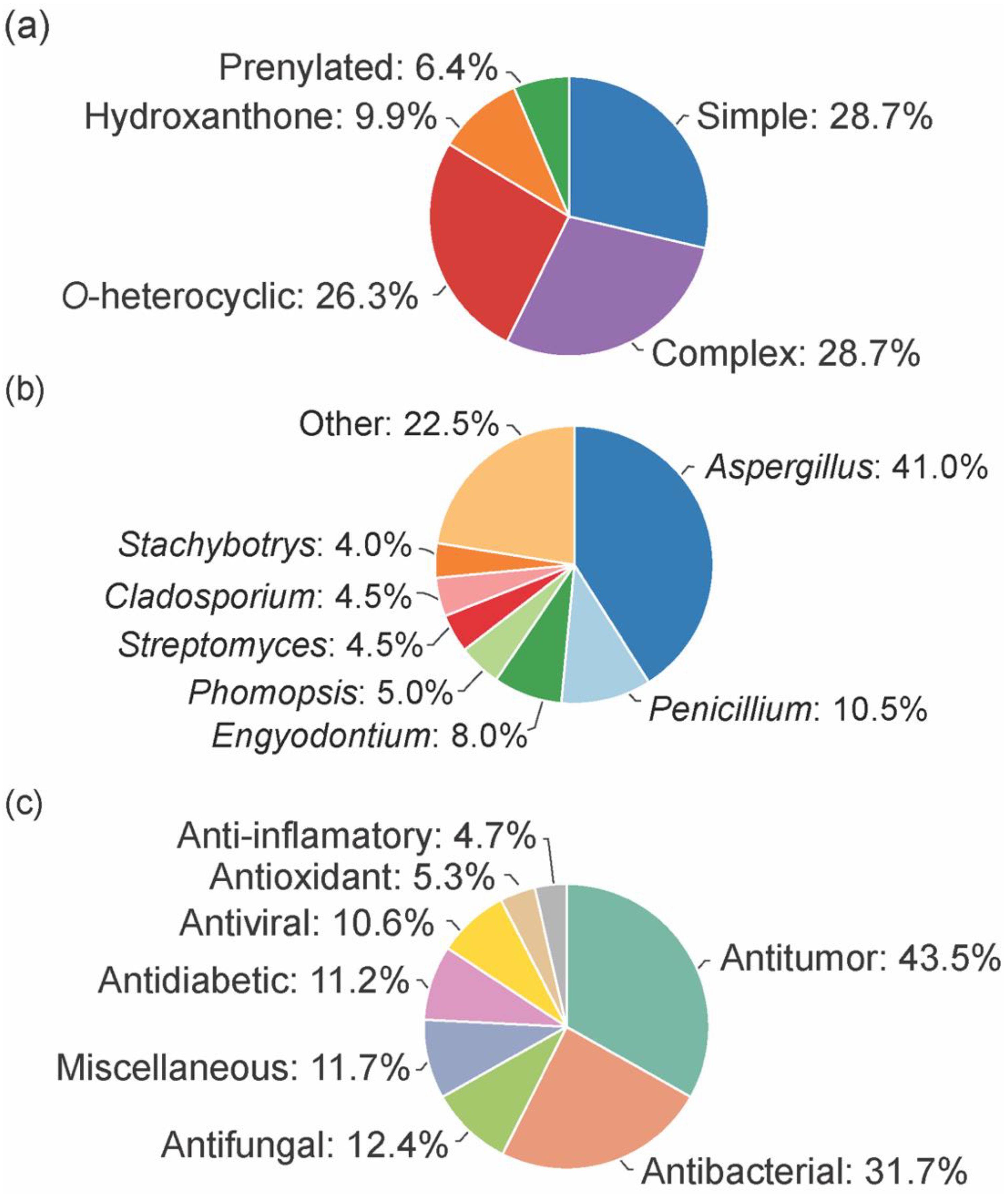
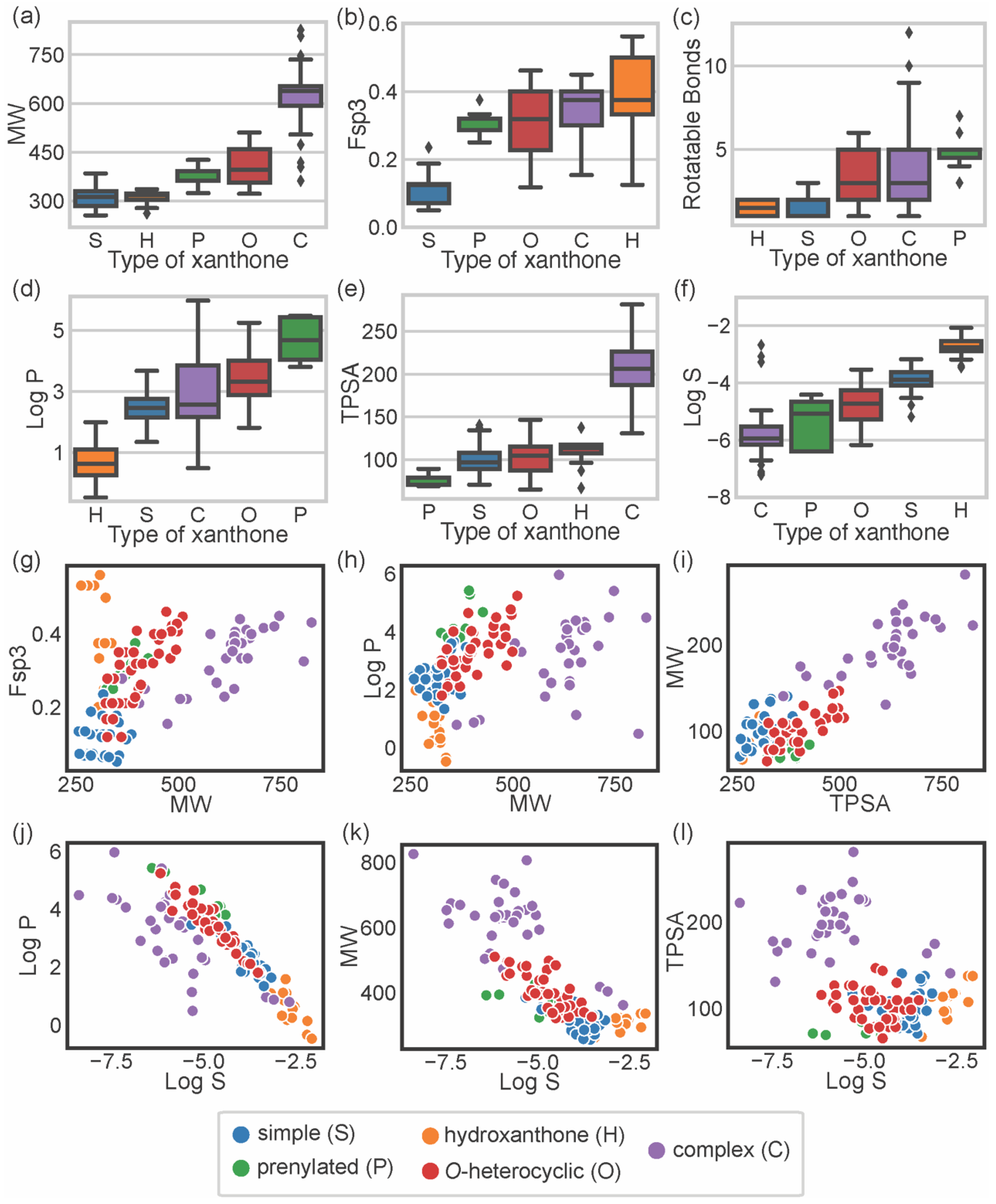
3. Chemical Space of Bioactive Marine Xanthones
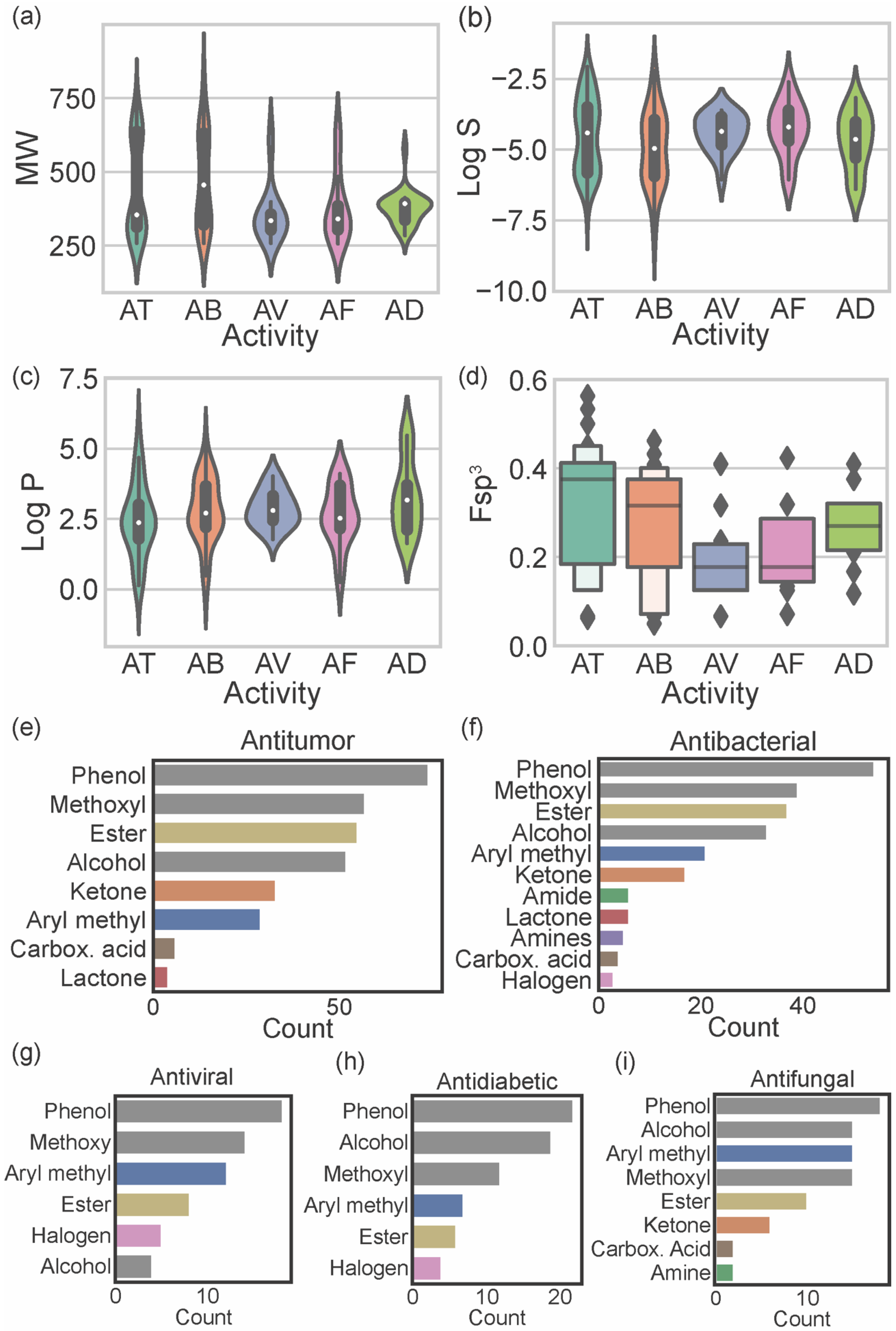
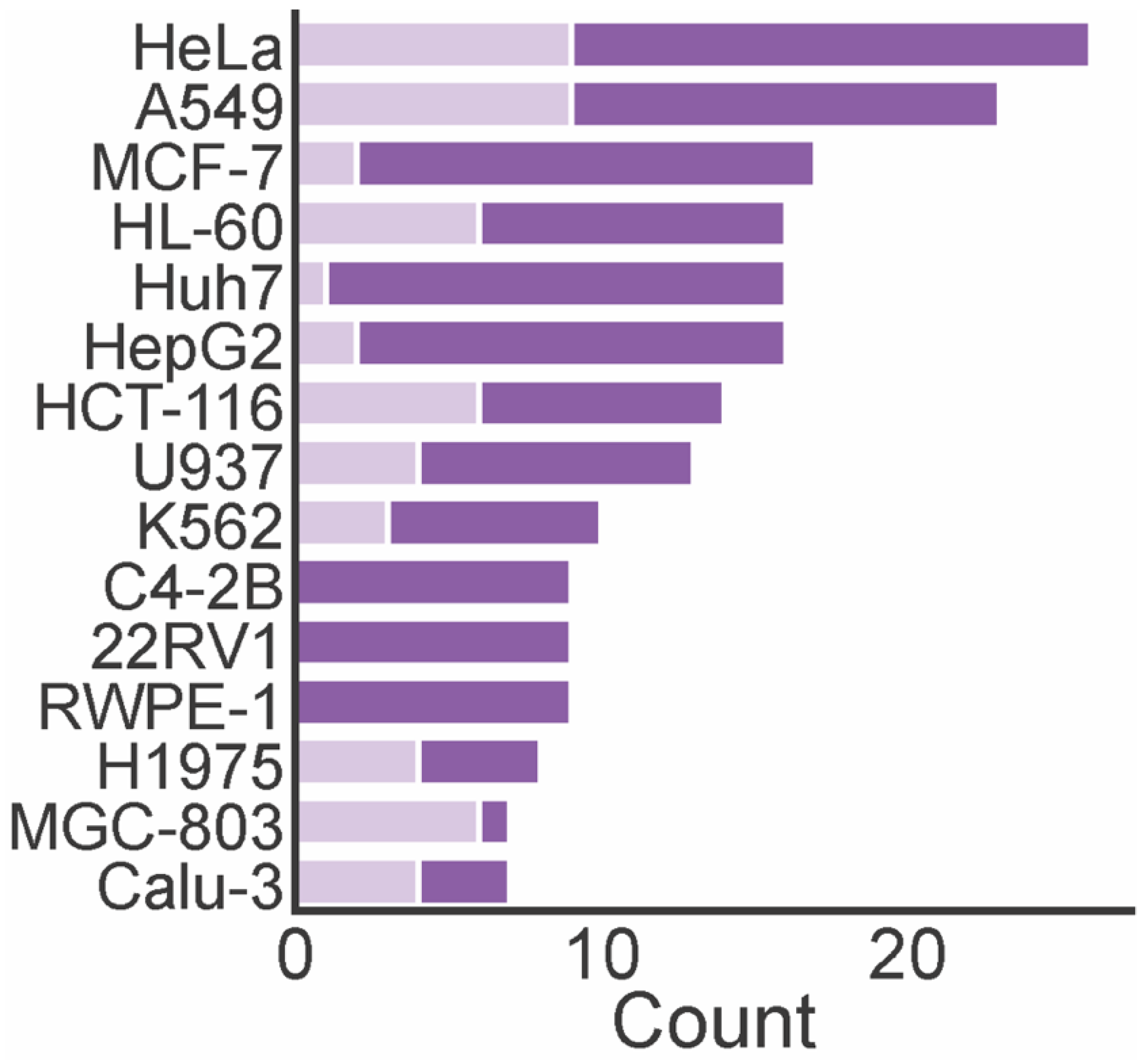
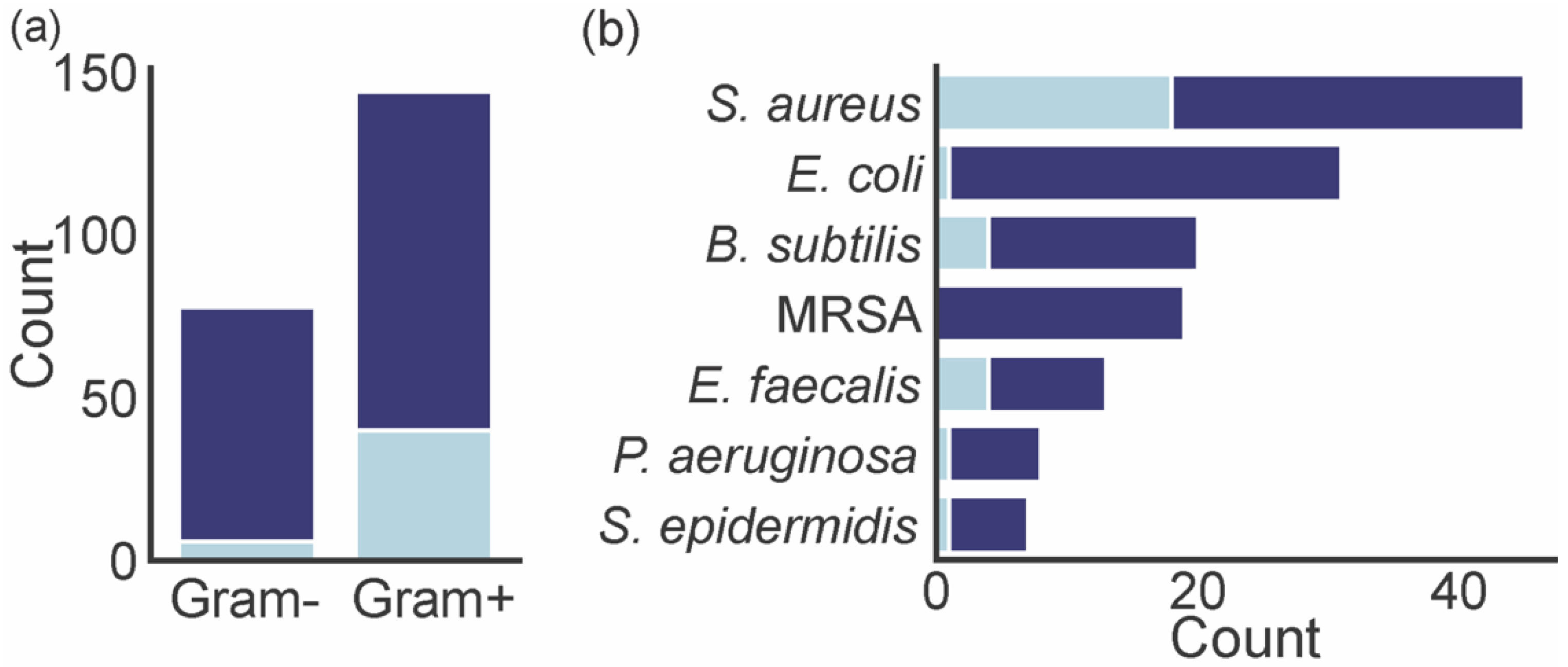
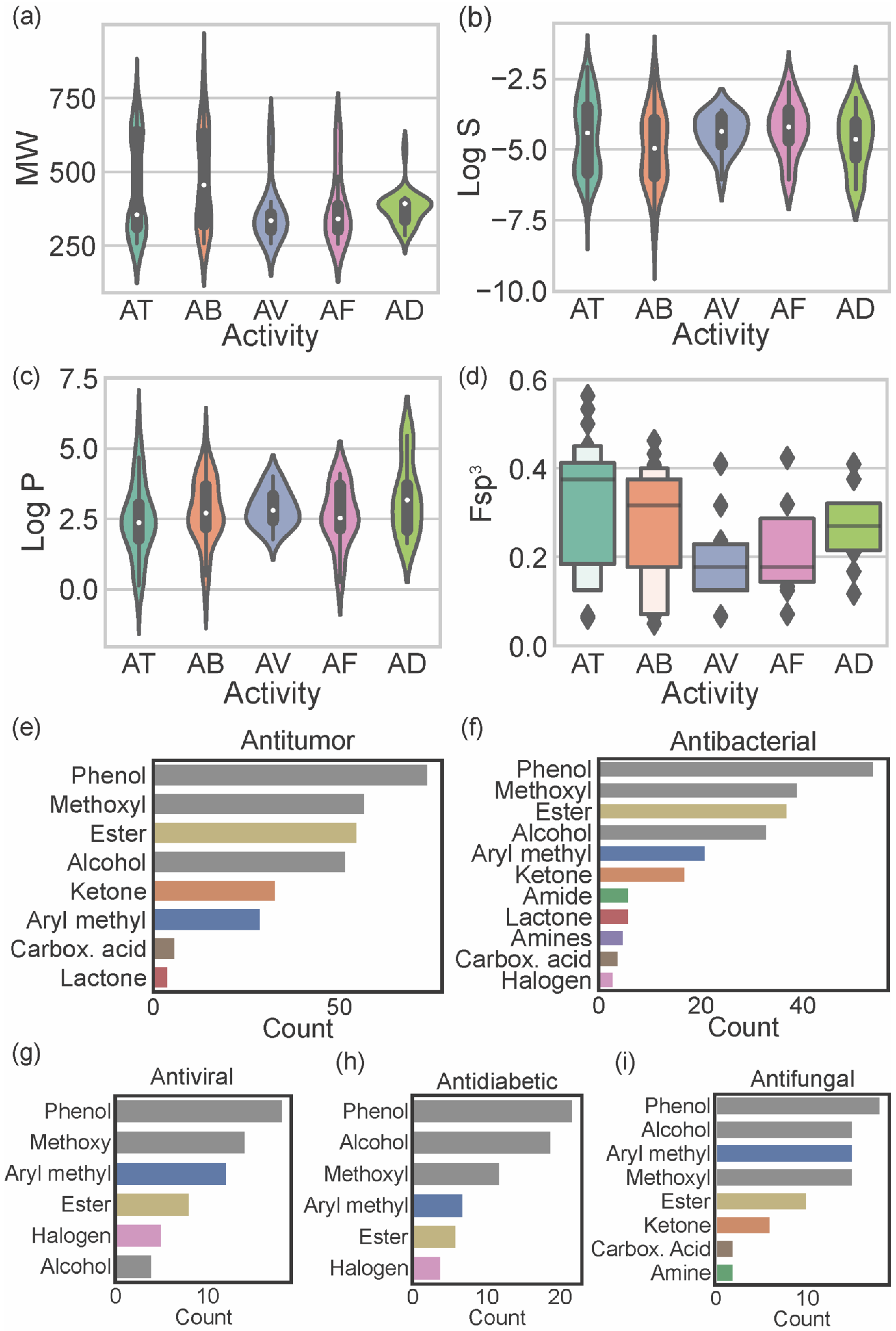
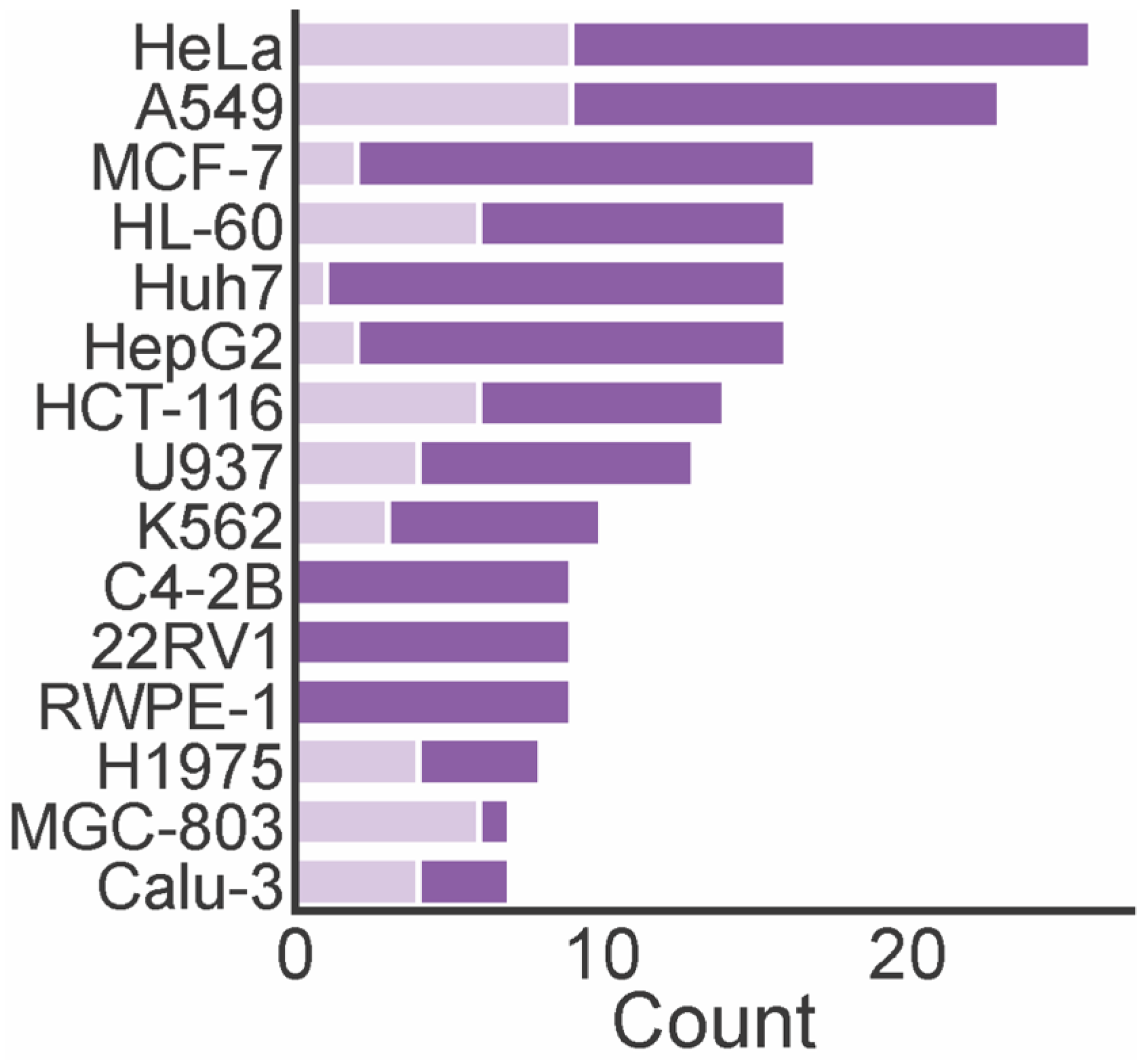
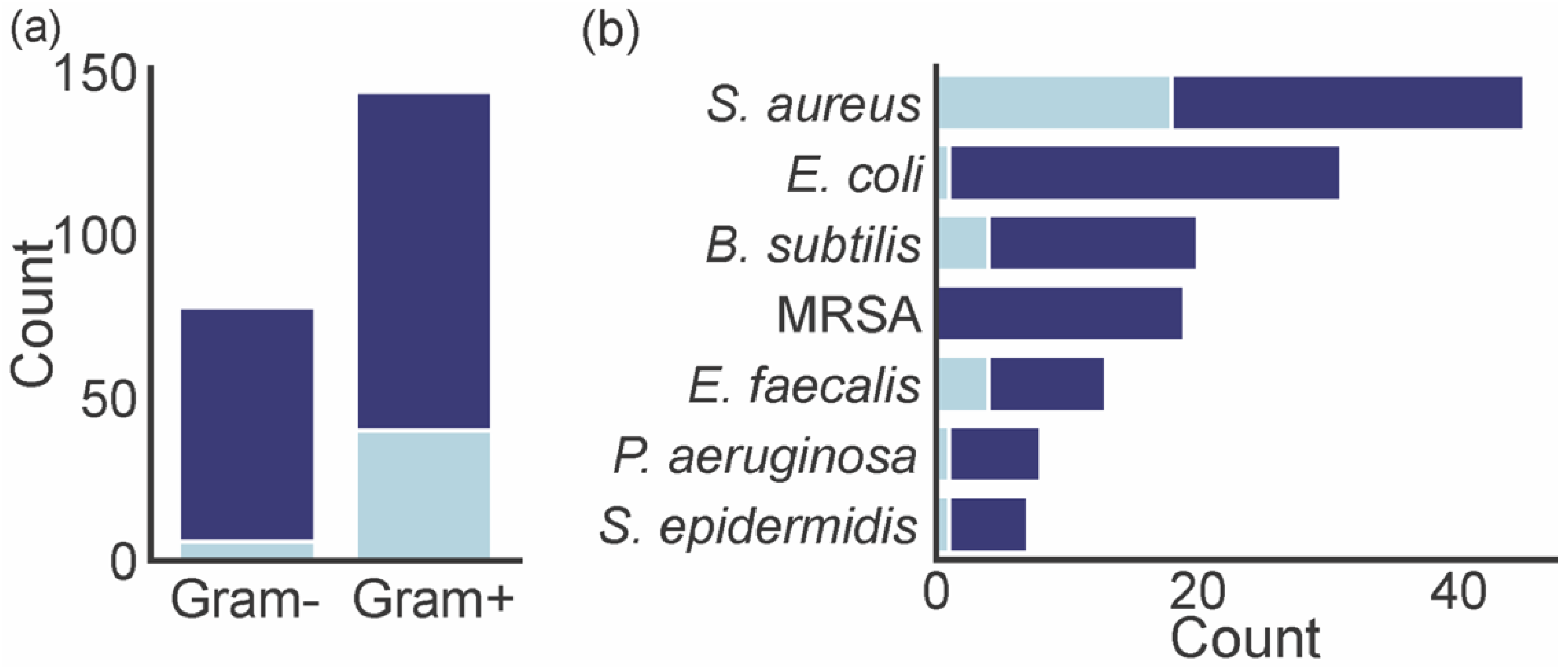
4. Biological Activities of Marine Xanthones
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shang, J.; Hu, B.; Wang, J.; Zhu, F.; Kang, Y.; Li, D.; Sun, H.; Kong, D.-X.; Hou, T. Cheminformatic Insight into the Differences between Terrestrial and Marine Originated Natural Products. J. Chem. Inf. Model. 2018, 58, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B.S. Lessons from the Past and Charting the Future of Marine Natural Products Drug Discovery and Chemical Biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, D.R.P.; Magalhães, Á.F.; Soares, J.X.; Pinto, J.; Azevedo, C.M.G.; Vieira, S.; Henriques, A.; Ferreira, H.; Neves, N.; Bousbaa, H.; et al. Yicathins B and C and Analogues: Total Synthesis, Lipophilicity and Biological Activities. ChemMedChem 2020, 15, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, D.R.P.; Soares, J.X.; Costa, J.C.; Magalhães, Á.F.; Azevedo, C.M.G.; Pinto, M.M.M.; Afonso, C.M.M. Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review. Molecules 2019, 24, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, M.M.M.; Castanheiro, R.A.P.; Kijjoa, A. Xanthones from marine-derived microorganisms: Isolation, structure elucidation and biological activities. In Encyclopedia of Analytical Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 1–21. ISBN 978-0-470-02731-8. [Google Scholar]
- Pinto, M.M.M.; Palmeira, A.; Fernandes, C.; Resende, D.I.S.P.; Sousa, E.; Cidade, H.; Tiritan, M.E.; Correia-da-Silva, M.; Cravo, S. From Natural Products to New Synthetic Small Molecules: A Journey through the World of Xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef]
- Oprea, T.I.; Gottfries, J. Chemography: The Art of Navigating in Chemical Space. J. Comb. Chem. 2001, 3, 157–166. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular Properties That Influence the Oral Bioavailability of Drug Candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A Knowledge-Based Approach in Designing Combinatorial or Medicinal Chemistry Libraries for Drug Discovery. 1. A Qualitative and Quantitative Characterization of Known Drug Databases. J. Comb. Chem. 1999, 1, 55–68. [Google Scholar] [CrossRef]
- Egan, W.J.; Merz, K.M.; Baldwin, J.J. Prediction of Drug Absorption Using Multivariate Statistics. J. Med. Chem. 2000, 43, 3867–3877. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.P. Generation of a Set of Simple, Interpretable ADMET Rules of Thumb. J. Med. Chem. 2008, 51, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Bickerton, G.R.; Paolini, G.V.; Besnard, J.; Muresan, S.; Hopkins, A.L. Quantifying the Chemical Beauty of Drugs. Nat. Chem. 2012, 4, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertl, P.; Roggo, S.; Schuffenhauer, A. Natural Product-Likeness Score and Its Application for Prioritization of Compound Libraries. J. Chem. Inf. Model. 2008, 48, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Jayaseelan, K.V.; Moreno, P.; Truszkowski, A.; Ertl, P.; Steinbeck, C. Natural Product-Likeness Score Revisited: An Open-Source, Open-Data Implementation. BMC Bioinform. 2012, 13, 106. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsson, P.; Kihlberg, J. How Big Is Too Big for Cell Permeability? J. Med. Chem. 2017, 60, 1662–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Cherukupalli, S.; Jing, L.; Liu, X.; Zhan, P. Fsp3: A New Parameter for Drug-Likeness. Drug Discov. 2020, 25, 1839–1845. [Google Scholar] [CrossRef]
- Hann, M.M.; Keserü, G.M. Finding the Sweet Spot: The Role of Nature and Nurture in Medicinal Chemistry. Nat. Rev. Drug Discov. 2012, 11, 355–365. [Google Scholar] [CrossRef]
- Caron, G.; Ermondi, G. Molecular Descriptors for Polarity: The Need for Going beyond Polar Surface Area. Future Med. Chem. 2016, 8, 2013–2016. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Edward, H.K.; Guy, T.C. Drug-Like Property Concepts in Pharmaceutical Design. Curr. Pharm. Des. 2009, 15, 2184–2194. [Google Scholar] [CrossRef]
- Meanwell, N.A. Improving Drug Candidates by Design: A Focus on Physicochemical Properties As a Means of Improving Compound Disposition and Safety. Chem. Res. Toxicol. 2011, 24, 1420–1456. [Google Scholar] [CrossRef] [PubMed]
- Sorokina, M.; Steinbeck, C. NaPLeS: A Natural Products Likeness Scorer—Web Application and Database. J. Cheminformatics 2019, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Stratton, C.F.; Newman, D.J.; Tan, D.S. Cheminformatic Comparison of Approved Drugs from Natural Product versus Synthetic Origins. Bioorg. Med. Chem. Lett. 2015, 25, 4802–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doak, B.C.; Over, B.; Giordanetto, F.; Kihlberg, J. Oral Druggable Space beyond the Rule of 5: Insights from Drugs and Clinical Candidates. Chem. Biol. 2014, 21, 1115–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A. The Impact of Natural Products upon Modern Drug Discovery. Curr. Opin. Chem. Biol. 2008, 12, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.F.; Wang, J.E.; Zhang, X.Y.; Nong, X.H.; Xu, X.Y.; Qi, S.H. Cytotoxic Polyketides from the Deep-Sea-Derived Fungus Engyodontium Album DFFSCS021. Mar. Drugs 2014, 12, 5902–5915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.X.; Qiu, S.; She, Z.; Lin, Y. A New Xanthone Derivative from the Marine Fungus Phomopsis sp. (No. SK7RN3G1). Chem. Nat. Compd. 2013, 49, 31–33. [Google Scholar] [CrossRef]
- Krick, A.; Kehraus, S.; Gerhäuser, C.; Klimo, K.; Nieger, M.; Maier, A.; Fiebig, H.H.; Atodiresei, I.; Raabe, G.; Fleischhauer, J.; et al. Potential Cancer Chemopreventive in Vitro Activities of Monomeric Xanthone Derivatives from the Marine Algicolous Fungus Monodictys Putredinis. J. Nat. Prod. 2007, 70, 353–360. [Google Scholar] [CrossRef]
- Wang, C.N.; Lu, H.M.; Gao, C.H.; Guo, L.; Zhan, Z.Y.; Wang, J.J.; Liu, Y.H.; Xiang, S.T.; Wang, J.; Luo, X.W. Cytotoxic Benzopyranone and Xanthone Derivatives from a Coral Symbiotic Fungus Cladosporium Halotolerans GXIMD 02502. Nat. Prod. Res. 2020, 27, 1–8. [Google Scholar] [CrossRef]
- Wang, W.Y.; Gao, M.L.; Luo, Z.H.; Liao, Y.Y.; Zhang, B.B.; Ke, W.Q.; Shao, Z.Z.; Li, F.; Chen, J.M. Secondary Metabolites Isolated from the Deep Sea-Derived Fungus Aspergillus Sydowii C1-S01-A7. Nat. Prod. Res. 2019, 33, 3077–3082. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.Q.; Li, D.; Peng, J.X.; Zhu, T.J.; Gu, Q.Q.; Li, D.H. Penicitols A-C and Penixanacid A from the Mangrove-Derived Penicillium Chrysogenum HDN11-24. J. Nat. Prod. 2015, 78, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Lin, X.P.; Lu, X.; Wan, J.T.; Zhou, X.F.; Liao, S.R.; Tu, Z.C.; Xu, S.H.; Liu, Y.H. Sesquiterpenoids and Xanthones Derivatives Produced by Sponge-Derived Fungus Stachybotry sp. HH1 ZSDS1F1-2. J. Antibiot. 2015, 68, 121–125. [Google Scholar] [CrossRef]
- Yamazaki, H.; Rotinsulu, H.; Kaneko, T.; Murakami, K.; Fujiwara, H.; Ukai, K.; Namikoshi, M. A New Dibenz[b,e]Oxepine Derivative, 1-Hydroxy-10-Methoxy-Dibenz[b,e]Oxepin-6,11-Dione, from a Marine-Derived Fungus, Beauveria Bassiana TPU942. Mar. Drugs 2012, 10, 2691–2697. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.M.; Wei, X.Y.; Lin, X.P.; Zhou, X.F.; Dong, J.D.; Yang, B. Penixanthones A and B, Two New Xanthone Derivatives from Fungus Penicillium sp. SYFz-1 Derived of Mangrove Soil Sample. Nat. Prod. Res. 2017, 31, 2218–2222. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.Q.; Qin, X.C.; Lin, X.P.; Kaliyaperumal, K.; Zhou, X.F.; Liu, J.; Ju, Z.R.; Tu, Z.C.; Liu, Y.H. Sydoxanthone C and Acremolin B Produced by Deep-Sea-Derived Fungus Aspergillus sp. SCSIO Ind09F01. J. Antibiot. 2015, 68, 703–706. [Google Scholar] [CrossRef]
- Elnaggar, M.S.; Ebada, S.S.; Ashour, M.L.; Ebrahim, W.; Muller, W.E.G.; Mandi, A.; Kurtan, T.; Singab, A.; Lin, W.H.; Liu, Z.; et al. Xanthones and Sesquiterpene Derivatives from a Marine-Derived Fungus Scopulariopsis sp. Tetrahedron 2016, 72, 2411–2419. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Guo, W.Q.; Che, Q.; Zhu, T.J.; Gu, Q.Q.; Li, D.H. Versicones E-H and Arugosin K Produced by the Mangrove-Derived Fungus Aspergillus Versicolor HDN11-84. J. Antibiot. 2017, 70, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Lin, Y.C.; She, Z.G.; Du, D.S.; Chan, W.L.; Zheng, Z.H. Paeciloxanthone, a New Cytotoxic Xanthone from the Marine Mangrove Fungus Paecilomyces sp. (Tree1-7). J. Asian Nat. Prod. Res. 2008, 10, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Pontius, A.; Krick, A.; Kehraus, S.; Brun, R.; König, G.M. Antiprotozoal Activities of Heterocyclic-Substituted Xanthones from the Marine-Derived Fungus Chaetomium sp. J. Nat. Prod. 2008, 71, 1579–1584. [Google Scholar] [CrossRef]
