
Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders


 and
and 
Abstract
1. Introduction
2. Results
2.1. Amino Acid Analysis
2.2. Evaluation of Protein Quality
2.3. Macronutrients and Further Components
2.4. Fatty Acid Distribution
2.5. Minerals, Trace Elements, and Heavy Metals
2.6. Analysis of Vitamins
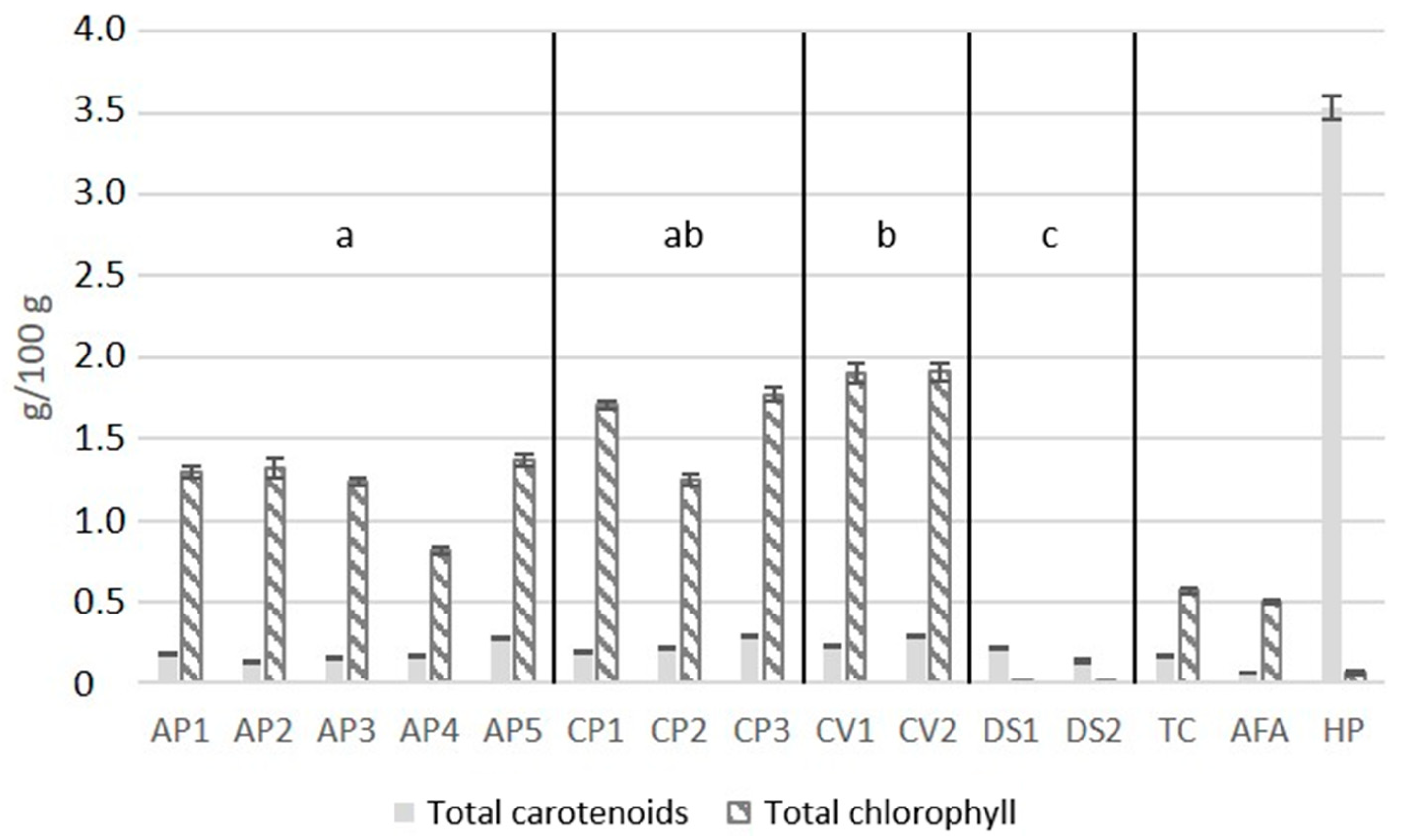
2.7. Total Chlorophylls and Carotenoids
3. Discussion
4. Materials and Methods
4.1. Microalgae Powders
4.2. Amino Acids and Ammonium Quantification
4.3. N-Factor Calculation
4.4. Quantification of Macronutrients
4.5. Lipid Extraction and Fatty Acid Analysis
4.6. Quantification of Minerals, Trace Elements, and Heavy Metals
4.7. Analysis of Vitamin Content
4.8. Total Carotenoids and Chlorophylls
- Chlaa = Concentration of Chlorophyll a
- Chlb = Concentration of Chlorophyll b
- E = Emission
- C = Concentration
- V = Volume
- m = Mass
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Wiley Online Library: Hoboken, NJ, USA, 2004; Volume 577. [Google Scholar]
- Di Lena, G.; Casini, I.; Lucarini, M.; Lombardi-Boccia, G. Carotenoid profiling of five microalgae species from large-scale production. Food Res. Int. 2019, 120, 810–818. [Google Scholar] [CrossRef]
- Grewe, C.; Griehl, C. Time- and media-dependent secondary carotenoid accumulation in Haematococcus pluvialis. Biotechnol. J. 2008, 3, 1232–1244. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, F.; Sun, Z.; Sun, P.; Chen, T.; Chen, F. Microalgal carotenoids: Beneficial effects and potential in human health. Food Funct. 2014, 5, 413–425. [Google Scholar] [CrossRef]
- Brown, M.R.; Mular, M.; Miller, I.; Farmer, C.; Trenerry, C. The vitamin content of microalgae used in aquaculture. Environ. Biol. Fishes 1999, 11, 247–255. [Google Scholar] [CrossRef]
- Fabregas, J.; Herrero, C. Vitamin content of four marine microalgae. Potential use as source of vitamins in nutrition. J. Ind. Microbiol. Biotechnol. 1990, 5, 259–263. [Google Scholar] [CrossRef]
- Fábregas, J.; Herrero, C. Marine microalgae as a potential source of minerals in fish diets. Aquaculture 1986, 51, 237–243. [Google Scholar] [CrossRef]
- Breuer, G.; Evers, W.A.C.; De Vree, J.H.; Kleinegris, D.M.M.; Martens, D.E.; Wijffels, R.H.; Lamers, P.P. Analysis of Fatty Acid Content and Composition in Microalgae. J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [PubMed]
- Pierre, G.; Delattre, C.; Dubessay, P.; Jubeau, S.; Vialleix, C.; Cadoret, J.-P.; Probert, I.; Michaud, P. What is in store for eps microalgae in the next decade? Molecules 2019, 24, 4296. [Google Scholar] [CrossRef] [PubMed]
- The European Commission. Commission implementing regulation (eu) 2017/2470 of 20 december 2017 establishing the union list of novel foods in accordance with regulation (eu) 2015/2283 of the european parliament and of the council on novel foods. Off. J. Eur. Union 2017, 351, 72. [Google Scholar]
- Ermis, H.; Guven-Gulhan, U.; Cakir, T.; Altinbas, M. Effect of iron and magnesium addition on population dynamics and high value product of microalgae grown in anaerobic liquid digestate. Sci. Rep. 2020, 10, 3510. [Google Scholar] [CrossRef]
- Loganathan, B.G.; Orsat, V.; Lefsrud, M.; Wu, B.S. A comprehensive study on the effect of light quality imparted by light-emitting diodes (LEDs) on the physiological and biochemical properties of the microalgal consortia of Chlorella variabilis and Scenedesmus obliquus cultivated in dairy wastewater. Bioprocess Biosyst. Eng. 2020, 43, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Michelon, W.; Da Silva, M.L.B.; Mezzari, M.P.; Pirolli, M.; Prandini, J.M.; Soares, H.M. Effects of Nitrogen and Phosphorus on Biochemical Composition of Microalgae Polyculture Harvested from Phycoremediation of Piggery Wastewater Digestate. Appl. Biochem. Biotechnol. 2016, 178, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Block, R.J.; Mitchell, H.H. Correlation of the amino acid composition of proteins with their nutritive value. Nutr. Abstr. Rev. 1946, 16, 249–278. [Google Scholar]
- WHO. Energy and Protein Requirements: Report of a Joint FAO/WHO/UNU Expert Consultation [Held in Rome from 5 to 17 October 1981]; World Health Organization: Geneva, Switzerland, 1985. [Google Scholar]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Marquez, U.M.L.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae: Calculation of new nitrogen-to-protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Rzymski, P.; Budzulak, J.; Niedzielski, P.; Klimaszyk, P.; Proch, J.; Kozak, L.; Poniedzialek, B. Essential and toxic elements in commercial microalgal food supplements. J. Appl. Phycol. 2019, 31, 3581. [Google Scholar] [CrossRef]
- Brown, M.R. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Da Silva Gorgonio, C.M.; Aranda, D.A.G.; Couri, S. Morphological and chemical aspects of Chlorella pyrenoidosa, Dunaliella tertiolecta, Isochrysis galbana and Tetraselmis gracilis microalgae. Nat. Sci. 2013, 05, 783–791. [Google Scholar] [CrossRef]
- Seville, P.; Learoyd, T.; Li, H.-Y.; Williamson, I.; Birchall, J. Amino acid-modified spray-dried powders with enhanced aerosolisation properties for pulmonary drug delivery. Powder Technol. 2007, 178, 40–50. [Google Scholar] [CrossRef]
