
Oyster-Derived Tyr-Ala (YA) Peptide Prevents Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Suppressing Inflammatory, Apoptotic, Ferroptotic, and Pyroptotic Signals

 , , , , , , , and
, , , , , , , and 
Abstract
:1. Introduction
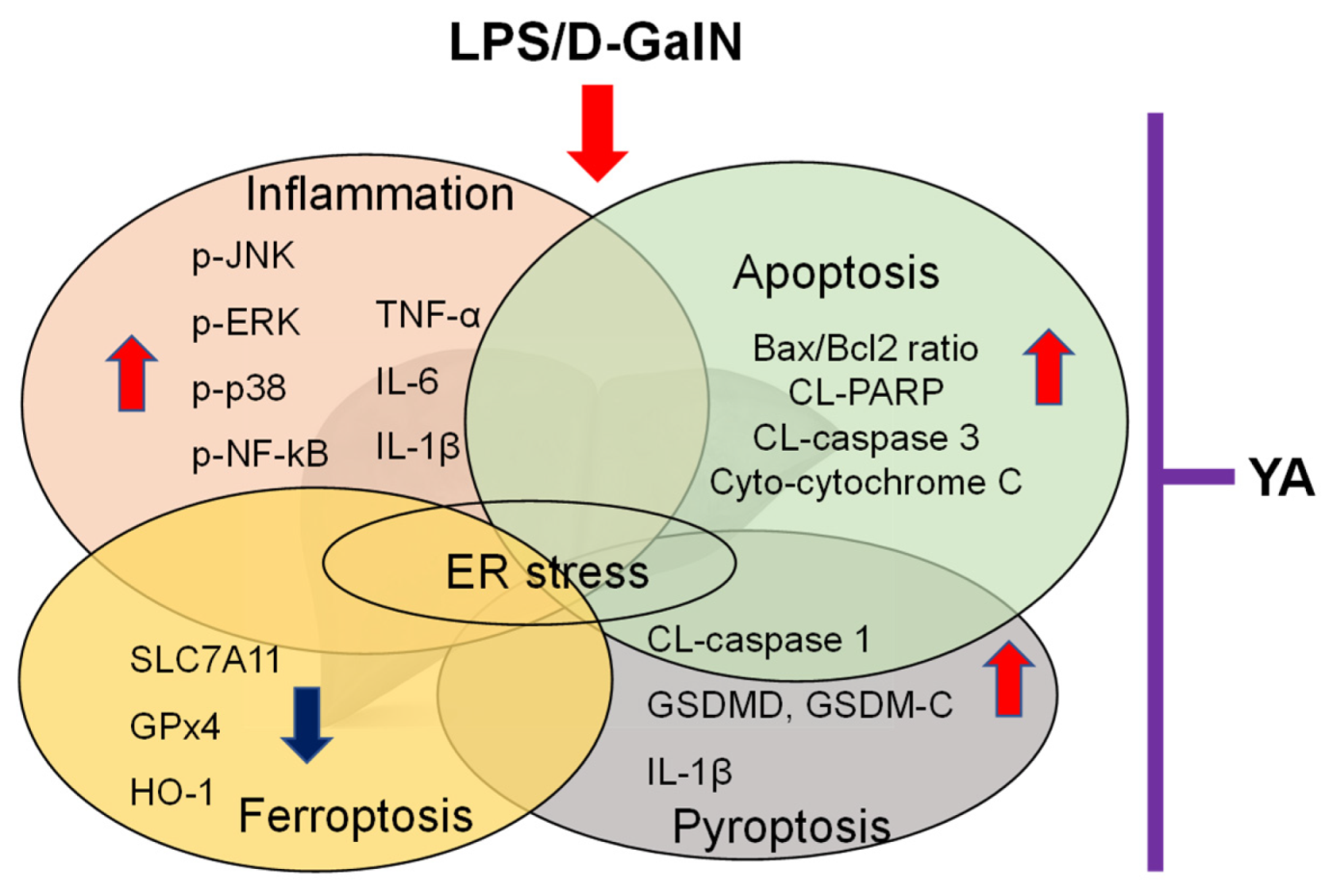
2. Results
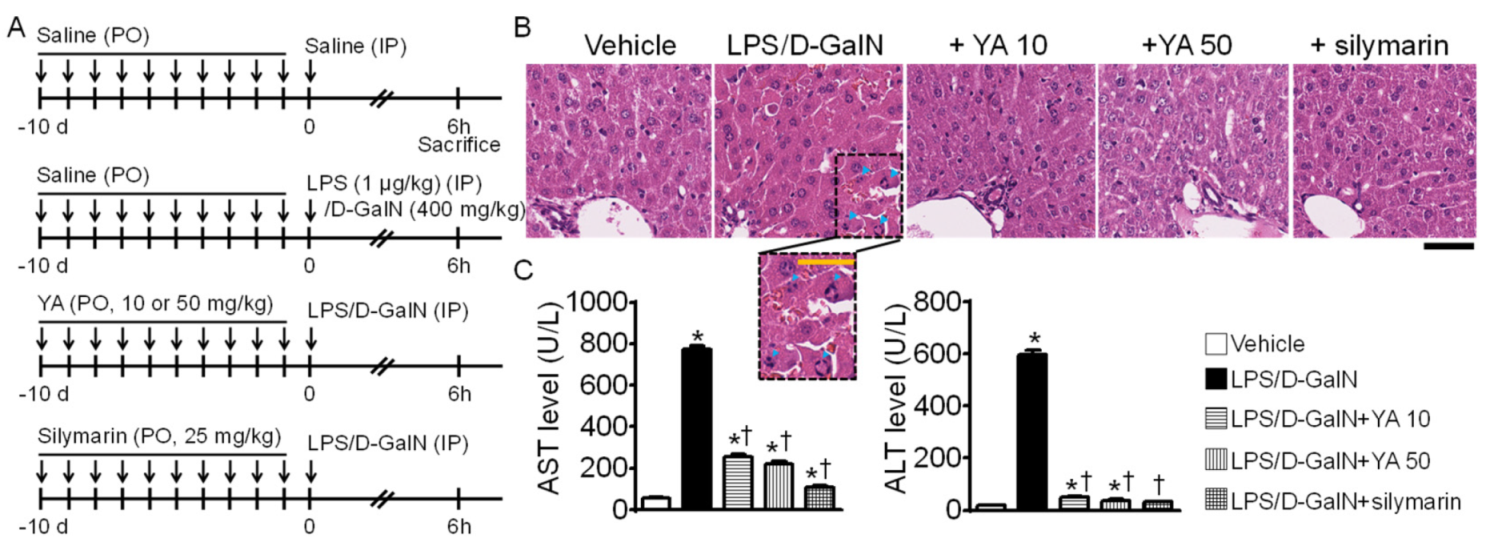
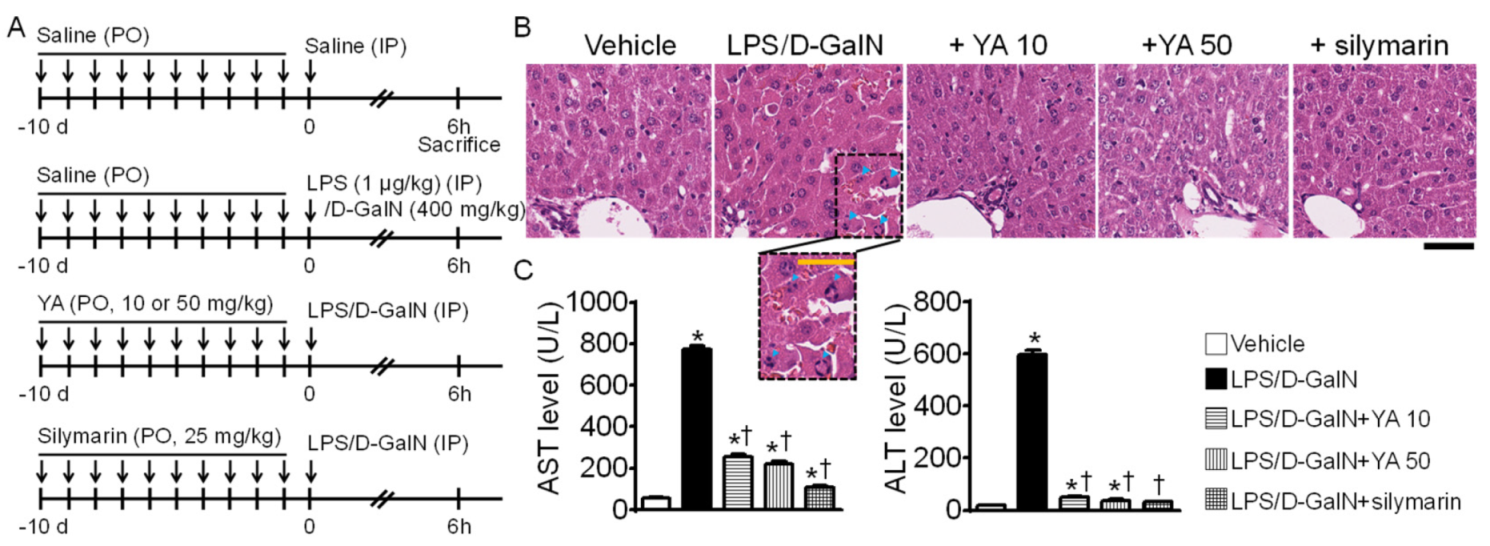
2.1. Generation of Acute Liver Failure (ALF) Mouse Model
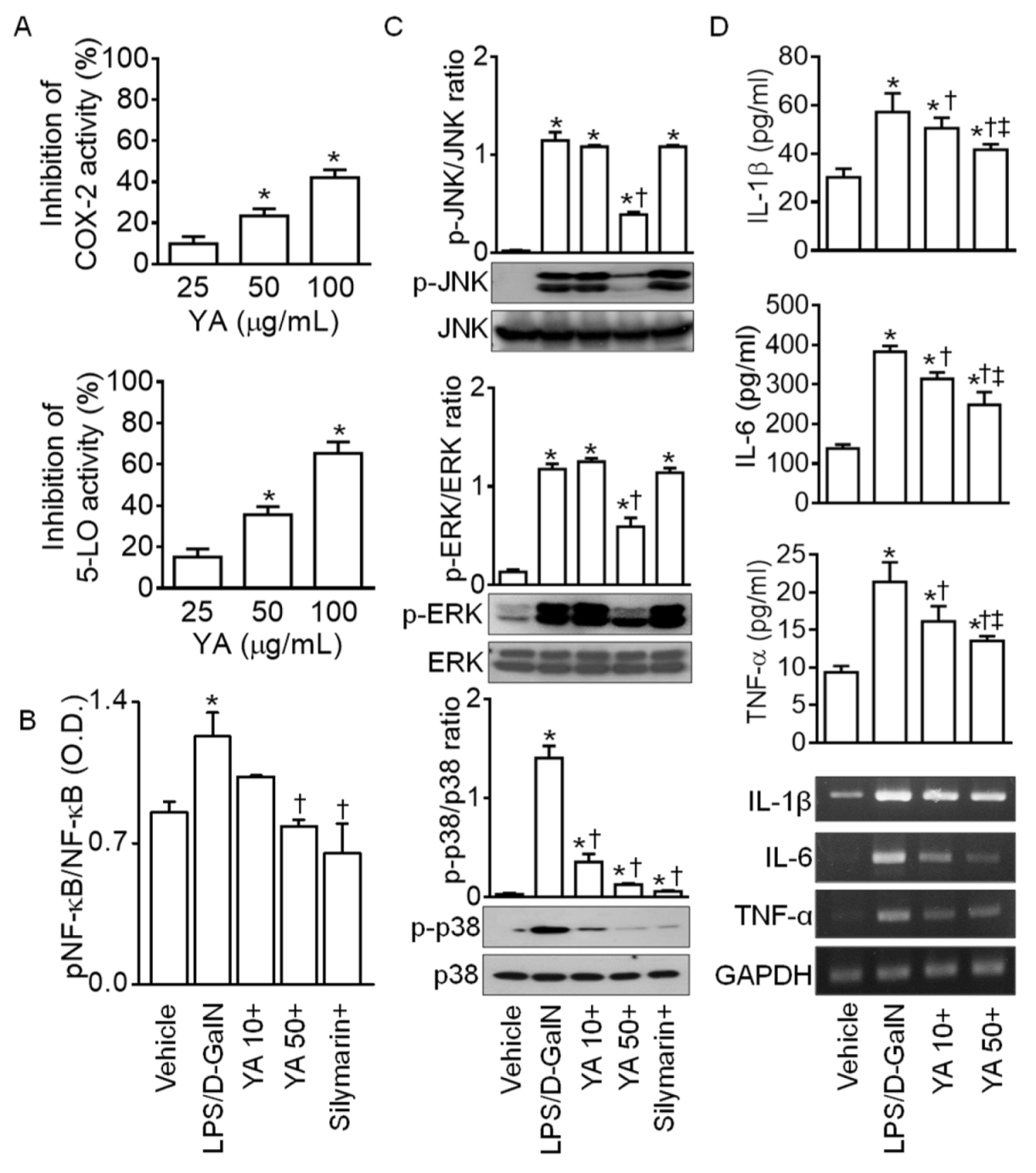
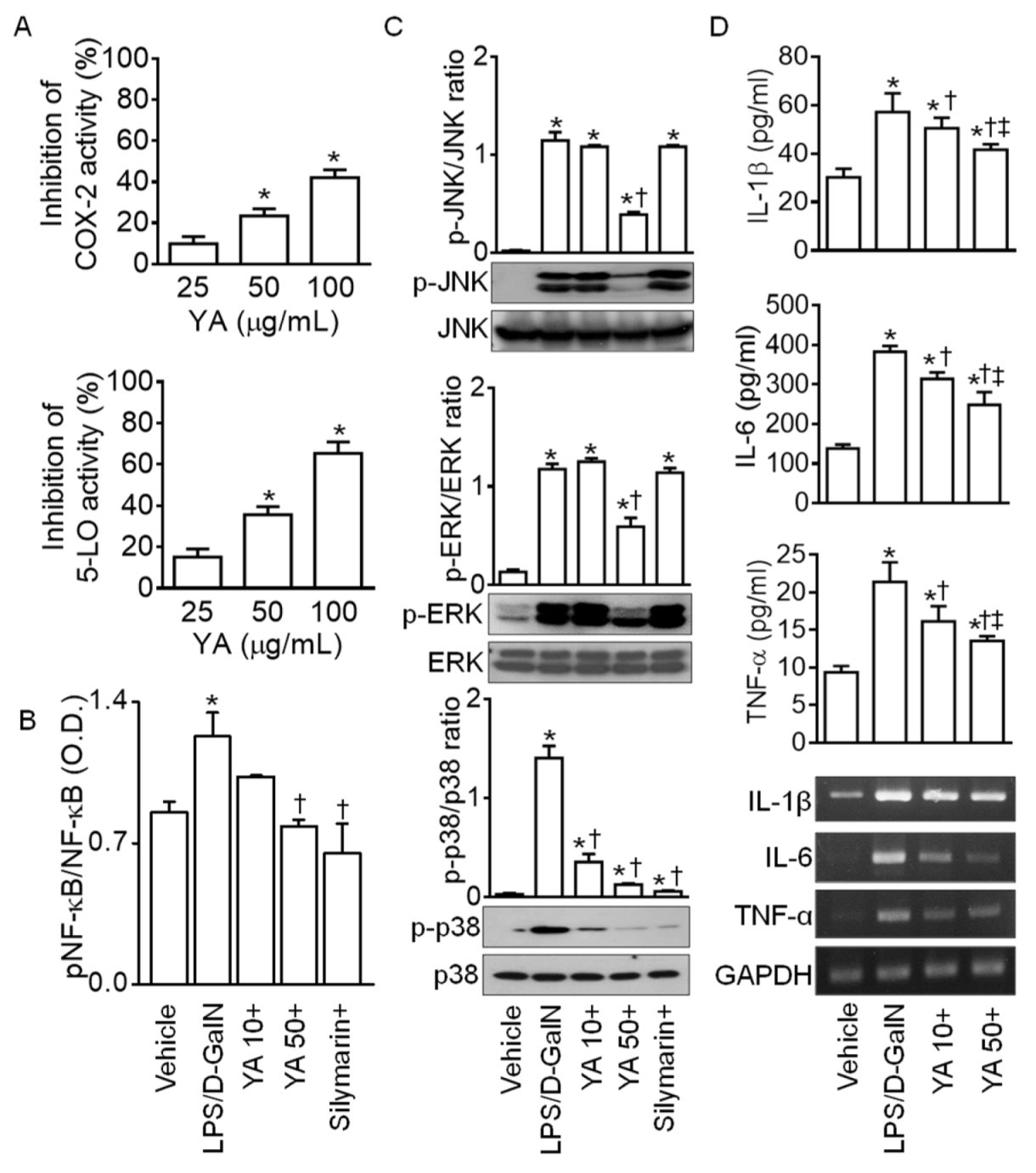
2.2. YA Pre-Administration Attenuated Inflammatory Signals in ALF Model
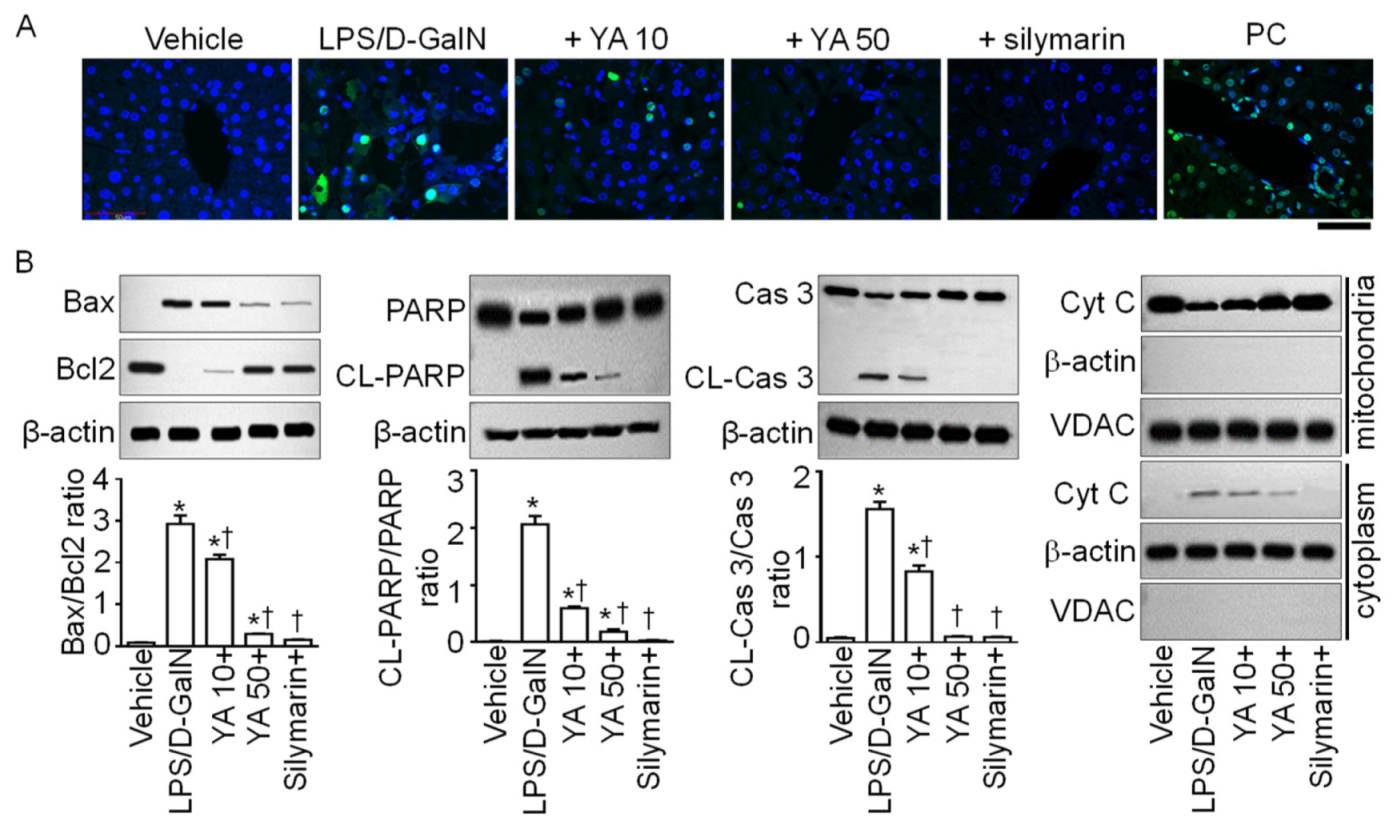
2.3. YA Pre-Administration Attenuated Apoptotic Signals in ALF Model
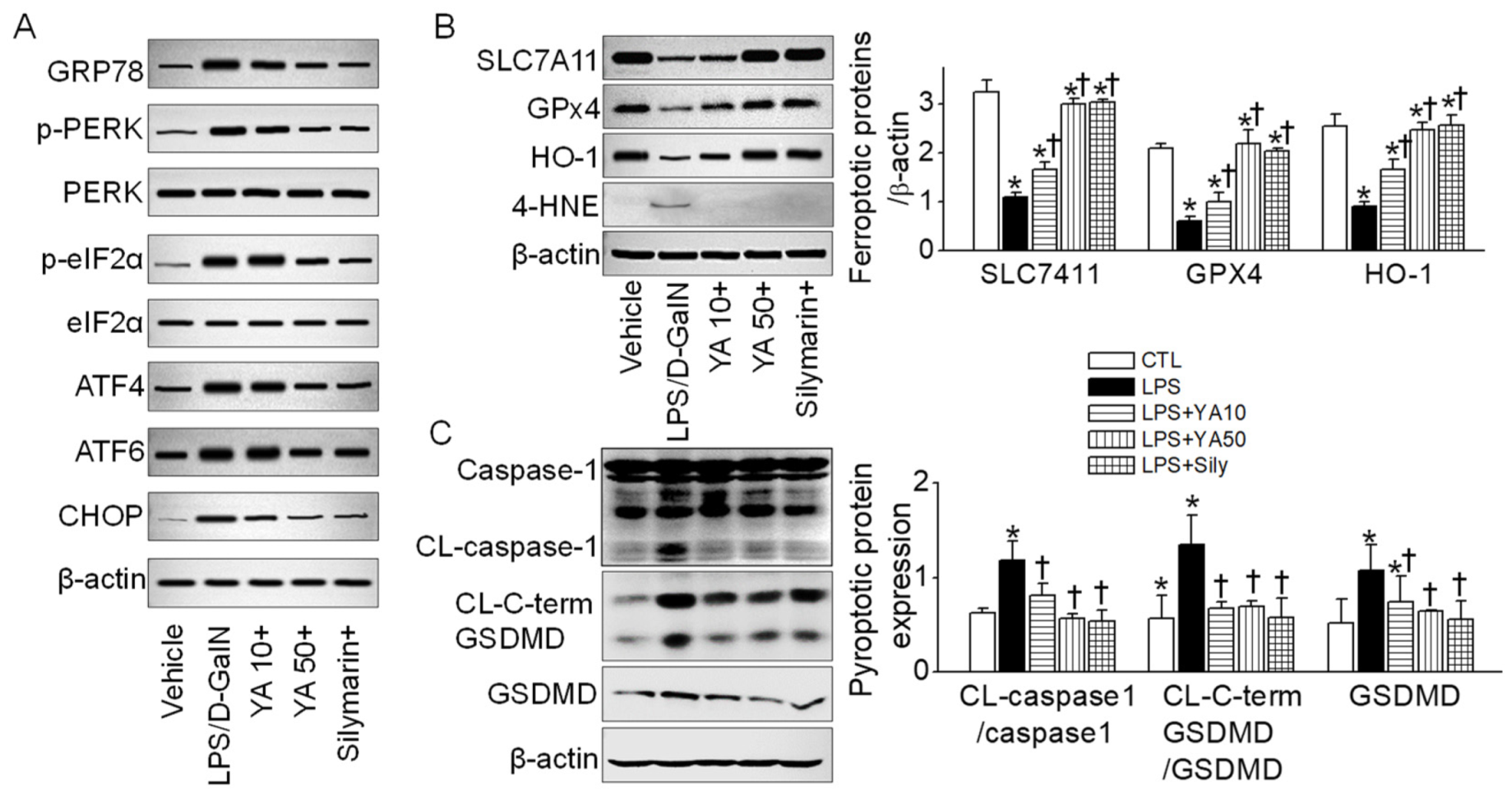
2.4. YA Pre-Administration Attenuated Endoplasmic Reticulum (ER) Stress and Ferroptosis and Pyroptosis Signals in ALF Model
3. Discussion
4. Materials and Methods
4.1. Preparation of YA Peptide
4.2. Measurement of Cyclooxygenase-2 (COX-2) and 5-Lipoxygenase (5-LO) Inhibition Activity
4.3. LPS/D-GalN-Induced ALF Model
4.4. Measurement of Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) Levels
4.5. Hematoxylin and Eosin (H&E) Staining
4.6. TUNEL Staining
4.7. RT-PCR
4.8. Western Blot Analysis
4.9. Measurement of IL-1β, IL-6, and TNF-α Concentrations in Liver Tissues
4.10. Measurement of Total and Phospho-NF-ĸB p65 Protein
4.11. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernal, W.; Wendon, J. Acute liver failure. N. Engl. J. Med. 2013, 369, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Toma, D.; Lazar, O.; Bontas, E. Acute Liver Failure; Springer Nature Switzerland AG: Cham, Switzerland, 2020; p. 12. [Google Scholar]
- Rolando, N.; Wade, J.; Davalos, M.; Wendon, J.; Philpott-Howard, J.; Williams, R. The systemic inflammatory response syndrome in acute liver failure. Hepatology 2000, 32, 734–739. [Google Scholar] [CrossRef]
- Rolando, N.; Harvey, F.; Brahm, J.; Philpott-Howard, J.; Alexander, G.; Gimson, A.; Casewell, M.; Fagan, E.; Williams, R. Prospective study of bacterial infection in acute liver failure: An analysis of fifty patients. Hepatology 1990, 11, 49–53. [Google Scholar] [CrossRef]