- Zhu, F.; Lin, Y.C. Three Xanthones from a Marine-Derived Mangrove Endophytic Fungus. Chem. Nat. Compd. 2007, 43, 132–135. [Google Scholar] [CrossRef]
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.A.; Kim, D.K.; Jung, J.H. Bioactive Metabolites from the Sponge-Derived Fungus Aspergillus Versicolor. Arch. Pharm. Res. 2010, 33, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Jiang, X.; Liu, X.P.; He, C.W.; Di, Y.X.; Lu, S.J.; Huang, H.L.; Lin, B.; Wang, D.; Fan, B.Y. Antibacterial Anthraquinone Dimers from Marine Derived Fungus Aspergillus sp. Fitoterapia 2019, 133, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Yang, M.Y.; Zhang, Y.H.; Shao, C.L.; Wang, C.Y.; Hu, L.D.; Cao, F.; Zhu, H.J. Absolute Configurations of 14,15-Hydroxylated Prenylxanthones from a Marine-Derived Aspergillus sp. Fungus by Chiroptical Methods. Sci. Rep. 2018, 8, 10621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.X.; Zhu, T.J.; Du, L.; Zhao, B.Y.; Li, D.H.; Gu, Q.Q. Sterigmatocystins from the Deep-Sea-Derived Fungus Aspergillus Versicolor. J. Antibiot. 2011, 64, 193–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.A.; Lin, X.; Zhou, X.; Chen, M.; Huang, X.; Yang, B.; Tao, H. Xanthones and Quinolones Derivatives Produced by the Deep-Sea-Derived Fungus Penicillium sp. SCSIO Ind16F01. Molecules 2017, 22, 1999. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.; Yuan, J.; Huang, X.S.; Wen, W.T.; Zhu, X.; Liu, Y.Y.; Li, H.X.; Lu, Y.J.; He, L.; Tan, H.M.; et al. New Dimeric Members of the Phomoxanthone Family: Phomolactonexanthones A, B and Deacetylphomoxanthone C Isolated from the Fungus Phomopsis sp. Mar. Drugs 2013, 11, 4961–4972. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.; Kimishima, A.; Setiawan, A.; Arai, M. Secalonic Acid D as a Selective Cytotoxic Substance on the Cancer Cells Adapted to Nutrient Starvation. J. Nat. Med. 2020, 74, 495–500. [Google Scholar] [CrossRef]
- Zhen, X.; Gong, T.; Wen, Y.H.; Yan, D.J.; Chen, J.J.; Zhu, P. A Chrysoxanthones A-C, Three New Xanthone-Chromanone Heterdimers from Sponge-Associated Penicillium Chrysogenum HLS111 Treated with Histone Deacetylase Inhibitor. Mar. Drugs 2018, 16, 357. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, Y.P.; Li, X.X.; Lu, Z.H.; Zheng, Q.H.; Liu, Q.Y. Isolation of 4,4′-Bond Secalonic Acid D from the Marine-Derived Fungus Penicillium Oxalicum with Inhibitory Property against Hepatocellular Carcinoma. J. Antibiot. 2019, 72, 34–44. [Google Scholar] [CrossRef]
- Wu, G.W.; Qi, X.; Mo, X.M.; Yu, G.H.; Wang, Q.; Zhu, T.J.; Gu, Q.Q.; Liu, M.; Li, J.; Li, D.H. Structure-Based Discovery of Cytotoxic Dimeric Tetrahydroxanthones as Potential Topoisomerase I Inhibitors from a Marine-Derived Fungus. Eur. J. Med. Chem. 2018, 148, 268–278. [Google Scholar] [CrossRef]
- Liu, L.L.; He, L.S.; Xu, Y.; Han, Z.; Li, Y.X.; Zhong, J.L.; Guo, X.R.; Zhang, X.X.; Ko, K.M.; Qian, P.Y. Caspase-3-Dependent Apoptosis of Citreamicin ε-Induced HeLa Cells Is Associated with Reactive Oxygen Species Generation. Chem. Res. Toxicol. 2013, 26, 1055–1063. [Google Scholar] [CrossRef]
- Yeon, J.T.; Kim, H.; Kim, K.J.; Lee, J.; Won, D.H.; Nam, S.J.; Kim, S.H.; Kang, H.; Son, Y.J. Acredinone C and the Effect of Acredinones on Osteoclastogenic and Osteoblastogenic Activity. J. Nat. Prod. 2016, 79, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Pavão, G.B.; Venâncio, V.P.; de Oliveira, A.L.L.; Hernandes, L.C.; Almeida, M.R.; Antunes, L.M.G.; Debonsi, H.M. Differential Genotoxicity and Cytotoxicity of Phomoxanthone A Isolated from the Fungus Phomopsis Longicolla in HL60 Cells and Peripheral Blood Lymphocytes. Toxicol. Vitro 2016, 37, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Ueda, J.Y.; Takagi, M.; Shin-Ya, K. New Xanthoquinodin-like Compounds, JBIR-97,-98 and-99, Obtained from Marine Sponge-Derived Fungus Tritirachium sp. SpB081112MEf2. J. Antibiot. 2010, 63, 615–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, K.; Chung, B.; Shin, Y.; Rheingold, A.L.; Moore, C.E.; Park, S.J.; Park, S.; Lee, S.K.; Oh, K.B.; Shin, J.; et al. Pentacyclic Antibiotics from a Tidal Mud Flat-Derived Actinomycete. J. Nat. Prod. 2015, 78, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, Y.; Kawahara, T.; Hitora, Y.; Tsukamoto, S. Ukixanthomycin A: A Hexacyclic Xanthone from the Mudflat-Derived Actinomycete Streptomyces sp. Heterocycles 2020, 100, 1686–1693. [Google Scholar] [CrossRef]
- Li, H.L.; Li, X.M.; Liu, H.; Meng, L.H.; Wang, B.G. Two New Diphenylketones and a New Xanthone from Talaromyces Islandicus EN-501, an Endophytic Fungus Derived from the Marine Red Alga Laurencia Okamurai. Mar. Drugs 2016, 14, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Cai, X.L.; Yang, H.; Xia, X.K.; Guo, Z.Y.; Yuan, J.; Li, M.F.; She, Z.G.; Lin, Y.C. The Bioactive Metabolites of the Mangrove Endophytic Fungus Talaromyces sp. ZH-154 Isolated from Kandelia Candel (L.) Druce. Planta Med. 2010, 76, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.R.; Miao, F.P.; Zhang, J.; Wang, G.; Yin, X.L.; Ji, N.Y. Three New Xanthone Derivatives from an Algicolous Isolate of Aspergillus Wentii. Magn. Reson. Chem. 2013, 51, 65–68. [Google Scholar] [CrossRef]
- Wang, J.H.; Ding, W.J.; Wang, R.M.; Du, Y.P.; Liu, H.L.; Kong, X.H.; Li, C.Y. Identification and Bioactivity of Compounds from the Mangrove Endophytic Fungus Alternaria sp. Mar. Drugs 2015, 13, 4492–4504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.B.; Chen, W.J.; Shan, T.Z.; Sun, B.Y.; Yan, P.C.; Jiang, W. Antibacterial Diphenyl Ether, Benzophenone and Xanthone Derivatives from Aspergillus Flavipes. Chem. Biodivers. 2020, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Gao, J.; Hu, J.; He, H.; Huang, P.; Zhang, L.; Song, F. One New Xanthenone from the Marine-Derived Fungus Aspergillus Versicolor MF160003. Nat. Prod. Res. 2020, 34, 2907–2912. [Google Scholar] [CrossRef]
- Khoshbakht, M.; Thanaussavadate, B.; Zhu, C.X.; Cao, Y.; Zakharov, L.N.; Loesgen, S.; Blakemore, P.R. Total Synthesis of Chalaniline B: An Antibiotic Aminoxanthone from Vorinostat-Treated Fungus Chalara sp. 6661. J. Org. Chem. 2021, 86, 7773–7780. [Google Scholar] [CrossRef]
- Zhang, W.; Krohn, K.; Zia-Ullah, F.U.; Pescitelli, G.; Di Bari, L.; Antus, S.; Kurtán, T.; Rheinheimer, J.; Draeger, S.; Schulz, B. New Mono- and Dimeric Members of the Secalonic Acid Family: Blennolides A–G Isolated from the Fungus Blennoria sp. Chem. Eur. J. 2008, 14, 4913–4923. [Google Scholar] [CrossRef]
- Song, F.H.; Ren, B.; Chen, C.X.; Yu, K.; Liu, X.R.; Zhang, Y.H.; Yang, N.; He, H.T.; Liu, X.T.; Dai, H.Q.; et al. Three New Sterigmatocystin Analogues from Marine-Derived Fungus Aspergillus Versicolor MF359. Appl. Microbiol. Biotechnol. 2014, 98, 3753–3758. [Google Scholar] [CrossRef] [PubMed]
- Fredimoses, M.; Zhou, X.F.; Ai, W.; Tian, X.P.; Yang, B.; Lin, X.P.; Liu, J.; Liu, Y.H. Emerixanthone E, a New Xanthone Derivative from Deep Sea Fungus Emericella sp. SCSIO 05240. Nat. Prod. Res. 2019, 33, 2088–2094. [Google Scholar] [CrossRef] [PubMed]
- Fredimoses, M.; Zhou, X.F.; Lin, X.P.; Tian, X.P.; Ai, W.; Wang, J.F.; Liao, S.R.; Liu, J.; Yang, B.; Yang, X.W.; et al. New Prenylxanthones from the Deep-Sea Derived Fungus Emericella sp. SCSIO 05240. Mar. Drugs 2014, 12, 3190–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmstrøm, J.; Christophersen, C.; Barrero, A.F.; Enrique Oltra, J.; Justicia, J.; Rosales, A. Bioactive Metabolites from a Marine-Derived Strain of the Fungus Emericella Variecolor. J. Nat. Prod. 2002, 65, 364–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.Q.; Lin, S.T.; Kumaravel, K.; Zhou, H.; Wang, S.Y.; Liu, Y.H. Polyketide-Derived Metabolites from the Sponge-Derived Fungus Aspergillus sp. F40. Phytochem. Lett. 2018, 27, 74–77. [Google Scholar] [CrossRef]
- Erbert, C.; Lopes, A.A.; Yokoya, N.S.; Furtado, N.; Conti, R.; Pupo, M.T.; Lopes, J.L.C.; Debonsi, H.M. Antibacterial Compound from the Endophytic Fungus Phomopsis Longicolla Isolated from the Tropical Red Seaweed Bostrychia Radicans. Bot. Mar. 2012, 55, 435–440. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, X.P.; Ju, Z.R.; Liao, X.J.; Huang, X.J.; Zhang, C.; Zhao, B.X.; Xu, S.H. Aspergchromones A and B, Two New Polyketides from the Marine Sponge-Associated Fungus Aspergillus sp. SCSIO XWS03F03. J. Asian Nat. Prod. Res. 2017, 19, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Sun, Y.L.; Zhang, X.Y.; Han, Z.; Gao, H.C.; He, F.; Qian, P.Y.; Qi, S.H. Antifouling and Antibacterial Polyketides from Marine Gorgonian Coral-Associated Fungus Penicillium sp. SCSGAF 0023. J. Antibiot. 2013, 66, 219–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, Antibiotic Polyketides from the Marine Fungus Engyodontium Album Strain LF069. Chem. Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef] [PubMed]
