
Marine-Derived Xanthone from 2010 to 2021: Isolation, Bioactivities and Total Synthesis




Abstract
:
1. Introduction
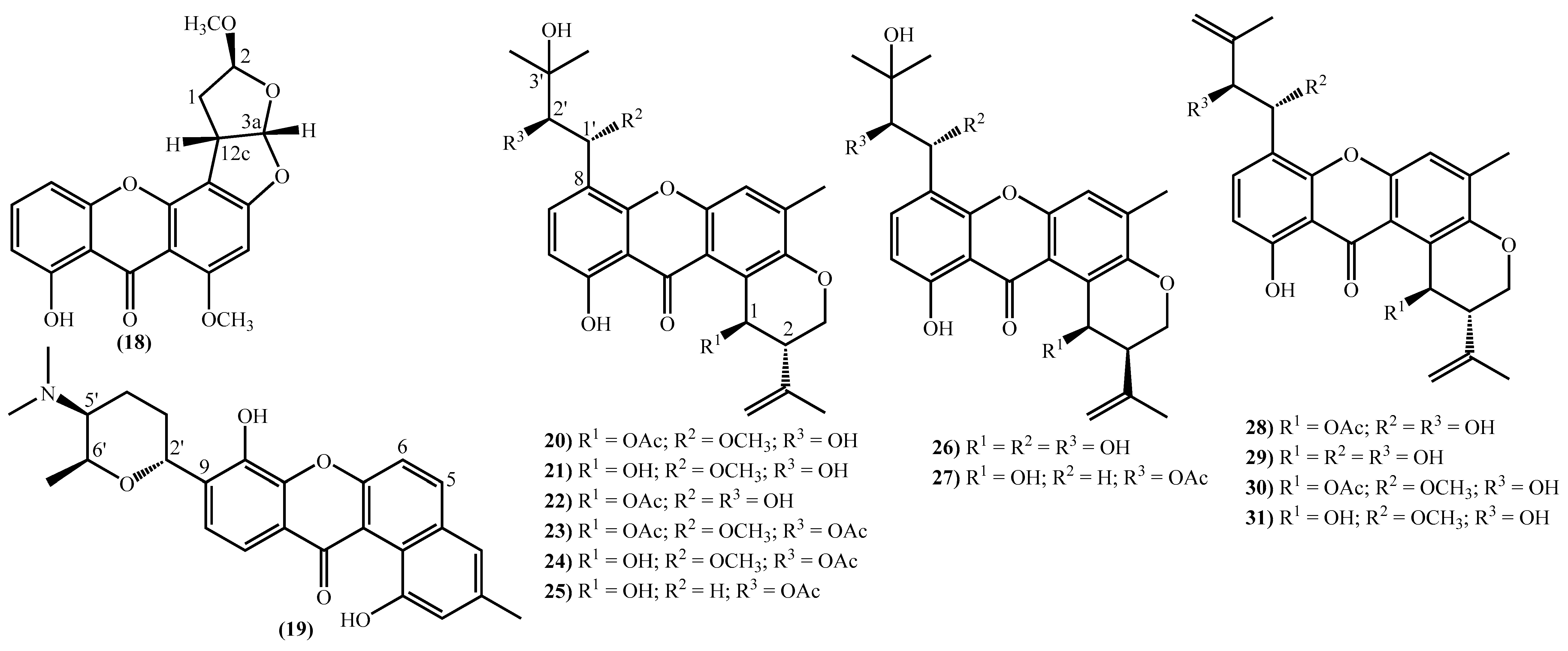
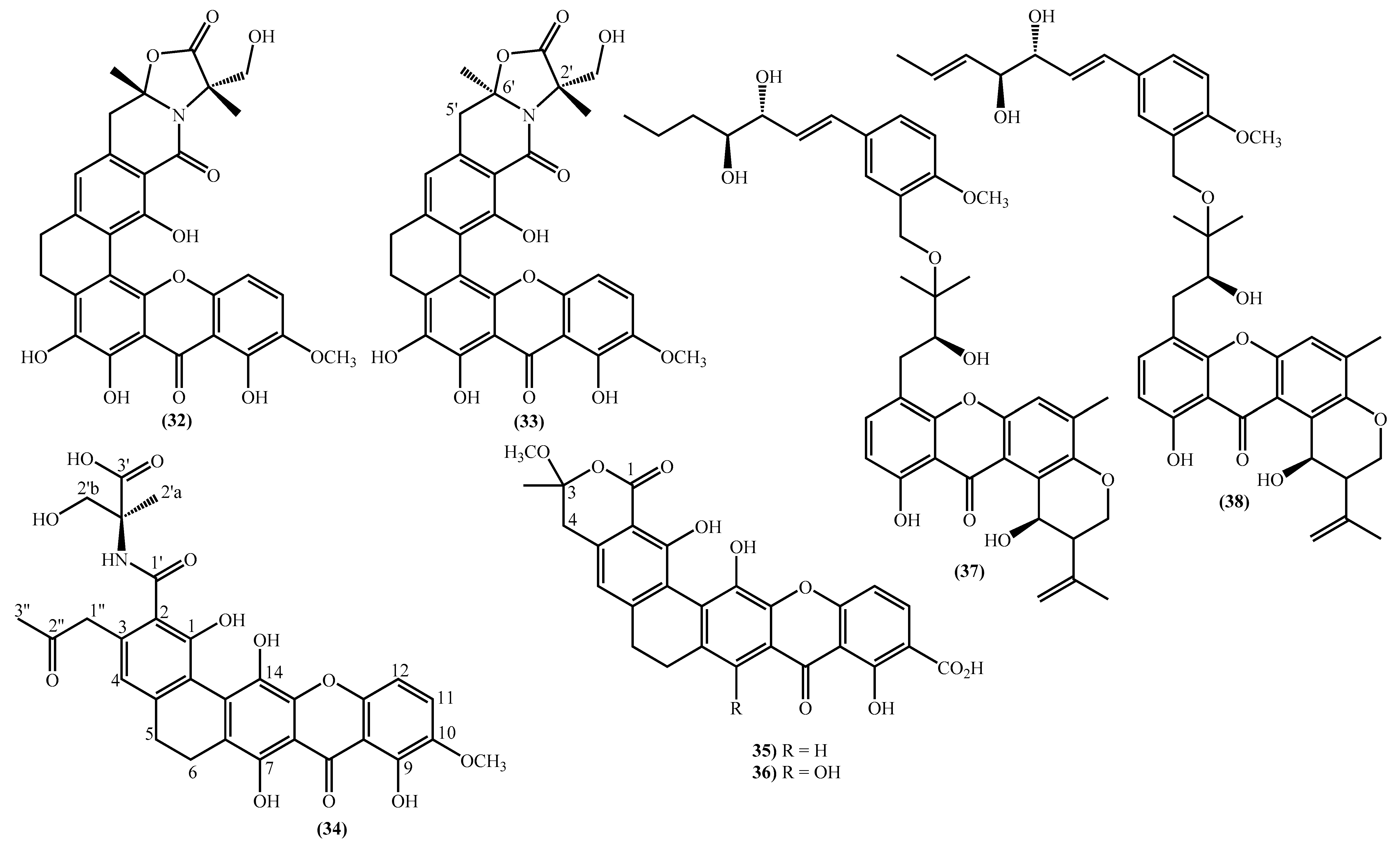
2. New Marine Xanthones Isolated since 2010
3. Bioactivities Presented by Marine Xanthones
4. Xanthones Obtained by Total Synthesis
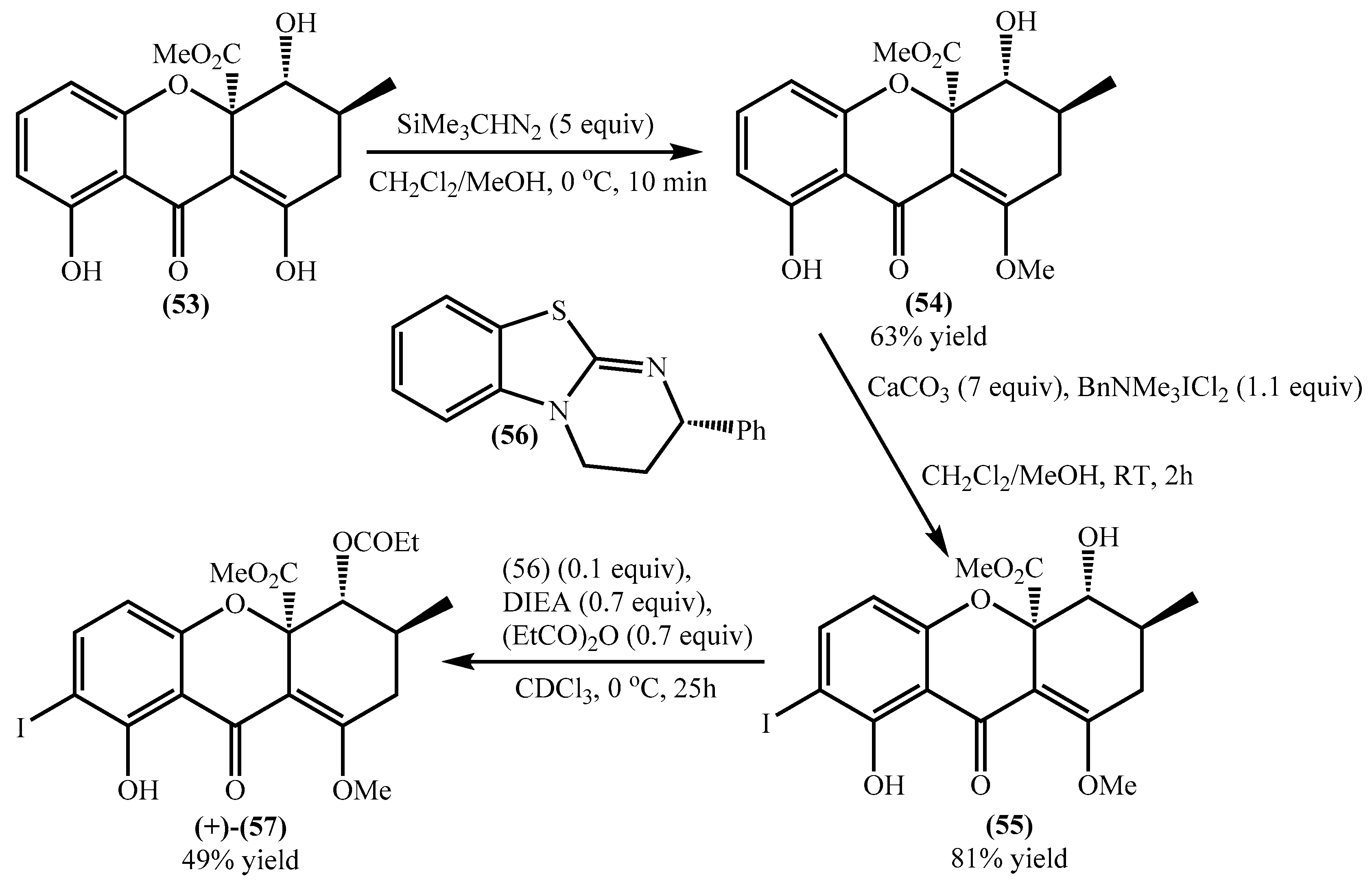
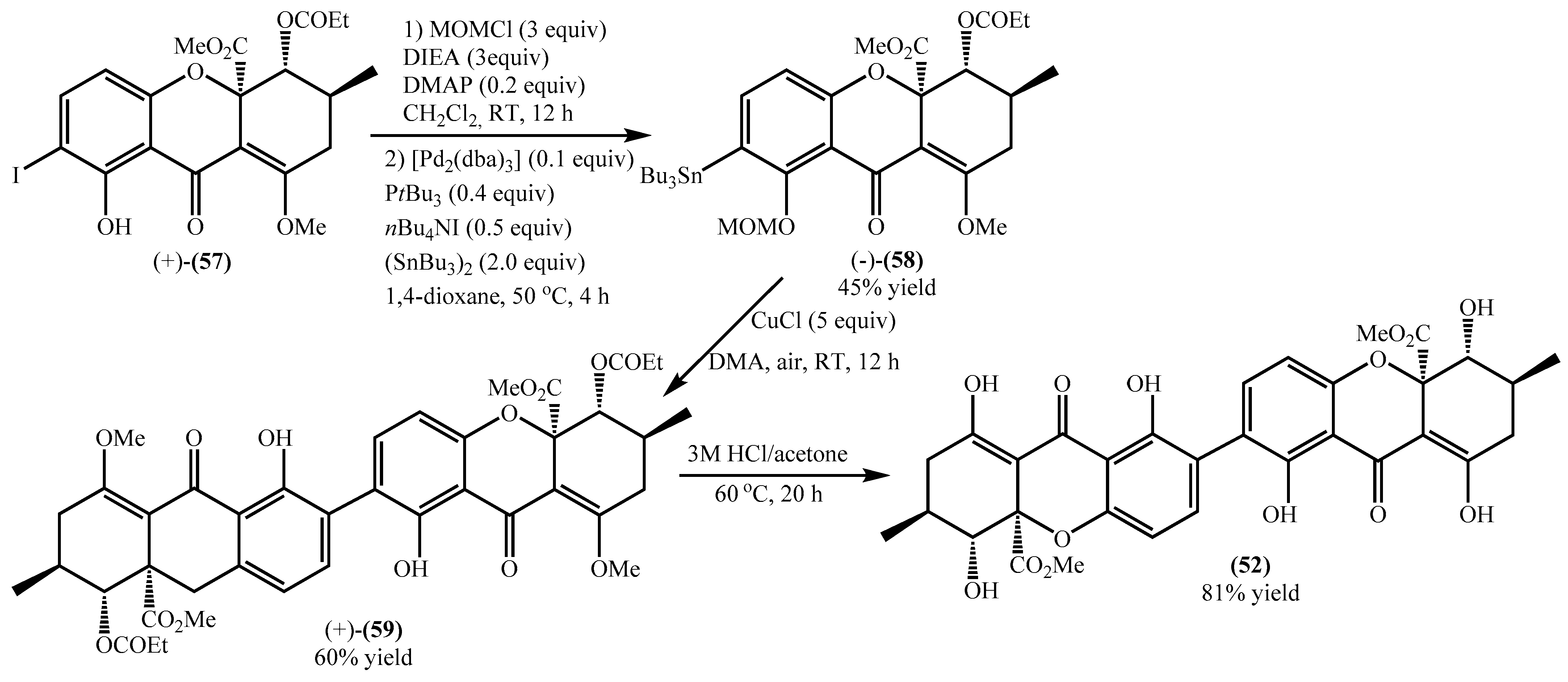
4.1. Total Synthesis of Secalonic Acid D
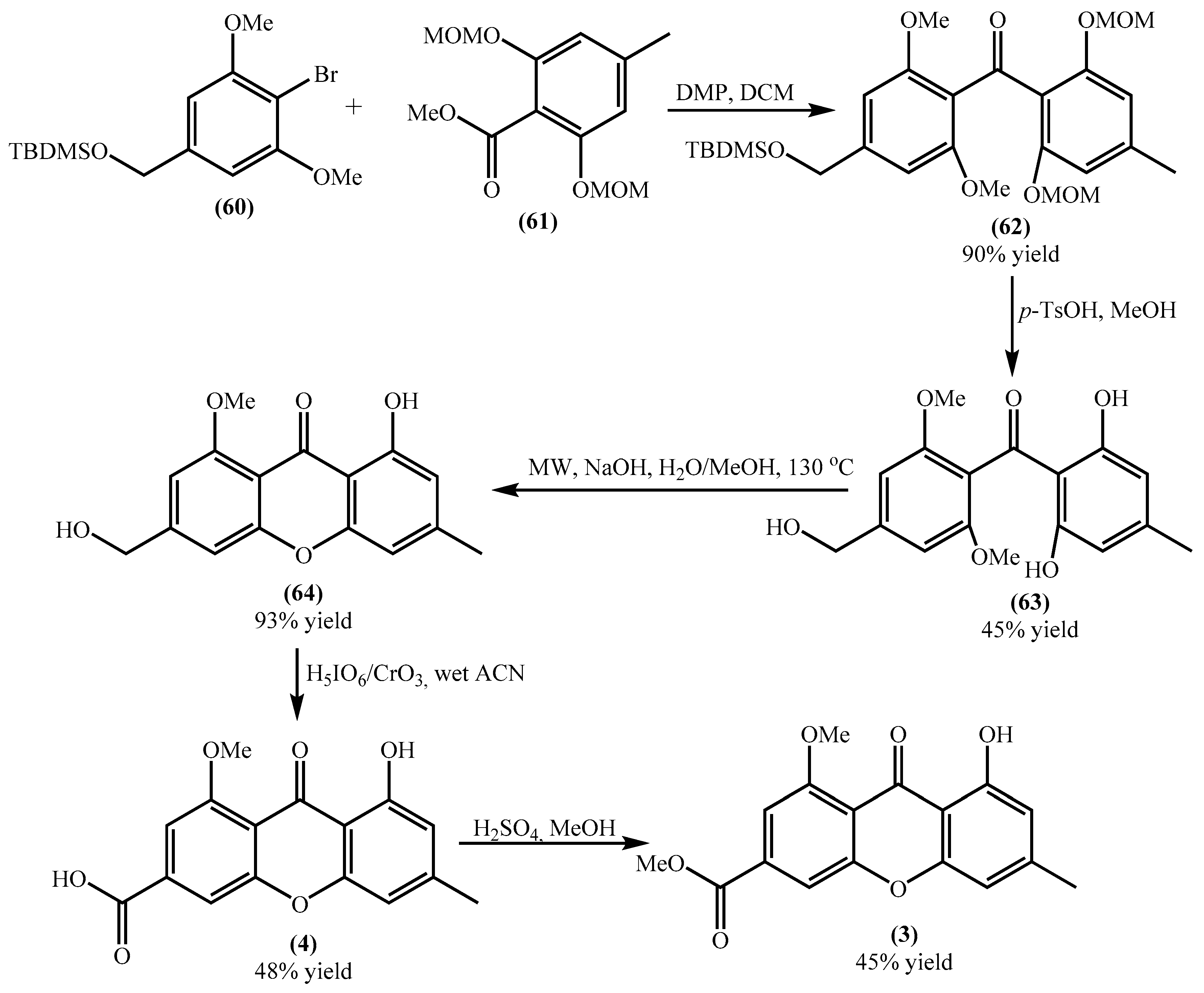
4.2. Total Synthesis of Yicathins B and C and Analogues
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khattab, A.R.; Farag, M.A. Marine and terrestrial endophytic fungi: A mine of bioactive xanthone compounds, recent progress, limitations, and novel applications. Crit. Rev. Biotechnol. 2022, 42, 403–430. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.M.; Palmeira, A.; Fernandes, C.; Resende, D.I.S.P.; Sousa, E.; Cidade, H.; Tiritan, M.E.; Correia-da-Silva, M.; Cravo, S. From natural products to new synthetic small molecules: A journey through the world of xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.S.; Chand, M.; Sakhuja, R.; Jain, S.C. Xanthones as potential antioxidants. Curr. Med. Chem. 2013, 20, 4481–4507. [Google Scholar] [CrossRef] [PubMed]
- Negi, J.S.; Bisht, V.K.; Singh, P.; Rawat, M.S.M.; Joshi, G.P. Naturally occurring xanthones: Chemistry and biology. J. Appl. Chem. 2013, 2013, 621459. [Google Scholar] [CrossRef] [Green Version]
- Masters, K.-S.; Bräse, S. Xanthones from fungi, lichens, and bacteria: The natural products and their synthesis. Chem. Rev. 2012, 112, 3717–3776. [Google Scholar] [CrossRef]
- Resende, D.; Pereira-Terra, P.; Inácio, Â.; Costa, P.; Pinto, E.; Sousa, E.; Pinto, M. Lichen xanthones as models for new antifungal agents. Molecules 2018, 23, 2617. [Google Scholar] [CrossRef] [Green Version]
- Peres, V.; Nagem, T.J.; de Oliveira, F.F. Tetraoxygenated naturally occurring xanthones. Phytochemistry 2000, 55, 683–710. [Google Scholar] [CrossRef] [Green Version]
- Khattab, A.R.; Farag, M.A. Current status and perspectives of xanthones production using cultured plant biocatalyst models aided by in-silico tools for its optimization. Crit. Rev. Biotechnol. 2020, 40, 415–431. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Ghorab, D.M.H.; El-Barbary, M.A.; Zayed, M.F.; Göransson, U.; Larsson, S.; Verpoorte, R. Naturally occurring xanthones; latest investigations: Isolation, structure elucidation and chemosystematic significance. Curr. Med. Chem. 2009, 16, 2581–2626. [Google Scholar] [CrossRef]
- Loureiro, D.R.P.; Soares, J.X.; Costa, J.C.; Magalhães, Á.F.; Azevedo, C.M.G.; Pinto, M.M.M.; Afonso, C.M.M. Structures, activities and drug-likeness of anti-Infective xanthone derivatives isolated from the marine environment: A review. Molecules 2019, 24, 243. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.M.M.; Castanheiro, R.A.P.; Kijjoa, A. Xanthones from marine-derived microorganisms: Isolation, structure elucidation and biological activities. In Encyclopedia of Analytical Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 1–21. [Google Scholar]