- Zhai, X.R.; Tong, J.J.; Wang, H.M.; Xu, X.; Mu, X.Y.; Chen, J.; Liu, Z.F.; Wang, Y.; Su, H.B.; Hu, J.H. Infection deteriorating hepatitis B virus related acute-on-chronic liver failure: A retrospective cohort study. BMC Gastroenterol. 2020, 20, 320. [Google Scholar] [CrossRef]
- Yin, X.; Gong, X.; Zhang, L.; Jiang, R.; Kuang, G.; Wang, B.; Chen, X.; Wan, J. Glycyrrhetinic acid attenuates lipopolysaccharide-induced fulminant hepatic failure in D-galactosamine-sensitized mice by up-regulating expression of interleukin-1 receptor-associated kinase-M. Toxicol. Appl. Pharmacol. 2017, 320, 8–16. [Google Scholar] [CrossRef]
- Seo, M.J.; Hong, J.M.; Kim, S.J.; Lee, S.M. Genipin protects D-galactosamine and lipopolysaccharide-induced hepatic injury through suppression of the necroptosis-mediated inflammasome signaling. Eur. J. Pharmacol. 2017, 812, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Kemelo, M.K.; Wojnarova, L.; Kutinova Canova, N.; Farghali, H. D-galactosamine/lipopolysaccharide-induced hepatotoxicity downregulates sirtuin 1 in rat liver: Role of sirtuin 1 modulation in hepatoprotection. Physiol. Res. 2014, 63, 615–623. [Google Scholar] [CrossRef]
- Nowak, M.; Gaines, G.C.; Rosenberg, J.; Minter, R.; Bahjat, F.R.; Rectenwald, J.; MacKay, S.L.; Edwards, C.K., 3rd; Moldawer, L.L. LPS-induced liver injury in D-galactosamine-sensitized mice requires secreted TNF-alpha and the TNF-p55 receptor. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R1202–R1209. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Martich, G.D.; Danner, R.L.; Ceska, M.; Suffredini, A.F. Detection of interleukin 8 and tumor necrosis factor in normal humans after intravenous endotoxin: The effect of antiinflammatory agents. J. Exp. Med. 1991, 173, 1021–1024. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xu, D.-X.; Lv, J.-W.; Ning, H.; Wei, W. Melatonin attenuates lipopolysaccharide (LPS)-induced apoptotic liver damage in D-galactosamine-sensitized mice. Toxicology 2007, 237, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Newsome, P.N.; Plevris, J.N.; Nelson, L.J.; Hayes, P.C. Animal models of fulminant hepatic failure: A critical evaluation. Liver Transplant. 2000, 6, 21–31. [Google Scholar] [CrossRef]