- Eltamany, E.E.; Abdelmohsen, U.R.; Ibrahim, A.K.; Hassanean, H.A.; Hentschel, U.; Ahmed, S.A. New Antibacterial Xanthone from the Marine Sponge-Derived Micrococcus sp. EG45. Bioorg. Med. Chem. Lett. 2014, 24, 4939–4942. [Google Scholar] [CrossRef]
- Liu, L.L.; Xu, Y.; Han, Z.; Li, Y.X.; Lu, L.; Lai, P.Y.; Zhong, J.L.; Guo, X.R.; Zhang, X.X.; Qian, P.Y. Four New Antibacterial Xanthones from the Marine-Derived Actinomycetes Streptomyces Caelestis. Mar. Drugs 2012, 10, 2571–2583. [Google Scholar] [CrossRef] [Green Version]
- Malet-Cascón, L.; Romero, F.; Espliego-Vázquez, F.; Grávalos, D.; Fernández-Puentes, J.L. IB-00208, a New Cytotoxic Polycyclic Xanthone Produced by a Marine-Derived Actinomadura. I. Isolation of the Strain, Taxonomy and Biological Activities. J. Antibiot. 2003, 56, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peoples, A.J.; Zhang, Q.B.; Millett, W.P.; Rothfeder, M.T.; Peseatore, B.C.; Madden, A.A.; Ling, L.L.; Moore, C.M. Neocitreamicins I and II, Novel Antibiotics with Activity against Methicillin-Resistant Staphylococcus Aureus and Vancomycin-Resistant Enterococci. J. Antibiot. 2008, 61, 457–463. [Google Scholar] [CrossRef]
- Maiese, W.M.; Lechevalier, M.P.; Lechevalier, H.A.; Korshalla, J.; Goodman, J.; Wildey, M.J.; Kuck, N.; Greenstein, M. LL-E19085 Alpha, a Novel Antibiotic from Micromonospora Citrea: Taxonomy, Fermentation and Biological Activity. J. Antibiot. 1989, 42, 846–851. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Lateff, A.; Klemke, C.; König, G.M.; Wright, A.D. Two New Xanthone Derivatives from the Algicolous Marine Fungus Wardomyces Anomalus. J. Nat. Prod. 2003, 66, 706–708. [Google Scholar] [CrossRef]
- Shao, C.; Wang, C.; Wei, M.; Gu, Y.; Xia, X.; She, Z.; Lin, Y. Structure Elucidation of Two New Xanthone Derivatives from the Marine Fungus Penicillium sp. (ZZF 32#) from the South China Sea. Magn. Reson. Chem. 2008, 46, 1066–1069. [Google Scholar] [CrossRef]
- Wang, X.; Mao, Z.G.; Song, B.B.; Chen, C.H.; Xiao, W.W.; Hu, B.; Wang, J.W.; Jiang, X.B.; Zhu, Y.H.; Wang, H.J. Advances in the Study of the Structures and Bioactivities of Metabolites Isolated from Mangrove-Derived Fungi in the South China Sea. Mar. Drugs 2013, 11, 3601–3616. [Google Scholar] [CrossRef] [Green Version]
- Höller, U.; König, G.M.; Wright, A.D. A New Tyrosine Kinase Inhibitor from a Marine Isolate of Ulocladium Botrytis and New Metabolites from the Marine Fungi Asteromyces Cruciatus and Varicosporina Ramulosa. Eur. J. Org. Chem. 1999, 2949–2955. [Google Scholar] [CrossRef]
- Li, C.Y.; Zhang, J.; Shao, C.L.; Ding, W.J.; She, Z.G.; Lin, Y.C. A New Xanthone Derivative from the Co-Culture Broth of Two Marine Fungi (Strain No. E33 and K38). Chem. Nat. Compd. 2011, 47, 382–384. [Google Scholar] [CrossRef]
- Marmann, A.; Aly, A.H.; Lin, W.H.; Wang, B.G.; Proksch, P. Co-Cultivation-A Powerful Emerging Tool for Enhancing the Chemical Diversity of Microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.F.; He, W.J.; Huang, X.L.; Tian, X.P.; Liao, S.R.; Yang, B.; Wang, F.Z.; Zhou, X.J.; Liu, Y.H. Antifungal New Oxepine-Containing Alkaloids and Xanthones from the Deep-Sea-Derived Fungus Aspergillus Versicolor SCSIO 05879. J. Agric. Food Chem. 2016, 64, 2910–2916. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.W.; Yang, J.; Chen, F.M.; Lin, X.P.; Chen, C.M.; Zhou, X.F.; Liu, S.W.; Liu, Y.H. Structurally Diverse Polyketides From the Mangrove-Derived Fungus Diaporthe Sp SCSIO 41011 With Their Anti-Influenza A Virus Activities. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.H.; Zhang, H.B.; Zhong, M.J.; Ma, L.Y.; Liu, D.S.; Liu, W.Z.; Ren, H. Potential Antiviral Xanthones from a Coastal Saline Soil Fungus Aspergillus Iizukae. Mar. Drugs 2018, 16, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.Z.; Peng, S.; Yang, J.; Cong, Z.W.; Lin, X.P.; Liao, S.R.; Yang, B.; Zhou, X.F.; Zhou, X.J.; Liu, Y.H.; et al. Structurally Diverse Sesquiterpenoids and Polyketides from a Sponge-Associated Fungus Aspergillus Sydowii SCSI041301. Fitoterapia 2019, 135, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Tang, X.L.; Luo, X.C.; Sun, C.X.; Liu, K.C.; Zhang, Y.; Li, P.L.; Li, G.Q. Isolation and Identification of Three New Sterigmatocystin Derivatives from the Fungus Aspergillus Versicolor Guided by Molecular Networking Approach. Chem. Biodivers. 2020, 17, 8. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.J.; Ouyang, M.A.; Tan, Q.W. New Asperxanthone and Asperbiphenyl from the Marine Fungus Aspergillus sp. Pest Manag. Sci. 2009, 65, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, B.; Liu, J.; Xun, T.; Liu, Y.; Zhou, X. Penicillixanthone A, a Marine-Derived Dual-Coreceptor Antagonist as Anti-HIV-1 Agent. Nat. Prod. Res. 2019, 33, 1467–1471. [Google Scholar] [CrossRef]
- Wang, J.F.; Zhou, L.M.; Chen, S.T.; Yang, B.; Liao, S.R.; Kong, F.D.; Lin, X.P.; Wang, F.Z.; Zhou, X.F.; Liu, Y.H. New Chlorinated Diphenyl Ethers and Xanthones from a Deep-Sea-Derived I Fungus Penicillium Chrysogenum SCSIO 41001. Fitoterapia 2018, 125, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Lin, C.Y.; Lu, C.J.; Chang, Y.M.; Che, Q.; Zhang, G.J.; Zhu, T.J.; Gu, Q.Q.; Wu, Z.Q.; Li, M.Y.; et al. Staprexanthones, Xanthone-Type Stimulators of Pancreatic Beta-Cell Proliferation from a Mangrove Endophytic Fungus. J. Nat. Prod. 2020, 83, 2996–3003. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Huang, Z.H.; Ma, X.; Zheng, Z.H.; Zhang, X.X.; Lu, X.H.; Qi, S.H. Mycotoxins as Inhibitors of Protein Tyrosine Phosphatases from the Deep-Sea-Derived Fungus Aspergillus Puniceus SCSIO Z021. Bioorganic Chem. 2021, 107, 104571. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; He, F.; Yu, J.H.; Zhai, H.J.; Cheng, Z.Q.; Jiang, C.S.; Zhang, Y.Y.; Zhang, Y.; Zhang, X.Y.; Chen, G.Y.; et al. New Chromones from a Marine-Derived Fungus, Arthrinium sp., and Their Biological Activity. Molecules 2018, 23, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.H.; Liu, D.; Xu, Y.; Chen, J.L.; Lin, W.H. Antioxidant Xanthones and Anthraquinones Isolated from a Marine-Derived Fungus Aspergillus Versicolor. Chin. J. Nat. Med. 2018, 16, 219–224. [Google Scholar] [CrossRef]
- Du, X.W.; Liu, D.; Huang, J.; Zhang, C.J.; Proksch, P.; Lin, W.H. Polyketide Derivatives from the Sponge Associated Fungus Aspergillus Europaeus with Antioxidant and NO Inhibitory Activities. Fitoterapia 2018, 130, 190–197. [Google Scholar] [CrossRef]
- Wang, J.F.; Xu, F.Q.; Wang, Z.; Lu, X.; Wan, J.T.; Yang, B.; Zhou, X.F.; Zhang, T.Y.; Tu, Z.C.; Liu, Y.H. A New Naphthalene Glycoside from the Sponge-Derived Fungus Arthrinium sp. ZSDS1-F3. Nat. Prod. Res. 2014, 28, 1070–1074. [Google Scholar] [CrossRef]
- Liu, H.J.; Chen, S.H.; Liu, W.Y.; Liu, Y.Y.; Huang, X.S.; She, Z.G. Polyketides with Immunosuppressive Activities from Mangrove Endophytic Fungus Penicillium sp. ZJ-SY2. Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Huang, J.X.; Zhou, X.F.; Lin, X.P.; Liu, J.; Liao, S.R.; Wang, J.F.; Liu, F.A.; Tao, H.M.; Liu, Y.H. The Fungal Metabolites with Potential Antiplasmodial Activity. Curr. Med. Chem. 2018, 25, 3796–3825. [Google Scholar] [CrossRef] [PubMed]
- Tempone, A.G.; de Oliveira, C.M.; Berlinck, R.G.S. Current Approaches to Discover Marine Antileishmanial Natural Products. Planta Med. 2011, 77, 572–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.; Zhang, X.W.; Zhang, M.; Li, W.; Ma, Z.Y.; Zhu, H.J.; Cao, F. Aspergixanthones I–K, New Anti-Vibrio Prenylxanthones from the Marine-Derived Fungus Aspergillus sp. ZA-01. Mar. Drugs 2018, 16, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.Q.; Li, X.M.; Xu, G.M.; Li, X.; An, C.Y.; Wang, B.G. Antibacterial Anthraquinone Derivatives Isolated from a Mangrove-Derived Endophytic Fungus Aspergillus Nidulans by Ethanol Stress Strategy. J. Antibiot. 2018, 71, 778–784. [Google Scholar] [CrossRef] [PubMed]
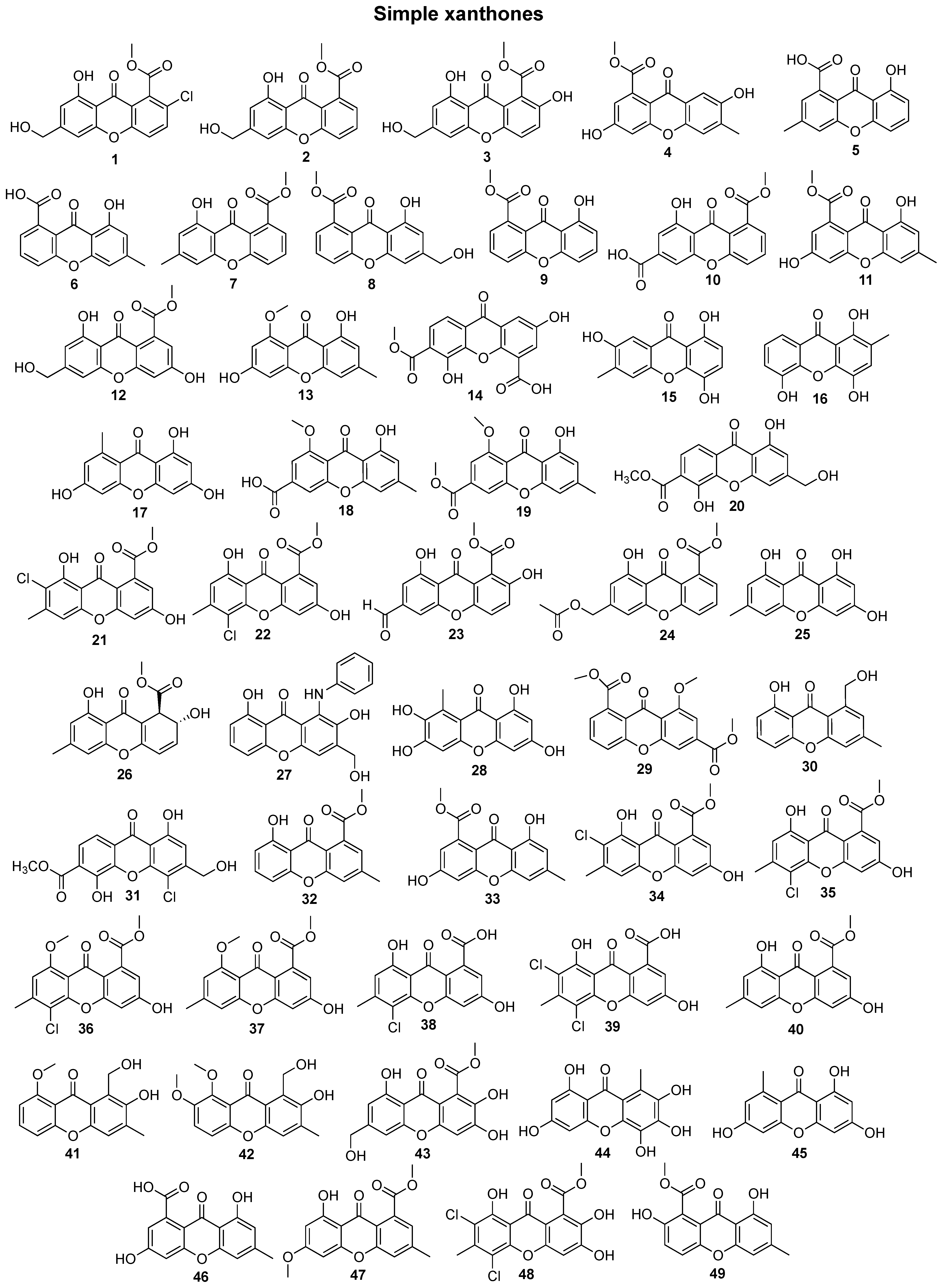

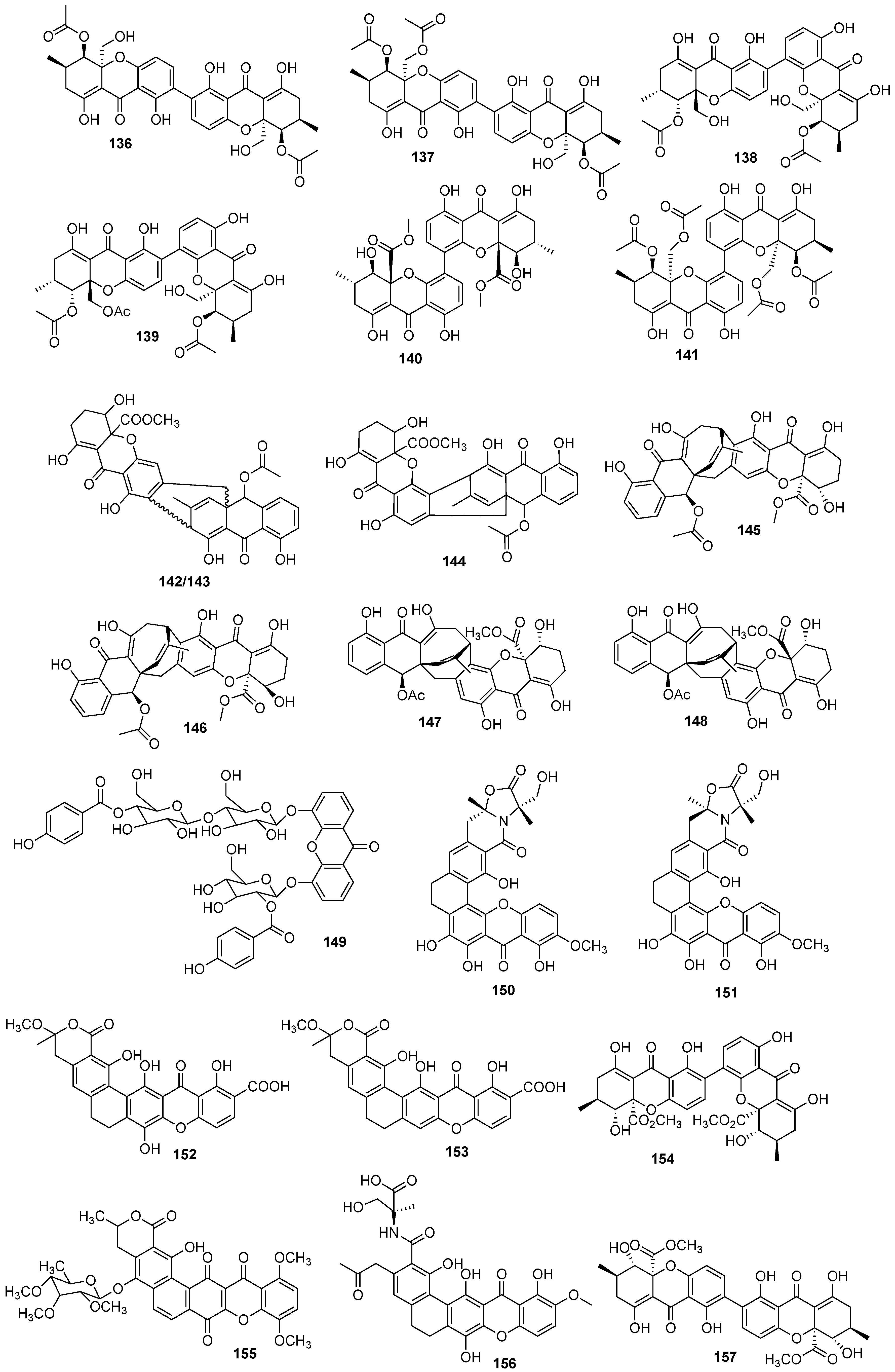



{kind=link}
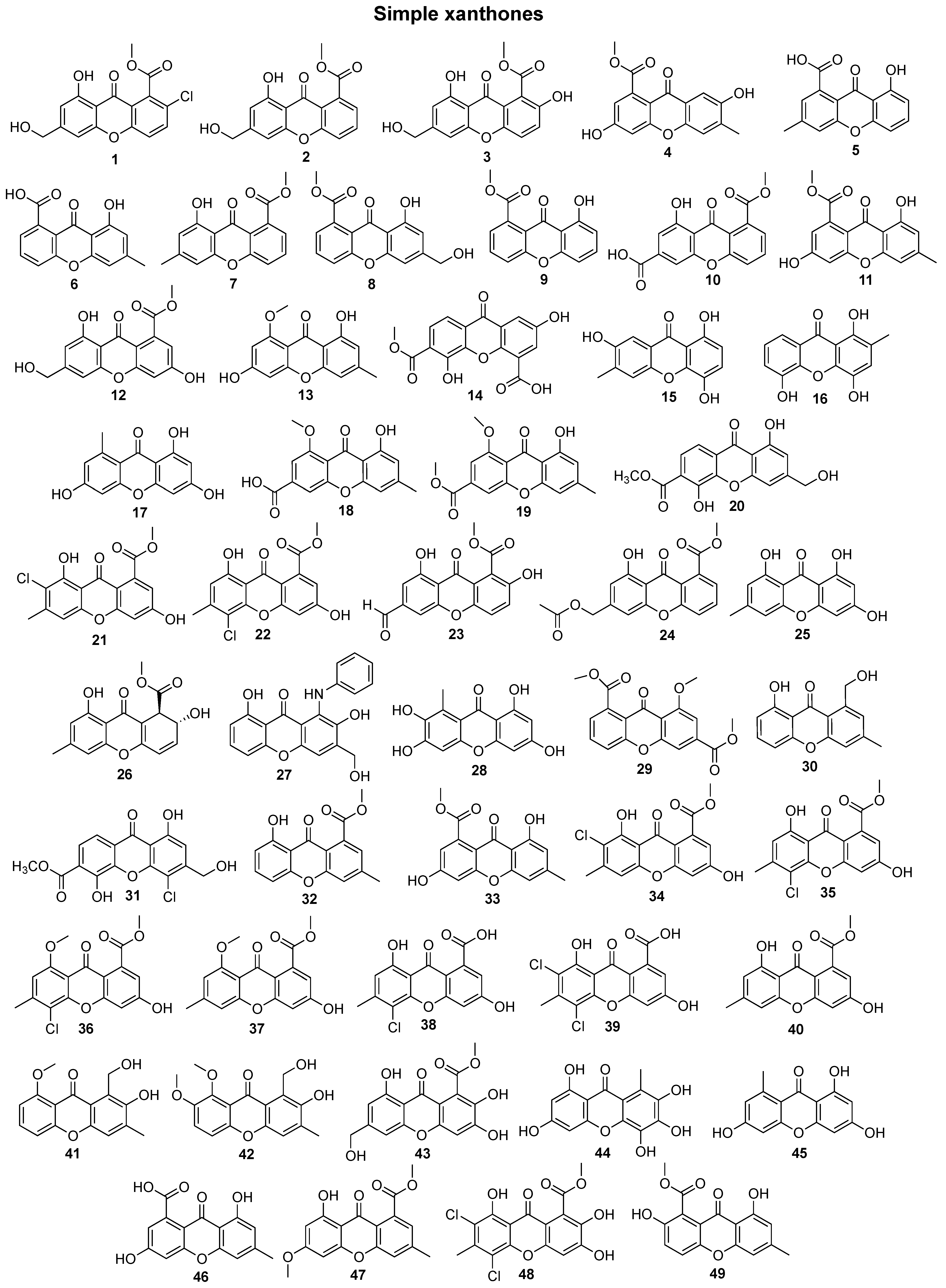{kind=link}
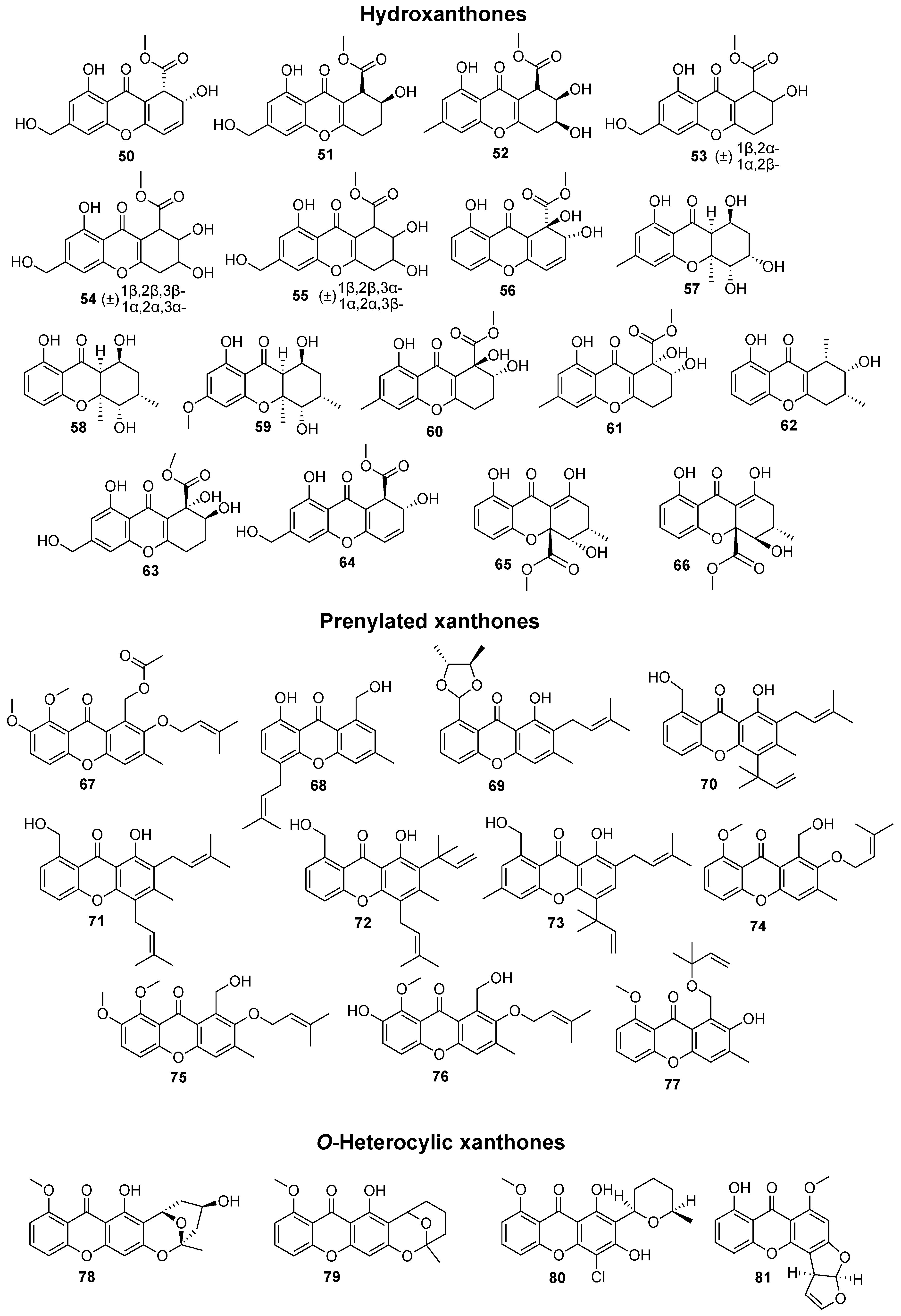{kind=link}
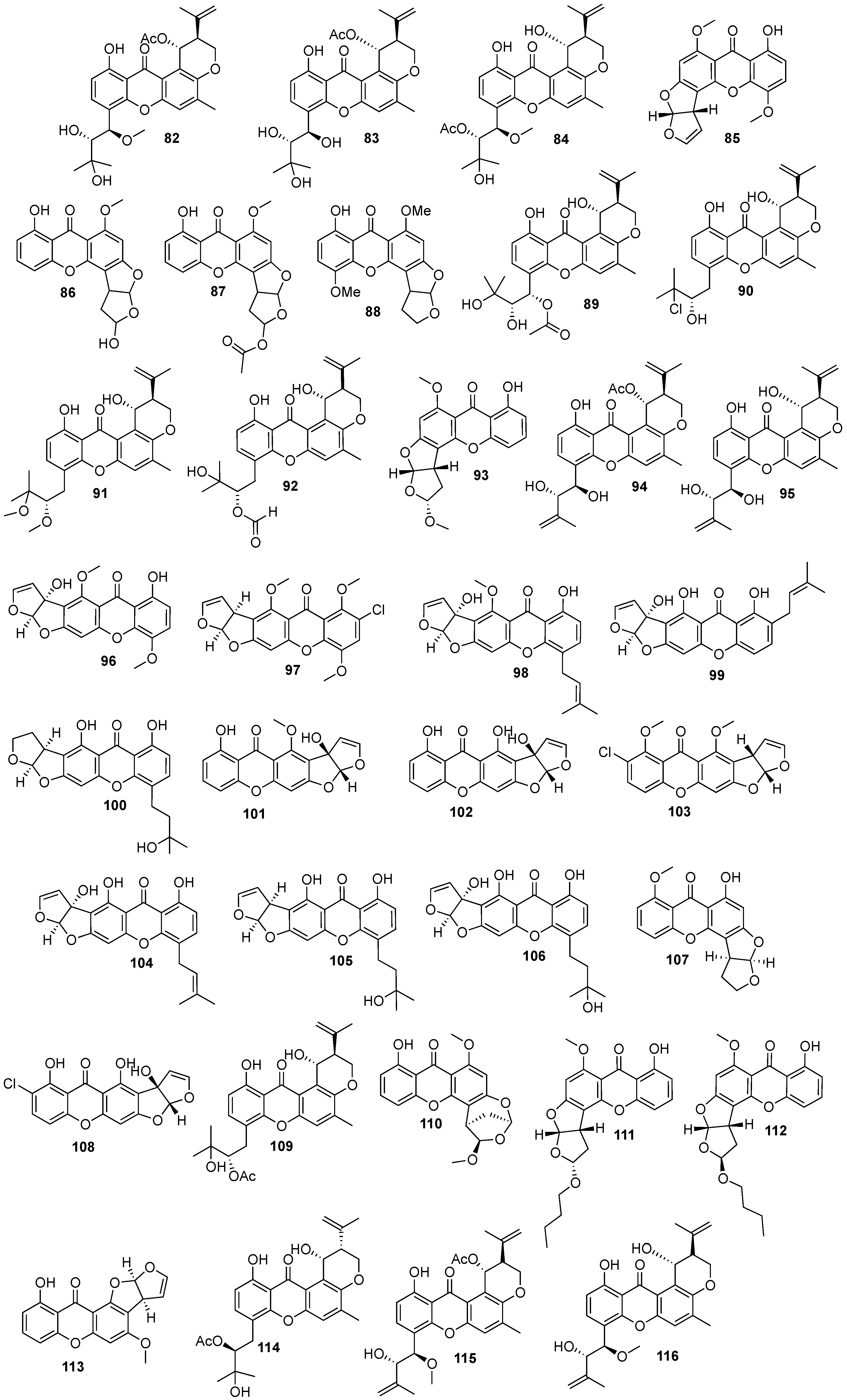{kind=link}
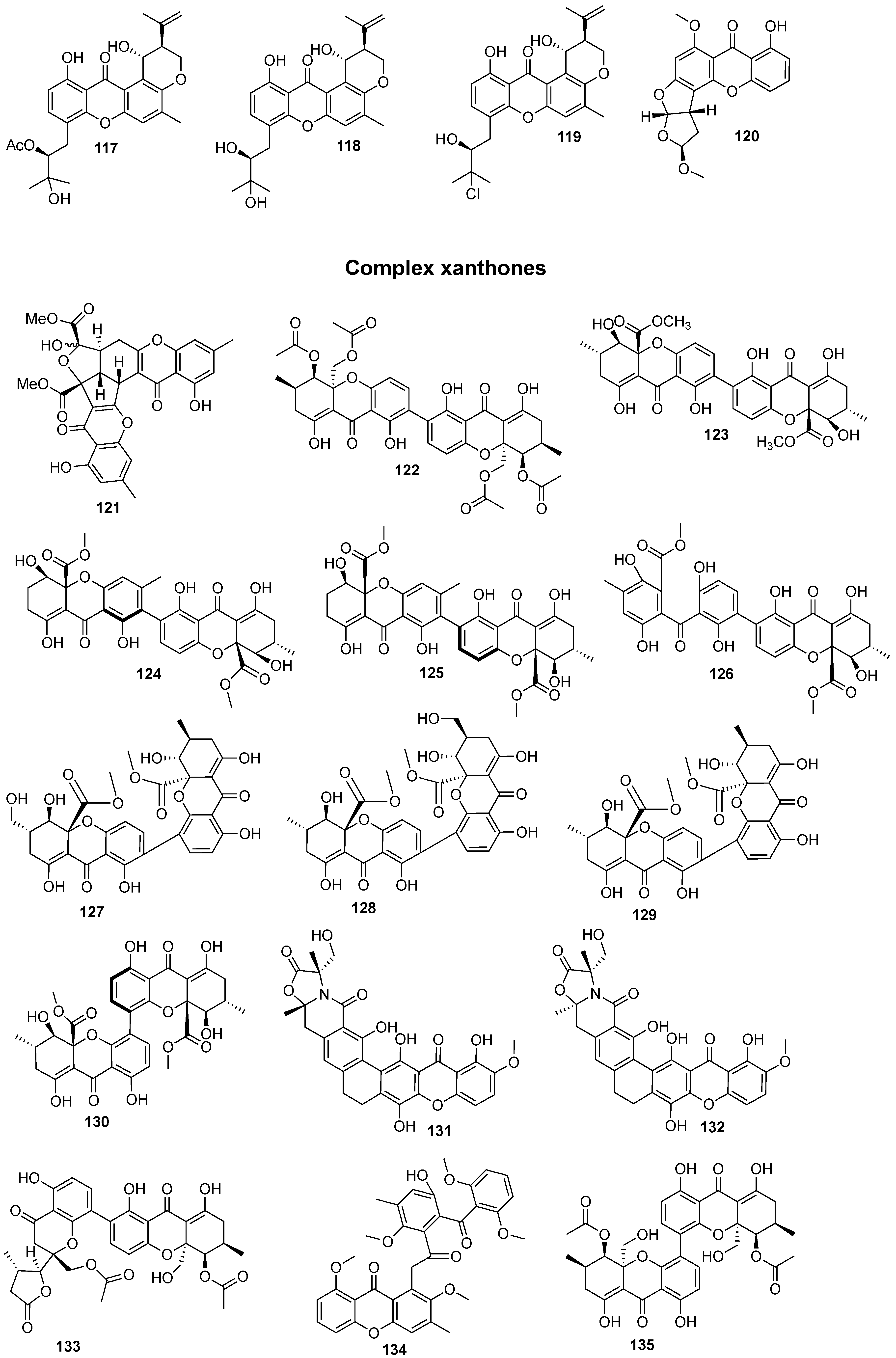{kind=link}
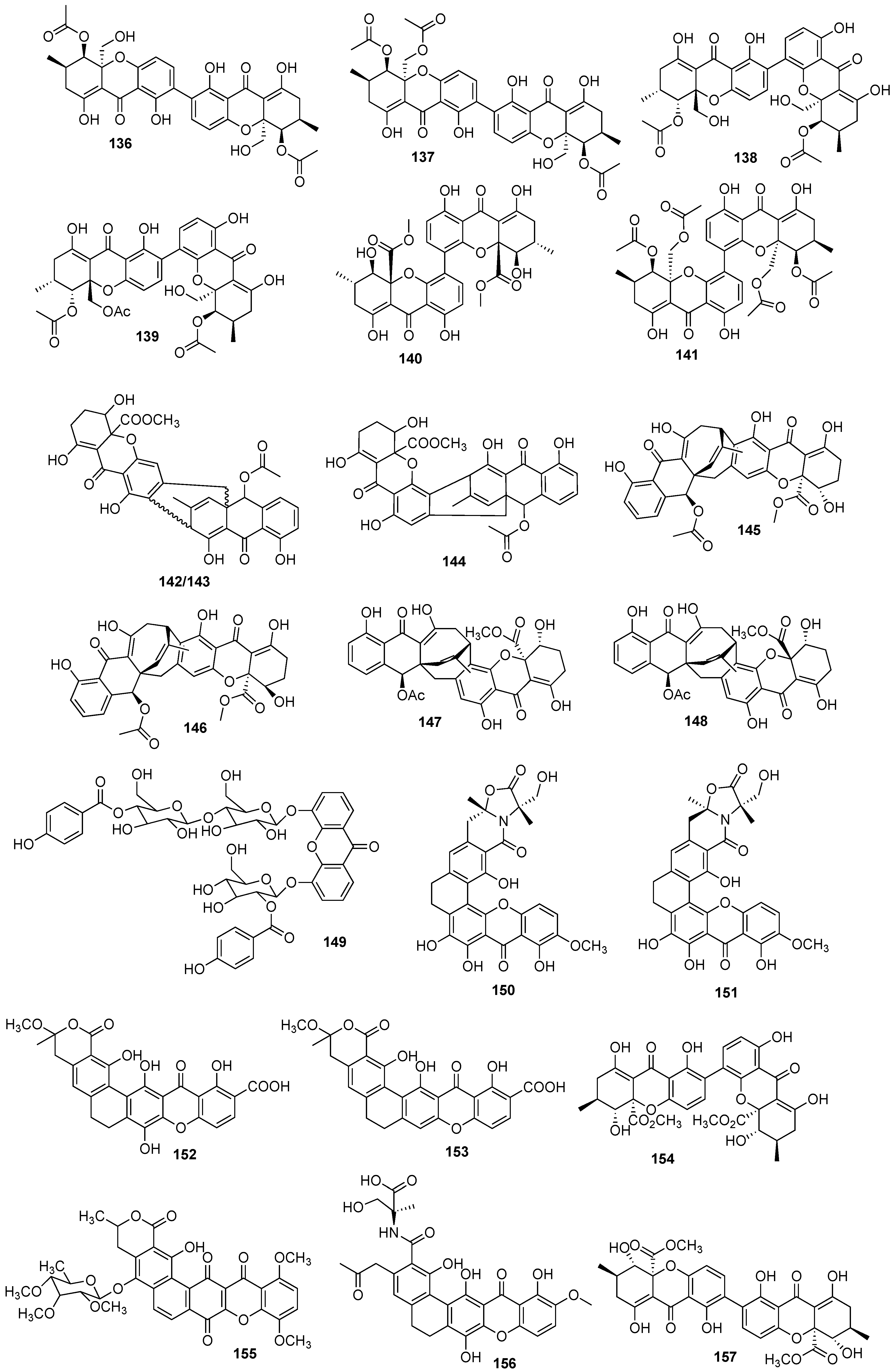{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Engyodontiumone B (1) | U937 (IC50 = 55.5 µM); Hela (IC50 = 96.1 µM); MCF-7 (IC50 = 172.3 µM); HepG2 (IC50 = 73.8 µM); Huh7 (IC50 ≥ 300 µM) | Engyodontium album (DFFSCS02) isolated from sediment collected in the South China Sea | [28] |
| Sydowinin A (2) | U937 (IC50 = 75.6 µM); Hela (IC50 ≥ 300 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| Sydowinin B (3) | U937 (IC50 = 127.0 µM); Hela (IC50 ≥ 300 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| 2,6-Dihydroxy-3-methyl-9-oxoxanthene-8-carboxylic acid methyl ester (4) | HEp-2 (IC50 = 8 µg mL−1); HepG2 (IC50 = 9 µg mL−1) | Endophytic fungus (SK7RN3G1) isolated from mangrove collected in the South China Sea | [29] |