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.A.; Kim, D.-K.; Jung, J.H. Bioactive metabolites from the sponge-derived fungus Aspergillus versicolor. Arch. Pharm. Res. 2010, 33, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, B.; Liu, J.; Xun, T.; Liu, Y.; Zhou, X. Penicillixanthone A, a marine-derived dual-coreceptor antagonist as anti-HIV-1 agent. Nat. Prod. Res. 2019, 33, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Zhang, X.-W.; Zhang, M.; Li, W.; Ma, Z.-Y.; Zhu, H.-J.; Cao, F. Aspergixanthones I–K, new anti-vibrio prenylxanthones from the marine-derived fungus Aspergillus sp. ZA-01. Mar. Drugs 2018, 16, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elnaggar, M.S.; Ebada, S.S.; Ashour, M.L.; Ebrahim, W.; Müller, W.E.G.; Mándi, A.; Kurtán, T.; Singab, A.; Lin, W.; Liu, Z.; et al. Xanthones and sesquiterpene derivatives from a marine-derived fungus Scopulariopsis sp. Tetrahedron 2016, 72, 2411–2419. [Google Scholar] [CrossRef] [Green Version]
- Li, J.L.; Jiang, X.; Liu, X.; He, C.; Di, Y.; Lu, S.; Huang, H.; Lin, B.; Wang, D.; Fan, B. Antibacterial anthraquinone dimers from marine derived fungus Aspergillus sp. Fitoterapia 2019, 133, 1–4. [Google Scholar] [CrossRef]
- Zhu, A.; Yang, M.-Y.; Zhang, Y.-H.; Shao, C.-L.; Wang, C.-Y.; Hu, L.-D.; Cao, F.; Zhu, H.-J. Absolute configurations of 14,15-hydroxylated prenylxanthones from a marine-derived Aspergillus sp. fungus by chiroptical methods. Sci. Rep. 2018, 8, 10621. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Wang, K.; Xue, Y.; Chen, J.; Su, H.; Zhang, J.; Wu, Y.; Jia, J.; Bi, H.; Wang, H.; et al. Three new metabolites from the marine-derived fungus Aspergillus sp. WHUF03110. Nat. Prod. Commun. 2021, 16, 1934578X2110550. [Google Scholar] [CrossRef]
- Liu, L.-L.; Xu, Y.; Han, Z.; Li, Y.-X.; Lu, L.; Lai, P.-Y.; Zhong, J.-L.; Guo, X.-R.; Zhang, X.-X.; Qian, P.-Y. Four new antibacterial xanthones from the marine-derived Actinomycetes Streptomyces caelestis. Mar. Drugs 2012, 10, 2571–2583. [Google Scholar] [CrossRef] [Green Version]
- Ningsih, B.N.S.; Rukachaisirikul, V.; Pansrinun, S.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. New aromatic polyketides from the marine-derived fungus Pseudopithomyces maydicus PSU-AMF350 and their antimicrobial activity. Nat. Prod. Res. 2022, 36, 1–8. [Google Scholar] [CrossRef]
- Chang, Y.; Xing, L.; Sun, C.; Liang, S.; Liu, T.; Zhang, X.; Zhu, T.; Pfeifer, B.A.; Che, Q.; Zhang, G.; et al. Monacycliones G–K and ent-gephyromycin A, angucycline derivatives from the marine-derived Streptomyces sp. HDN15129. J. Nat. Prod. 2020, 83, 2749–2755. [Google Scholar] [CrossRef]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Liu, J.; Liu, Y. Emerixanthone E, a new xanthone derivative from deep sea fungus Emericella sp SCSIO 05240. Nat. Prod. Res. 2019, 33, 2088–2094. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Qi, X.; Mo, X.; Yu, G.; Wang, Q.; Zhu, T.; Gu, Q.; Liu, M.; Li, J.; Li, D. Structure-based discovery of cytotoxic dimeric tetrahydroxanthones as potential topoisomerase I inhibitors from a marine-derived fungus. Eur. J. Med. Chem. 2018, 148, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yang, J.; Chen, F.; Lin, X.; Chen, C.; Zhou, X.; Liu, S.; Liu, Y. Structurally diverse polyketides from the mangrove-derived fungus Diaporthe sp. SCSIO 41011 with their anti-influenza A virus activities. Front. Chem. 2018, 6, 282. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, S.; Liu, W.; Liu, Y.; Huang, X.; She, Z. Polyketides with immunosuppressive activities from mangrove endophytic fungus Penicillium sp. ZJ-SY2 Hongju. Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, antibiotic polyketides from the marine fungus Engyodontium album strain LF069. Chem.-Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Yu, G.; Kurtán, T.; Mándi, A.; Peng, J.; Mo, X.; Liu, M.; Li, H.; Sun, X.; Li, J.; et al. Versixanthones A–F, cytotoxic xanthone–chromanone dimers from the marine-derived fungus Aspergillus versicolor HDN1009. J. Nat. Prod. 2015, 78, 2691–2698. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, C.; Long, H.; Chen, R.; Liu, D.; Proksch, P.; Guo, P.; Lin, W. Varioxiranols A–G and 19-O-methyl-22-methoxypre-shamixanthone, PKS and hybrid PKS-derived metabolites from a sponge-associated Emericella variecolor fungus. J. Nat. Prod. 2015, 78, 2461–2470. [Google Scholar] [CrossRef]
