
Docosahexaenoic Acid-Acylated Astaxanthin Esters Exhibit Superior Renal Protective Effect to Recombination of Astaxanthin with DHA via Alleviating Oxidative Stress Coupled with Apoptosis in Vancomycin-Treated Mice with Nephrotoxicity



Abstract
:1. Introduction
2. Results
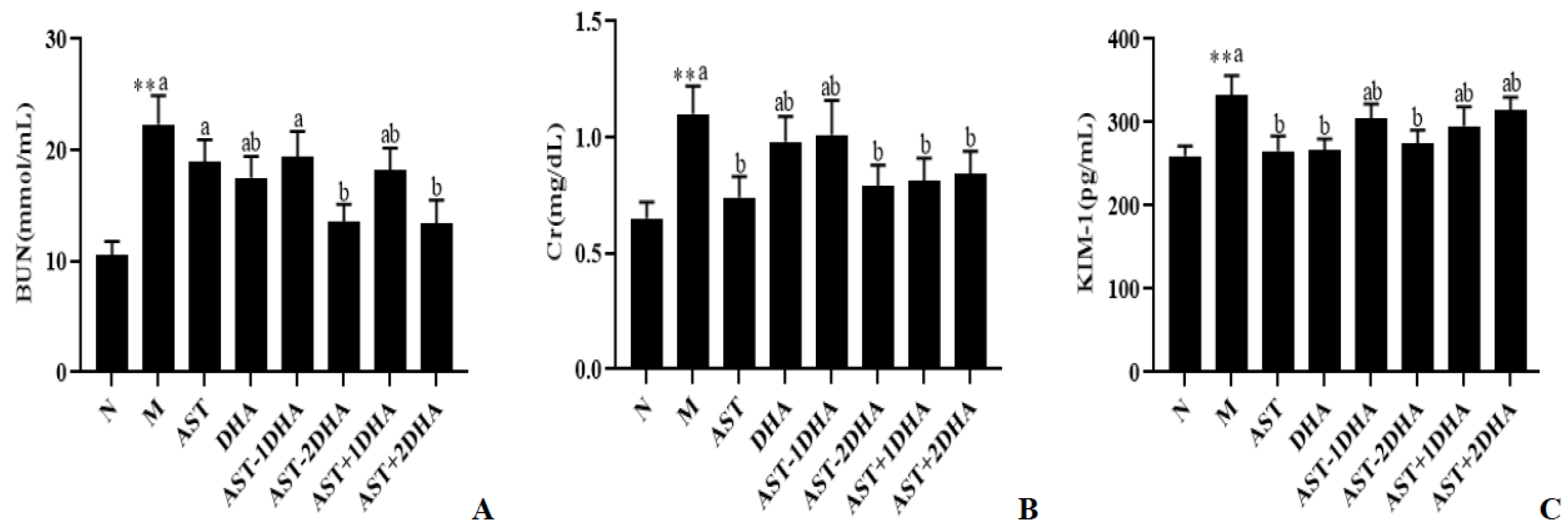
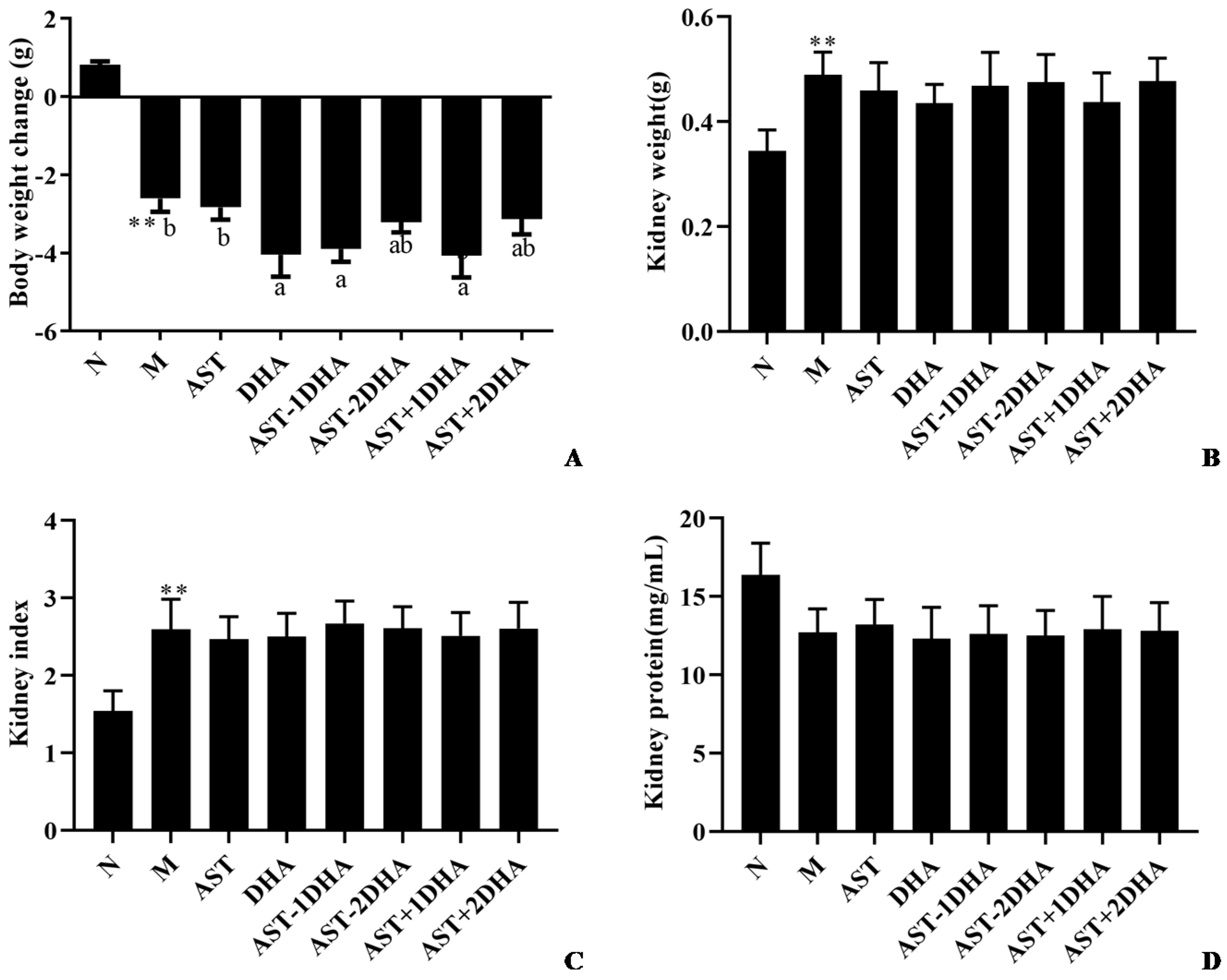
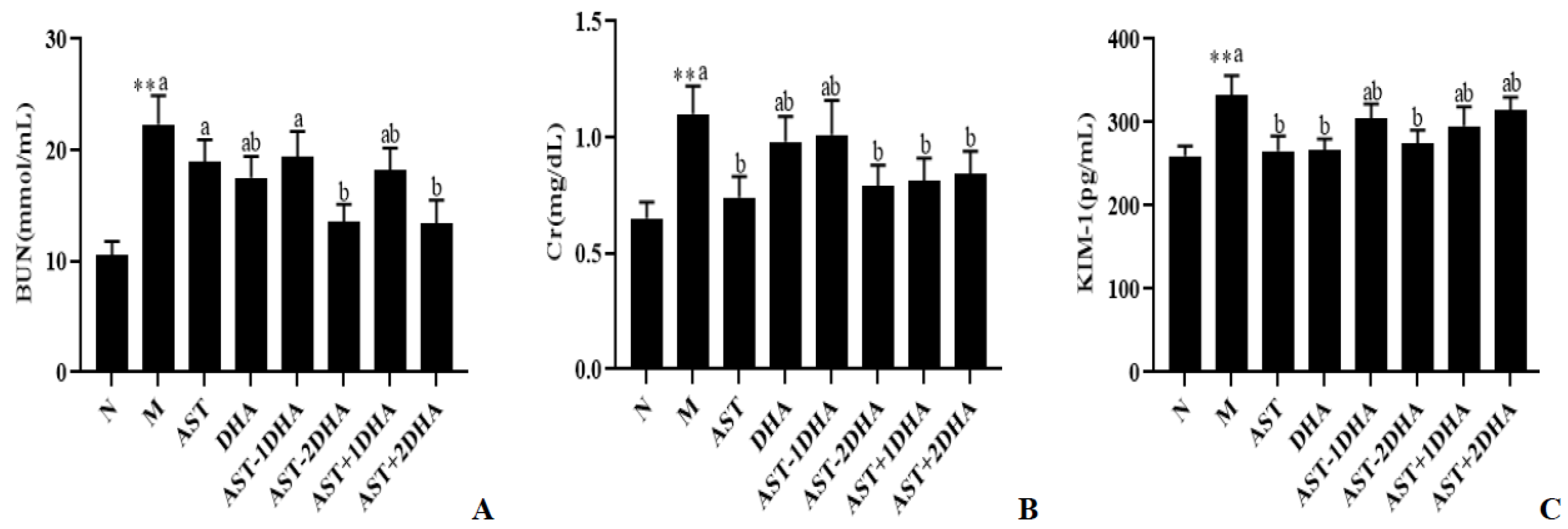
2.1. Effects on Vancomycin-Induced Kidney Dysfunction
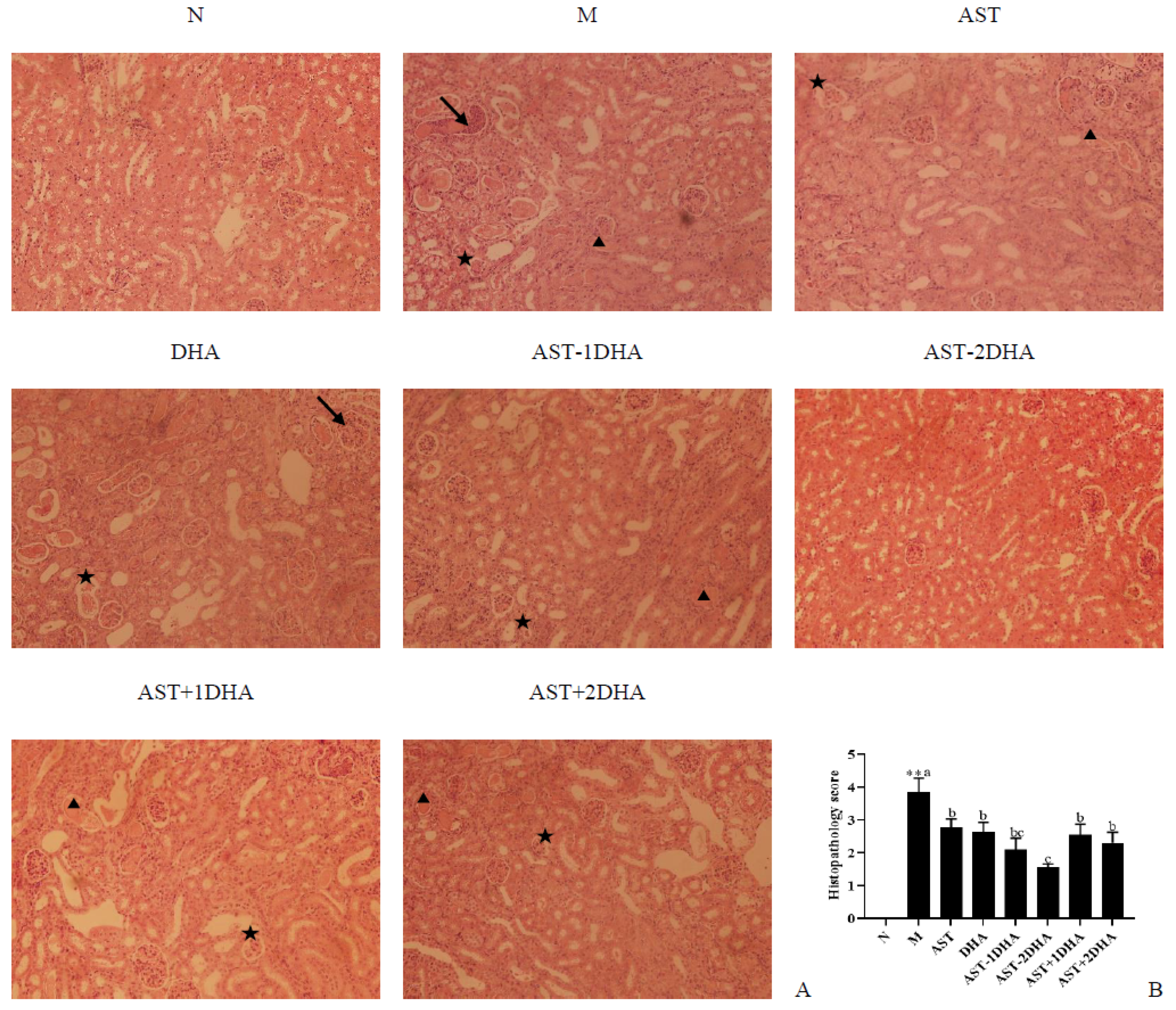
2.2. Effects of Different Samples on Vancomycin-Induced Renal Pathological Change
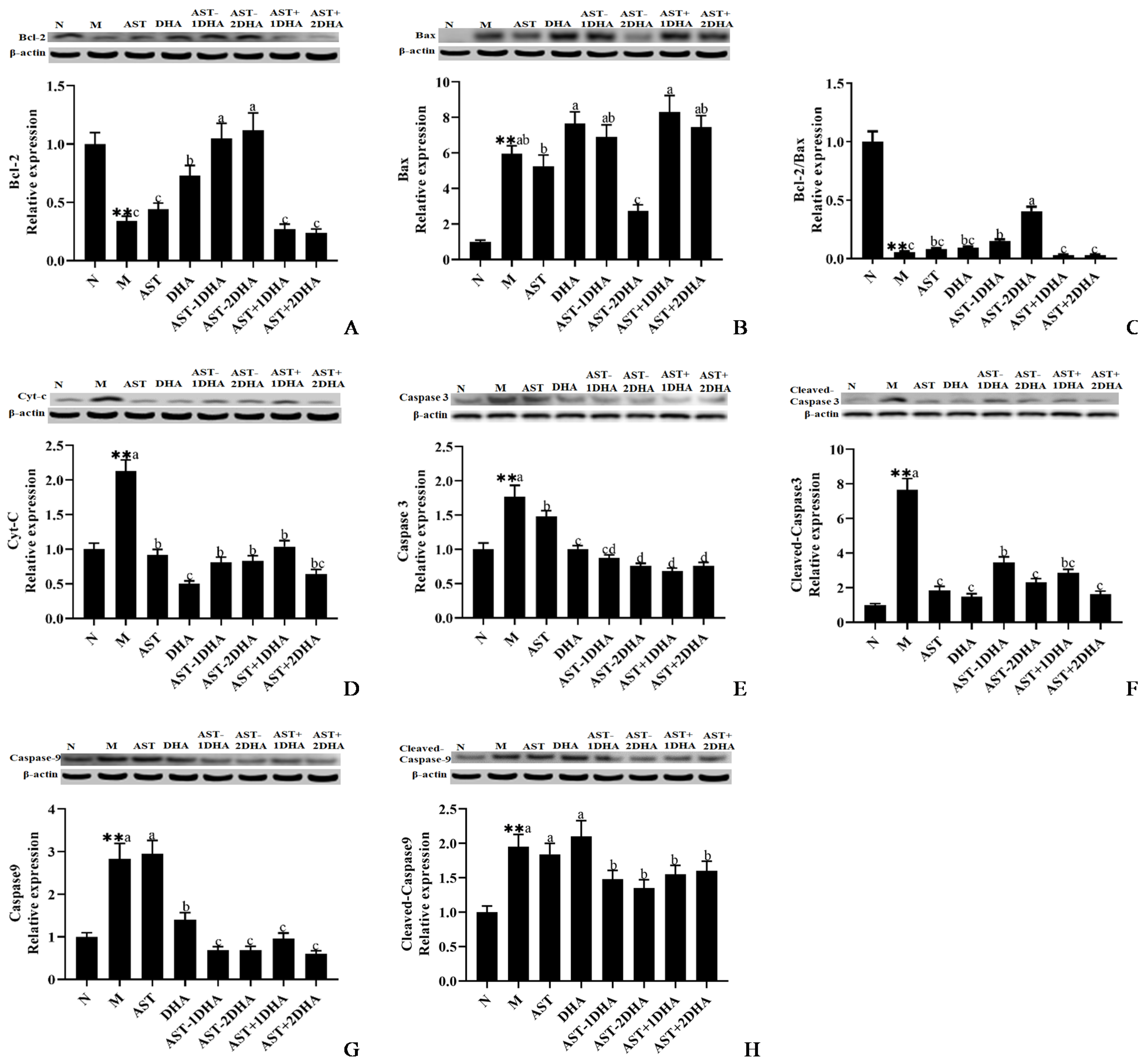
2.3. Effects on Vancomycin-Induced Activation of The Mitochondrial Apoptosis Pathway
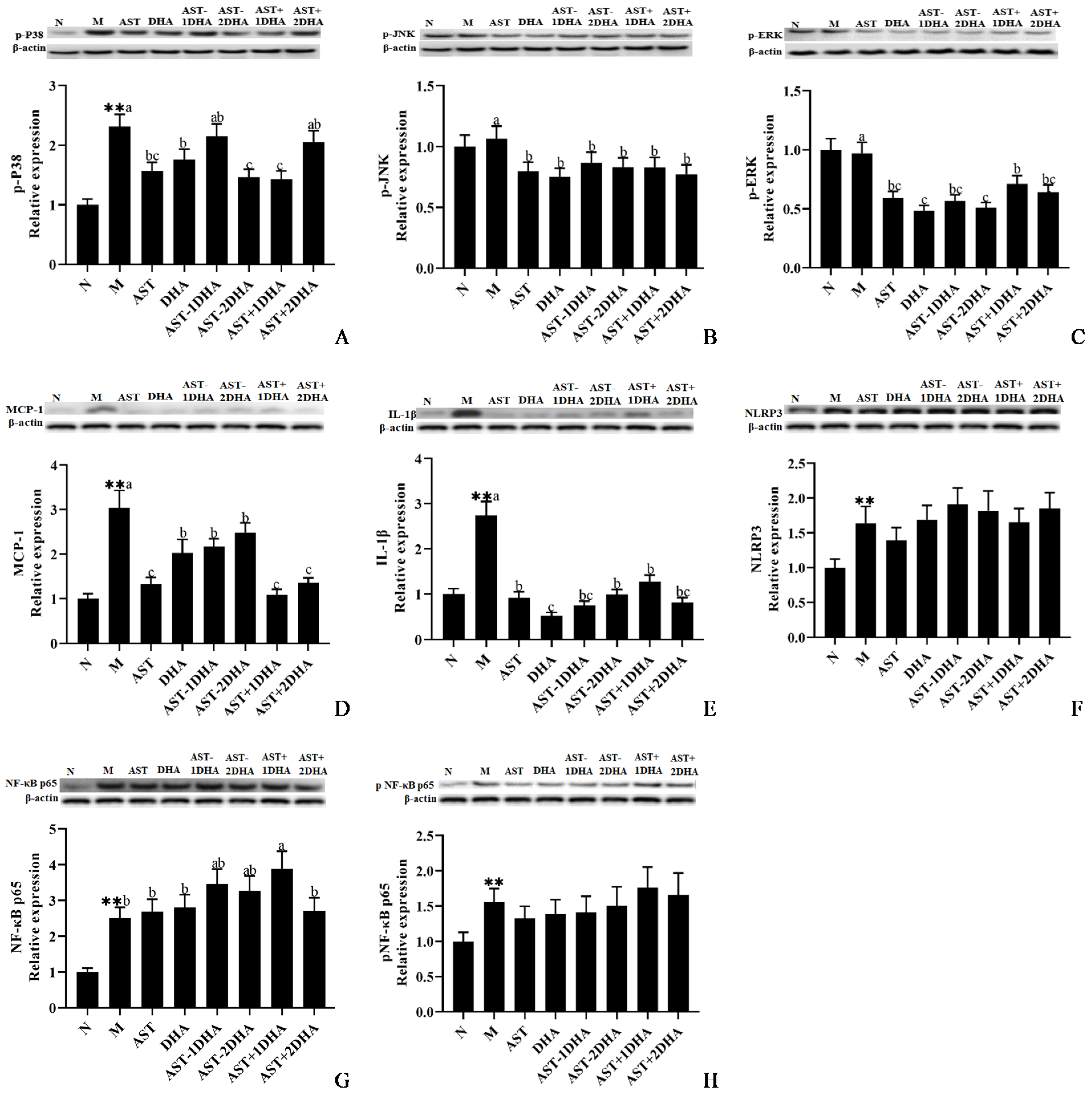
2.4. Effects on Vancomycin-Induced Activation of MAPKs Signal Pathway
2.5. Effects on Vancomycin-Induced Inflammatory Pathway
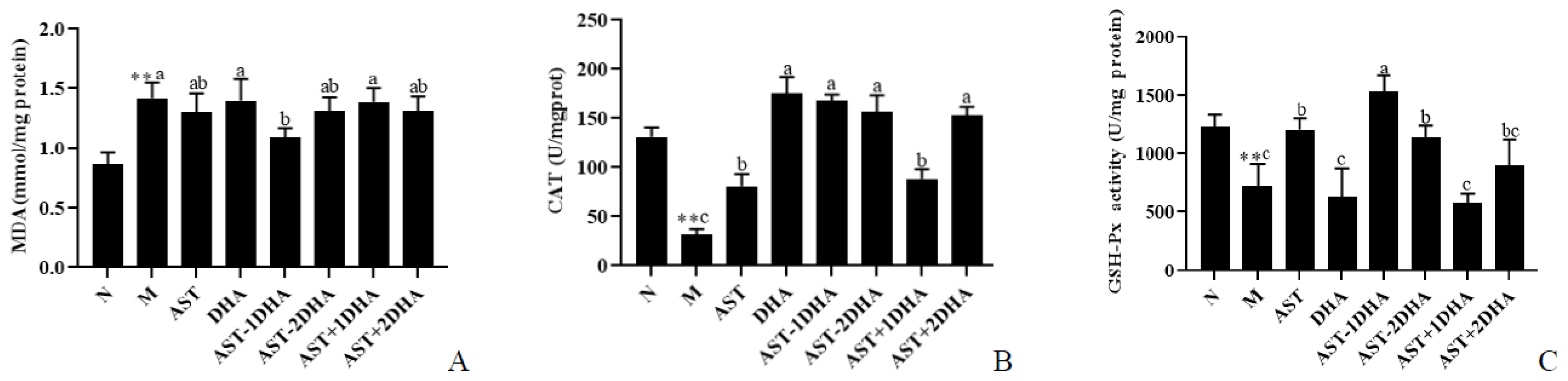
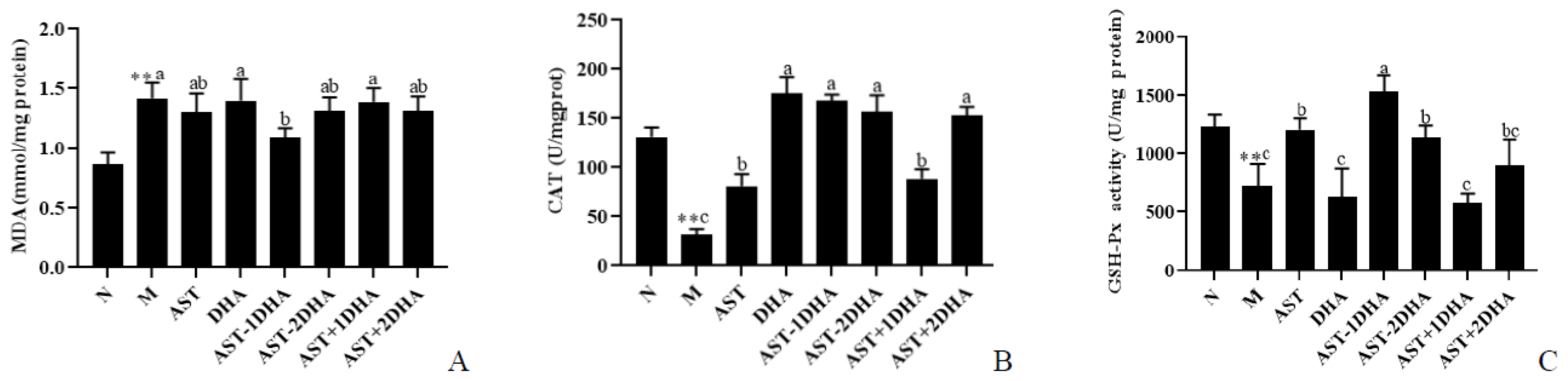
2.6. Effects on Vancomycin-Induced Oxidative Stress
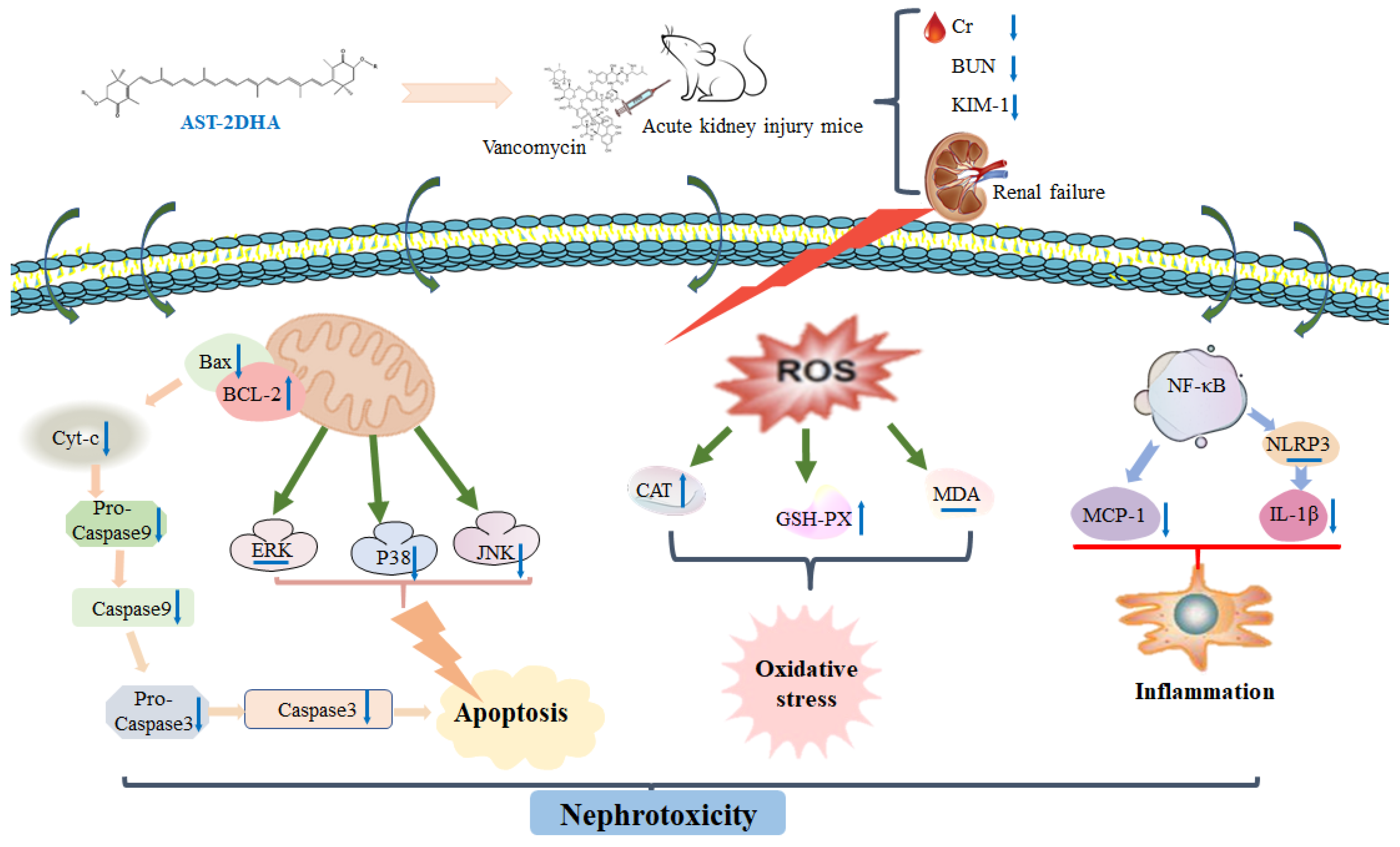
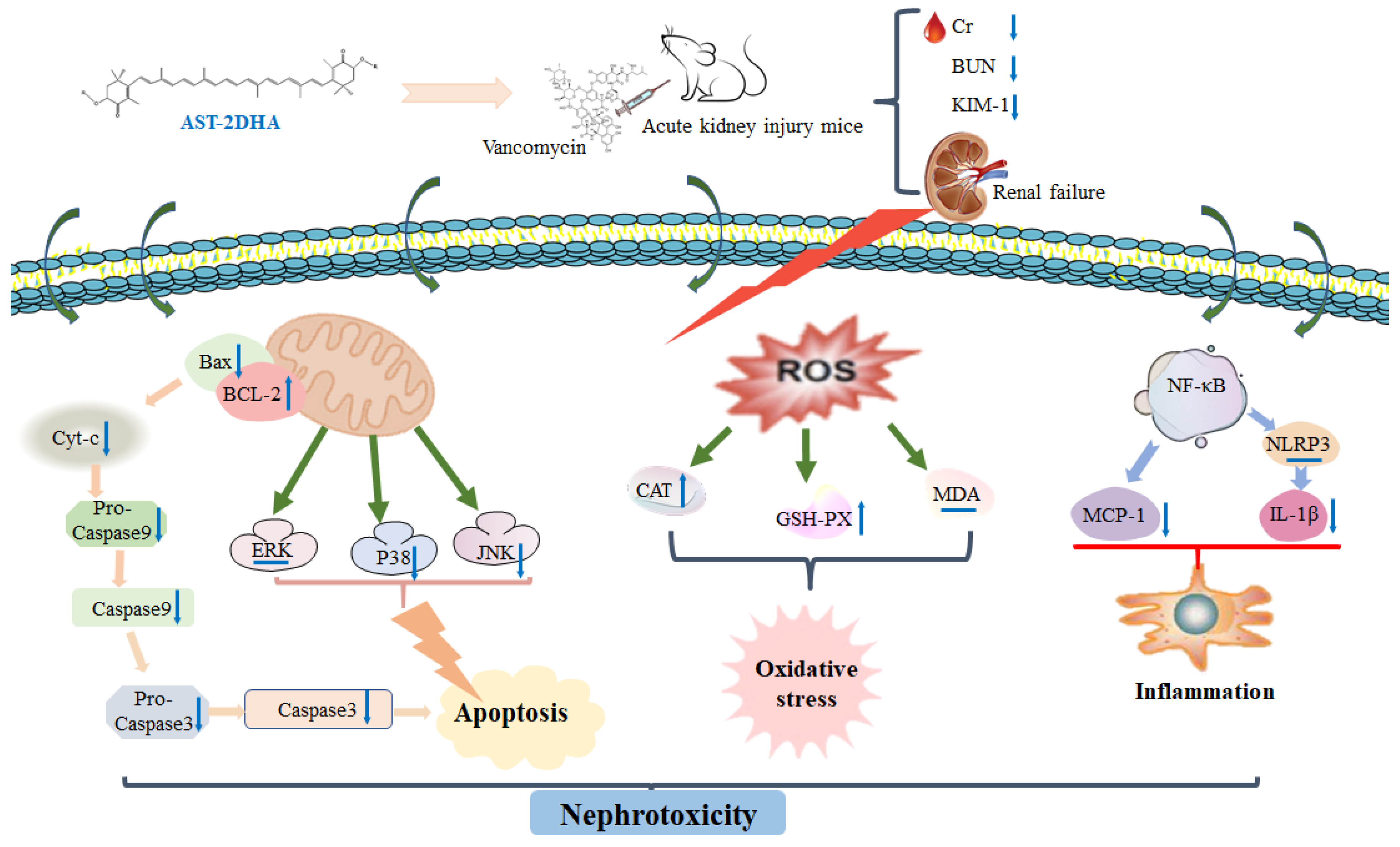
3. Discussion
4. Materials and methods
4.1. Chemicals
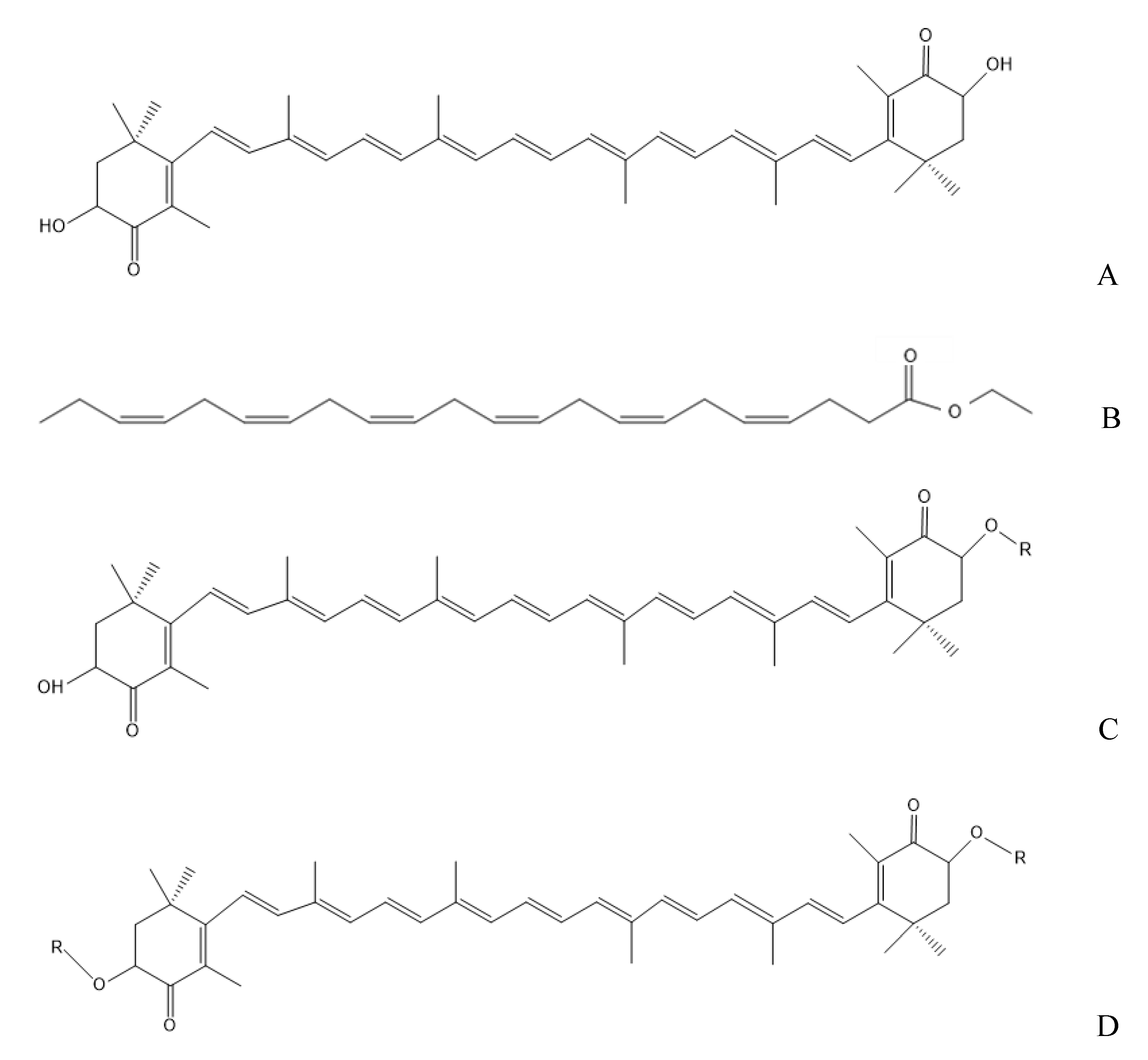
4.2. Preparation and Determination of DHA-Acylated AST Esters
4.3. Animals and Treatment
4.4. Biochemical Assays
4.5. Histological Analysis
4.6. Western Blotting Assessment
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hoste, E.A.J.; Kellum, J.A.; Selby, N.M.; Zarbock, A.; Palevsky, P.M.; Bagshaw, S.M.; Goldstein, S.L.; Cerda, J.; Chawla, L.S. Global epidemiology and outcomes of acute kidney injury. Nat. Rev. Nephrol. 