| Monodictyxanthone (5) | Hepa-1c1c7(Cyp1A inhibition (IC50 = 34.8 ± 7.4 µM); NAD(P)H:quinone reductase induction (CD ≥ 50 (1.4) µM, IC50 ≥ 50 µM) | Monodictys putredinis isolated from the inner tissue of a green alga collected at Tenerife | [30] |
| 8-Hydroxy-6-methylxanthone-1-carboxylic acid (6) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (71.3 ± 1.2%); C4-2B (60.7 ± 5.1%); RWPE-1 (19.7 ± 4.9%) | Cladosporium halotolerans (GXIMD 02502) isolated from a coral collected in Beibu Gulf | [31] |
| Methyl 8-hydroxy-6-methyl-9-oxo-9H-xanthene-1- carboxylate (7) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (55.8 ± 3.0%); C4-2B (8.1 ± 20.6%); RWPE-1 (5.3 ± 3.1%) | ||
| Methyl 8-hydroxy-6-(hydroxymethyl)-9-oxo-9H-xanthene-1-carboxylate (8) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (68.1 ± 1.9%); C4-2B (20.2 ± 0.1%); RWPE-1 (19.0 ± 8.5%) | ||
| Vertixanthone (9) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (27.1 ± 6.9%); C4-2B (−0.1 ± 4.6%); RWPE-1 (25.0 ± 7.9%) | ||
| 8-(Methoxycarbonyl)-1-hydroxy-9-oxo-9H-xanthene-3-carboxylic acid (10) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (63.9 ± 2.2%); C4-2B (12.2 ± 5.2%); RWPE-1 (27.0 ± 5.1%) | ||
| 3,8-Dihydroxy-6-methyl-9-oxo-9H-xanthene-1-carboxylate (11) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (82.1 ± 0.9%); C4-2B (77.7 ± 0.5%); RWPE-1 (11.5 ± 1.5%) | ||
| Conioxanthone A (12) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (36.8 ± 13.3%); C4-2B (3.3 ± 11.3%); RWPE-1 (20.3 ± 9.0%) | ||
| Questin (13) | A549 (IC50 = 40.0 ± 0.3 µM); HepG2 (IC50 = 42.2 ± 0.5 µM); HeLa (IC50 = 36.2 ± 0.9 µM) | Aspergillus sydowii (C1-S01-A7) collected in the West Pacific Ocean | [32] |
| Penixanacid A (14) | HeLa (IC50 = 10.0 µM); BEL-7402 (IC50 = 30.6 µM); HEK-293 (IC50 = 28.5 µM); HCT-116 (IC50 = 19.0 µM); A-549 (IC50 = 16.9 µM) | Penicillium chrysogenum (HND11-24) isolated from a mangrove | [33] |
| Norlichexanthone (17) | K562 (IC50 = 74.6 μM); A549 (IC50 = 64.6 μM); Huh-7 (IC50 >30 μM); H1975 (IC50 = 79.1 μM); MCF-7 (IC50 = 56.7 μM); U937 (IC50 > 30 μM); BGC823 (IC50 = 697.6 μM); HL-60 (IC50 > 30 μM); MOLT-4 (IC50 = 135.4 μM); Hela (IC50 = 7.2 μM) | Stachybotry sp. (ZSDS1F1-2) isolated from a sponge collected at Xisha Island | [34] |
| Yicathin C (18) | A549 (IC50 = 37.7 ± 0.3 µM) | Aspergillus sydowii (C1-S01-A7) collected in the West Pacific Ocean | [32] |
| A375-C5 (IC50 = 48.70 ± 4.24 µM); MCF-7 (IC50 = 98.93 ± 9.83 µM); NCI-H460 (IC50 = 79.83 ± 18.45 µM) | Aspergillus wentii isolated from Gymnogongrus flabelliformis collected at Pingtan Island | [4] | |
| Yicathin B (19) | A375-C5 (IC50 = 47.70 ± 2.62 µM); MCF-7 (IC50 = 73.92 ± 2.28 µM); NCI-H460 (IC50 = 86.21 ± 2.30 µM) | ||
| 2-Hydroxy-6-formyl-vertixanthone (23) | HepG2 (IC50 = 32.7 ± 0.9 µM) | Aspergillus sydowii (C1-S01-A7) collected in the West Pacific Ocean | [32] |
| 12-O-Acetyl-sydowinin A (24) | A549 (IC50 = 25.2 ± 0.9 µM); HepG2 (IC50 = 42.3 ± 0.6 µM); HeLa (IC50 = 33.6 ± 0.7 µM) | ||
| Emodin (25) | HeLa (IC50 = 27.1 ± 0.8 µM) | ||
| Engyodontiumone H (50) | U937 (IC50 = 4.9 µM); Hela (IC50 = 24.8 µM); MCF-7 (IC50 = 38.5 µM); HepG2 (IC50 = 60.5 µM); Huh7 (IC50 = 53.3 µM) | Engyodontium album (DFFSCS021) from isolated sediment collected in the South China Sea | [28] |
| Engyodontiumone C (51) | U937 (IC50 = 218.4 µM); Hela (IC50 ≥ 300 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| Engyodontiumone D (52) | U937 (IC50 = 208.6 µM); Hela (IC50 ≥ 300 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| Engyodontiumone E (53) | U937 (IC50 = 15.9 µM); Hela (IC50 = 205.9 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| Engyodontiumone F (54) | U937 (IC50 = 192.7 µM); Hela (IC50 ≥ 300 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| Engyodontiumone G (55) | U937 (IC50 = 287.2 µM); Hela (IC50 ≥ 300 µM); MCF-7 (IC50 ≥ 300 µM); HepG2 (IC50 ≥ 300 µM); Huh7 (IC50 ≥ 300 µM) | ||
| Globosuxanthone A (56) | HCT-15 (IC50 = 10.7 µM); T-cell leukemia Jurkat cells (IC50 = 2.3 µM) | Beauveria bassiana (TPU942) isolated from a piece of an unidentified sponge collected at Iriomote Island | [35] |
| Monodictysin A (57) | Hepa-1c1c7(Cyp1A inhibition IC50 ≥ 50 µM); NAD(P)H:quinone reductase induction (CD = 191.1 µM, IC50 ≥ 400 µM) | Monodictys putredinis isolated from the inner tissue of a green alga collected at Tenerife | [30] |
| Monodictysin B (58) | Hepa-1c1c7(Cyp1A inhibition IC50 = 23.3 ± 3.9 µM); NAD(P)H:quinone reductase induction (CD = 12.0 ± 4.8 µM, IC50 ≥ 50 µM) | ||
| Monodictysin C (59) | Hepa-1c1c7(Cyp1A inhibition IC50 = 3.0 ± 0.7 µM); NAD(P)H:quinone reductase induction (CD = 12.8 ± 2.6 µM, IC50 ≥ 50 µM) | ||
| α-Diversonolic ester (60) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (28.8 ± 10.3%); C4-2B (12.9 ± 12.6%); RWPE-1 (24.3 ± 3.3%) | Cladosporium halotolerans (GXIMD 02502) isolated from a coral collected in Beibu Gulf | [31] |
| β-Diversonolic ester (61) | % Inhibitions on the cell proliferation at 10 μM: 22RV1 (40.2 ± 1.5%); C4-2B (2.8 ± 2.2%); RWPE-1 (10.3 ± 3.8%) | ||
| Penixanthone A (62) | Weak cytotoxicity against H1975, MCF-7, K562, HL7702 at concentration of 30 μM. | Penicillium sp. (SYFz-1) isolated from a mangrove sample | [36] |
| AGI-B4 (64) | K562 (IC50 = 6.97 µM); MCF-7 (IC50 = 11.7 µM); HeLa (IC50 = 1.39 µM); DU145 (IC50 = 2.69 µM); U937 (IC50 = 0.463 µM); H1975 (IC50 = 8.53 µM); SGC-7901 (IC50 = 9.43 µM); A549 (IC50 = 7.01 µM); MOLT-4 (IC50 = 5.26 µM); HL-60 (IC50 = 6.20 µM) | Aspergillus sp. (SCSIO Ind09F01) | [37] |
| U937 (IC50 = 8.8 µM); Hela (IC50 = 60.0 µM); MCF-7 (IC50 = 102.2 µM); HepG2 (IC50 = 52.7 µM); Huh7 (IC50 = 133.3 µM) | Engyodontium album (DFFSCS021) isolated from sediment collected in the South China Sea | [28] | |
| L5178Y (IC50 = 1.5 µM) | Scopulariopsis sp. isolated from solid rice cultures obtained from the Red Sea hard coral Stylophora sp. | [38] | |
| Versicone G (67) | NB4 (IC50 = 15.6 µM); HL-60 (IC50 = 21.7 µM); HeLa (IC50 = 16.9 µM) | Aspergillus versicolor (HDN11-84) isolated from mangrove | [39] |
| Paeciloxanthone (68) | HepG2 (IC50 = 1.08 µg mL−1) | Paecilomyces sp. isolated from mangrove collected in the Taiwan Strait | [40] |
| Chaetoxanthone A (78) | L6-cells (IC50 = 59.1 μg/mL) | Chaetomium sp. isolated from the Greek alga collected at Santorini Island. | [41] |
| Chaetoxanthone B (79) | L6-cells (IC50 > 90 μg/mL) | ||
| Chaetoxanthone C (80) | L6-cells (IC50 = 46.7 μg/mL) | ||
| Sterigmatocystin (81) | Bel-7402 (IC50 = 96.53 µg mL−1); NCIH-460 (IC50 = 72.52 µg mL−1) | Fungal strain (isolate 1850) isolated from a leaf of Kandelia candel collected in Hong Kong | [42] |
| A-549 (IC50 = 1.86 µg mL−1); SK-OV-3 (IC50 = 2.53 µg mL−1); SK-MEL-2 (IC50 = 1.22 µg mL−1); XF- 498 (IC50 = 2.75 µg mL−1); HCT-15 (IC50 = 4.61 µg mL−1) | Aspergillus versicolor isolated from Petrosia sp. | [43] | |
| A-549 (IC50 = 11.25 μg mL−1); SK-OV-3 (IC50 = 17.36 μg mL−1); SK-MEL-2 (IC50 = 14.33 μg mL−1); XF-498 (IC50 = 15.12 μg mL−1); HCT-15 (IC50 ≥ 30 μg mL−1) | Aspergillus versicolor | [44] | |
| Aspergixanthone A (82) | A-549 (IC50 = 1.8 µM) | Aspergillus sp. (ZA-01) isolated from sediment collected in the Bohai Sea | [45] |
| Aspergixanthone C (83) | MDA-MB-231 (IC50 = 3.3 µM); MCF-7 (IC50 = 2.8 µM); MGC-803 (IC50 = 3.6 µM); HeLa (IC50 = 2.9 µM); A-549 (IC50 = 3.2 µM) | ||
| Aspergixanthone F (84) | MDA-MB-231 (IC50 = 9.8 µM); MCF-7 (IC50 = 2.7 µM); MGC-803 (IC50 = 3.6 µM); HeLa (IC50 = 1.7 µM); A-549 (IC50 = 1.1 µM) | ||
| 5-Methoxysterigmatocystin (85) | A-549 (IC50 = 3.86 µM); HL-60 (IC50 = 5.32 µM) | Aspergillus versicolor | [46] |
| Epiremisporine B (121) | K562 (IC50 = 16.6 µM); MCF-7(IC50 = 16.3 µM); SGC7901 (IC50 = 15.8 µM) | Penicillium sp. (SCSIO Ind16F01) isolated from a deep-sea sediment collected in the Indian Ocean | [47] |
| Dicerandrol C (122) | MDA-MB-435 (IC50 = 44.10 ± 2.45 µM); HCT-116 (IC50 = 42.63 ± 2.90 µM); Calu-3 (IC50 = 36.52 ± 3.32 µM); Huh7 (IC50 ≥ 50 µM); MCF-10A (IC50 = 33.05 ± 2.74 µM) | Phomopsis sp. (HNY29-2B) isolated from Acanthus ilicifolius collected in the South China Sea | [48] |
| Secalonic acid D (123) | PANC-1 Glucose (−) (IC50 = 0.6 µM); PANC-1 Glucose (+) (IC50 ≥ 1000 µM) | Penicilliumoxalicum (16A08-1-1) isolated from a sponge collected at Pramuka Island | [49] |
| U87 MG (IC50 = 5.64 µM); NCI-H1650 (IC50 = 4.93 µM); HT29 (IC50 = 1.46 µM)A498 (IC50 = 8.88 µM); HL-60 (IC50 = 0.41 µM) | Penicillium chrysogenum (HLS111) isolated from a sponge | [50] | |
| SK- HEP (IC50 = 1.504 µM); Hela (IC50 = 1.322 µM); A549 (IC50 = 1.625 µM); SK-MES-1 (IC50 = 1.314 µM); SPC-A1 (IC50 = 1.679 µM); 95D (IC50 = 1.003 µM); Jeko-1 (IC50 = 0.915 µM); Raji (IC50 = 0.955 µM); U937 (IC50 = 1.119 µM); A375 (IC50 = 1.598 µM); HFF (IC50 = 24.1 µM); H22 (IC50 = 1.007µM) | Penicilliumoxalicum isolated from sediments collected on the southeast coastal region of China | [51] | |
