Characterization of the CAZy Repertoire from the Marine-Derived Fungus Stemphylium lucomagnoense in Relation to Saline Conditions

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
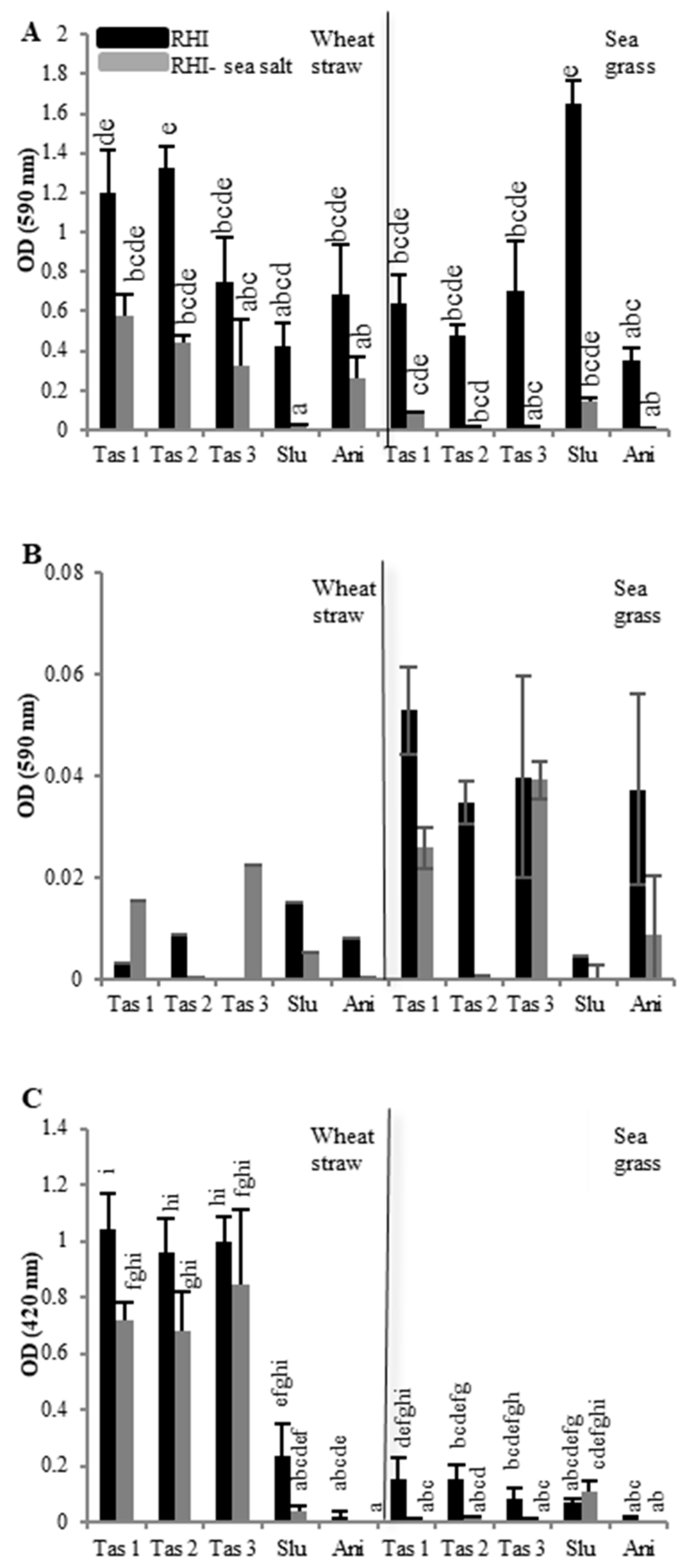
2.1. Effect of Carbon Sources and Sea Salt on Fungal Growth
2.2. Effect of Carbon Sources and Sea Salt on Lignocellulolytic Activities
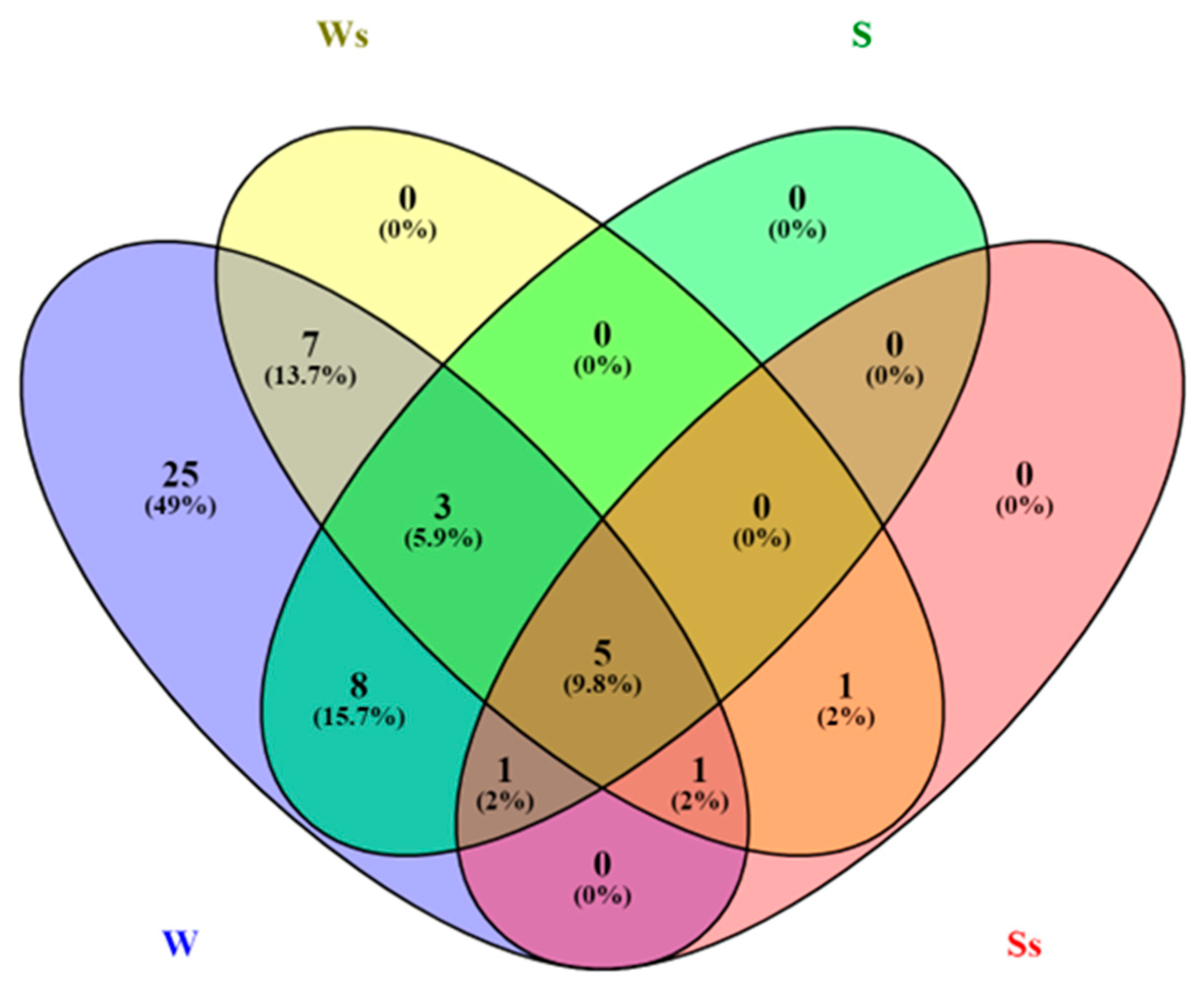
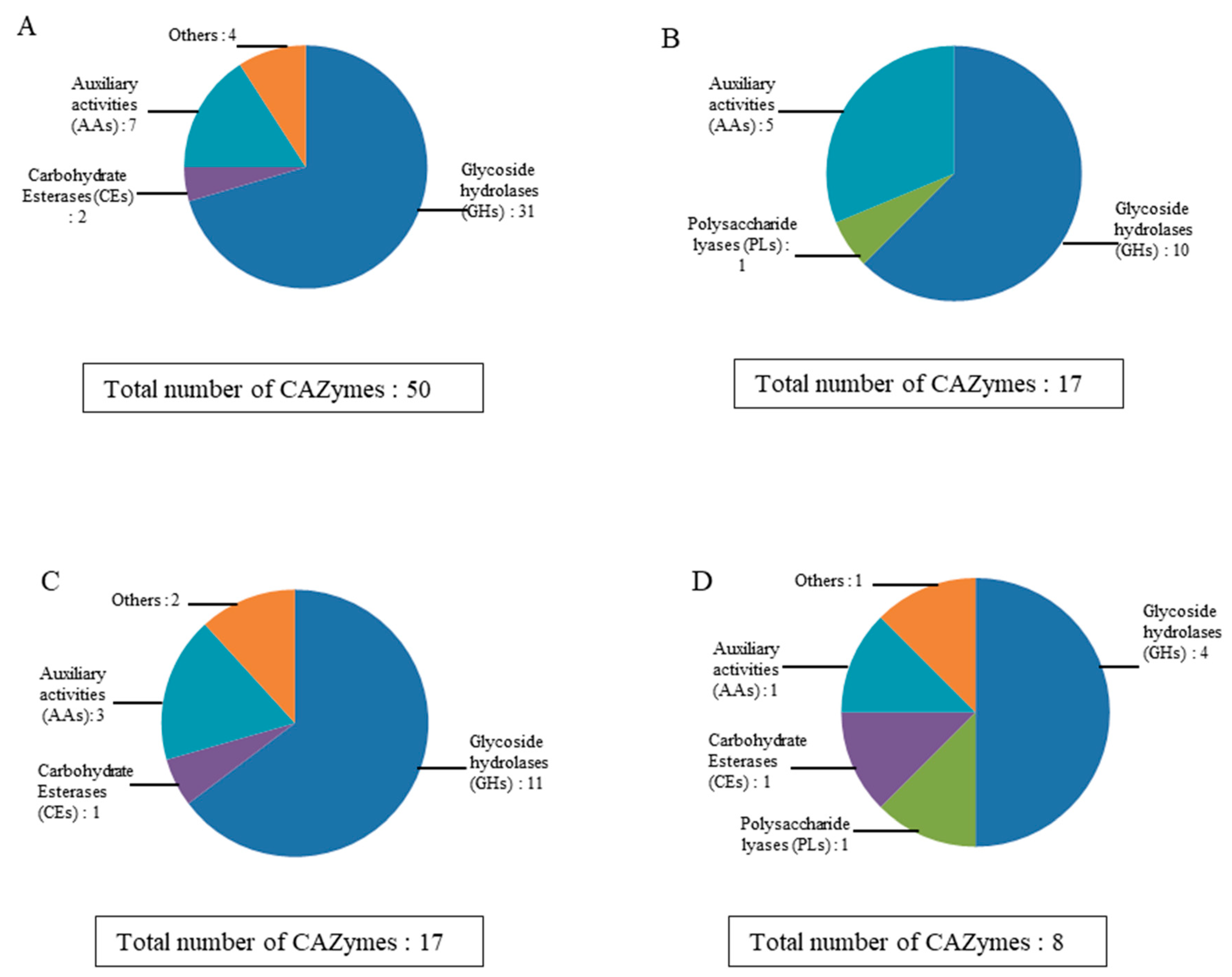
2.3. cDNA Library Construction and Sequencing, and Annotation of the Lignocellulolytic Enzyme Set from the Stemphylium Lucomagnoense Proteomes
2.4. Enzyme Distribution in Secretomes from Cultures on Wheat Straw and Seagrass, in Saline and Non-Saline Conditions
3. Discussion
4. Materials and Methods
4.1. Radial Growth Rate Determination
4.2. Screening of Lignocellulolytic Activities on Wheat Straw and Seagrass
4.3. Statistical Analysis of Data
4.4. RNA Extraction, cDNA Library Construction, Sequencing, Assembling and Annotation
4.5. Preparation of Stemphylium Lucomagnoense Secretomes
4.6. Proteomic Analysis of Secretomes
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Grosberg, R.K.; Vermeij, G.J.; Wainwright, P.C. Biodiversity in water and on land. Curr. Biol. 2012, 22, R900–R903. [Google Scholar] [CrossRef] [PubMed]