2018, 14, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Mody, H.; Ramakrishnan, V.; Chaar, M.; Lezeau, J.; Rump, A.; Taha, K.; Lesko, L.; Ait-Oudhia, S. A Review on Drug-Induced Nephrotoxicity: Pathophysiological Mechanisms, Drug Classes, Clinical Management, and Recent Advances in Mathematical Modeling and Simulation Approaches. Clin. Pharmacol. Drug Dev. 2020, 9, 896–909. [Google Scholar] [CrossRef]
- Bamgbola, O. Review of vancomycin-induced renal toxicity: An update. Ther. Adv. Endocrinol. Metab. 2016, 7, 136–147. [Google Scholar] [CrossRef]
- Ahmida, M.H.S.; Pathology, T. Protective role of curcumin in nephrotoxic oxidative damage induced by vancomycin in rats. Exp. Toxicol. Pathol. 2012, 64, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Fu, G.; Shen, J.; Shen, K.; Xu, Z.; Wang, Y.; Jin, B.; Pan, H. Ameliorative Effect of Daidzein on Cisplatin-Induced Nephrotoxicity in Mice via Modulation of Inflammation, Oxidative Stress, and Cell Death. Oxid. Med. Cell. Longevity 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications-A Review. Mar. Drugs. 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Zuluaga, M.; Gueguen, V.; Letourneur, D.; Pavon-Djavid, G. Astaxanthin-antioxidant impact on excessive Reactive Oxygen Species generation induced by ischemia and reperfusion injury. Chem.-Biol. Interact. 2018, 279, 145–158. [Google Scholar] [CrossRef]