- Eipel, C.; Kidess, E.; Abshagen, K.; Leminh, K.; Menger, M.D.; Burkhardt, H.; Vollmar, B. Antileukoproteinase protects against hepatic inflammation, but not apoptosis in the response of D-galactosamine-sensitized mice to lipopolysaccharide. Br. J. Pharmacol. 2007, 151, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhong, L.; Zhu, H.; Wang, F. The Protective Effect of Cordycepin on D-Galactosamine/Lipopolysaccharide-Induced Acute Liver Injury. Mediat. Inflamm. 2017, 2017, 3946706. [Google Scholar] [CrossRef]
- Li, L.; Yin, H.; Zhao, Y.; Zhang, X.; Duan, C.; Liu, J.; Huang, C.; Liu, S.; Yang, S.; Li, X. Protective role of puerarin on LPS/D-Gal induced acute liver injury via restoring autophagy. Am. J. Transl. Res. 2018, 10, 957–965. [Google Scholar]
- Li, H.; Zhao, X.K.; Cheng, Y.J.; Zhang, Q.; Wu, J.; Lu, S.; Zhang, W.; Liu, Y.; Zhou, M.Y.; Wang, Y.; et al. Gasdermin D-mediated hepatocyte pyroptosis expands inflammatory responses that aggravate acute liver failure by upregulating monocyte chemotactic protein 1/CC chemokine receptor-2 to recruit macrophages. World J. Gastroenterol. 2019, 25, 6527–6540. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Kong, M.; Duan, Z.; Liu, S.; Zheng, S.; Chen, Y. M2-like macrophages exert hepatoprotection in acute-on-chronic liver failure through inhibiting necroptosis-S100A9-necroinflammation axis. Cell Death Dis. 2021, 12, 93. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Shi, C.; Jiao, F.; Gong, Z. Mechanism of glycyrrhizin on ferroptosis during acute liver failure by inhibiting oxidative stress. Mol. Med. Rep. 2019, 20, 4081–4090. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Jangle, S. Hepatoprotective activity of herbal preparation (HP-4) against alcohol induced hepatotoxicity in mice. Int. J. Appl. Sci. Biotechnol. 2014, 2, 50–58. [Google Scholar] [CrossRef]