- Jang, N.Y.; Peng, F.; Fang, Z.K.; Wu, Y.D.; Chen, M.H.; Xie, Y.; Wang, H.R.; Jiang, H.; Lian, Y.Y. Study on aromatic polyketide metabolite with antibacterial activity from the marine-derived Actinomadura sp. FIM95-F26. Chin. J. Antibiot. 2015, 40, 161–165. [Google Scholar]
- Eltamany, E.E.; Abdelmohsen, U.R.; Ibrahim, A.K.; Hassanean, H.A.; Hentschel, U.; Ahmed, S.A. New antibacterial xanthone from the marine sponge-derived Micrococcus sp. EG45. Bioorg. Med. Chem. Lett. 2014, 24, 4939–4942. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Zhang, X.-Y.; Zheng, Z.-H.; Xu, X.-Y.; Qi, S.-H. Three new polyketides from marine-derived fungus Penicillium citrinum SCSGAF 0167. Nat. Prod. Res. 2014, 28, 239–244. [Google Scholar] [CrossRef]
- Yang, J.X.; Qiu, S.; She, Z.; Lin, Y. A new xanthone derivative from the marine fungus Phomopsis sp. (No. SK7RN3G1). Chem. Nat. Compd. 2013, 49, 246–248. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Shao, C.; Ding, W.; She, Z.; Lin, Y. A new xanthone derivative from the co-culture broth of two marine fungi (strain No. E33 and K38). Chem. Nat. Compd. 2011, 47, 382–384. [Google Scholar] [CrossRef]
- Gao, H.; Zhou, L.; Li, D.; Gu, Q.; Zhu, T.-J. New cytotoxic metabolites from the marine-derived fungus Penicillium sp. ZLN29. Helv. Chim. Acta 2013, 96, 514–519. [Google Scholar] [CrossRef]
- Li, H.-L.; Li, X.-M.; Liu, H.; Meng, L.-H.; Wang, B.-G. Two New diphenylketones and a new xanthone from Talaromyces islandicus EN-501, an endophytic fungus derived from the marine red alga Laurencia okamurai. Mar. Drugs 2016, 14, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Gao, J.; Hu, J.; He, H.; Huang, P.; Zhang, L.; Song, F. One new xanthenone from the marine-derived fungus Aspergillus versicolor MF160003. Nat. Prod. Res. 2020, 34, 2907–2912. [Google Scholar] [CrossRef]
- Wu, Z.-H.; Liu, D.; Xu, Y.; Chen, J.-L.; Lin, W.-H. Antioxidant xanthones and anthraquinones isolated from a marine-derived fungus Aspergillus versicolor. Chin. J. Nat. Med. 2018, 16, 219–224. [Google Scholar] [CrossRef]
- Liu, S.; Wang, H.; Su, M.; Hwang, G.J.; Hong, J.; Jung, J.H. New metabolites from the sponge-derived fungus Aspergillus sydowii J05B-7F-4. Nat. Prod. Res. 2017, 31, 1682–1686. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, X.-P.; Ju, Z.-R.; Liao, X.-J.; Huang, X.-J.; Zhang, C.; Zhao, B.-X.; Xu, S.-H. Aspergchromones A and B, two new polyketides from the marine sponge-associated fungus Aspergillus sp. SCSIO XWS03F03. J. Asian Nat. Prod. Res. 2017, 19, 684–690. [Google Scholar] [CrossRef]
- Ueda, J.; Takagi, M.; Shin-ya, K. New xanthoquinodin-like compounds, JBIR-97, -98 and -99, obtained from marine sponge-derived fungus Tritirachium sp. SpB081112MEf2. J. Antibiot. 2010, 63, 615–618. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, C. Marine natural products in medicinal chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [Green Version]
- Wilson, Z.E.; Brimble, M.A. Molecules derived from the extremes of life. Nat. Prod. Rep. 2009, 26, 44–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, H.-F.; Zhang, L.-H.; Liu, F.; He, F.-J.; Bai, J.; Hua, H.-M.; Chen, G.; Pei, Y.-H. New compound with DNA Topo I inhibitory activity purified from Penicillium oxalicum HSY05. Nat. Prod. Res. 2015, 29, 2197–2202. [Google Scholar] [CrossRef] [PubMed]