- Qiu, X.; Fu, K.; Zhao, X.; Zhang, Y.; Yuan, Y.; Zhang, S.; Gu, X.; Guo, H. Protective effects of astaxanthin against ischemia/reperfusion induced renal injury in mice. J. Transl. Med. 2015, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Yang, L.; Xu, J.; Qiao, X.; Li, Z.; Wang, Y.; Xue, C. Evaluation of the physicochemical stability and digestibility of microencapsulated esterified astaxanthins using in vitro and in vivo models. Food Chem. 2018, 260, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Gu, J.; Luan, T.; Qiao, X.; Cao, Y.; Xue, C.; Xu, J. Influence of oil matrixes on stability, antioxidant activity, bioaccessibility and bioavailability of astaxanthin ester. J. Sci. Food Agric. 2020, 101, 1609–1617. [Google Scholar] [CrossRef]
- Shubha, P.; Medha, P.; Arivarasu, N.A.; Neelam, F.; Sheeba, K.; Khan, S.A.; Md Wasim, K.; Yusufi, A.N.K. Studies on the protective effect of dietary fish oil on gentamicin-induced nephrotoxicity and oxidative damage in rat kidney. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 35–44. [Google Scholar]
- Shi, H.; Zou, J.; Zhang, T.; Che, H.; Gao, X.; Wang, C.; Wang, Y.; Xue, C. Protective Effects of DHA-PC against Vancomycin-Induced Nephrotoxicity through the Inhibition of Oxidative Stress and Apoptosis in BALB/c Mice. J. Agric. Food Chem. 2018, 66, 475–484. [Google Scholar] [CrossRef]
- Che, H.; Li, Q.; Zhang, T.; Wang, D.; Yang, L.; Xu, J.; Yanagita, T.; Xue, C.; Chang, Y.; Wang, Y.M. The effects of astaxanthin and docosahexaenoic acid-acylated astaxanthin on Alzheimer’s disease in APP/PS1 double transgenic mice. J. Agric. Food Chem. 2018, 66, 4948–4957. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Shi, H.H.; Xu, J.; Yanagita, T.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Docosahexaenoic acid-acylated astaxanthin ester exhibits superior performance over non-esterified astaxanthin in preventing behavioral deficits coupled with apoptosis in MPTP-induced mice with Parkinson’s disease. Food Funct. 2020, 11, 8038–8050. [Google Scholar] [CrossRef]