- Yang, S.; Kuang, G.; Zhang, L.; Wu, S.; Zhao, Z.; Wang, B.; Yin, X.; Gong, X.; Wan, J. Mangiferin Attenuates LPS/D-GalN-Induced Acute Liver Injury by Promoting HO-1 in Kupffer Cells. Front. Immunol. 2020, 11, 285. [Google Scholar] [CrossRef] [PubMed]
- Gillessen, A.; Schmidt, H.H. Silymarin as Supportive Treatment in Liver Diseases: A Narrative Review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef] [Green Version]
- Doehmer, J.; Tewes, B.; Klein, K.U.; Gritzko, K.; Muschick, H.; Mengs, U. Assessment of drug-drug interaction for silymarin. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2008, 22, 610–617. [Google Scholar] [CrossRef]
- Ryu, J.H.; Kim, E.J.; Xie, C.; Nyiramana, M.M.; Siregar, A.S.; Park, S.H.; Cho, S.B.; Song, D.H.; Kim, N.G.; Choi, Y.J.; et al. Hepatoprotective Effects of Oyster Hydrolysate on Lipopolysaccharide/D-galactosamine-Induced Acute Liver Injury in Mice. J. Korean Soc. Food Sci. Nutr. 2017, 46, 659–670. [Google Scholar]
- Siregar, A.S.; Nyiramana, M.M.; Kim, E.J.; Shin, E.J.; Woo, M.S.; Kim, J.M.; Kim, J.H.; Lee, D.K.; Hahm, J.R.; Kim, H.J.; et al. Dipeptide YA is Responsible for the Positive Effect of Oyster Hydrolysates on Alcohol Metabolism in Single Ethanol Binge Rodent Models. Mar. Drugs 2020, 18, 512. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.H.; Ahmad, K.; Saeed, M.; Alharbi, A.M.; Barreto, G.E.; Ashraf, G.M.; Choi, I. Peptide based therapeutics and their use for the treatment of neurodegenerative and other diseases. Biomed. Pharm. 2018, 103, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.; Han, J.R.; Park, H.J.; Hah, J.S.; Kang, J.L. Synthetic RGDS peptide attenuates lipopolysaccharide-induced pulmonary inflammation by inhibiting integrin signaled MAP kinase pathways. Respir. Res. 2009, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Schon, H.T.; Bartneck, M.; Borkham-Kamphorst, E.; Nattermann, J.; Lammers, T.; Tacke, F.; Weiskirchen, R. Pharmacological Intervention in Hepatic Stellate Cell Activation and Hepatic Fibrosis. Front. Pharmacol. 