- Shinde, P.; Banerjee, P.; Mandhare, A. Marine natural products as source of new drugs: A patent review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Khamthong, N.; Rukachaisirikul, V.; Tadpetch, K.; Kaewpet, M.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Tetrahydroanthraquinone and xanthone derivatives from the marine-derived fungus Trichoderma aureoviride PSU-F95. Arch. Pharm. Res. 2012, 35, 461–468. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Li, Z.; Wang, Y.; Tian, L.; Pei, Y.; Hua, H. A new sulfo-xanthone from the marine-derived fungus Penicillium sacculum. Chem. Nat. Compd. 2012, 48, 771–773. [Google Scholar] [CrossRef]
- Sun, R.-R.; Miao, F.-P.; Zhang, J.; Wang, G.; Yin, X.-L.; Ji, N.-Y. Three new xanthone derivatives from an algicolous isolate of Aspergillus wentii. Magn. Reson. Chem. 2013, 51, 65–68. [Google Scholar] [CrossRef]
- Liu, F.; Lin, X.; Zhou, X.; Chen, M.; Huang, X.; Yang, B.; Tao, H. Xanthones and quinolones derivatives produced by the deep-sea-derived fungus Penicillium sp. SCSIO Ind16F01. Molecules 2017, 22, 1999. [Google Scholar] [CrossRef] [Green Version]
- Soares, J.X.; Loureiro, D.R.P.; Dias, A.L.; Reis, S.; Pinto, M.M.M.; Afonso, C.M.M. Bioactive marine xanthones: A review. Mar. Drugs 2022, 20, 58. [Google Scholar] [CrossRef]
- Loureiro, D.R.P.; Magalhães, Á.F.; Soares, J.X.; Pinto, J.; Azevedo, C.M.G.; Vieira, S.; Henriques, A.; Ferreira, H.; Neves, N.; Bousbaa, H.; et al. Yicathins B and C and analogues: Total synthesis, lipophilicity and biological activities. ChemMedChem 2020, 15, 749–755. [Google Scholar] [CrossRef]
- Moon, K.; Chung, B.; Shin, Y.; Rheingold, A.L.; Moore, C.E.; Park, S.J.; Park, S.; Lee, S.K.; Oh, K.-B.; Shin, J.; et al. Pentacyclic antibiotics from a tidal mud flat-derived actinomycete. J. Nat. Prod. 2015, 78, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Ngan, N.T.T.; Quang, T.H.; Kim, K.-W.; Kim, H.J.; Sohn, J.H.; Kang, D.G.; Lee, H.S.; Kim, Y.-C.; Oh, H. Anti-inflammatory effects of secondary metabolites isolated from the marine-derived fungal strain Penicillium sp. SF-5629. Arch. Pharm. Res. 2017, 40, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Y.-P.; Li, X.-X.; Lu, Z.-H.; Zheng, Q.-H.; Liu, Q.-Y. Isolation of 4,4′-bond secalonic acid D from the marine-derived fungus Penicillium oxalicum with inhibitory property against hepatocellular carcinoma. J. Antibiot. 2019, 72, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Quang, T.H.; Huong, P.T.M.; Ngan, N.T.T.; Hanh, T.T.H.; Cuong, N.X.; Nam, N.H.; Van Minh, C. Secondary metabolites from a marine sponge-associated fungus Xenomyrothecium sp. IMBC-FP2.11. Vietnam J. Chem. 2020, 58, 752–758. [Google Scholar] [CrossRef]
- Hong, R. Secalonic acid D as a novel DNA topoisomerase I inhibitor from marine lichen-derived fungus Gliocladium sp. T31. Pharm. Biol. 2011, 49, 796–799. [Google Scholar] [CrossRef]
- Hu, Y.; Tao, L.; Wang, F.; Zhang, J.; Liang, Y.; Fu, L. Secalonic acid D reduced the percentage of side populations by down-regulating the expression of ABCG2. Biochem. Pharmacol. 2013, 85, 1619–1625. [Google Scholar] [CrossRef]
- Chen, L.; Cheng, M.; Li, Y.; Lin, S.; Zheng, Q.; Liu, Q. 4,4’-Bond secalonic acid D targets SP cells and inhibits metastasis in hepatocellular carcinoma. Mol. Med. Rep. 2020, 21, 2624–2632. [Google Scholar] [CrossRef]
- Steyn, P.S. The separation and detection of several mycotoxins by thin-layer chromatography. J. Chromatogr. A 1969, 45, 473–475. [Google Scholar] [CrossRef]