- Stoyanoff, T.R.; Todaro, J.S.; Aguirre, M.V.; Zimmermann, M.C.; Brandan, N.C.J.T. Amelioration of lipopolysaccharide-induced acute kidney injury by erythropoietin: Involvement of mitochondria-regulated apoptosis. Prostag. Leukotr. Ess. 2014, 318, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Suchal, K.; Gamad, N.; Dinda, A.K.; Arya, D.S.; Bhatia, J. Telmisartan ameliorates cisplatin-induced nephrotoxicity by inhibiting MAPK mediated inflammation and apoptosis. Eur. J. Pharmacol. 2015, 748, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.A.; Edrees, G.M.; El-Gamel, E.M.; El-Sayed, E.A. Amelioration of cisplatin-induced nephrotoxicity by grape seed extract and fish oil is mediated by lowering oxidative stress and DNA damage. Cytotechnology 2014, 66, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Rad, A.K.; Mohebbati, R.; Hosseinian, S. Drug-induced Nephrotoxicity and Medicinal Plants. Iran. J. Kidney. Dis. 2017, 11, 169–179. [Google Scholar]
- Kharroubi, W.; Dhibi, M.; Mekni, M.; Haouas, Z.; Chreif, I.; Neffati, F.; Hammami, M.; Skly, R. Sodium arsenate induce changes in fatty acids profiles and oxidative damage in kidney of rats. Environ. Sci. Pollut. Res. 2014, 21, 12040–12049. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, J.; Xu, J.; Che, H.; Wang, Y.; Xue, C.; Huang, Q.; Zhang, T. Comparison of the Digestion and Absorption Characteristics of Docosahexaenoic Acid-Acylated Astaxanthin Monoester and Diester in Mice. J. Ocean. Univ. China 2021, 20, 973–984. [Google Scholar] [CrossRef]