2016, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-kappaB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Xiao, K.; Liu, C.; Tu, Z.; Xu, Q.; Chen, S.; Zhang, Y.; Wang, X.; Zhang, J.; Hu, C.A.; Liu, Y. Activation of the NF-kappaB and MAPK Signaling Pathways Contributes to the Inflammatory Responses, but Not Cell Injury, in IPEC-1 Cells Challenged with Hydrogen Peroxide. Oxid. Med. Cell. Longev. 2020, 2020, 5803639. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Hu, S.Q.; Liu, J.; Cao, H.C.; Xu, W.; Li, Y.J.; Li, L.J. Nature and mechanisms of hepatocyte apoptosis induced by D-galactosamine/lipopolysaccharide challenge in mice. Int. J. Mol. Med. 2014, 33, 1498–1506. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Song, K.; Huang, X.; Fu, S.; Zeng, Q. GDF-15 prevents LPS and D-galactosamine-induced inflammation and acute liver injury in mice. Int. J. Mol. Med. 2018, 42, 1756–1764. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Yang, B.; Zhang, X.; Wen, T.; Wang, X.; Yin, J.; Piao, Z.; Zheng, S.; Zhang, J.; Chen, Y.; et al. Role of endoplasmic reticulum stress in D-GalN/LPS-induced acute liver failure. Zhonghua Gan Zang Bing Za Zhi Chin. J. Hepatol. Abs 2014, 22, 364–368. [Google Scholar] [CrossRef]
- Wang, H.; Chen, L.; Zhang, X.; Xu, L.; Xie, B.; Shi, H.; Duan, Z.; Zhang, H.; Ren, F. Kaempferol protects mice from D-GalN/LPS-induced acute liver failure by regulating the ER stress-Grp78-CHOP signaling pathway. Biomed. Pharmacother. 2019, 111, 468–475. [Google Scholar] [CrossRef]
- Chen, L.; Ren, F.; Zhang, H.; Wen, T.; Piao, Z.; Zhou, L.; Zheng, S.; Zhang, J.; Chen, Y.; Han, Y.; et al. Inhibition of glycogen synthase kinase 3beta ameliorates D-GalN/LPS-induced liver injury by reducing endoplasmic reticulum stress-triggered apoptosis. PLoS ONE 2012, 7, 45202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Kaufman, R.J. From endoplasmic-reticulum stress to the inflammatory response. Nature 2008, 454, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebeaupin, C.; Proics, E.; de Bieville, C.H.; Rousseau, D.; Bonnafous, S.; Patouraux, S.; Adam, G.; Lavallard, V.J.; Rovere, C.; Le Thuc, O.; et al. ER stress induces NLRP3 inflammasome activation and hepatocyte death. Cell Death Dis. 2015, 6, 1879. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Lee, D.H.; Choudry, H.A.; Bartlett, D.L.; Lee, Y.J. Ferroptosis-Induced Endoplasmic Reticulum Stress: Cross-Talk between Ferroptosis and Apoptosis. Mol. Cancer Res. MCR 2018, 16, 1073–1076. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Zhao, T.; Song, Y.; Lin, L.; Fan, X.; Cui, B.; Feng, H.; Wang, X.; Yu, Q.; Zhang, J.; et al. The emerging role of ferroptosis in non-cancer liver diseases: Hype or increasing hope? Cell Death Dis. 2020, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Si, W.; Zeng, J.; Huang, L.; Huang, Z.; Zhao, L.; Liu, J.; Zhu, M.; Kuang, W. Niujiaodihuang Detoxify Decoction Inhibits Ferroptosis by Enhancing Glutathione Synthesis in Acute Liver Failure Models. J. Ethnopharmacol. 2021, 279, 114305. [Google Scholar] [CrossRef] [PubMed]
- Demarco, B.; Chen, K.W.; Broz, P. Cross talk between intracellular pathogens and cell death. Immunol. Rev. 2020, 297, 174–193. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-derived bioactive peptides on inflammation and oxidative stress. BioMed. Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.L.; Kang, S.S.; Lu, C.; Choi, Y.J. Quantification of Multifunctional Dipeptide YA from Oyster Hydrolysate for Quality Control and Efficacy Evaluation. BioMed. Res. Int. 2018, 2018, 8437379. [Google Scholar] [CrossRef]
- Xie, C.L.; Kim, J.S.; Ha, J.M.; Choung, S.Y.; Choi, Y.J. Angiotensin I-converting enzyme inhibitor derived from cross-linked oyster protein. BioMed. Res. Int. 2014, 2014, 379234. [Google Scholar] [CrossRef]
- Byun, J.-H.; Shin, J.E.; Choi, Y.-J.; Choung, S.-Y. Oyster hydrolysate ameliorates ethanol diet–induced alcoholic fatty liver by regulating lipid metabolism in rats. Int. J. Food Sci. Technol. 2021, 56, 11. [Google Scholar] [CrossRef]
- Felig, P. The glucose-alanine cycle. Metab. Clin. Exp. 1973, 22, 179–207. [Google Scholar] [CrossRef]