- Ren, H.; Tian, L.; Gu, Q.; Zhu, W. Secalonic acid D; A cytotoxic constituent from marine lichen-derived fungusGliocladium sp. T31. Arch. Pharm. Res. 2006, 29, 59–63. [Google Scholar] [CrossRef]
- Iwaguchi, T.; Kitagawa, H.; Hirose, K.; Ishida, T.; Yamamoto, T. 5-Di-(2’-tetrahydropyranyl)secalonic acid D as a new antibiotic derivative with anticancer activity. Gan 1980, 71, 900–906. [Google Scholar] [CrossRef]
- Tang, R.; Kimishima, A.; Setiawan, A.; Arai, M. Secalonic acid D as a selective cytotoxic substance on the cancer cells adapted to nutrient starvation. J. Nat. Med. 2020, 74, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhou, J.; Wang, H.; Mao, Z.; Xiao, W.; Wang, H.; She, Z.; Zhu, Y. The cell toxicity effect of secalonic acid D on GH3 cells and the related mechanisms. Oncol. Rep. 2010, 23, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Porco, J.A. Total syntheses of secalonic acids A and D. Angew. Chemie Int. Ed. 2014, 53, 3107–3110, including the supporting information. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, T.; Johnson, R.P.; Porco, J.A. Vinylogous addition of siloxyfurans to benzopyryliums: A concise approach to the tetrahydroxanthone natural products. J. Am. Chem. Soc. 2011, 133, 1714–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Source | Antibacterial (Positive Control Value) | Cytotoxic (Positive Control Value) |
|---|---|---|---|
| JBIR-99 (45a) | marine sponge-derived fungus Tritirachium sp. Sp081112MEf2 and marine fungus Engyodontium album Strain LF069 | Staphylococcus epidermidis IC50 0.21 ± 0.09 μM (chloramphenicol IC50 1.8 ± 0.04 μM) Staphylococcus aureus IC50 0.25 ± 0.07 μM (chloramphenicol IC50 2.9 ± 0.5 μM) | NIH 3T3 IC50 13.2 ± 2.7 μM (amoxifen citrate IC50 16.5 ± 0.2 μM) |
| Engyodontochone B (45b) | marine fungus Engyodontium album Strain LF069 | Staphylococcus epidermidis IC50 0.22 ± 0.03 μM (chloramphenicol IC50 1.8 ± 0.04 μM) Staphylococcus aureus IC50 0.24 ± 0.04 μM (chloramphenicol IC50 2.9 ± 0.5 μM) | NIH 3T3 IC50 14.4 ± 2.6 μM (amoxifen citrate IC50 16.5 ± 0.2 μM) |
| JBIR-97/98 (46a) | marine sponge-derived fungus Tritirachium sp. Sp081112MEf2 and marine fungus Engyodontium album Strain LF069 | Staphylococcus epidermidis IC50 0.20 ± 0.04 μM (chloramphenicol IC50 1.8 ± 0.04 μM) Staphylococcus aureus IC50 0.19 ± 0.02 μM (chloramphenicol IC50 2.9 ± 0.5 μM) | NIH 3T3 IC50 14.0 ± 1.1 μM (amoxifen citrate IC50 16.5 ± 0.2 μM) |
| Engyodontochone A (46b) | marine fungus Engyodontium album Strain LF069 | Staphylococcus epidermidis IC50 0.19 ± 0.04 μM (chloramphenicol IC50 1.8 ± 0.04 μM) Staphylococcus aureus IC50 0.17 ± 0.02 μM (chloramphenicol IC50 2.9 ± 0.5 μM) | NIH 3T3 IC50 11.0 ± 1.6 μM (amoxifen citrate IC50 16.5 ± 0.2 μM) |
| Compound (8) | marine fungus Talaromyces islandicus EN-501 found in red alga Laurencia okamurai | Escherichia coli MIC 32 μg/mL (chloramphenicol MIC 1 μg/mL) Pseudomonas aeruginosa MIC 32 μg/mL (chloramphenicol MIC 4 μg/mL) Staphylococcus aureus MIC > 64 μg/mL (chloramphenicol MIC 2 μg/mL) Vibrio alginolyticus MIC 32 μg/mL (chloramphenicol MIC 0.5 μg/mL) Vibrio harveyi MIC 32 μg/mL (chloramphenicol MIC 2 μg/mL) Vibrio parahaemolyticus MIC 32 μg/mL (chloramphenicol MIC 2 μg/mL) | |