- Gong, J.; Noel, S.; Pluznick, J.L.; Hamad, R.A.; Rabb, H. Gut Microbiota-Kidney Cross-Talk in Acute Kidney Injury. Semin. Nephrol. 2019, 39, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Wei, W.; Huang, J.; Liu, X.; Ci, X. Isoorientin Attenuates Cisplatin-Induced Nephrotoxicity Through the Inhibition of Oxidative Stress and Apoptosis via Activating the SIRT1/SIRT6/Nrf-2 Pathway. Front. Pharmacol. 2020, 11, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, L.; Wen, M.; Du, L.; Gao, X.; Xue, C.; Xu, J.; Wang, Y. Enhanced neuroprotective effect of DHA and EPA-enriched phospholipids against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) induced oxidative stress in mice brain. J. Funct. Foods 2016, 25, 385–396. [Google Scholar] [CrossRef]
- Ajami, M.; Davoodi, S.H.; Habibey, R.; Namazi, N.; Soleimani, M.; Pazoki-Toroudi, H. Effect of DHA plus EPA on oxidative stress and apoptosis induced by ischemia-reperfusion in rat kidneys. Fundam. Clin. Pharmacol. 2013, 27, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, E.; Abdel-Wahab, B.A.; Ali, F.; El-Ghafar, O.; Sharkawi, S. Trans-ferulic acid ameliorates cisplatin-induced testicular damage via suppression of TLR4, P38-MAPK, and ERK1/2 signaling pathways. Environ. Sci. Pollut. Res. 2021, 28, 41948–41964. [Google Scholar] [CrossRef] [PubMed]