- Takagi, H.; Sakai, K.; Morida, K.; Nakamori, S. Proline accumulation by mutation or disruption of the proline oxidase gene improves resistance to freezing and desiccation stresses in saccharomyces cerevisiae. FEMS Microbiol. Lett. 2000, 184, 103–108. [Google Scholar] [CrossRef]
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223. [Google Scholar] [CrossRef]
- Huang, T.; Jander, G. Abscisic acid-regulated protein degradation causes osmotic stress-induced accumulation of branched-chain amino acids in Arabidopsis thaliana. Planta 2017, 246, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Cui, X. Metabolic profiling of dunaliella salina shifting cultivation conditions to nitrogen deprivation. J. Postgenomics Drug Biomark. Dev. 2016, 6. [Google Scholar] [CrossRef]
- Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, G. The Protein Digestibility–Corrected Amino Acid Score. J. Nutr. 2000, 130, 1865S–1867S. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.I.B.; Chagas, B.M.E.; Sassi, R.; Medeiros, G.F.; Aguiar, E.M.; Borba, L.H.F.; Silva, E.P.E.; Neto, J.C.A.; Rangel, A.H.N. Mixotrophic cultivation of Spirulina platensis in dairy wastewater: Effects on the production of biomass, biochemical composition and antioxidant capacity. PLoS ONE 2019, 14, e0224294. [Google Scholar] [CrossRef]
- Wild, K.J.; Steingaß, H.; Rodehutscord, M. Variability in nutrient composition and in vitro crude protein digestibility of 16 microalgae products. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1306–1319. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.M.R.; Abd El Baky, H.H.; El Baroty, G.S.; El Baz, F.K.; Murad, S.A. Antioxidant activity of protein hydrolysates derived from blue-green algae spirulina platensis extracted with three different methods treated with enzymes. Biosci. Res. 2017, 14, 485–497. [Google Scholar]
- Leal, M.J.R.; Daschner, Á.; Oliag, P.T.; Marí, J.A.T. Informe del comité científico de la agencia española de seguridad alimentaria y nutrición (aesan) en relación a una solicitud de evaluación inicial para la comercialización de la microalga marina tetraselmis chuiien el marco del reglamento (ce) nº 258/97 sobre nuevos alimentos y nuevos ingredientes alimentarios. Revista del Comité Científico de la AESAN 2013, 18, 11–27. [Google Scholar]
- Ummalyma, S.B.; Mathew, A.K.; Pandey, A.; Sukumaran, R.K. Harvesting of microalgal biomass: Efficient method for flocculation through pH modulation. Bioresour. Technol. 2016, 213, 216–221. [Google Scholar] [CrossRef]
- Molino, A.; Rimauro, J.; Casella, P.; Cerbone, A.; Larocca, V.; Chianese, S.; Karatza, D.; Mehariya, S.; Ferraro, A.; Hristoforou, E.; et al. Extraction of astaxanthin from microalga Haematococcus pluvialis in red phase by using generally recognized as safe solvents and accelerated extraction. J. Biotechnol. 2018, 283, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Muhaemin, M. Biomass nutrient profiles of marine microalgae dunaliella salina. J. Penelit. Sains 2010, 13, 64–67. [Google Scholar]
- Meyer, K.A.; Kushi, L.H.; Jacobs, D.R., Jr.; Slavin, J.; Sellers, T.A.; Folsom, A.R. Carbohydrates, dietary fiber, and incident type 2 diabetes in older women. Am. J. Clin. Nutr. 2000, 71, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Praznik, W.; Loeppert, R.; Viernstein, H.; Haslberger, A.G.; Unger, F.M. Dietary Fiber and Prebiotics; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2015; pp. 891–925. [Google Scholar]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Dawczynski, C.; Dittrich, M.; Neumann, T.; Goetze, K.; Welzel, A.; Oelzner, P.; Völker, S.; Schaible, A.; Troisi, F.; Thomas, L.; et al. Docosahexaenoic acid in the treatment of rheumatoid arthritis: A double-blind, placebo-controlled, randomized cross-over study with microalgae vs. sunflower oil. Clin. Nutr. 2018, 37, 494–504. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.; Guihéneuf, F.; Schmid, M.; Stengel, D.; Fitzgerald, G.; Ross, P.; Stanton, C. Algae-derived polyunsaturated fatty acids: Implications for human health. In Polyunsaturated Fatty Acids: Sources, Antioxidant Properties and Health Benefits; Nova Sciences Publishers, Inc.: Hauppauge, NY, USA, 2013. [Google Scholar]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef]
- Chen, H.; Qin, J.; Wei, P.; Zhang, J.; Li, Q.; Fu, L.; Li, S.; Ma, C.; Cong, B. Effects of leukotriene B4 and prostaglandin E2 on the differentiation of murine Foxp3+ T regulatory cells and Th17 cells. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 195–200. [Google Scholar] [CrossRef]
- Sheibanie, A.F.; Yen, J.-H.; Khayrullina, T.; Emig, F.; Zhang, M.; Tuma, R.F.; Ganea, D. The Proinflammatory Effect of Prostaglandin E2 in Experimental Inflammatory Bowel Disease Is Mediated through the IL-23→IL-17 Axis. J. Immunol. 2007, 178, 8138–8147. [Google Scholar] [CrossRef]
- Sheibanie, A.F.; Khayrullina, T.; Safadi, F.F.; Ganea, D. Prostaglandin E2 exacerbates collagen-induced arthritis in mice through the inflammatory interleukin-23/interleukin-17 axis. Arthritis Rheum. 2007, 56, 2608–2619. [Google Scholar] [CrossRef]
- Crooks, S.; Stockley, R. Leukotriene B4. Int. J. Biochem. Cell Biol. 1998, 30, 173–178. [Google Scholar] [CrossRef]