- Okun, J.G.; Rusu, P.M.; Chan, A.Y.; Wu, Y.; Yap, Y.W.; Sharkie, T.; Schumacher, J.; Schmidt, K.V.; Roberts-Thomson, K.M.; Russell, R.D.; et al. Liver alanine catabolism promotes skeletal muscle atrophy and hyperglycaemia in type 2 diabetes. Nat. Metab. 2021, 3, 394–409. [Google Scholar] [CrossRef]
- McCommis, K.S.; Chen, Z.; Fu, X.; McDonald, W.G.; Colca, J.R.; Kletzien, R.F.; Burgess, S.C.; Finck, B.N. Loss of Mitochondrial Pyruvate Carrier 2 in the Liver Leads to Defects in Gluconeogenesis and Compensation via Pyruvate-Alanine Cycling. Cell Metab. 2015, 22, 682–694. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.R.; Flamm, S.L.; di Bisceglie, A.M.; Bodenheimer, H.C.; Public Policy Committee of the American Association for the Study of Liver, D. Serum Activity of Alanine Aminotransferase (ALT) as an Indicator of Health and Disease. Hepatology 2008, 47, 1363–1370. [Google Scholar] [CrossRef]
- Maezono, K.; Mawatari, K.; Kajiwara, K.; Shinkai, A.; Maki, T. Effect of alanine on D-galactosamine-induced acute liver failure in rats. Hepatology 1996, 24, 1211–1216. [Google Scholar] [CrossRef]
- Maezono, K.; Kajiwara, K.; Mawatari, K.; Shinkai, A.; Torii, K.; Maki, T. Alanine protects liver from injury caused by F-galactosamine and CCl4. Hepatology 1996, 24, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Litwack, G. Metabolism of Amino Acids. In Human Biochemistry; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar] [CrossRef]
- Levine, R.J.; Conn, H.O. Tyrosine metabolism in patients with liver disease. J. Clin. Investig. 1967, 46, 2012–2020. [Google Scholar] [CrossRef]
- Siregar, A.S.; Nyiramana, M.M.; Kim, E.-J.; Shin, E.-J.; Kim, C.-W.; Lee, D.; Hong, S.-G.; Han, J.; Kang, D. TRPV1 Is Associated with Testicular Apoptosis in Mice. J. Anim. Reprod. Biotechnol. 2019, 34, 7. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Siregar, A.S.; Kim, E.J.; Nyiramana, M.M.; Shin, E.J.; Han, J.; Sohn, J.T.; Kim, J.W.; Kang, D. Involvement of TREK-1 Channel in Cell Viability of H9c2 Rat Cardiomyoblasts Affected by Bupivacaine and Lipid Emulsion. Cells 2019, 8, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Acc. No. | Primer Sequences (5′–3′) | Expected Size (bp) |