- Shuang, W.A.; Shan, T.A.; Xuan, C.A.; Xin, L.A.; Shuang, J.; Hpla, B.; Phj, A.; Mjsa, B.; Peng, D. Pulchinenoside B4 exerts the protective effects against cisplatin-induced nephrotoxicity through NF-κB and MAPK mediated apoptosis signaling pathways in mice. Chem.-Biol. Interact. 2020, 331, 109233. [Google Scholar]
- Che, H.; Zhou, M.; Zhang, T.; Zhang, L.; Ding, L.; Yanagita, T.; Xu, J.; Xue, C.; Wang, Y. Comparative study of the effects of phosphatidylcholine rich in DHA and EPA on Alzheimer’s disease and the possible mechanisms in CHO-APP/PS1 cells and SAMP8 mice. Food Funct. 2018, 9, 643–654. [Google Scholar] [CrossRef]
- Li, J.; Wang, F.; Xia, Y.; Dai, W.; Guo, C. Astaxanthin Pretreatment Attenuates Hepatic Ischemia Reperfusion-Induced Apoptosis and Autophagy via the ROS/MAPK Pathway in Mice. Mar. Drugs. 2015, 13, 3368–3387. [Google Scholar] [CrossRef] [Green Version]
- Arunkumar, E.; Bhuvaneswari, S.; Anuradha, C. An intervention study in obese mice with astaxanthin, a marine carotenoid—effects on insulin signaling and pro-inflammatory cytokines. Food Funct. 2012, 7, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Sun, L.; Niu, X.T.; Chen, X.M.; Tian, J.X.; Kong, Y.D.; Wang, G.Q. Astaxanthin protects lipopolysaccharide-induced inflammatory response in Channa argus through inhibiting NF-kappaB and MAPKs signaling pathways. Fish Shellfish Immunol. 2019, 86, 280–286. [Google Scholar] [CrossRef]
- Mehmet, K.F.; Serkan, Y.; Sefa, K.; Cuneyt, C.; Amdia, M.; Bahaeddin, D.M. Therapeutic efficacy of zingerone against vancomycin-induced oxidative stress, inflammation, apoptosis and aquaporin 1 permeability in rat kidney. Biomed. Pharmacother. 2018, 105, 981–991. [Google Scholar]