- Ünver, N.; Çelik, Ş.; Yakar, Y. Storage stability and fatty acid composition of Sanliurfa butterfat. Mljekarstvo 2021, 71, 124–131. [Google Scholar] [CrossRef]
- Khaled, A.; Hessein, A.; Abdel-Halim, A.M.; Morsy, F.M. Distribution of heavy metals in seaweeds collected along Marsa-Matrouh beaches, Egyptian Mediterranean Sea. Egypt. J. Aquat. Res. 2014, 40, 363–371. [Google Scholar] [CrossRef]
- Ratnaike, R.N. Acute and chronic arsenic toxicity. Postgrad. Med. J. 2003, 79, 391–396. [Google Scholar] [CrossRef]
- Prüser, T.F.; Braun, P.; Wieacek, C. Mikroalgen als neuartiges lebensmittel. Ernährungs Umschau 2021, 4, M206–M213. [Google Scholar]
- Chen, J.; Wang, Y.; Benemann, J.R.; Zhang, X.; Hu, H.; Qin, S. Microalgal industry in China: Challenges and prospects. Environ. Biol. Fishes 2016, 28, 715–725. [Google Scholar] [CrossRef]
- Liu, L.; Wang, F.; Yang, J.; Li, X.; Cui, J.; Liu, J.; Shi, M.; Wang, K.; Chen, L.; Zhang, W. Nitrogen Feeding Strategies and Metabolomic Analysis To Alleviate High-Nitrogen Inhibition on Docosahexaenoic Acid Production in Crypthecodinium cohnii. J. Agric. Food Chem. 2018, 66, 10640–10650. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, R.; Muñoz, R.; Taboada, M.E.; Vega, M.; Bolado, S. Comparative uptake study of arsenic, boron, copper, manganese and zinc from water by different green microalgae. Bioresour. Technol. 2018, 263, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.J.; Lee, Y.K. Determination of biomass dry weight of marine microalgae. Environ. Biol. Fishes 1997, 9, 189–194. [Google Scholar] [CrossRef]
- Trehan, N.; Afonso, L.; Levine, D.L.; Levy, P.D. Vitamin D Deficiency, Supplementation, and Cardiovascular Health. Crit. Pathw. Cardiol. A J. Evid. Based Med. 2017, 16, 109–118. [Google Scholar] [CrossRef]
- Rao, D.S.; Raghuramulu, N. Food chain as origin of vitamin D in fish. Comp. Biochem. Physiol. Part A Physiol. 1996, 114, 15–19. [Google Scholar] [CrossRef]
- Takeuchi, A.; Okano, T.; Tanda, M.; Kobayashi, T. Possible origin of extremely high contents of vitamin-d3 in some kinds of fish liver. Comp. Biochem. Physiol. A Physiol. 1991, 100, 483–487. [Google Scholar]
- Safafar, H.; van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef]
- Watanabe, F.; Takenaka, S.; Kittaka-Katsura, H.; Ebara, S.; Miyamoto, E. Characterization and Bioavailability of Vitamin B12-Compounds from Edible Algae. J. Nutr. Sci. Vitaminol. 2002, 48, 325–331. [Google Scholar] [CrossRef]
- Beale, S.I. Enzymes of chlorophyll biosynthesis. Photosynth. Res. 1999, 60, 43–73. [Google Scholar] [CrossRef]
- Kang, Y.-R.; Park, J.; Jung, S.K.; Chang, Y.H. Synthesis, characterization, and functional properties of chlorophylls, pheophytins, and Zn-pheophytins. Food Chem. 2018, 245, 943–950. [Google Scholar] [CrossRef]
- Morsy, E.; Abou-El-Souod, G.; Hassan, L. Comparison of different media formulations and the optimal growing conditions on growth, morphology and chlorophyll content of green alga, chlorella vulgaris. J. Am. Sci. 2016, 12, 86–95. [Google Scholar]
- Domínguez-Bocanegra, A.; Legarreta, I.G.; Jeronimo, F.M.; Campocosio, A.T. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2004, 92, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Richer, S.; Stiles, W.; Statkute, L.; Pulido, J.; Frankowski, J.; Rudy, D.; Pei, K.; Tsipursky, M.; Nyland, J. Double-masked, placebo-controlled, randomized trial of lutein and antioxidant supplementation in the intervention of atrophic age-related macular degeneration: The Veterans LAST study (Lutein Antioxidant Supplementation Trial). Optom. J. Am. Optom. Assoc. 2004, 75, 216–229. [Google Scholar] [CrossRef]
- Arab, L.; Steck, S. Lycopene and cardiovascular disease. Am. J. Clin. Nutr. 2000, 71, 1691S–1695S. [Google Scholar] [CrossRef]
- Giovannucci, E. Tomatoes, Tomato-Based Products, Lycopene, and Cancer: Review of the Epidemiologic Literature. J. Natl. Cancer Inst. 1999, 91, 317–331. [Google Scholar] [CrossRef]
- Giovannucci, E.; Rimm, E.B.; Liu, Y.; Stampfer, M.J.; Willett, W.C. A prospective study of tomato products, lycopene, and prostate cancer risk. J. Natl. Cancer Inst. 2002, 94, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Kulczyński, B.; Gramza-Michałowska, A.; Kobus-Cisowska, J.; Kmiecik, D. The role of carotenoids in the prevention and treatment of cardiovascular disease—Current state of knowledge. J. Funct. Foods 2017, 38, 45–65. [Google Scholar] [CrossRef]
- Kim, L.; Rao, A.V.; Rao, L.G. Lycopene II—Effect on Osteoblasts: The Carotenoid Lycopene Stimulates Cell Proliferation and Alkaline Phosphatase Activity of SaOS-2 Cells. J. Med. Food 2003, 6, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Shi, J.; Pan, Z.; Cheng, Y.; Zhang, Y.; Li, N. Effect of heat treatment, pH, sugar concentration, and metal ion addition on green color retention in homogenized puree of Thompson seedless grape. LWT 2014, 55, 595–603. [Google Scholar] [CrossRef]
- Boon, C.S.; McClements, D.J.; Weiss, J.; Decker, E. Factors Influencing the Chemical Stability of Carotenoids in Foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Koca, N.; Karadeniz, F.; Burdurlu, H.S. Effect of pH on chlorophyll degradation and colour loss in blanched green peas. Food Chem. 2007, 100, 609–615. [Google Scholar] [CrossRef]