| Versixanthone G (124) | HL-60 (IC50 = 13.4 µM); K562 (IC50 = 20.9 µM); A549 (IC50 = 17.8 µM); H1975 (IC50 = 9.8 µM); MGC803 (IC50 = 4.6 µM); HEK293 (IC50 ≥ 50 µM); HO-8910 (IC50 = 9.6 µM); HCT-116 (IC50 = 16.2 µM) | Aspergillus versicolor isolated from mangrove | [52] |
| Versixanthone H (125) | HL-60 (IC50 = 6.9 µM); K562 (IC50 = 22.1 µM); A549 (IC50 = 19.2 µM); H1975 (IC50 = 5.3 µM); MGC803 (IC50 = 6.2 µM); HEK293 (IC50 ≥ 50 µM); HO-8910 (IC50 = 6.9 µM); HCT-116 (IC50 = 15.2 µM) | ||
| Versixanthone I (126) | HL-60 (IC50 = 27.8 µM); K562 (IC50 ≥ 50.0 µM); A549 (IC50 ≥ 50.0 µM); H1975 (IC50 ≥ 50.0 µM); HEK293 (IC50 ≥ 50 µM); HO-8910 (IC50 ≥ 50.0 µM); HCT-116 (IC50 ≥ 50.0 µM) | ||
| Versixanthone J (127) | HL-60 (IC50 = 47.3 µM); K562 (IC50 ≥ 50.0 µM); A549 (IC50 ≥ 50.0 µM); H1975 (IC50 ≥ 50.0 µM); HEK293 (IC50 ≥ 50 µM); HO-8910 (IC50 ≥ 50.0 µM); HCT-116 (IC50 ≥ 50.0 µM) | ||
| Versixanthone K (128) | HL-60 (IC50 = 49.5 µM); K562 (IC50 ≥ 50.0 µM); A549 (IC50 ≥ 50.0µM); H1975 (IC50 = 49.5 µM); MGC803 (IC50 ≥ 50.0µM); HEK293 (IC50 ≥ 50 µM); HO-8910 (IC50 ≥ 50.0 µM); HCT-116 (IC50 ≥ 50.0 µM) | ||
| Versixanthone L (129) | HL-60 (IC50 = 0.5 µM); K562 (IC50 = 1.1 µM); A549 (IC50 = 1.6 µM); MGC803 (IC50 = 1.1 µM); HO-8910 (IC50 = 1.5 µM); HCT-116 (IC50 = 1.2 µM) | ||
| Versixanthone M (130) | HL-60 (IC50 = 0.9 µM); K562 (IC50 = 0.4 µM); A549 (IC50 = 11.7 µM); H1975 (IC50 = 3.5 µM); MGC803 (IC50 = 0.9 µM); HO-8910 (IC50 = 1.4 µM); line HCT-116 (IC50 = 0.5 µM) | ||
| Citreamicin ε A (131) | HeLa (IC50 = 0.032 ± 0.0062 µM); HepG2 (IC50 = 0.079 ± 0.031 µM) | Streptomyces caelestis collected on the coastal water of the Red Sea | [53] |
| Citreamicin ε B (132) | HeLa (IC50 = 0.031 ± 0.0081 µM); HepG2 (IC50 = 0.10 ± 0.0053 µM) | ||
| Acredinone C (133) | Inhibited the RANKL- induced formation of TRAP+-MNCs in a dose-dependent manner without any cytotoxicity up to 10 μM | Acremonium sp. isolated from the inner tissue of Suberites japonicas | [54] |
| Phomolactonexanthone A (134) | Calu-3 (IC50 = 43.45 ± 2.51 µM) | Phomopsis sp. (HNY29-2B) isolated from Acanthus ilicifolius collected in the South China Sea | [48] |
| Deacetylphomoxanthone C (135) | HCT-116 (IC50 = 44.06 ± 3.29 µM); Calu-3 (IC50 = 43.35 ± 2.09µM) | ||
| Dicerandrol A (136) | MDA-MB-435 (IC50 = 3.03 ± 0.12 µM); HCT-116 (IC50 = 2.64 ± 0.03 µM); Calu-3 (IC50 = 1.76 ± 0.02 µM); Huh7 (IC50 = 4.19 ± 0.08 µM); MCF-10A (IC50 = 28.32 ± 3.57µM) | ||
| Dicerandrol B (137) | MDA-MB-435 (IC50 = 8.65 ± 0.66 µM); HCT-116 (IC50 = 3.94 ± 0.39µM); Calu-3 (IC50 = 4.10 ± 0.08 µM); Huh7 (IC50 = 30.37 ± 1.10 µM); MCF-10A (IC50 = 8.14 ± 1.27 µM) | ||
| Deacetylphomoxanthone B (138) | MDA-MB-435 (IC50 = 14.40 ± 1.18 µM); HCT-116 (IC50 = 7.12 ± 0.70 µM); Calu-3 (IC50 = 4.14 ± 0.02 µM); Huh7 (IC50 = 29.20 ± 1.19 µM) | ||
| Penexanthone A (139) | MDA-MB-435 (IC50 = 7.90 ± 0.58µM); HCT-116 (IC50 = 6.92 ± 0.38 µM); Calu-3 (IC50 = 6.44 ± 0.86 µM); Huh7 (IC50 = 42.82 ± 3.58 µM); MCF-10A (IC50 = 16.13 ± 1.57 µM) | ||
| 4,4′-bond Secalonic acid D (140) | SK- HEP (IC50 = 1.342 µM); Hela (IC50 = 0.827 µM); A549 (IC50 = 1.353 µM); SK-MES-1 (IC50 = 0.640 µM); SPC-A1 (IC50 = 1.205 µM); 95D (IC50 = 0.978 µM); Jeko-1 (IC50 = 0.705 µM); Raji (IC50 = 0.484 µM); U937 (IC50 = 0.960 µM); A375 (IC50 = 1.085 µM); HFF (IC50 = 26.6 µM); H22 (IC50 = 1.211 µM) | Penicilliumoxalicum isolated from sediments collected on the southeast coastal region of China | [51] |
| Phomoxanthone A (141) | HL-60 (cytotoxic at 0.1 to 0.01 μg mL−1) | Phomopsis longicolla isolated from Bostrychia radicans | [55] |
| JBIR-97 (142/143) | HeLa (IC50 = 11 µM); ACC-MESO-1 (IC50 = 31 µM) | Tritirachium sp. (SpB081112MEf2) isolated from Pseudoceratina purpurea collected at Ishigaki Island | [56] |
| JBIR-98 (142/143) | HeLa (IC50 = 17 µM); ACC-MESO-1 (IC50 = 63 µM) | ||
| JBIR-99 (144) | HeLa (IC50 = 17 µM); ACC-MESO-1 (IC50 = 59 µM) | ||
| Buanmycin (156) | A549 (IC50 = 1.7 µM); HCT116 (IC50 = 0.9 µM); SNU638 (IC50 = 0.8 µM); SK-HEP1 (IC50 = 1.9 µM); MDA-MB231 (IC50 = 1.2 µM) | Streptomyces sp. isolated from a tidal mudflat collected in Buan | [57] |
| A549 (IC50 = 0.8 µM); HeLa (IC50 = 0.9 µM) | Streptomyces sp. (HGMA004) isolated from a mudflat collected at Uki | [58] | |
| Chrysoxanthone A (162) | U87 MG (IC50 = 22.6 µM); NCI-H1650 (IC50 = 42.2 µM); HT29 (IC50 = 41.8 µM); A498 (IC50 = 28.5 µM); HL-60 (IC50 = 37.2 µM) | Penicillium chrysogenum (HLS111) isolated from a sponge | [50] |
| Chrysoxanthone B (163) | U87 MG (IC50 ≥ 50 µM); NCI-H1650 (IC50 ≥ 50 µM); HT29 (IC50 = 30.8 µM); A498 (IC50 ≥ 50 µM); HL-60 (IC50 = 16.2 µM) | ||
| Chrysoxanthone C (164) | U87 MG (IC50 = 47.0 µM); NCI-H1650 (IC50 ≥ 50 µM); HT29 (IC50 = 43.2 µM); A498 (IC50 ≥ 50 µM); HL-60 (IC50 = 22.7 µM) | ||
| Ukixanthomycin A (165) | A549 (IC50 ≥ 200 µM); HeLa (IC50 ≥ 200 µM) | Streptomyces sp. (HGMA004) isolated from a mudflat collected at Uki | [58] |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Sydowinin B (3) | V. rotiferianus (MCCC E385) (MIC = 32.6 ± 1.1 µg mL−1) | Aspergillus sydowii (C1-S01-A7) collected in the West Pacific Ocean | [32] |
| 1,4,7-Trihydroxy-6-methylxanthone (15) | E. coli (MIC = 32 μg mL−1); P. aeruginosa (MIC = 32 μg mL−1); S. aureus (MIC > 64 μg mL−1); V. alginolyticus (MIC = 32 μg mL−1); V. harveyi (MIC = 32 μg mL−1); V. parahaemolyticus (MIC = 32 μg mL−1) | Talaromyces islandicus (EN-501) isolated from Laurencia okamurai | [59] |
| 1,4,5-Trihydroxy-2-methylxanthone (16) | E. coli (MIC = 4 μg mL−1); P. aeruginosa (MIC = 4 μg mL−1); S. aureus (MIC = 8 μg mL−1); V. alginolyticus (MIC = 4 μg mL−1); V. harveyi (MIC = 8 μg mL−1); V. parahaemolyticus (MIC = 4 μg mL−1) | ||
| Norlichexanthone (17) | S. aureus (ATCC 27154) (MIC = 12.5 µg mL−1); E. coli (ATCC 25922) (MIC > 100 µg mL−1); S. ventriculi (ATCC 29068) (MIC = 25.0 µg mL−1); P. aeruginosa (ATCC 25668) (MIC = 25.0 µg mL−1) | Talaromyces sp. (ZH-154) collected in the South China Sea | [60] |
| Yicathin C (18) | E. coli (zone of inhibition 12.0 mm); S. aureus (zone of inhibition 7.5 mm) | Aspergillus wentii isolated from Gymnogongrus flabelliformis collected at Pingtan Island | [61] |
| Yicathin B (19) | E. coli (zone of inhibition 9 mm) | ||
| Fischexanthone (20) | E. coli (MIC > 1265.82 µM); S. aureus (MIC > 1265.82 µM) | Alternaria sp. (R6) isolated from mangrove collected at Leizhou peninsula | [62] |
| Methyl (2-chloro-l,6-dihydroxy-3-methylxanthone)-8-carboxylate (21) | S. aureus (ATCC43300) (MIC = 6.25 μg mL−1); S. aureus (ATCC29213) (MIC = 6.25 μg mL−1); S. aureus (ATCC33591) (MIC = 3.13 μg mL−1); S. aureus (ATCC25923) (MIC = 3.13 μg mL−1); E. faecalis (ATCC51299) (MIC = >100 μg mL−1); E. faecium (ATCC35667) (MIC = >100 μg mL−1); V. parahaemolyticus (ATCC17802) (MIC > 100 μg mL−1) | Aspergillus flavipes (DL-11) isolated from coastal sediment collected in Dalian | [63] |
| Methyl (4- chloro-l,6-dihydroxy-3-methylxanthone)-8-carboxylate (22) | S. aureus (ATCC43300) (MIC = 3.13 μg mL−1); S. aureus (ATCC29213) (MIC = 3.13 μg mL−1); S. aureus (ATCC33591) (MIC = 1.56 μg mL−1); S. aureus (ATCC25923) (MIC = 3.13 μg mL−1); E. faecalis (ATCC51299) (MIC 25 μg mL−1); E. faecium (ATCC35667) (MIC 50 μg mL−1); V. parahaemolyticus (ATCC17802) (MIC > 100 μg mL−1) | ||
| 2-Hydroxy-6-formyl-vertixanthone (23) | MRSA (ATCC 43300) (MIC = 16.3 ± 0.9 µg mL−1); MRSA (CGMCC 1.12409) (MIC = 16.1 ± 0.5 µg mL−1) | Aspergillus sydowii (C1-S01-A7) collected in the West Pacific Ocean | [32] |
| 12-O-Acetyl-sydowinin A (24) | MRSA (ATCC 43300) (MIC = 32.6 ± 0.8 µg mL−1); MRSA (CGMCC 1.12409) (MIC = 31.8 ± 0.8 µg mL−1) | ||
| Emodin (25) | V. vulnificus (MCCC E1758) (MIC = 16.1 ± 0.7 µg mL−1); MRSA (ATCC 43300) (MIC = 15.4 ± 0.3 µg mL−1); MRSA (CGMCC 1.12409) (MIC = 15.7 ± 0.5 µg mL−1) | ||
| Aspergillusone A (26) | MRSA (ATCC 43300) (MIC = 32.2 ± 0.3 µg mL−1); MRSA (CGMCC 1.12409) (MIC = 32.4 ± 0.1 µg mL−1) | ||
| BCG (M. bovis Pasteur 1173P2) (MIC = 20 µg mL−1) | Aspergillus versicolor (MF160003) | [64] | |
| Chalaniline B (27) | Percent (%) growth of treated bacteria: B. subtilis (ATCC 49343) (67 ± 17%); S. aureus (ATCC 25923) (64 ± 14%); MRSA (ATCC BAA-41) (57 ± 8%); MRSA (ATCC BAA-44) (40 ± 2%) | Endophytic ascomycete with Chalara sp. (6661) | [65] |
| Engyodontiumone H (50) | E. coli (zone of inhibition 13.8 mm); B. subtilis (zone of inhibition 16.5 mm) | Engyodontium album (DFFSCS02) isolated from a sediment collected in the South China Sea | [28] |
| E. coli (MIC = 64 μg mL−1); B. subtilis (MIC = 32 μg mL−1) | |||
| Aspergillusone B (63) | E. coli (zone of inhibition 11.0 mm); B. subtilis (zone of inhibition 14.4 mm) | Engyodontium album (DFFSCS02) isolated from a sediment collected in the South China Sea | |
| E. coli (MIC = 64 μg mL−1); B. subtilis (MIC = 64 μg mL−1) | |||