- Kohlmeyer, J.; Kohlmeyer, E. Marine fungi from tropical America and Africa. Mycologia 1971, 63, 831–861. [Google Scholar] [CrossRef] [PubMed]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the marine environment: Open questions and unsolved problems. mBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; De Gruyter, J.; De Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G. Marine fungi and fungal-like organisms. In Marine and Freshwater Botany; Pang, K.-L., Ed.; Walter De Gruyter Incorporated: Berlin, Germany, 2012; ISBN 3-11-026406-4. [Google Scholar]
- Kohlmeyer, J.; Volkmann-Kohlmeyer, B. Fungi from coral reefs: A commentary. Mycol. Res. 2003, 107, 385–387. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G. Diversity of fungal isolates from three Hawaiian marine sponges. Microbiol Res. 2009, 164, 233–241. [Google Scholar] [CrossRef]
- Jones, E.G. Are there more marine fungi to be described? Bot. Mar. 2011, 54, 343–354. [Google Scholar] [CrossRef]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef]
- Bonugli-Santos, R.C.; Dos Santos Vasconcelos, M.R.; Passarini, M.R.Z.; Vieira, G.A.L.; Lopes, V.C.P.; Mainardi, P.H.; Dos Santos, J.A.; de Azevedo Duarte, L.; Otero, I.V.R.; da Silva Yoshida, A.M.; et al. Marine-derived fungi: Diversity of enzymes and biotechnological applications. Front. Microbiol. 2015, 6, 269. [Google Scholar] [CrossRef]