- Oelshlegel, F.; Schroeder, J.; Stahmann, M. A simple procedure for basic hydrolysis of proteins and rapid determination of tryptophan using a starch column. Anal. Biochem. 1970, 34, 331–337. [Google Scholar] [CrossRef]
- Fountoulakis, M.; Lahm, H.-W. Hydrolysis and amino acid composition analysis of proteins. J. Chromatogr. A 1998, 826, 109–134. [Google Scholar] [CrossRef]
- (NREL). Nitrogen-to-Protein Factor Calculator. Available online: https://www.nrel.gov/bioenergy/assets/docs/calculation_sheet_n_factor.xls (accessed on 2 June 2020).
- Lee, S.C.; Prosky, L.; De Vries, J.W. Determination of Total, Soluble, and Insoluble Dietary Fiber in Foods—Enzymatic-Gravimetric Method, MES-TRIS Buffer: Collaborative Study. J. AOAC Int. 1992, 75, 395–416. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Degen, C.; Ecker, J.; Piegholdt, S.; Liebisch, G.; Schmitz, G.; Jahreis, G. Metabolic and growth inhibitory effects of conjugated fatty acids in the cell line HT-29 with special regard to the conversion of t11,t13-CLA. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2011, 1811, 1070–1080. [Google Scholar] [CrossRef]
- Kühn, J.; Hirche, F.; Geissler, S.; Stangl, G.I. Oral intake of 7-dehydrocholesterol increases vitamin D3 concentrations in the liver and kidney. J. Steroid Biochem. Mol. Biol. 2016, 164, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Kühn, J.; Schröter, A.; Hartmann, B.M.; Stangl, G.I. Cocoa and chocolate are sources of vitamin D2. Food Chem. 2018, 269, 318–320. [Google Scholar] [CrossRef] [PubMed]
- Klocker, H.; Hååg, P.; Eder, I.E.; Bartsch, G. Targeting the androgen receptor in hormone-refractory prostate cancer? new concepts. Futur. Oncol. 2005, 1, 93–101. [Google Scholar] [CrossRef]
- Baur, A.C.; Brandsch, C.; König, B.; Hirche, F.; Stangl, G.I. Plant Oils as Potential Sources of Vitamin D. Front. Nutr. 2016, 3, 29. [Google Scholar] [CrossRef]
- Rowan, K.S. Photosynthetic Pigments of Algae/Kingsley s. Rowan; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1989. [Google Scholar]
- Jeffrey, S.T.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Danesi, E.; Rangel-Yagui, C.; Carvalho, J.; Sato, S. Effect of reducing the light intensity on the growth and production of chlorophyll by Spirulina platensis. Biomass Bioenergy 2004, 26, 329–335. [Google Scholar] [CrossRef]
- De Nobel, W.T.; Matthijs, H.C.P.; Von Elert, E.; Mur, L.R. Comparison of the light-limited growth of the nitrogen-fixing cyanobacteria Anabaena and Aphanizomenon. New Phytol. 1998, 138, 579–587. [Google Scholar] [CrossRef]


{kind=link}
| AP | CP | CV | DS | TC | AFA | HP | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ◊ | 1 | 2 | 3 | ◊ | 1 | 2 | ◊ | 1 | 2 | ◊ | 1 | 1 | 1 | |
| Ala | 3.70 ± 0.34 | 3.65 ± 0.22 | 3.87 ± 0.10 | 3.39 ± 0.13 | 3.77 ± 0.23 | a | 3.88 ± 0.10 | 3.440 ± 0.001 | 3.82 ± 0.11 | a | 3.81 ± 0.24 | 3.59 ± 0.19 | a | 0.009 ± 0.009 | <LOQ | b | 0.72 ± 0.10 | 3.66 ± 0.27 | 0.300 ± 0.001 |
| Arg | 3.24 ± 0.35 | 3.29 ± 0.22 | 3.45 ± 0.08 | 3.21 ± 0.12 | 3.27 ± 0.21 | a | 2.99 ± 0.06 | 2.58 ± 0.01 | 2.87 ± 0.09 | b | 2.95 ± 0.19 | 2.69 ± 0.15 | b | <LOQ | <LOQ | b | 0.52 ± 0.07 | 2.52 ± 0.19 | 0.320 ± 0.003 |
| Asp | 4.67 ± 0.44 | 4.70 ± 0.29 | 4.90 ± 0.13 | 4.38 ± 0.19 | 4.70 ± 0.33 | a | 4.16 ± 0.09 | 3.76 ± 0.03 | 4.23 ± 0.15 | b | 4.06 ± 0.25 | 3.94 ± 0.21 | b | 0.010 ± 0.009 | <LOQ | c | 0.89 ± 0.13 | 4.41 ± 0.30 | 0.450 ± 0.003 |
| Cys | 0.06 ± 0.01 | 0.037 ± 0.002 | 0.071 ± 0.004 | 0.037 ± 0.003 | 0.030 ± 0.003 | a | 0.0486 ± 0.0004 | 0.023 ± 0.001 | 0.037 ± 0.004 | a | 0.046 ± 0.002 | 0.035 ± 0.003 | a | <LOQ | <LOQ | b | 0.013 ± 0.002 | 0.016 ± 0.002 | <LOQ |
| Glu | 5.48 ± 0.42 | 5.40 ± 0.32 | 5.52 ± 0.16 | 5.08 ± 0.25 | 5.44 ± 0.33 | a | 4.96 ± 0.17 | 4.46 ± 0.01 | 4.80 ± 0.11 | b | 4.78 ± 0.29 | 4.49 ± 0.23 | b | 0.027 ± 0.008 | 0.014 ± 0.001 | c | 1.22 ± 0.17 | 5.21 ± 0.36 | 0.84 ± 0.02 |
| Gly | 2.44 ± 0.23 | 2.41 ± 0.14 | 2.55 ± 0.07 | 2.22 ± 0.07 | 2.49 ± 0.12 | a | 2.66 ± 0.06 | 2.25 ± 0.01 | 2.63 ± 0.07 | a | 2.55 ± 0.17 | 2.48 ± 0.14 | a | 0.010 ± 0.003 | 0.004 ± 0.004 | b | 0.58 ± 0.08 | 2.23 ± 0.18 | 0.260 ± 0.004 |
| His | 0.81 ± 0.08 | 0.80 ± 0.05 | 0.81 ± 0.02 | 0.66 ± 0.03 | 0.79 ± 0.04 | a | 0.98 ± 0.01 | 0.84 ± 0.01 | 0.91 ± 0.03 | a | 0.94 ± 0.06 | 0.85 ± 0.05 | a | 0.011 ± 0.019 | <LOQ | b | 0.15 ± 0.02 | 0.76 ± 0.07 | 0.091 ± 0.004 |
| Ile | 2.34 ± 0.24 | 2.32 ± 0.16 | 2.43 ± 0.03 | 2.09 ± 0.04 | 2.36 ± 0.13 | a | 1.59 ± 0.03 | 1.336 ± 0.003 | 1.49 ± 0.02 | b | 1.46 ± 0.10 | 1.39 ± 0.08 | b | 0.004 ± 0.007 | <LOQ | c | 0.40 ± 0.06 | 2.13 ± 0.18 | 0.21 ± 0.03 |
| Leu | 4.00 ± 0.39 | 3.98 ± 0.25 | 4.16 ± 0.10 | 3.63 ± 0.14 | 4.04 ± 0.26 | a | 3.82 ± 0.08 | 3.38 ± 0.02 | 3.74 ± 0.11 | a | 3.70 ± 0.25 | 3.51 ± 0.19 | a | 0.027 ± 0.013 | 0.020 ± 0.019 | b | 0.84 ± 0.12 | 3.72 ± 0.27 | 0.44 ± 0.02 |
| Lys | 2.23 ± 0.24 | 2.26 ± 0.14 | 2.35 ± 0.07 | 2.07 ± 0.09 | 2.30 ± 0.16 | a | 3.81 ± 0.08 | 2.17 ± 0.01 | 3.40 ± 0.11 | a | 3.72 ± 0.23 | 3.08 ± 0.17 | a | <LOQ | <LOQ | b | 0.47 ± 0.05 | 2.42 ± 0.18 | 0.12 ± 0.01 |