| AGI-B4 (64) | E. coli (zone of inhibition 15.8 mm); B. subtilis (zone of inhibition 17.5 mm) | Engyodontium album (DFFSCS02) isolated from sediment collected in the South China Sea | |
| E. coli (MIC = 64 μg mL−1); B. subtilis (MIC = 64 μg mL−1) | Engyodontium album (DFFSCS02) isolated from sediment collected in the South China Sea | ||
| V. vulnificus (MCCC E1758) (MIC = 32.5 ± 0.4 µg mL−1); MRSA (ATCC 43300) (MIC = 32.9 ± 0.3 µg mL−1); MRSA (CGMCC 1.12409) (MIC = 16.3 ± 0.5 µg mL−1) | Deep sea-derived fungus Aspergillus sydowii C1-S01-A7 isolated in the West Pacific Ocean | [32] | |
| Blennolide A (65) | E. coli (zone of inhibition 7 mm); B. megaterium (zone of inhibition 8 mm) | Blennoria sp. isolated from Carpobrotus edulis collected at Gomera | [66] |
| Blennolide B (66) | E. coli (zone of inhibition 8 mm); B. megaterium (zone of inhibition 8 mm) | ||
| Paeciloxanthone (68) | E. coli (zone of inhibition 12 mm) | Paecilomyces sp. isolated from a mangrove collected in the Taiwan Strait | [40] |
| Stergimatocystin (81) | S. aureus (zone of inhibition 9.0 mm) | Aspergillus versicolor | [44] |
| Hemi-acetal sterigmatocystin (86) | S. aureus (ATCC 6538) (MIC > 100 μg mL−1); B. subtilis (ATCC 6633) (MIC > 100 μg mL−1); MRSA (MIC >100 μg mL−1); P. aeruginosa (ATCC 15692) (MIC > 100 μg mL−1) | Aspergillus versicolor (MF359) isolated from Hymeniacidon perleve collected in the Bohai Sea | [67] |
| Acyl-hemiacetal sterigmatocystin (87) | S. aureus (ATCC 6538) (MIC > 100 μg mL−1); B. subtilis (ATCC 6633) (MIC > 100 μg mL−1); MRSA (MIC > 100 μg/mL); P. aeruginosa (ATCC 15692) (MIC > 100 μg mL−1) | ||
| 5-Methoxydihydrosterigmatocystin (88) | S. aureus (ATCC 6538) (MIC = 12.5 μg mL−1); B. subtilis (ATCC 6633) (MIC = 3.125 μg mL−1); MRSA (MIC > 100 μg mL−1); P. aeruginosa (ATCC 15692) (MIC > 100 μg mL−1) | ||
| Emerixanthone E (89) | E. coli (ATCC 29922); K. pneumoniae (ATCC 13883); S. aureus (ATCC 29213), E. faecalis (ATCC 29212); A. baumannii (ATCC 19606); A. hydrophila (ATCC 7966): Diameters of the inhibition zones ranged between 9 and 11 mm | Emericella sp. collected in the South China Sea | [68] |
| Emerixanthone A (90) | E. coli (ATCC 29922); K. pneumoniae (ATCC 13883); S. aureus (ATCC 29213); E. faecalis (ATCC 29212); A. baumannii (ATCC 19606); A. hydrophila (ATCC 7966): Diameters of inhibition zones were all 4–6 mm | Emericella sp. (SCSIO 05240) collected in the South China Sea | [69] |
| Emerixanthone C (91) | E. coli (ATCC 29922); K. pneumoniae (ATCC 13883); S. aureus (ATCC 29213); E. faecalis (ATCC 29212); A. baumannii (ATCC 19606); A. hydrophila (ATCC 7966): Diameters of inhibition zones were all 4–6 mm | ||
| Varixanthone (92) | E. coli (MIC = 12.5 µg mL−1); Proteus sp. (MIC = 12.5 µg mL−1); B. subtilis (MIC = 12.5 µg mL−1); S. aureus (MIC = 12.5 µg mL−1); E. faecalis (MIC = 50 µg mL−1) | Emericella variecolor (M75-2) was isolated from a Porifera sp. collected in the Caribbean Sea | [70] |
| Oxisterigmatocystin C (93) | S. aureus (ATCC25923) (MIC < 48 μg mL−1) | Aspergillus sp. (F40) isolated from Callyspongia sp. | [71] |
| Aspergixanthone G (94) | M. luteus (MIC = 0.78 µg mL−1); B. anthracis (MIC = 12.5 µg mL−1); S. typhi (MIC = 6.13 µg mL−1); E. aerogenes (MIC = 6.13 µg mL−1) | Aspergillus sp. (ZA-01) isolated from sediment collected in the Bohai Sea | [45] |
| Aspergixanthone H (95) | M. luteus (MIC = 6.13 µg mL−1); B. anthracis (MIC = 12.5 µg mL−1); S. typhi (MIC = 6.13 µg mL−1); E. aerogenes (MIC = 6.13 µg mL−1) | ||
| Dicerandrol C (122) | S. aureus (ATCC 6538) (MIC = 1.33 µM); S. saprophyticus (ATCC 15305) (MIC = 2.66 µM) | Phomopsis longicolla isolated from Bostrychia radicans collected in Brazil | [72] |
| Secalonic acid D (123) | S. aureus (ATCC 29,213) (IC50 = 7.19 μM); M. tuberculosis (IC50 = 1.26 μM) | Aspergillus sp. (SCSIO XWS03F03) isolated from a sponge | [73] |
| B. subtilis (MIC = 24.4 µg mL−1); E. coli J(VC1228) (MIC = 24.4 µg mL−1); M. luteus (UST950701-006) (MIC = 24.4 µg mL−1); P. nigrifaciens (UST010620-005) (MIC = 97.5 µg mL−1) | Penicillium sp. (SCSGAF0023) isolated from Dichotella gemmacea collected in the South China Sea | [74] | |
| JBIR-97/98 (145) | S. epidermidis (IC50 = 0.20 ± 0.04 μM); MRSA (IC50 = 0.19 ± 0.02 μM); P. acnes (IC50 = 11.0 ± 1.3 μM) | Engyodontium album isolated from Cacospinga scalaris collected at the Limski Fjord | [75] |
| Engyodontochone A (146) | S. epidermidis (IC50 = 0.19 ± 0.04 μM); MRSA (IC50 = 0.17 ± 0.02 μM); P. acnes (IC50 = 13.8 ± 1.7 μM) | ||
| JBIR-99 (147) | S. epidermidis (IC50 = 0.21 ± 0.09 μM); MRSA (IC50 = 0.25 ± 0.07μM); P. acnes (IC50 = 14.1 ± 2.7 μM) | ||
| Engyodontochone B (148) | S. epidermidis (IC50 = 0.22 ± 0.03 μM); MRSA (IC50 = 0.24 ± 0.04 μM); P. acnes (IC50 = 11.7 ± 2.4 μM) | ||
| Microluside A (149) | E. faecalis (JH212) (MIC = 10 μM); S. aureus (NCTC 8325) (MIC = 13 μM) | Micrococcus sp. (EG45) isolated from Spheciospongia vagabunda collected in the Red Sea | [76] |
| Citreamicin θ A (150) | S. haemolyticus (MIC = 0.5 μg mL−1); S. aureus (UST950701-005) (MIC = 1.0 μg mL−1); B. subtillis (769) (MIC = 0.25 μg mL−1); S. aureus (ATCC43300) (MIC = 0.25 μg mL−1) | Streptomyces caelestis collected in the Red Sea | [77] |
| Citreamicin θ B (151) | S. haemolyticus (UST950701-004) (MIC = 0.5 μg mL−1); S. aureus (UST950701-005) (MIC = 1.0 μg mL−1); B. subtilis (769) (MIC = 0.25 μg mL−1); S. aureus (ATCC43300) (MIC = 0.25 μg mL−1) | ||
| Citreaglycon A (152) | S. haemolyticus (MIC = 8.0 μg mL−1); S. aureus (UST950701-005) (MIC = 16 μg mL−1); B. subtilis (769) (MIC = 8.0 μg mL−1); S. aureus (ATCC43300) (MIC = 8.0 μg mL−1) | ||
| Dehydrocitreaglycon A (153) | S. haemolyticus (UST950701-004) (MIC = 8.0 μg mL−1); S. aureus (UST950701-005) (MIC = 16 μg mL−1); B. subtilis (769) (MIC = 8.0 μg mL−1) | ||
| Penicillixanthone A (154) | B. subtilis (MIC = 24.4 µg mL−1); E. coli (JVC1228) (MIC = 24.4 µg mL−1); M. luteus (UST950701- 006) (MIC = 24.4 µg mL−1); P. nigrifaciens (UST010620-005) (MIC = 97.5 µg mL−1) | Penicillium sp. (SCSGAF0023) isolated from Dichotella gemmacea collected in the South China Sea | [74] |
| IB-00208 (155) | E. coli (ATCC 10536) (MIC ≥ 150 nM); K. pneumonie (ATCC 29665) (MIC ≥ 150 nM); P. aeruginosa (ATCC 10145) (MIC ≥ 150 nM); B. subtilis (ATCC 6051) (MIC = 1.4 nM); S. aureus (ATCC 6538P) (MIC = 1.4 nM); M. luteus (ATCC 9341) (MIC = 0.09 nM) | Actinomadura sp. collected at the northern coast of Spain | [78] |
| Buanmycin (156) | S. aureus (MIC = 10.5 μM, sortase A inhibition IC50 = 43.2 μM); B. subtilis (MIC = 0.7 μM); K. rhizophila (MIC = 10.5 μM); S. enterica (MIC = 0.7 μM); P. hauseri (MIC = 21.1 μM) | Streptomyces sp. isolated from a tidal mudflat collected in Buan | [57] |
| B. cereus (IC50 = 3.0 µM); E. coli (IC50 = 6.0 µM) | Streptomyces sp. (HGMA004) isolated from a mudflat collected at Uki | [58] | |
| Secalonic acid A (157) | S. aureus (ATCC 27154) (MIC = 12.5 μg mL−1); E. coli (ATCC 25922) (MIC = 25 μg mL−1); S. ventriculi (ATCC 29068) (MIC = 12.5 μg mL−1); P. aeruginosa (ATCC 25668) (MIC = 12.5 μg mL−1) | Talaromyces sp. (ZH-154) collected in the South China Sea | [60] |
| Secalonic acid B (158) | B. subtilis (MIC = 97.5 µg mL−1); E. coli (JVC1228) (MIC = 97.5 µg mL−1); M. luteus (UST950701-006) (MIC = 97.5 µg mL−1); P. nigrifaciens (UST010620-005) (MIC = 390.5 µg mL−1) | Penicillium sp. (SCSGAF0023) isolated from Dichotella gemmacea collected in the South China Sea | [74] |
| B. megaterium (zone of inhibition 15 mm) | Blennoria sp. isolated from Carpobrotus edulis collected at Gomera | [66] | |
| Neocitreamicin I (159) | B. subtilis 1A1 (MIC = 0.06 μg mL−1); S. aureus (MRSA NRS1) (MIC = 0.50 μg mL−1); S. aureus (MRSA NRS2) (MIC = 0.12 μg mL−1); S. aureus (MRSA NRS71) (MIC = 0.12 μg mL−1); E. faecalis (VRE 51299) (MIC = 0.06 μg mL−1); E. faecalis (VRE 51575) (MIC = 0.12 μg mL−1); E. coli K-12 (MIC ≥ 8.0 μg mL−1) | Nocardia sp. (G0655) isolated from a sandy soil sample collected in Falmouth | [79] |
| Neocitreamicin II (160) | B. subtilis 1A1 (MIC = 0.12 μg mL−1); S. aureus (MRSA NRS1) (MIC = 1.0 μg mL−1); S. aureus (MRSA NRS2) (MIC = 0.50 μg mL−1); S. aureus (MRSA NRS71) (MIC = 0.50 μg mL−1); E. faecalis (VRE 51299) (MIC = 0.06 μg mL−1); E. faecalis (VRE 51575) (MIC = 0.25 μg mL−1); E. coli K-12 (MIC ≥ 8.0 μg mL−1) | ||
| Citreamicin α (161) | E. coli (MIC > 128 μg mL−1); K. pneumoniae (MIC > 128 μg mL−1); Serratia sp. (MIC > 128 μg mL−1); Citrobacter sp. (MIC > 128 μg mL−1); P. aeruginosa (MIC ≥ 128 μg mL−1); S. aureus (MIC < 0.06~0.12 μg mL−1); S. epidermidis (MIC < 0.06 μg mL−1); Enterococcus sp. (MIC < 0.06~0.12 μg mL−1); Streptococcus sp. (MIC < 0.06 μg mL−1); S. pneumoniae (MIC < 0.06 μg mL−1); B. fragilis (MIC = 16 μg mL−1); B. thetaiotaomicron (MIC = 4 μg mL−1); Clostridium perfringens (MIC < 0.06 μg mL−1); C. difficile (MIC < 0.06 μg mL−1) | Culture LL-E19085 was isolated from a soil sample collected at Lake Manyara | [80] |