| Aspergixanthone A (20) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus Bacillus anthraci Salmonella typhi Enterobacter aerogenes | MDA-MB-231 IC50 > 10.0 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 > 10.0 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 > 10.0 μM (cisplatin IC50 1.1 μM) HeLa IC50 > 10.0 μM (cisplatin IC50 0.82 μM) A-549 IC50 1.80 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone B (21) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus Bacillus anthraci Salmonella typhi Enterobacter aerogenes | MDA-MB-231 IC50 > 10.0 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 > 10.0 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 > 10.0 μM (cisplatin IC50 1.1 μM) HeLa IC50 > 10.0 μM (cisplatin IC50 0.82 μM) A-549 IC50 >10.0 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone C (22) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus Bacillus anthraci Salmonella typhi Enterobacter aerogenes | MDA-MB-231 IC50 3.3 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 2.8 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 3.6 μM (cisplatin IC50 1.1 μM) HeLa IC50 2.9 μM (cisplatin IC50 0.82 μM) A-549 IC50 3.2 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone E (23) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus Bacillus anthraci Salmonella typhi Enterobacter aerogenes | MDA-MB-231 IC50 > 10.0 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 > 10.0 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 > 10.0 μM (cisplatin IC50 1.1 μM) HeLa IC50 > 10.0 μM (cisplatin IC50 0.82 μM) A-549 IC50 > 10.0 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone F (24) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus Bacillus anthraci Salmonella typhi Enterobacter aerogenes | MDA-MB-231 IC50 9.8 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 2.7 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 3.6 μM (cisplatin IC50 1.1 μM) HeLa IC50 1.7 μM (cisplatin IC50 0.82 μM) A-549 IC50 1.1 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone D (26) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus Bacillus anthraci Salmonella typhi Enterobacter aerogenes | MDA-MB-231 IC50 > 10.0 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 > 10.0 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 > 10.0 μM (cisplatin IC50 1.1 μM) HeLa IC50 > 10.0 μM (cisplatin IC50 0.82 μM) A-549 IC50 > 10.0 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone G (28) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus MIC 0.78 μg/mL Bacillus anthraci MIC 12.5 μg/mL Salmonella typhi MIC 6.13 μg/mL Enterobacter aerogenes MIC 6.13 μg/mL | MDA-MB-231 IC50 > 10.0 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 > 10.0 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 > 10.0 μM (cisplatin IC50 1.1 μM) HeLa IC50 > 10.0 μM (cisplatin IC50 0.82 μM) A-549 IC50 > 10.0 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone H (29) | marine fungus Aspergillus sp. ZA-01 | Micrococcus lysodeikticus MIC 6.13 μg/mL Bacillus anthraci MIC 12.5 μg/mL Salmonella typhi MIC 6.13 μg/mL Enterobacter aerogenes MIC 6.13 μg/mL | MDA-MB-231 IC50 > 10.0 μM (cisplatin IC50 1.3 μM) MCF-7 IC50 > 10.0 μM (cisplatin IC50 0.97 μM) MGC-803 IC50 > 10.0 μM (cisplatin IC50 1.1 μM) HeLa IC50 > 10.0 μM (cisplatin IC50 0.82 μM) A-549 IC50 > 10.0 μM (cisplatin IC50 0.74 μM) |
| Aspergixanthone I (27) | marine fungus Aspergillus sp. ZA-01 | Vibrio anguillarum MIC 1.56 µM (ciprofloxacin MIC 0.312 µM) Vibrio parahemolyticus MIC 1.56 µM (ciprofloxacin MIC 0.078 µM) Vibrio alginolyticus MIC 3.12 µM (ciprofloxacin MIC 0.625 µM) | |
| Aspergixanthone J (30) | marine fungus Aspergillus sp. ZA-01 | Vibrio anguillarum MIC 6.25 µM (ciprofloxacin MIC 0.312 µM) Vibrio parahemolyticus MIC 25 µM (ciprofloxacin MIC 0.078 µM) Vibrio alginolyticus MIC 25 µM (ciprofloxacin MIC 0.625 µM) | |
| Aspergixanthone K (31) | marine fungus Aspergillus sp. ZA-01 | Vibrio anguillarum MIC 3.12 µM (ciprofloxacin MIC 0.312 µM) Vibrio parahemolyticus MIC 25.0 µM (ciprofloxacin MIC 0.078 µM) Vibrio alginolyticus MIC 12.5 µM (ciprofloxacin MIC 0.625 µM) | |