| Met | 1.09 ± 0.21 | 0.82 ± 0.08 | 0.80 ± 0.04 | 0.65 ± 0.13 | 0.89 ± 0.14 | a | 0.73 ± 0.06 | 0.41 ± 0.07 | 0.86 ± 0.01 | a | 0.77 ± 0.11 | 0.81 ± 0.03 | a | <LOQ | <LOQ | b | 0.04 ± 0.07 | 0.43 ± 0.01 | 0.01 ± 0.02 |
| Phe | 2.16 ± 0.22 | 2.16 ± 0.14 | 2.22 ± 0.05 | 1.90 ± 0.07 | 2.16 ± 0.14 | a | 2.24 ± 0.04 | 1.97 ± 0.01 | 2.20 ± 0.06 | a | 2.17 ± 0.14 | 2.07 ± 0.12 | a | 0.006 ± 0.010 | <LOQ | b | 0.57 ± 0.08 | 1.96 ± 0.18 | 0.270 ± 0.004 |
| Pro | 2.00 ± 0.17 | 2.06 ± 0.11 | 2.10 ± 0.07 | 1.81 ± 0.05 | 2.02 ± 0.12 | a | 2.57 ± 0.18 | 1.960 ± 0.003 | 2.41 ± 0.07 | a | 2.52 ± 0.06 | 2.30 ± 0.07 | a | <LOQ | <LOQ | b | 0.59 ± 0.11 | 1.71 ± 0.08 | 0.220 ± 0.005 |
| Ser | 2.75 ± 0.28 | 2.76 ± 0.17 | 2.92 ± 0.07 | 2.49 ± 0.10 | 2.80 ± 0.19 | a | 2.08 ± 0.05 | 1.89 ± 0.01 | 2.15 ± 0.07 | b | 2.07 ± 0.13 | 2.01 ± 0.11 | b | <LOQ | <LOQ | b | 0.49 ± 0.07 | 2.55 ± 0.16 | 0.280 ± 0.006 |
| Thr | 2.38 ± 0.25 | 2.40 ± 0.16 | 2.50 ± 0.06 | 2.12 ± 0.09 | 2.43 ± 0.17 | a | 2.08 ± 0.05 | 1.73 ± 0.01 | 2.17 ± 0.07 | a | 2.01 ± 0.13 | 2.05 ± 0.12 | a | <LOQ | <LOQ | b | 0.49 ± 0.07 | 2.58 ± 0.18 | 0.190 ± 0.002 |
| Trp | 0.51 ± 0.03 | 0.37 ± 0.04 | 0.42 ± 0.16 | 0.55 ± 0.26 | 0.41 ± 0.04 | a | 0.37 ± 0.03 | 0.31 ± 0.07 | 0.42 ± 0.05 | a | 0.33 ± 0.04 | 0.38 ± 0.05 | a | 0.028 ± 0.008 | <LOQ | b | 0.08 ± 0.02 | 0.39 ± 0.04 | 0.06 ± 0.02 |
| Tyr | 2.40 ± 0.27 | 2.38 ± 0.16 | 2.50 ± 0.05 | 2.18 ± 0.09 | 2.41 ± 0.16 | a | 2.52 ± 0.07 | 3.54 ± 0.01 | 2.54 ± 0.09 | a | 2.53 ± 0.16 | 2.27 ± 0.13 | a | 0.011 ± 0.019 | <LOQ | b | 0.37 ± 0.05 | 2.43 ± 0.18 | 0.46 ± 0.02 |
| Val | 2.43 ± 0.22 | 2.41 ± 0.16 | 2.55 ± 0.05 | 2.25 ± 0.05 | 2.48 ± 0.14 | a | 2.39 ± 0.05 | 2.10 ± 0.01 | 2.30 ± 0.05 | a | 2.23 ± 0.15 | 2.16 ± 0.13 | a | 0.024 ± 0.004 | 0.019 ± 0.003 | b | 0.55 ± 0.08 | 2.28 ± 0.19 | 0.31 ± 0.02 |
| Tau | 0.045 ± 0.004 | 0.034 ± 0.004 | 0.033 ± 0.003 | 0.078 ± 0.002 | 0.0295 ± 0.0005 | a | 0.05 ± 0.01 | 0.043 ± 0.001 | 0.045 ± 0.001 | a | 0.051 ± 0.003 | 0.035 ± 0.003 | a | 0.306 ± 0.037 | 0.313 ± 0.017 | b | 0.18 ± 0.03 | 0.035 ± 0.005 | 0.301 ± 0.008 |
| MetS | 0.16 ± 0.09 | 0.31 ± 0.05 | 0.36 ± 0.04 | 0.34 ± 0.15 | 0.27 ± 0.14 | a | 0.23 ± 0.05 | 0.45 ± 0.05 | 0.13 ± 0.03 | a | 0.22 ± 0.06 | 0.14 ± 0.01 | a | <LOQ | <LOQ | b | 0.10 ± 0.06 | 0.33 ± 0.05 | 0.094 ± 0.003 |
| (Cys)2 | 0.43 ± 0.04 | 0.42 ± 0.02 | 0.42 ± 0.01 | 0.39 ± 0.02 | 0.427 ± 0.004 | a | 0.40 ± 0.02 | 0.40 ± 0.01 | 0.451 ± 0.001 | a | 0.44 ± 0.03 | 0.43 ± 0.02 | a | 0.047 ± 0.004 | 0.053 ± 0.003 | b | 0.10 ± 0.02 | 0.29 ± 0.03 | 0.110 ± 0.001 |
| GABA | 0.059 ± 0.002 | 0.062 ± 0.001 | 0.059 ± 0.002 | 0.05 ± 0.01 | 0.054 ± 0.003 | a | 0.08 ± 0.01 | 0.083 ± 0.003 | 0.071 ± 0.001 | a | 0.09 ± 0.01 | 0.063 ± 0.003 | a | <LOQ | 0.003 ± 0.005 | b | 0.014 ± 0.002 | 0.02 ± 0.03 | 0.021 ± 0.001 |
| Orn | 0.027 ± 0.005 | 0.021 ± 0.005 | 0.022 ± 0.002 | 0.021 ± 0.002 | 0.023 ± 0.001 | a | 0.025 ± 0.001 | 0.040 ± 0.002 | 0.036 ± 0.01 | a | 0.026 ± 0.001 | 0.026 ± 0.001 | a | <LOQ | <LOQ | a | <LOQ | 0.50 ± 0.04 | 0.028 ± 0.001 |
| NH4+ | 0.76 ± 0.10 | 0.71 ± 0.05 | 0.75 ± 0.01 | 0.66 ± 0.02 | 0.76 ± 0.03 | a | 0.92 ± 0.02 | 0.95 ± 0.01 | 0.80 ± 0.01 | b | 0.80 ± 0.05 | 0.79 ± 0.05 | b | 0.034 ± 0.011 | 0.033 ± 0.002 | c | 0.14 ± 0.01 | 0.86 ± 0.07 | 0.174 ± 0.008 |
| Evaluation of protein quality | |||||||||||||||||||
| NEA | 23.50 ± 0.85 | 23.40 ± 0.57 | 24.44 ± 0.26 | 21.58 ± 0.37 | 23.67 ± 0.69 | a | 22.87 ± 0.30 | 21.33 ± 0.04 | 22.61 ± 0.26 | a | 22.36 ± 0.53 | 21.13 ± 0.43 | a | 0.089 ± 0.024 | 0.024 ± 0.004 | b | 4.87 ± 0.28 | 22.21 ± 0.62 | 2.80 ± 0.03 |
| SEA | 4.05 ± 0.35 | 4.09 ± 0.22 | 4.26 ± 0.08 | 3.87 ± 0.12 | 4.06 ± 0.22 | a | 3.97 ± 0.06 | 3.42 ± 0.02 | 3.78 ± 0.10 | a | 3.89 ± 0.20 | 3.54 ± 0.16 | a | 0.012 ± 0.019 | 0.002 ± 0.001 | b | 0.67 ± 0.08 | 3.28 ± 0.20 | 0.41 ± 0.01 |
| EAA | 17.14 ± 0.69 | 16.72 ± 0.43 | 17.43 ± 0.22 | 15.25 ± 0.36 | 17.08 ± 0.44 | a | 17.03 ± 0.16 | 13.40 ± 0.10 | 16.59 ± 0.19 | a | 16.39 ± 0.45 | 15.45 ± 0.34 | a | 0.103 ± 0.020 | 0.045 ± 0.019 | b | 3.45 ± 0.21 | 15.92 ± 0.49 | 1.61 ± 0.05 |
| AAS (%) | 100 ± 1.5 | 94 ± 0.74 | 94 ± 0.22 | 71 ± 0.37 | 94 ± 0.76 | a | 82 ± 0.3 | 59 ± 0.2 | 100 ± 0.3 | a | 88 ± 0.9 | 94 ± 0.6 | a | 6 ± 3.1 | 13 ± 0.02 | b | 29 ± 1.4 | 59 ± 1.1 | 18 ± 0.8 |
| N-factor | 4.87 | 4.72 | 4.76 | 4.75 | 4.62 | a | 4.63 | 4.67 | 4.73 | a | 4.67 | 4.54 | a | 0.71 | 0.42 | b | 4.90 | 4.63 | 4.85 |
| AP | CP | CV | DS | TC | AFA | HP | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ◊ | 1 | 2 | 3 | ◊ | 1 | 2 | ◊ | 1 | 2 | ◊ | 1 | 1 | 1 | |
| Protein | |||||||||||||||||||
| Nitrogen (Kjeldahl) | 9.25 ± 0.05 | 10.33 ± 0.07 | 10.0 ± 0.2 | 10.4 ± 0.1 | 8.8 ± 0.4 | a | 9.12 ± 0.07 | 8.5 ± 0.1 | 9.83 ± 0.08 | a | 9.6 ± 0.2 | 9.6 ± 0.1 | a | 1.5 ± 0.2 | 1.0 ± 0.8 | b | 1.94 ± 0.06 | 9.3 ± 0.2 | 1.04 ± 0.04 |
| Crude protein (N-factor 6.25) | 57.8 ± 0.3 | 64.5 ± 0.5 | 62.7 ± 1.2 | 65.2 ± 0.7 | 55.1 ± 2.2 | a | 57.0 ± 0.4 | 53.0 ± 0.6 | 61.4 ± 0.5 | a | 60.0 ± 1.2 | 60.1 ± 0.4 | a | 0.9 ± 0.1 | 0.6 ± 0.5 | b | 12.1 ± 0.4 | 58.4 ± 1.5 | 6.5 ± 0.3 |
| Crude protein (N-factor 4.78) | 44.2 ± 0.2 | 49.3 ± 0.4 | 48.0 ± 1.0 | 49.9 ± 0.6 | 42.2 ± 1.7 | a | 43.6 ± 0.3 | 40.6 ± 0.5 | 47.0 ± 0.4 | a | 45.9 ± 1.0 | 46.0 ± 0.6 | a | 0.7 ± 0.1 | 0.5 ± 0.4 | b | 9.3 ± 0.3 | 44.6 ± 1.1 | 5.0 ± 0.2 |