- Zhang, J.; Zhang, S.; Bi, J.; Gu, J.; Deng, Y.; Liu, C. Astaxanthin pretreatment attenuates acetaminophen-induced liver injury in mice. Int. Immunopharmacol. 2017, 45, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Matejovic, M.; Ince, C.; Chawla, L.S.; Blantz, R.; Molitoris, B.A.; Rosner, M.H.; Okusa, M.D.; Kellum, J.A.; Ronco, C. Renal Hemodynamics in AKI: In Search of New Treatment Targets. J. Am. Soc. Nephrol. 2016, 27, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.; Wu, M.; Qin, J.; Yang, J.; Ye, C.; Wang, C. Magnesium lithospermate B improves renal hemodynamics and reduces renal oxygen consumption in 5/6th renal ablation/infarction rats. BMC Nephrol. 2019, 20, 49. [Google Scholar] [CrossRef]
- Yang, L.; Qiao, X.; Liu, J.; Wu, L.; Cao, Y.; Xu, J.; Xue, C. Preparation, characterization and antioxidant activity of astaxanthin esters with different molecular structures. J. Sci. Food Agric. 2020, 101, 2576–2583. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Abdelrahman, A.; Al Suleimani, Y.; Manoj, P.; Ali, H.; Nemmar, A.; Al Za’abi, M. Effect of concomitant treatment of curcumin and melatonin on cisplatin-induced nephrotoxicity in rats. Biomed. Pharmacother. 2020, 131, 110761. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (mg/g) | N | M | AST | DHA | AST-1DHA | AST-2DHA | AST+1DHA | AST+2DHA |
|---|---|---|---|---|---|---|---|---|