| Citreamicin θ A (32) | marine Streptomyces sp. | Staphylococcus haemolyticus UST950701-004 MIC 0.5 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus UST950701-005 MIC 1.0 μg/mL (penicillin G MIC 0.25 μg/mL, streptomycin MIC 8.0 μg/mL) Bacillus subtillis 769 MIC 0.25 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus ATCC43300 MIC 0.25 μg/mL (penicillin G MIC > 50 μg/mL, streptomycin MIC > 50 μg/mL) | HeLa IC50 0.055 μg/mL (cisplatin IC50 18.14 μg/mL) |
| Citreamicin θ B (33) | marine Streptomyces sp. | Staphylococcus haemolyticus UST950701-004 MIC 0.5 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus UST950701-005 MIC 1.0 μg/mL (penicillin G MIC 0.25 μg/mL, streptomycin MIC 8.0 μg/mL) Bacillus subtillis 769 MIC 0.25 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus ATCC43300 MIC 0.25 μg/mL (penicillin G MIC > 50 μg/mL, streptomycin MIC > 50 μg/mL) | HeLa IC50 0.072 μg/mL (cisplatin IC50 18.14 μg/mL) |
| Citreaglycon A (35) | marine Streptomyces sp. | Staphylococcus haemolyticus UST950701-004 MIC 8.0 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus UST950701-005 MIC 16.0 μg/mL (penicillin G MIC 0.25 μg/mL, streptomycin MIC 8.0 μg/mL) Bacillus subtillis 769 MIC 8.0 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus ATCC43300 MIC 8.0 μg/mL (penicillin G MIC > 50 μg/mL, streptomycin MIC > 50 μg/mL) | HeLa IC50 > 40 μg/mL (cisplatin IC50 18.14 μg/mL) |
| Dehydrocitreaglycon A (36) | marine Streptomyces sp. | Staphylococcus haemolyticus UST950701-004 MIC 8.0 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus UST950701-005 MIC 16.0 μg/mL (penicillin G MIC 0.25 μg/mL, streptomycin MIC 8.0 μg/mL) Bacillus subtillis 769 MIC > 50 μg/mL (penicillin G MIC 0.13 μg/mL, streptomycin MIC 8.0 μg/mL) Staphylococcus aureus ATCC43300 MIC 0.25 μg/mL (penicillin G MIC > 50 μg/mL, streptomycin MIC > 50 μg/mL) | HeLa IC50 > 40 μg/mL (cisplatin IC50 18.14 μg/mL) |
| Buanmycin (34) | marine Streptomyces sp. | Staphylococcus aureus MIC 10.5 μΜ (ampicillin MIC 4.5 μΜ) Bacillus subtilis MIC 0.7 μM (ampicillin MIC 1.1 μΜ) Kocuria rhizophila MIC 10.5 μM (ampicillin MIC 0.6 μΜ) Salmonella enterica MIC 0.7 μM (ampicillin MIC 4.5 μΜ) Proteus hauseri MIC 21.1 μM (ampicillin MIC < 0.6 μΜ) | A-549 IC50 1.7 μΜ (etoposide IC50 0.8 μΜ) HCT-116 IC50 0.9 μM (etoposide IC50 1.9 μΜ) SNU-638 IC50 0.8 μM (etoposide IC50 0.5 μΜ) K-562 IC50 > 100 μM (etoposide IC50 3.1 μΜ) SK-HEP1 IC50 1.9 μM (etoposide IC50 1.1 μΜ) MDA-MB-231 IC50 1.2 μM (etoposide IC50 10.6 μΜ) |
| Compound (7) | marine fungus Phomopsis sp. (No. SK7RN3G1) | Hep-2 IC50 8 μg/mL HepG2 IC50 9 μg/mL | |
| Versixanthone A (39) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 2.6 μM (doxorubicin hydrochloride IC50 0.02 μM) K562 IC50 7.1 μM (doxorubicin hydrochloride IC50 0.3 μM) A549 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.2 μM) H1975 IC50 11.2 μM (doxorubicin hydrochloride IC50 0.8 μM) 803 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.2 μM) HO-8910 IC50 10.1 μM (doxorubicin hydrochloride IC50 0.5 μM) | |
| Versixanthone B (40) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 9.9 μM (doxorubicin hydrochloride IC50 0.02 μM) A549 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.2 μM) H1975 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.8 μM) 803 IC50 21.6 μM (doxorubicin hydrochloride IC50 0.2 μM) HO-8910 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.5 μM) | |