| Crude protein (specific N-factor) | 45.1 ± 0.3 | 48.7 ± 0.4 | 47.8 ± 1.2 | 49.6 ± 0.7 | 40.8 ± 2.0 | a | 42.4 ± 0.4 | 39.6 ± 0.6 | 46.5 ± 0.5 | a | 44.8 ± 1.1 | 43.7 ± 0.7 | a | 0.21 ± 0.01 | 0.24 ± 0.03 | b | 9.5 ± 0.4 | 43.2 ± 1.4 | 5.1 ± 0.3 |
| Pure protein (specific N-factor) | 38.3 ± 1.6 | 39.7 ± 0.2 | 39.9 ± 0.4 | 43.4 ± 1.7 | 37.8 ± 0.7 | a | 40.7 ± 0.5 | 34.0 ± 0.1 | 41.9 ± 0.3 | a | 41.0 ± 0.5 | 39.2 ± 0.3 | a | 0.12 ± 0.2 | 0.10 ± 0.1 | b | 9.1 ± 0.1 | 33.0 ± 1.4 | 3.3 ± 0.5 |
| NPN | 1.4 ± 0.3 | 1.90 ± 0.09 | 4.7 ± 0.2 | 5.6 ± 0.3 | 4.2 ± 0.1 | a | 0.3 ± 0.1 | 1.2 ± 0.1 | 1.0 ± 0.1 | a | 0.8 ± 0.2 | 1.0 ± 0.1 | a | 0.09 ± 0.20 | 0.14 ± 0.10 | a | 0.08 ± 0.06 | 2.2 ± 0.4 | 0.4 ± 0.1 |
| Further macronutrients | |||||||||||||||||||
| Total fiber | 30.9 ± 3.5 | 13.0 ± 1.1 | 10.9 ± 1.4 | 13.2 ± 1.5 | 38.7 ± 1.5 | a | 36.6 ± 2.6 | 21.9 ± 0.2 | 18.6 ± 1.7 | a | 29.8 ± 1.4 | 21.9 ± 1.1 | a | 6.3 ± 0.1 | 6.8 ± 0.1 | a | 9.2 ± 0.3 | 19.1 ± 2.2 | 15.8 ± 0.5 |
| Total fat | 4.32 ± 0.03 | 4.8 ± 0.2 | 4.7 ± 0.2 | 5.6 ± 0.3 | 4.2 ± 0.1 | a | 8.1 ± 0.5 | 7.7 ± 0.3 | 8.6 ± 0.2 | b | 8.4 ± 0.2 | 8.7 ± 0.2 | b | 3.0 ± 0.1 | 3.05 ± 0.05 | c | 8.0 ± 0.1 | 4.4 ± 0.1 | 2.1 ± 0.1 |
| Carbohydrates | 15.4 ± 3.9 | 34.9 ± 1.1 | 36.9 ± 1.4 | 31.2 ± 2.3 | 13.6 ± 1.6 | a | 8.8 ± 2.7 | 28.6 ± 0.4 | 25.1 ± 1.8 | a | 15.9 ± 1.5 | 24.9 ± 1.2 | a | 86.6 ± 0.2 | 85.8 ± 0.2 | b | 55.9 ± 0.4 | 36.6 ± 2.6 | 77.9 ± 0.8 |
| Further main components | |||||||||||||||||||
| Ash | 11.0 ± 0.2 | 7.6 ± 0.1 | 7.62 ± 0.03 | 6.59 ± 0.03 | 5.77 ± 0.01 | a | 5.83 ± 0.02 | 7.8 ± 0.04 | 5.70 ± 0.02 | a | 4.90 ± 0.03 | 5.329 ± 0.001 | a | 3.985 ± 0.003 | 4.24 ± 0.05 | a | 17.74 ± 0.04 | 6.9 ± 0.1 | 0.91 ± 0.03 |
| Fatty Acids | AP | CP | CV | DS | TC | AFA | HP | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ◊ | 1 | 2 | 3 | ◊ | 1 | 2 | ◊ | 1 | 2 | ◊ | 1 | 1 | 1 | |
| SFAs | |||||||||||||||||||
| C12:0 | 4.11 | 6.69 | 0.46 | 3.12 | 4.46 | a | 1.59 | 1.08 | 1.56 | a | 1.42 | 2.36 | a | 8.73 | 7.65 | b | 2.49 | 4.40 | 0.49 |
| C14:0 | 65.6 | 15.7 | 6.97 | 14.5 | 24.2 | a | 25.3 | 21.3 | 25.6 | a | 21.0 | 23.4 | a | 15.8 | 15.6 | a | 22.9 | 275 | 5.95 |
| C16:0 | 1278 | 1895 | 2076 | 1790 | 1809 | a | 857 | 1467 | 1195 | ab | 813 | 942 | b | 776 | 802 | b | 1486 | 1662 | 177 |
| C18:0 | 435 | 97.0 | 44.8 | 249 | 388 | ab | 288 | 53.1 | 855 | ab | 273 | 160 | a | 453 | 486 | b | 40.6 | 75.5 | 37.6 |
| C20:0 | 6.55 | 2.87 | 2.10 | 4.69 | 7.59 | a | 6.18 | 2.50 | 25.0 | a | 5.46 | 7.36 | a | 7.71 | 9.43 | a | 2.93 | 1.51 | 1.67 |
| C22:0 | 2.09 | 0.65 | <LOQ | 0.73 | 2.44 | a | 1.50 | 3.56 | 13.9 | ab | 1.53 | 5.57 | a | 8.73 | 12.9 | b | <LOQ | <LOQ | 0.65 |
| C24:0 | 9.57 | <LOQ | <LOQ | 1.08 | 4.42 | ab | 6.61 | 14.0 | 35.7 | a | 5.62 | 6.59 | a | 2.54 | 4.96 | b | 2.03 | <LOQ | 1.28 |
| MUFAs | |||||||||||||||||||
| C14:1n5 | 0.96 | 0.18 | 0.41 | 0.53 | 0.59 | a | 0.59 | 1.09 | <LOQ | a | <LOQ | 0.80 | a | <LOQ | <LOQ | a | <LOQ | 17.4 | 0.28 |
| C16:1n7 | 146 | 146 | 152 | 110 | 129 | a | 45.4 | 33.3 | 31.5 | b | 40.3 | 115 | ab | 1.86 | 2.68 | c | 42.5 | 122 | 29.5 |
| C17:1n7 | 3.90 | 1.20 | 7.99 | 1.96 | 4.71 | a | 14.9 | 35.3 | 9.85 | b | 18.2 | 7.01 | ab | 0.51 | 0.55 | c | 2.97 | 7.56 | 3.36 |
| C18:1n9 | 134 | 190 | 115 | 136 | 175 | a | 137.7 | 547 | 3430 | ab | 158 | 179 | a | 777 | 797 | b | 1844 | 136 | 37.8 |
| C18:1n7 | 31.5 | 20.8 | 16.0 | 23.6 | <LOQ | ac | 47.5 | 90.8 | 26.9 | ab | 50.5 | 92.1 | b | 13.9 | 14.3 | c | 173 | 22.7 | 24.7 |
| C20:1n9 | 8.07 | <LOQ | <LOQ | 0.86 | 4.13 | a | 3.06 | 14.06 | 8.86 | a | 3.88 | 2.79 | a | 3.64 | 3.94 | a | 165 | 0.76 | 0.63 |
| C22:1n9 | 5.22 | 1.49 | <LOQ | <LOQ | 1.81 | a | <LOQ | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | <LOQ | <LOQ |
| n6—PUFAs | |||||||||||||||||||
| C18:2n6 | 635 | 1392 | 942 | 896 | 1489 | a | 2592 | 1971 | 2184 | b | 3143 | 2196 | b | 895 | 739 | a | 482 | 217 | 512 |
| C18:3n6 | 699 | 628 | 781 | 489 | 915 | a | 1.85 | 18.3 | <LOQ | b | <LOQ | 2.53 | b | 0.67 | 0.30 | b | 44.6 | <LOQ | 8.33 |
| C20:2n6 | 12.8 | 6.59 | 4.13 | 5.17 | 15.2 | a | 9.17 | 3.12 | 5.85 | a | 9.12 | 5.70 | a | 0.17 | <LOQ | b | 4.20 | 0.78 | 1.22 |
| C20:3n6 | 59.5 | 12.0 | 5.99 | 6.45 | 20.4 | a | <LOQ | <LOQ | <LOQ | b | <LOQ | <LOQ | b | <LOQ | <LOQ | b | 3.50 | <LOQ | <LOQ |
| C20:4n6 | 37.5 | 6.08 | 1.98 | 3.00 | 19.2 | a | <LOQ | <LOQ | <LOQ | b | <LOQ | 1.19 | b | <LOQ | <LOQ | b | 44.4 | 2.45 | 0.29 |
| C22:5n6 | 2.46 | 0.43 | <LOQ | 0.74 | 6.51 | a | <LOQ | <LOQ | 1.61 | a | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | <LOQ | <LOQ |
| n-3 PUFAs | |||||||||||||||||||
| C18:3n3 | 2.23 | 1.54 | 4.61 | 3.28 | 4.02 | a | 1247 | 868 | 35.5 | b | 624 | 1299 | b | 1.28 | 1.40 | a | 972 | 880 | 326 |
| C20:5n3 | 2.52 | 0.60 | 4.31 | 1.40 | 9.30 | a | 7.14 | 8.85 | 3.55 | a | 5.95 | 4.61 | a | 4.39 | 6.94 | a | 406 | 6.74 | 1.12 |
| C22:6n3 | 0.54 | <LOQ | 0.47 | <LOQ | 2.54 | a | <LOQ | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | <LOQ | <LOQ |
| Sum | |||||||||||||||||||
| SFAs | 1801 | 2018 | 2130 | 2063 | 2240 | a | 1186 | 1563 | 2152 | a | 1121 | 1147 | a | 1273 | 1339 | a | 1557 | 2018 | 225 |