|---|---|---|---|
| IL1b | NM_008361.4 | Sense: GTTGACGGACCCCAAAAGAT Antisense: TCGTTGCTTGGTTCTCCTTG | 440 |
| IL6 | NM_031168 | Sense: CTTCACAAGTCCGGAGAGGAG Antisense: TGGTCTTGGTCCTTAGCCACT | 489 |
| Tnf | D84199 | Sense: CAGCCTCTTCTCATTCCTGCT Antisense: TGTCCCTTGAAGAGAACCTGG | 339 |
| GAPDH | NM_017008.4 | Sense: CTA AAG GGC ATC CTG GGC Antisense: TTA CTC CTT GGA GGC CAT | 201 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siregar, A.S.; Nyiramana, M.M.; Kim, E.-J.; Cho, S.B.; Woo, M.S.; Lee, D.K.; Hong, S.-G.; Han, J.; Kang, S.S.; Kim, D.R.; et al. Oyster-Derived Tyr-Ala (YA) Peptide Prevents Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Suppressing Inflammatory, Apoptotic, Ferroptotic, and Pyroptotic Signals. Mar. Drugs 2021, 19, 614. https://doi.org/10.3390/md19110614
Siregar AS, Nyiramana MM, Kim E-J, Cho SB, Woo MS, Lee DK, Hong S-G, Han J, Kang SS, Kim DR, et al. Oyster-Derived Tyr-Ala (YA) Peptide Prevents Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Suppressing Inflammatory, Apoptotic, Ferroptotic, and Pyroptotic Signals. Marine Drugs. 2021; 19(11):614. https://doi.org/10.3390/md19110614
Chicago/Turabian StyleSiregar, Adrian S., Marie Merci Nyiramana, Eun-Jin Kim, Soo Buem Cho, Min Seok Woo, Dong Kun Lee, Seong-Geun Hong, Jaehee Han, Sang Soo Kang, Deok Ryong Kim, and et al. 2021. "Oyster-Derived Tyr-Ala (YA) Peptide Prevents Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Suppressing Inflammatory, Apoptotic, Ferroptotic, and Pyroptotic Signals" Marine Drugs 19, no. 11: 614. https://doi.org/10.3390/md19110614
APA StyleSiregar, A. S., Nyiramana, M. M., Kim, E.-J., Cho, S. B., Woo, M. S., Lee, D. K., Hong, S.-G., Han, J., Kang, S. S., Kim, D. R., Choi, Y. J., & Kang, D. (2021). Oyster-Derived Tyr-Ala (YA) Peptide Prevents Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Suppressing Inflammatory, Apoptotic, Ferroptotic, and Pyroptotic Signals. Marine Drugs, 19(11), 614. https://doi.org/10.3390/md19110614