| DHA | 3.46 ± 0.41 | 1.06 ± 0.15c ** | 1.13 ± 0.09c | 1.36 ± 0.11b | 1.42 ± 0.13b | 1.96 ± 0.21a | 1.26 ± 0.09bc | 1.76 ± 0.15a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.-H.; Guo, Y.; Chen, L.-P.; Wang, C.-C.; Huang, Q.-R.; Xue, C.-H.; Wang, Y.-M.; Zhang, T.-T. Docosahexaenoic Acid-Acylated Astaxanthin Esters Exhibit Superior Renal Protective Effect to Recombination of Astaxanthin with DHA via Alleviating Oxidative Stress Coupled with Apoptosis in Vancomycin-Treated Mice with Nephrotoxicity. Mar. Drugs 2021, 19, 499. https://doi.org/10.3390/md19090499
Shi H-H, Guo Y, Chen L-P, Wang C-C, Huang Q-R, Xue C-H, Wang Y-M, Zhang T-T. Docosahexaenoic Acid-Acylated Astaxanthin Esters Exhibit Superior Renal Protective Effect to Recombination of Astaxanthin with DHA via Alleviating Oxidative Stress Coupled with Apoptosis in Vancomycin-Treated Mice with Nephrotoxicity. Marine Drugs. 2021; 19(9):499. https://doi.org/10.3390/md19090499
Chicago/Turabian StyleShi, Hao-Hao, Ying Guo, Li-Pin Chen, Cheng-Cheng Wang, Qing-Rong Huang, Chang-Hu Xue, Yu-Ming Wang, and Tian-Tian Zhang. 2021. "Docosahexaenoic Acid-Acylated Astaxanthin Esters Exhibit Superior Renal Protective Effect to Recombination of Astaxanthin with DHA via Alleviating Oxidative Stress Coupled with Apoptosis in Vancomycin-Treated Mice with Nephrotoxicity" Marine Drugs 19, no. 9: 499. https://doi.org/10.3390/md19090499
APA StyleShi, H.-H., Guo, Y., Chen, L.-P., Wang, C.-C., Huang, Q.-R., Xue, C.-H., Wang, Y.-M., & Zhang, T.-T. (2021). Docosahexaenoic Acid-Acylated Astaxanthin Esters Exhibit Superior Renal Protective Effect to Recombination of Astaxanthin with DHA via Alleviating Oxidative Stress Coupled with Apoptosis in Vancomycin-Treated Mice with Nephrotoxicity. Marine Drugs, 19(9), 499. https://doi.org/10.3390/md19090499