| Chrysoxanthone A (162) | B. subtilis (ATCC 63501) (MIC = 5 µg mL−1); E. coli (ATCC 25922) (MIC > 100 µg mL−1) | Penicillium chrysogenum (HLS111) isolated from a sponge | [50] |
| Chrysoxanthone B (163) | S. epidermidis (ATCC 12228, MSSE) (MIC = 10 µg mL−1); S. aureus (ATCC 29213, MSSA) (MIC = 20 µg mL−1); B. subtilis (ATCC 63501) (MIC = 5 µg mL−1); E. faecalis (ATCC 29212, VSE) (MIC ≥ 100 µg mL−1); E. coli (ATCC 25922) (MIC ≥ 100 µg mL−1) | ||
| Chrysoxanthone C (164) | S. epidermidis (ATCC 12228, MSSE) (MIC = 20 µg mL−1); S. aureus (ATCC 29213, MSSA) (MIC = 80 µg mL−1); B. subtilis (ATCC 63501) (MIC = 10 µg mL−1); E. faecalis (ATCC 29212, VSE) (MIC > 100 µg mL−1); E. coli (ATCC 25922) (MIC > 100 µg mL−1) | ||
| Ukixanthomycin A (165) | B. cereus (IC50 > 200 µM); E. coli (IC50 > 200 µM) | Streptomyces sp. (HGMA004) isolated from a mudflat collected at Uki | [58] |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Norlichexanthone (17) | B. megaterium (zone of inhibition 1 mm) | Enteromorpha sp. collected at Fehmarn Island | [81] |
| C. albicans (ATCC 10231) (MIC = 6.25 μg mL−1); A. niger (ATCC 13496) (MIC = 25.0 μg mL−1); F. oxysporum f. sp. cubense (MIC = 50.0 μg mL−1) | Talaromyces sp. (ZH-154) collected in the South China Sea | [60] | |
| Yicathin C (18) | C. lagenarium (zone of inhibition 11.0 mm) | Aspergillus wentii isolated from Gymnogongrus flabelliformis collected at Pingtan Island | [61] |
| Fischexanthone (20) | F. graminearum (MIC = 474.68 µM); C. musae (MIC = 474.68 µM) | Alternaria sp. (R6) isolated from mangrove collected at Leizhou peninsula | [62] |
| 2,3,6,8-Tetrahydroxy-1-methylxanthone (28) | M. violaceum (zone of inhibition 1 mm) | Enteromorpha sp. collected at Fehmarn Island | [81] |
| Dimethyl 8-methoxy-9-oxo- 9H-xanthene-1, 6-dicarboxylate (29) | F. oxysporum f. sp. cubense (MIC = 12.5 µg mL−1) | Penicillium sp. ZZF 32# collected in the South China Sea | [82,83] |
| 1-Hydroxy-6-methyl-8-(hydroxymethyl)xanthone (30) | E. repens (zone of inhibition 2 mm) U. violacea (zone of inhibition 2 mm) | Ulocladium botrytis (193A4) isolated from the Callyspongia vaginalis collected at Dominica | [84] |
| 4-Chlorofischexanthone (31) | F. graminearum (MIC = 107 µM) C. musae (MIC = 214 µM) | Alternaria sp. (R6) isolated from mangrove collected at Leizhou peninsula | [62] |
| 8-Hydroxy-3-methyl-9-oxo-9H-xanthene-1-carboxylic acid methyl ether (32) | G. musae (Rate of inhibition 53%); P. cichoralearum (Rate of inhibition 48%); C. glocosporioides (Rate of inhibition 28%); B. graminearum (Rate of inhibition 4.6%); F. oxysporum (Rate of inhibition 9.5%) | Co-culture broth of mangrove fungi (strain No. K38 and E33) collected in the South China Sea | [85,86] |
| Globosuxanthone A (56) | C. albicans IFM 4954 (zone of inhibition 7 mm) | Beauveria bassiana (TPU942) isolated from a sponge collected at Iriomote Island | [35] |
| Blennolide A (65) | M. violaceum (zone of inhibition 9 mm) | Blennoria sp. isolated from Carpobrotus edulis collected at Gomera | [66] |
| Blennolide B (66) | M. violaceum (zone of inhibition 8 mm) | ||
| Paeciloxanthone (68) | C. lunata (zone of inhibition 6 mm); C. albicans (zone of inhibition 10 mm) | Paecilomyces sp. isolated from a mangrove collected in the Taiwan Strait | [40] |
| Versicone A (74) | C. acutatum (MIC = 32 μg mL−1); F. oxysporum (MIC = 128 μg mL−1); M. oryzae (MIC > 200 μg mL−1) | Aspergillus versicolor (SCSIO 05879) collected in the Indian Ocean | [87] |
| Versicone B (75) | C. acutatum (MIC > 200 μg mL−1); F. oxysporum (MIC > 200 μg mL−1); M. oryzae (MIC > 200 μg mL−1) | ||
| Versicone C (76) | C. acutatum (MIC > 200 μg mL−1); F. oxysporum (MIC > 200 μg mL−1); M. oryzae (MIC > 200 μg mL−1) | ||
| Versicone D (77) | C. acutatum (MIC > 200 μg mL−1); F. oxysporum (MIC > 200 μg mL−1); M. oryzae (MIC > 200 μg mL−1) | ||
| Emerixanthone D (109) | Fusarium sp., Penicillium sp., A. niger, R. solani, F. oxysporium f. sp. niveum, F. oxysporum f. sp. cucumeris: Diameters of inhibition zones of which were both 3–4 mm | Emericella sp. (SCSIO 05240) collected in the South China Sea | [69] |
| Buanmycin (156) | C. albicans (MIC = 21.1 μM); A. fumigatus (MIC = 84.3 μM) | Streptomyces sp. isolated from a tidal mudflat collected in Buan | [57] |
| C. albicans (IC50 = 0.4 μM) | Streptomyces sp. (HGMA004) isolated from a mudflat collected at Uki | [58] | |
| Secalonic acid A (157) | C. albicans (ATCC 10231) (MIC = 6.25 μg mL−1); A. niger (ATCC 13496) (MIC = 6.25 μg mL−1); F. oxysporum f. sp. cubense (MIC = 12.5 μg mL−1) | Talaromyces sp. (ZH-154) collected in the South China Sea | [60] |
| Secalonic acid B (158) | M. violaceum (zone of inhibition 13 mm) | Blennoria sp. isolated from Carpobrotus edulis collected at Gomera | [66] |
| Ukixanthomycin A (165) | C. albicans (IC50 = 11.5 µM) | Streptomyces sp. (HGMA004) isolated from a mudflat collected at Uki | [58] |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Norlichexanthone (17) | EV71 (IC50 = 40.3 μM) | Stachybotry sp. (ZSDS1F1-2) isolated from a sponge collected at Xisha Island | [34] |
| HIV-1-RT (82.9% inhibition at 66 μg mL−1) | Enteromorpha sp. collected at Fehmarn Island | [81] | |
| 2,3,6,8-Tetrahydroxy-1-methylxanthone (28) | HIV-1-RT (82.2% inhibition at 66 μg mL−1) | ||
| 3,8-Dihydroxy-6-methyl-9- oxo-9H-xanthene-1-carboxylate (33) | H1N1 (A/Puerto Rico/8/34 H274Y) (IC50 = 9.40 ± 1.96 µM); H1N1 (A/FM-1/1/47) (IC50 = 4.80 ± 1.28 µM); H3N2 (A/Aichi/2/68) (IC50 = 5.12 ± 1.49 µM) | Diaporthe sp. (SCSIO 41011), isolated from Rhizophora stylosa | [88] |
| Methyl-(2-chloro-l,6-dihydroxy-3-methylxanthone)-8-carboxylate (34) | H1N1 (IC50 = 133.4 µM); HSV-1 (IC50 = 55.5 µM); HSV-2 (IC50 = 175.5 µM) | Aspergillus iizukae collected from coastal saline soil | [89] |
| Methyl-(4-chloro-l,6-dihydroxy-3-methylxanthone)-8-carboxylate (35) | H1N1 (IC50 = 44.6 µM); HSV-1 (IC50 = 21.4 µM); HSV-2 (IC50 = 76.7 µM) | ||
| Methyl-(4-chloro-6-hydroxy-1-methoxy-3-methylxanthone)-8-carboxylate (36) | H1N1 (IC50 ≥ 200 µM); HSV-1 (IC50 = 139.4 µM); HSV-2 (IC50 ≥ 200 µM) | ||
| Methyl-(6-hydroxy-1-methoxy-3-methylxanthone)-8-carboxylate (37) | H1N1 (IC50 ≥ 200 µM); HSV-1 (IC50 = 157.7 µM); HSV-2 (IC50 = 163.3 µM) | ||
| 4-Chloro-1,6-dihydroxy-3-methylxanthone-8-carboxylic acid (38) | H1N1 (IC50 ≥ 200 µM); HSV-1 (IC50 = 183.3 µM); HSV-2 (IC50 ≥ 200 µM) | ||
| 2,4-Dichloro-1,6-dihydroxy-3-methylxanthone-8-carboxylic acid (39) | H1N1 (IC50 ≥ 200 µM); HSV-1 (IC50 = 144.4 µM); HSV-2 (IC50 ≥ 200 µM) | ||
| Methyl-(l,6-dihydroxy-3-methylxanthone)-8-carboxylate (40) | H1N1 (IC50 = 140.4 µM); HSV-1 (IC50 = 75.7 µM); HSV-2 (IC50 = 95.4 µM) | ||
| 2-Hydroxy-1-(hydroxymethyl)-8-methoxy-3-methyl-9H-xanthen-9- one (41) | H1N1 (A/PuertoRico/8/34) (IC50 = 4.70 ± 1.11 µM); H1N1 (A/FM-1/1/47) (IC50 = 4.04 ± 0.58 µM) | Aspergillus sydowii (SCSIO 41.301) isolated from Phakellia fusca | [90] |
| 2-Hydroxy-1-(hydroxymethyl)-7,8-dimethoxy-3-methyl-9H- xanthen-9-one (42) | H1N1 (A/PuertoRico/8/34) (IC50 = 2.17 ± 1.39 µM) | ||
| Sterigmatocystin A (110) | HSV-2 (IC50 = 47.11 µM) | Aspergillus versicolor (15XS43ZD-1) strain was isolated from sponge collected from Xisha Islands, China | [91] |
| Sterigmatocystin B (111) | HSV-2 (IC50 = 39.45 µM) | ||
| Sterigmatocystin C (112) | HSV-2 (IC50 = 38.73 µM) | ||
| Asperxanthone (113) | Tobacco mosaic virus: inhibitory rate 62.9% | Aspergillus sp. collected in Quan-Zhou Gulf | [92] |
| Epiremisporine B (121) | EV71 (IC50 = 19.8 μM); H3N2 (IC50 = 24.1 μM) | Penicillium sp. (SCSIO Ind16F01) isolated from sediment collected in the Indian Ocean | [47] |
| Penicillixanthone A (154) | HIV-1 (SF162) (10 μM, 90.86 ± 0.82%); HIV-1 (CCR5-tropic) (IC50 = 0.36 µM); HIV-1 (CXCR4-tropic) (IC50 = 0.26 µM) | Aspergillus fumigates isolated from a jellyfish | [93] |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Chrysoxanthone (48) | α-Glucosidase inhibition (IC50 = 0.04 mM) | Penicillium chrysogenum (SCSIO 41001) isolated from sediment collected in the Indian Ocean | [94] |
| Staprexanthone A (69) | Pancreatic β-cell number (zebrafish model): ~40 at 10 µM | Stachybotrys chartarum (HDN16-358) isolated from mangrove collected in Fujian Province | [95] |
| Staprexanthone B (70) | Pancreatic β-cell number (zebrafish model): 40 at 10 µM | ||
| Staprexanthone C (71) | Pancreatic β-cell number (zebrafish model): ~35 at 10 µM | ||
| Staprexanthone D (72) | Pancreatic β-cell number (zebrafish model): ~35 at 10 µM | ||
| Staprexanthone E (73) | Pancreatic β-cell number (zebrafish model): ~40 at 10 µM | ||
| Austocystin J (96) | Inhibitory effect against phosphatases: SHP1 (IC50 = 15 μM); MEG2 (IC50 = 77 μM) | Aspergillus puniceus (SCSIO z021) | [96] |
| Austocystin K (97) | Inhibitory effect against phosphatases: TCPTP (IC50 = 16 μM); SHP1 (IC50 = 3.8 μM) | ||