| MUFAs | 330 | 360 | 292 | 273 | 315 | a | 249 | 721 | 3507 | ab | 271 | 397 | a | 797 | 819 | b | 2228 | 306 | 96.2 |
| PUFAs | 1451 | 2047 | 1744 | 1405 | 2482 | a | 3857 | 2869 | 2230 | ab | 3782 | 3510 | b | 901 | 748 | c | 1956 | 1107 | 849 |
| Others | 739 | 305 | 624 | 409 | 544 | a | 3327 | 2557 | 191 | ab | 3516 | 3326 | b | 79.1 | 94.5 | c | 2300 | 960 | 880 |
| n6 | 1445 | 2045 | 1735 | 1400 | 2466 | a | 2603 | 1992 | 2191 | a | 3152 | 2206 | a | 895 | 740 | b | 578 | 220 | 522 |
| n3 | 5.29 | 2.14 | 9.39 | 4.69 | 15.9 | a | 1254 | 877 | 39.1 | b | 630 | 1304 | b | 5.67 | 8.34 | a | 1378 | 887 | 327 |
| n6/n3 | 273 | 956 | 185 | 299 | 155 | a | 2.08 | 2.27 | 56.1 | b | 5.01 | 1.69 | b | 158 | 88.7 | a | 0.42 | 0.25 | 1.59 |
| AP | CP | CV | DS | TC | AFA | HP | LOQ | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ◊ | 1 | 2 | 3 | ◊ | 1 | 2 | ◊ | 1 | 1 | 1 | 1 | |||
| Minerals | |||||||||||||||||||
| Ca | mg/100 g | 4.54 ± 0.41 | 12.5 ± 0.6 | 4227 ± 128 | 809.1 ± 31.2 | 149.1 ± 1.1 | a | 538.6 ± 25.4 | 101.6 ± 1.3 | 148.2 ± 4.6 | a | 87.1 ± 1.3 | 112.2 ± 0.7 | a | 75.7 ± 2.0 | 184.8 ± 6.2 | 183.5 ± 8.8 | 157.2 ± 5.0 | 0.11 |
| Mg | mg/100 g | 4.05 ± 0.05 | 22.4 ± 1.2 | 267.8 ± 10.7 | 228.5 ± 10.6 | 2304 ± 10 | a | 815.1 ± 35.0 | 235.6 ± 3.46 | 282.4 ± 2.5 | b | 286.8 ± 1.4 | 276.3 ± 2.6 | b | 267.2 ± 1.57 | 304.6 ± 15.4 | 294.8 ± 13.6 | 368.1 ± 13.2 | 0.48 |
| Trace elements | |||||||||||||||||||
| Fe | mg/100 g | 1.16 ± 0.01 | 4.96 ± 0.08 | 6.97 ± 0.10 | 49.2 ± 1.4 | 83.5 ± 1.5 | a | 20.7 ± 0.8 | 48.1 ± 0.9 | 21.98 ± 0.04 | a | 54.0 ± 0.7 | 53.1 ± 1.2 | a | 52.7 ± 1.1 | 51.8 ± 1.9 | 110.4 ± 2.5 | 97.2 ± 3.7 | 0.07 |
| Mn | µg/100 g | <LOQ | <LOQ | 3.89 ± 0.11 | <LOQ | 5.14 ± 0.15 | a | 9.13 ± 0.52 | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | 5.28 ± 0.25 | 5.67 ± 0.28 | 5.25 ± 0.19 | 3.23 |
| Ni | µg/100 g | 8.04 ± 1.92 | 16.6 ± 0.5 | 47.5 ± 1.5 | 72.9 ± 12.1 | 41.43 ± 0.07 | a | 61.2 ± 1.7 | 71.3 ± 3.2 | 75.4 ± 0.6 | b | 90.2 ± 1.6 | 71.5 ± 1.7 | b | 111.0 ± 7.8 | 13.23 ± 0.78 | 20.3 ± 2.6 | 32.5 ± 1.9 | 2.45 |
| Cu | µg/100 g | 11.0 ± 0.3 | 188.4 ± 4.6 | 575.4 ± 0.4 | 489.4 ± 3.8 | 300.0 ± 8.3 | a | 623.0 ± 13.4 | 56.6 ± 0.2 | 79.9 ± 1.3 | a | 65.3 ± 14.1 | 76.7 ± 5.1 | a | 182.7 ± 0.2 | 521.4 ± 3.8 | 499.4 ± 2.6 | 274.7 ± 18.3 | 5.83 |
| Zn | µg/100 g | <LOQ | 11943 ± 166 | 2404 ± 10 | 351.5 ± 82.6 | 2144 ± 75 | a | 1211 ± 120 | 607.6 ± 4.3 | 1790 ± 14 | a | 1032 ± 38 | 758.0 ± 63 | a | 2106 ± 21 | 2073 ± 99 | 1938 ± 3 | 1610 ± 54 | 39.85 |
| Mo | µg/100 g | 3.06 ± 0.02 | 36.9 ± 0.9 | 66.3 ± 2.0 | 558.8 ± 1.2 | 26.2 ± 0.9 | a | 34.8 ± 0.1 | 14.4 ± 0.1 | 11.08 ± 0.07 | a | 13.07 ± 0.28 | 11.8 ± 0.7 | a | 16.30 ± 0.02 | 50.9 ± 0.1 | 47.09 ± 0.5 | 27.7 ± 1.3 | 0.14 |
| Se | µg/100 g | <LOQ | 2.34 ± 0.09 | 0.30 ± 0.14 | 6.40 ± 0.04 | 7.74 ± 0.22 | a | <LOQ | 7.02 ± 0.65 | 13.2 ± 2.6 | a | 8.90 ± 0.47 | 8.17 ± 0.15 | a | 9.52 ± 0.44 | <LOQ | <LOQ | 6.15 ± 1.31 | 0.20 |
| I | µg/100 g | 273.9 ± 204.7 | 252.2 ± 319.7 | 431.4 ± 75.3 | 195.4 ± 214.9 | 372.8 ± 58.4 | a | 238.5 ± 140.9 | 308.1 ± 32.9 | 280.1 ± 210.8 | a | 211.9 ± 144.3 | 207.7 ± 107.1 | a | 213.9 ± 149.8 | 245.9 ± 288.4 | 173.3 ± 230.7 | 555.2 ± 129.0 | 0.05 |
| Heavy metals | |||||||||||||||||||
| As | µg/100 g | 0.89 ± 0.14 | 10.8 ± 0.2 | 49.4 ± 0.7 | 799.2 ± 14.8 | 47.0 ± 1.6 | a | 5.1 ± 0.1 | 70.0 ± 0.7 | 19.4 ± 0.2 | a | 40.5 ± 2.3 | 27.9 ± 0.3 | a | 213.7 ± 3.7 | 55.1 ± 0.5 | 60.37 ± 0.05 | 45.2 ± 1.3 | 0.03 |
| Cd | µg/100 g | 0.54 ± 0.26 | 9.61 ± 1.56 | 2.27 ± 1.91 | 1.96 ± 0.01 | 1.72 ± 0.50 | a | 0.46 ± 0.07 | 1.18 ± 0.26 | 1.64 ± 0.20 | a | 0.97 ± 0.02 | 1.66 ± 0.30 | a | 2.69 ± 0.84 | 0.59 ± 0.16 | 0.58 ± 0.30 | 1.48 ± 0.01 | 0.20 |
| Hg | µg/100 g | 0.58 ± 0.07 | 0.76 ± 0.16 | 0.25 ± 0.07 | 4.91 ± 1.02 | 3.96 ± 0.11 | a | 0.377 ± 0.001 | 0.59 ± 0.37 | 0.428 ± 0.001 | a | 0.55 ± 0.04 | 0.68 ± 0.07 | a | 1.51 ± 0.43 | 0.40 ± 0.14 | 0.3835 ± 0.0001 | 3.33 ± 0.64 | 0.2 |
| Pb | µg/100 g | 10.9 ± 7.72 | 6.8 ± 8.71 | 22.7 ± 16.06 | 4.25 ± 5.31 | 16.48 ± 10.3 | a | 4.16 ± 5.17 | 32.0 ± 10.7 | 31.1 ± 11.4 | a | 8.92 ± 9.20 | 30.2 ± 11.7 | a | 36.7 ± 18.7 | 5.90 ± 7.64 | 4.98 ± 6.33 | 11.2 ± 9.5 | 0.5 |
| AP | CP | CV | DS | TC | AFA | HP | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ◊ | 1 | 2 | 3 | ◊ | 1 | 2 | ◊ | 1 | 2 | ◊ | 1 | 1 | 1 | ||
| Vitamin D | ||||||||||||||||||||
| Vitamin D2 * | µg/100 g | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | a | 37.3 ± 2.9 | 68.7 ± 1.7 | 420.6 ± 1.7 | b | 55.4 ± 3.5 | 110.3 ± 7.7 | b | <LOQ | <LOQ | a | <LOQ | <LOQ | <LOQ |
| Vitamin D3 o | µg/100 g | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | a | <LOQ | <LOQ | 1.25 ± 0.03 | a | <LOQ | 0.639 ± 0.007 | a | <LOQ | <LOQ | a | <LOQ | 0.96 ± 0.07 | <LOQ |
| Tocopherols | ||||||||||||||||||||
| α-tocopherol # | mg/100 g | 1.20 ± 0.14 | <LOQ | <LOQ | <LOQ | 0.85 ± 0.40 | a | 3.62 ± 0.34 | 3.94 ± 0.55 | 4.78 ± 0.37 | b | 3.85 ± 0.23 | 4.50 ± 0.51 | b | 46.90 ± 1.96 | 37.47 ± 1.41 | c | 9.49 ± 0.07 | 1.98 ± 0.44 | <LOQ |
| δ-tocopherol # | mg/100 g | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | a | <LOQ | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | <LOQ | a | <LOQ | 1.90 ± 0.50 | <LOQ |
| AP | CP | CV | DS | TC | AFA | HP | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 1 | 2 | 1 | 2 | ||||