| Versixanthone C (41) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 7.8 μM (doxorubicin hydrochloride IC50 0.02 μM) K562 IC50 18.2 μM (doxorubicin hydrochloride IC50 0.3 μM) A549 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.2 μM) H1975 IC50 25.6 μM (doxorubicin hydrochloride IC50 0.8 μM) HO-8910 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.5 μM) HCT-116 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.2 μM) | |
| Versixanthone D (42) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 3.3 μM (doxorubicin hydrochloride IC50 0.02 μM) K562 IC50 9.1 μM (doxorubicin hydrochloride IC50 0.3 μM) A549 IC50 12.7 μM (doxorubicin hydrochloride IC50 0.2 μM) H1975 IC50 > 50 μM (doxorubicin hydrochloride IC50 0.8 μM) 803 IC50 9.8 μM (doxorubicin hydrochloride IC50 0.2 μM) HO-8910 IC50 13.9 μM (doxorubicin hydrochloride IC50 0.5 μM) HCT-116 IC50 6.1 μM (doxorubicin hydrochloride IC50 0.2 μM) | |
| Versixanthone E (43) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 1.6 μM (doxorubicin hydrochloride IC50 0.02 μM) K562 IC50 11.1 μM (doxorubicin hydrochloride IC50 0.3 μM) H1975 IC50 2.7 μM (doxorubicin hydrochloride IC50 0.8 μM) 803 IC50 2.2 μM (doxorubicin hydrochloride IC50 0.2 μM) HO-8910 IC50 2.0 μM (doxorubicin hydrochloride IC50 0.5 μM) | |
| Versixanthone F (44) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 2.7 μM (doxorubicin hydrochloride IC50 0.02 μM) K562 IC50 6.7 μM (doxorubicin hydrochloride IC50 0.3 μM) A549 IC50 10.6 μM (doxorubicin hydrochloride IC50 0.2 μM) HO-8910 IC50 20.8 μM (doxorubicin hydrochloride IC50 0.5 μM) HCT-116 IC50 0.7 μM (doxorubicin hydrochloride IC50 0.2 μM) | |
| Versixanthone G (47) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 13.4 μM K562 IC50 20.9 μM A549 IC50 17.8 μM H1975 IC50 9.8 μM MGC803 IC50 4.6 μM HEK293 IC50 > 50 μM HO-8910 IC50 9.6 μM HCT-116 IC50 16.2 μM | |
| Versixanthone H (48) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 6.9 μM K562 IC50 22.1 μM A549 IC50 19.2 μM H1975 IC50 5.3 μM MGC803 IC50 6.2 μM HEK293 IC50 > 50 μM HO-8910 IC50 6.9 μM HCT-116 IC50 15.2 μM | |
| Versixanthone I (49) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 27.8 μM K562 IC50 >50 μM A549 IC50 > 50 μM H1975 IC50 > 50 μM HEK293 IC50 > 50 μM HO-8910 IC50 > 50 μM HCT-116 IC50 > 50 μM | |
| Versixanthone J (50) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 47.3 μM K562 IC50 > 50 μM A549 IC50 > 50 μM H1975 IC50 > 50 μM HEK293 IC50 > 50 μM HO-8910 IC50 > 50 μM HCT-116 IC50 > 50 μM | |
| Versixanthone K (51) | fungus Aspergillus versicolor HDN1009 found in soil around a mangrove | HL-60 IC50 49.5 μM K562 IC50 > 50 μM A549 IC50 > 50 μM H1975 IC50 49.5 μM MGC803 IC50 > 50 μM HEK293 IC50 > 50 μM HO-8910 IC50 > 50 μM HCT-116 IC50 > 50 μM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veríssimo, A.C.S.; Pinto, D.C.G.A.; Silva, A.M.S. Marine-Derived Xanthone from 2010 to 2021: Isolation, Bioactivities and Total Synthesis. Mar. Drugs 2022, 20, 347. https://doi.org/10.3390/md20060347
Veríssimo ACS, Pinto DCGA, Silva AMS. Marine-Derived Xanthone from 2010 to 2021: Isolation, Bioactivities and Total Synthesis. Marine Drugs. 2022; 20(6):347. https://doi.org/10.3390/md20060347
Chicago/Turabian StyleVeríssimo, Ana C. S., Diana C. G. A. Pinto, and Artur M. S. Silva. 2022. "Marine-Derived Xanthone from 2010 to 2021: Isolation, Bioactivities and Total Synthesis" Marine Drugs 20, no. 6: 347. https://doi.org/10.3390/md20060347
APA StyleVeríssimo, A. C. S., Pinto, D. C. G. A., & Silva, A. M. S. (2022). Marine-Derived Xanthone from 2010 to 2021: Isolation, Bioactivities and Total Synthesis. Marine Drugs, 20(6), 347. https://doi.org/10.3390/md20060347