| Austocystin L (98) | Inhibitory effect against phosphatases: TCPTP (IC50 = 12 μM); SHP1 (IC50 = 20 μM); CDC25B (IC50 = 24 μM) | ||
| Austocystin M (99) | Inhibitory effect against phosphatases: TCPTP (IC50 = 12 μM); SHP2 (IC50 = 9.5 μM); PTP1B (IC50 = 4.6 μM) | ||
| Austocystin N (100) | Inhibitory effect against phosphatases: SHP1 (IC50 = 17 μM) | ||
| Austocystin I (101) | Inhibitory effect against phosphatases: MEG2 (IC50 = 16 μM); CDC25B (IC50 = 19 μM) | ||
| Austocystin F (102) | Inhibitory effect against phosphatases: SHP1 (IC50 = 6.7 μM); MEG2 (IC50 = 2.1 μM); CDC25B (IC50 = 6.7 μM); CD45 (IC50 = 20 μM) | ||
| Austocystin A (103) | Inhibitory effect against phosphatases: TCPTP (IC50 = 19 μM); MEG2 (IC50 = 8.1 μM); CDC25B (IC50 = 16 μM) | ||
| Austocystin H (104) | Inhibitory effect against phosphatases: TCPTP (IC50 = 3.0 μM); SHP1 (IC50 = 1.3 μM); SHP2 (IC50 = 1.3 μM); MEG2 (IC50 = 0.60 μM); PTP1B (IC50 = 0.90 μM); CDC25B (IC50 = 1.3 μM); CD45 (IC50 = 14 μM) | ||
| Austocystin B (105) | Inhibitory effect against phosphatases: TCPTP (IC50 = 8.8 μM); SHP2 (IC50 = 2.0 μM); MEG2 (IC50 = 1.3 μM); PTP1B (IC50 = 1.8 μM); CDC25B (IC50 = 1.3 μM) | ||
| Austocystin D (106) | Inhibitory effect against phosphatases: PTP1B (IC50 = 1.7 μM) | ||
| 8-O-Methyldihydrodemethylsterigmatocystin (107) | Inhibitory effect against phosphatases: TCPTP (IC50 = 11 μM); SHP1 (IC50 = 5.5 μM); MEG2 (IC50 = 4.6 μM); CDC25B (IC50 = 4.9 μM); CD45 (IC50 = 6.1 μM) | ||
| (1′ R,2′ R)-compound V (108) | Inhibitory effect against phosphatases: TCPTP (IC50 = 19 μM); SHP1 (IC50 = 6.9 μM); MEG2 (IC50 = 4.2 μM) |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| 1,4,7-Trihydroxy-6-methylxanthone (15) | DPPH (IC50 = 6.92 µg mL−1); ABTS (IC50 = 2.35 µg mL−1) | Talaromyces islandicus (EN-501) isolated from Laurencia okamurai | [59] |
| 1,4,5-Trihydroxy-2-methylxanthone (16) | DPPH (IC50 = 1.23 µg mL−1); ABTS (IC50 = 1.27 µg mL−1) | ||
| Norlichexanthone (17) | DPPH (% Scavenging effect: 6.2% at 25.0 μg mL−1; 12.9% at 50 μg mL−1; 25.3% at 100 μg mL−1; 90.6% at 500 μg mL−1) | Enteromorpha sp. collected at Fehmarn Island | [81] |
| 2,3,6,8-Tetrahydroxy-1-methylxanthone (28) | DPPH (% Scavenging effect: 94.7% at 25.0 μg mL−1; 94.8% at 50 μg mL−1; 95.2% at 100 μg mL−1; 95.4% at 500 μg mL−1) Linolenic acid peroxidation (% Inhibition: 17.0% at 7.4 μg mL−1; 37.0% at 37 μg mL−1) | ||
| Arthone C (43) | DPPH (IC50 = 16.9 µM); ABTS (IC50 = 18.7 µM) | Arthrinium sp. (UJNMF0008) | [97] |
| 2,3,4,6,8-Pentahydroxy-1-methylxanthone (44) | DPPH (IC50 = 22.1 µM); ABTS (IC50 = 18.0 µM) | ||
| Sterigmatocystin (81) | ABTS (0.65 ± 0.13 TEAC values) | Aspergillus versicolor (A-21-2-7) isolated from sediment collected in the South China Sea | [98] |
| Oxisterigmatocystin C (93) | ABTS (1.16 ± 0.18 TEAC values) | ||
| Oxisterigmatocystin D (120) | ABTS (0.55 ± 0.13 TEAC values) |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Norlichexanthone (17) | COX-2 (IC50 = 34.3 µM) | Stachybotry sp. (ZSDS1F1-2) isolated from a sponge collected at Xisha Island | [34] |
| Yicathin C (18) | NO inhibition (27.0 ± 3.2%); NF-κB inhibition (56.8 ± 5.7%) | Aspergillus europaeus (WZXY-SX-4-1) isolated from Xestospongia testudinaria | [99] |
| IL-6 cytokine % at 1 µM: 78.37 ± 7.78% | Aspergillus wentii isolated from Gymnogongrus flabelliformis collected at Pingtan Island | [4] | |
| Yicathin B (19) | IL-6 cytokine % at 10 µM: 95.65 ± 17.21% | ||
| NO inhibition (35.3 ± 3.9%); NF-κB inhibition (81.2 ± 8.3%) | Aspergillus europaeus (WZXY-SX-4-1) isolated from Xestospongia testudinaria | [99] | |
| 1,3,6-trihydroxy-8-methylxanthone (45) | COX-2 (IC50 = 12.2 µM) | Arthrinium sp. (ZSDS1-F3) isolated from a sponge collected at Xisha Islands | [100] |
| Calyxanthone (46) | NO inhibition (17.6 ± 5.1) NF-κB: 63.7 ± 5.6 | Aspergillus europaeus (WZXY-SX-4-1) isolated from Xestospongia testudinaria | [99] |
| Yicathin A (47) | NO inhibition (23.7 ± 4.8%); NF-κB inhibition (13.0 ± 9.8%) | ||
| Euroanthone A (166) | NO inhibition (42.2 ± 2.3%); NF-κB inhibition (68.8 ± 7.0%) | ||
| Euroanthone B (167) | NO inhibition (23.4 ± 3.3%); NF-κB inhibition (52.3 ± 10.6%) |
| Name | Activity | Source | Ref. |
|---|---|---|---|
| Sydowinin A (2) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 6.5 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 7.1 μg mL−1) | Penicillium sp. (ZJ-SY2) isolated from Sonneratia apetala | [101] |
| Sydowinin B (3) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 19.2 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 20.8 μg mL−1) | ||
| Methyl 8-hydroxy-6-methyl-9-oxo-9H-xanthene-1- carboxylate (7) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 25.7 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 26.4 μg mL−1) | ||
| Conioxanthone A (12) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 8.2 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 7.5 μg mL−1) | ||
| Pinselin (49) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 5.9 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 7.5 μg mL−1) | ||
| Epiremisporine B (121) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 30.8 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 31.2 μg mL−1) | ||
| Remisporine B (169) | Immunosuppressive: Inhibition of Con A-Induced proliferation (IC50 = 30.1 μg mL−1); Inhibition of LPS-Induced proliferation (IC50 = 32.4 μg mL−1) | ||
| Paeciloxanthone (68) | Anti-Alzheimer: acetylcholinesterase inhibition (IC50 = 2.25 µg mL−1) | Paecilomyces sp. isolated from a mangrove collected in the Taiwan Strait | [40] |
| Chaetoxanthone A (78) | Antiprotozoal: T. brucei rhodesiense (strain STIB 900) (IC50 = 4.7 μg mL−1); T. cruzi (strain Tulahuen C4) (IC50 ≥ 10 μg mL−1); L. donovani (strain MHOM-ET-67/L82) (IC50 = 5.3 μg mL−1); P. falciparum (IC50 3.5 μg mL−1) | Chaetomium sp. isolated from the Greek alga collected at Santorini Island | [41,102,103] |
| Chaetoxanthone B (79) | Antiprotozoal: T. brucei rhodesiense (strain STIB 900) (IC50 = 9.3 μg mL−1); T. cruzi (strain Tulahuen C4) (IC50 = 7.1 μg mL−1); L. donovani (strain MHOM-ET-67/L82) (IC50 = 3.4 μg mL−1); P. falciparum (IC50 = 0.5 μg mL−1) | ||
| Chaetoxanthone C (80) | Antiprotozoal: T. brucei rhodesiense (strain STIB 900) (IC50 = 42.6 μg mL−1); T. cruzi (strain Tulahuen C4) (IC50 = 1.5 μg mL−1); L. donovani (strain MHOM-ET-67/L82) (IC50 = 3.1 μg mL−1); P. falciparum (IC50 = 4.0 μg mL−1) | ||
| Aspergixanthone A (82) | Against aquatic pathogens: V. parahemolyticus (MIC = 25.0 µM); V. anguillarum (MIC = 25.0 µM); V. alginolyticus (MIC = 25.0 µM) | Aspergillus sp. (ZA-01) | [104] |
| Sterigmatocystin A (110) | Angiogenesis: Increase length of intersomitic vessels of transgenic zebrafish at 1.25 μM | Aspergillus versicolor (15XS43ZD-1) isolated from a sponge collected at Xisha Island | [91] |
| Aspergixanthone I (114) | Against aquatic pathogens: V. parahemolyticus (MIC = 1.56 µM); V. anguillarum (MIC = 1.56 µM); V. alginolyticus (MIC = 3.12 µM) | Aspergillus sp. (ZA-01) | [104] |
| Aspergixanthone J (115) | Against aquatic pathogens: V. parahemolyticus (MIC = 6.25µM); V. anguillarum (MIC = 25.0 µM); V. alginolyticus (MIC = 25.0 µM) | ||
| Aspergixanthone K (116) | Against aquatic pathogens: V. parahemolyticus (MIC = 3.12 µM); V. anguillarum (MIC = 25.0 µM); V. alginolyticus (MIC = 12.5 µM) | ||
| 15-Acetyl tajixanthone hydrate (117) | Against aquatic pathogens: V. parahemolyticus (MIC = 12.5 µM); V. anguillarum (MIC = 25.0 µM); V. alginolyticus (MIC = 12.5 µM) | ||
| Tajixanthone hydrate (118) | Against aquatic pathogens: V. parahemolyticus (MIC = 6.25 µM); V. anguillarum (MIC = 6.25 µM); V. alginolyticus (MIC = 12.5 µM) | ||
| 16-Chlorotajixanthone (119) | Against aquatic pathogens: V. parahemolyticus (MIC = 25.0 µM); V. anguillarum (MIC = 6.25 µM); V. alginolyticus (MIC = 25.0 µM) | ||
| Isosecosterigmatocystin (169) | Against aquatic pathogens: Ed. ictaluri (IC50 = 16 μg mL−1) | Aspergillus nidulans (MA-143) isolated from Rhizophora stylosa | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, J.X.; Loureiro, D.R.P.; Dias, A.L.; Reis, S.; Pinto, M.M.M.; Afonso, C.M.M. Bioactive Marine Xanthones: A Review. Mar. Drugs 2022, 20, 58. https://doi.org/10.3390/md20010058
Soares JX, Loureiro DRP, Dias AL, Reis S, Pinto MMM, Afonso CMM. Bioactive Marine Xanthones: A Review. Marine Drugs. 2022; 20(1):58. https://doi.org/10.3390/md20010058
Chicago/Turabian StyleSoares, José X., Daniela R. P. Loureiro, Ana Laura Dias, Salete Reis, Madalena M. M. Pinto, and Carlos M. M. Afonso. 2022. "Bioactive Marine Xanthones: A Review" Marine Drugs 20, no. 1: 58. https://doi.org/10.3390/md20010058
APA StyleSoares, J. X., Loureiro, D. R. P., Dias, A. L., Reis, S., Pinto, M. M. M., & Afonso, C. M. M. (2022). Bioactive Marine Xanthones: A Review. Marine Drugs, 20(1), 58. https://doi.org/10.3390/md20010058