| Energy (kJ) | 1463 | 1306 | 1626 | 1553 | 1782 | / | 1221 | / | 1693 | 1448 | 893 | / | 1421 | / | / |
| Protein | 60 | 56.6 | 62.8 | 60.40 | 60 | / | 60.70 | / | 55 | 61.3 | 7.4 | / | 38 | / | / |
| Fat total | 8.2 | 4.10 | 6.4 | 4.10 | 8.2 | / | 4.70 | / | 10 | 7.8 | 7 | / | 7.5 | / | / |
| PUFAs | / | 1.4 | / | 1.10 | / | / | 2.20 | / | / | 2.2 | / | / | 4.1 | / | / |
| MUFAs | / | 1.5 | / | 0.36 | / | / | 1.10 | / | / | 2.5 | / | / | 1.2 | / | / |
| SFAs | 1.1 | 1.3 | / | 1.41 | 1.1 | / | 1.40 | / | 0 | 3.1 | 3 | / | 2.2 | / | / |
| Carbohydrates | 15.1 | 11.3 | 18.9 | 20.42 | 27 | / | 60.70 | / | 23 | 6.9 | 29.7 | / | 31 | / | / |
| Sugar | / | 3.7 | / | 3.88 | 27 | / | / | / | 0 | 1.6 | 5 | / | 0 | / | / |
| Fiber | 7 | 16.4 | / | 3.50 | 13 | / | 13.00 | / | 0 | 16.1 | <1 | / | / | / | / |
| Salt | / | <0.1 | / | 2.12 | 3.7 | / | / | / | 0 | <0.1 | / | / | 4.2 | / | / |
| Vitamins | B1: 4.4 mg B2: 6.9 mg B3: 5.9 mg B6: 18.4 mg B12: 0.16 mg C: 1.4 mg D: 24 IU K: 1.3 mg | / | A: 68 mg C: 17 mg | A: 0.6 mg B6: 0.3 mg B12: 0.05 mg C: 5.1 mg K: 3.1 mg | B12: 350 µg | / | / | B12: 100 µg | Total vitamins: 60 µg | / | / | / | / | / | / |
| Minerals and trace elements | Ca: 270 mg Mg: 270 mg Fe: 82.7 mg K: 1040 mg Zn: 3.3 mg Na: 700 mg | / | Fe: 87 mg | Mg: 920 mg Mn: 4.6 mg Fe: 87 mg P: 102 mg Zn: 10.1 mg K: 1104 mg Cl: 112 mg F: 0.9 mg Cu: 1.84 mg I: 0.46 mg | Fe: 48 mg Ca: 130 mg | / | / | / | Fe: 102 mg | / | / | / | / | / | / |
| Additional information | Folic acid: 27 µg Chlorophyll: 1179 mg Carotenoids: 451 mg | / | / | / | / | / | Chlorophyll: 2.88 g | / | / | / | / | / | Ash: 15 g | / | / |
| Origin | / | China | China | China | / | / | / | Germany | / | China | China | China | Spain | / | / |
| Expiration date | 10/2020 | 04/2021 | 07/2019 | 04/2020 | 05/2019 | 05/2020 | 04/2020 | 04/2019 | 07/2020 | 11/2020 | 06/2020 | 12/2019 | 04/2020 | 03/2020 | 01/2020 |
| Daily reference intake | <5 g | 3 Tea-spoons | 3 g | 5 g | 5 g | 4 g | 5 g | 3 g | 4.5 g | 3 Tea-spoons | 2 g | 1 Tea-spoon | 0.25 g | 2 g | 0.012 g |
| Ranking | Name | Positiv Characteristics | Negative Characteristics |
|---|---|---|---|
| 1. | Chlorella pyrenoidosa No.3 | ↑↑AAS, ↑↑ protein, ↑↑ total fat, ↑↑↑ vitamin D2 + D3 | ↑ n6/n3 ratio |
| 2. | Chlorella vulgaris No.2 | ↑↑↑ AAS, ↑↑ fiber, ↑↑ protein, ↑↑↑ chlorophyll, ↑↑↑ vitamin D2, ↓↓↓ n6/n3 ratio | |
| 3. | Aphanizomenon flos-aquae | ↑↑ protein, ↑↑ Fe + Cu, ↑↑ vitamin D3 + δ-tocopherol, ↓↓↓ n6/n3 ratio | ↑↑↑ C16:0 |
| 4. | Chlorella pyrenoidosa No.1 | ↑↑ AAS, ↑↑↑ fiber, ↑↑ minerals + Cu, ↓↓↓ heavy metals, ↑↑ vitamin D2, ↓↓ n6/n3 ratio | |
| 5. | Chlorella vulgaris No.1 | ↑↑ AAS, ↑↑ protein, ↑↑↑ fiber, ↑↑↑ chlorophyll, ↑↑ vitamin D2, ↓↓ n6/n3 ratio | |
| 6. | Tetraselmis chuii | ↑↑↑ carbohydrates, ↑↑ α-tocopherol, ↑↑ EPA, ↓↓↓ n6/n3 ratio | ↑↑ As, ↓↓ chlorophylls |
| 7. | Chlorella pyrenoidosa No.2 | ↑↑↑ fiber, ↑↑ vitamin D2, ↓↓ n6/n3 ratio | ↓↓ trace elements, ↑↑ As |
| 8. | Arthrospira platensis No.1 | ↑↑↑ AAS, ↑↑ protein, ↑↑ fiber, ↓↓↓ heavy metals | ↓↓ minerals + trace elements, ↓↓↓ vitamins, ↑ n6/n3 ratio |
| 9. | Arthrospira platensis No.5 | ↑↑ AAS, ↑↑ protein, ↑↑↑ fiber, ↑ Mg, Cu, Zn | ↑ As, ↓↓↓ vitamins, ↑↑ n6/n3 ratio |
| 10. | Haematococcus pluvialis | ↑↑↑ carbohydrates, ↑↑↑ carotenoids, ↑↑ minerals, ↓↓ n6/n3 ratio | ↓↓ total fat + protein, ↓↓ AAS, ↓↓↓ vitamins |
| 11. | Arthrospira platensis No.3 | ↑↑↑ protein, ↑↑↑ minerals + Zn | ↑↑↑ C16:0, ↑↑ n6/n3 ratio, ↓↓↓ vitamins |
| 12. | Arthrospira platensis No.2 | ↑↑↑ protein, ↑↑↑ Zn | ↑↑↑ C16:0, ↑↑ n6/n3 ratio, ↓↓↓ vitamins |
| 13. | Arthrospira platensis No.4 | ↑↑↑ protein, ↑↑↑ Mo + Ca | ↑↑↑ C16:0, ↑↑ n6/n3 ratio, ↑↑↑ As, ↓↓↓ vitamins |
| 14. | Dunaliella salina No.2 | ↑↑↑ carbohydrates, ↑↑ Zn, ↑↑ α-tocopherol | ↓↓↓ macronutrients, ↓↓↓ AAS, ↑ n6/n3 ratio |
| 15. | Dunaliella salina No.1 | ↑↑↑ carbohydrates, ↑↑ Zn, ↑↑↑ α-tocopherol | ↓↓↓ macronutrients, ↓↓↓ AAS, ↑ n6/n3 ratio, ↑↑ As+Pb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandgruber, F.; Gielsdorf, A.; Baur, A.C.; Schenz, B.; Müller, S.M.; Schwerdtle, T.; Stangl, G.I.; Griehl, C.; Lorkowski, S.; Dawczynski, C. Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders. Mar. Drugs 2021, 19, 310. https://doi.org/10.3390/md19060310
Sandgruber F, Gielsdorf A, Baur AC, Schenz B, Müller SM, Schwerdtle T, Stangl GI, Griehl C, Lorkowski S, Dawczynski C. Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders. Marine Drugs. 2021; 19(6):310. https://doi.org/10.3390/md19060310
Chicago/Turabian StyleSandgruber, Fabian, Annekathrin Gielsdorf, Anja C. Baur, Benjamin Schenz, Sandra Marie Müller, Tanja Schwerdtle, Gabriele I. Stangl, Carola Griehl, Stefan Lorkowski, and Christine Dawczynski. 2021. "Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders" Marine Drugs 19, no. 6: 310. https://doi.org/10.3390/md19060310
APA StyleSandgruber, F., Gielsdorf, A., Baur, A. C., Schenz, B., Müller, S. M., Schwerdtle, T., Stangl, G. I., Griehl, C., Lorkowski, S., & Dawczynski, C. (2021). Variability in Macro- and Micronutrients of 15 Commercially Available Microalgae Powders. Marine Drugs, 19(6), 310. https://doi.org/10.3390/md19060310