- Batista-García, R.A.; Sutton, T.; Jackson, S.A.; Tovar-Herrera, O.E.; Balcázar-López, E.; del Rayo Sanchez-Carbente, M.; Sánchez-Reyes, A.; Dobson, A.D.; Folch-Mallol, J.L. Characterization of lignocellulolytic activities from fungi isolated from the deep-sea sponge Stelletta normani. PLoS ONE 2017, 12, e0173750. [Google Scholar] [CrossRef]
- Arfi, Y.; Chevret, D.; Henrissat, B.; Berrin, J.-G.; Levasseur, A.; Record, E. Characterization of salt-adapted secreted lignocellulolytic enzymes from the mangrove fungus Pestalotiopsis sp. Nat. Commun. 2013, 4, 1810. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Henrissat, B.; Arvas, M.; Syed, M.F.; Thieme, N.; Benz, J.P.; Sørensen, J.L.; Record, E.; Pöggeler, S.; Kempken, F. De novo assembly and genome analyses of the marine-derived Scopulariopsis brevicaulis strain LF580 unravels life-style traits and anticancerous scopularide biosynthetic gene cluster. PLoS ONE 2015, 10, e0140398. [Google Scholar] [CrossRef] [PubMed]
- Rédou, V.; Kumar, A.; Hainaut, M.; Henrissat, B.; Record, E.; Barbier, G.; Burgaud, G. Draft genome sequence of the deep-sea ascomycetous filamentous fungus Cadophora malorum Mo12 from the Mid-Atlantic Ridge reveals its biotechnological potential. Genome Announc. 2016, 4, e00467-16. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sørensen, J.L.; Hansen, F.T.; Arvas, M.; Syed, M.F.; Hassan, L.; Benz, J.P.; Record, E.; Henrissat, B.; Pöggeler, S. Genome sequencing and analyses of two marine fungi from the North Sea unraveled a plethora of novel biosynthetic gene clusters. Sci. Rep. 2018, 8, 10187. [Google Scholar] [CrossRef]
- Couturier, M.; Navarro, D.; Olivé, C.; Chevret, D.; Haon, M.; Favel, A.; Lesage-Meessen, L.; Henrissat, B.; Coutinho, P.M.; Berrin, J.-G. Post-genomic analyses of fungal lignocellulosic biomass degradation reveal the unexpected potential of the plant pathogen Ustilago maydis. BMC Genom. 2012, 13, 57. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Eisen, J.A. Characterization of the mycobiome of the seagrass, Zostera marina, reveals putative associations with marine chytrids. Front. Microbiol. 2019, 10, 2476. [Google Scholar] [CrossRef]
- Trevathan-Tackett, S.M.; Thomson, A.C.G.; Ralph, P.J.; Macreadie, P.I. Fresh carbon inputs to seagrass sediments induce variable microbial priming responses. Sci. Total Environ. 2018, 621, 663–669. [Google Scholar] [CrossRef]
- Hurtado-McCormick, V.; Kahlke, T.; Petrou, K.; Jeffries, T.; Ralph, P.J.; Seymour, J.R. Regional and microenvironmental scale characterization of the Zostera muelleri seagrass microbiome. Fron. Microbiol. 2019, 10, 1011. [Google Scholar] [CrossRef]
- Pilgaard, B.; Wilkens, C.; Herbst, F.-A.; Vuillemin, M.; Rhein-Knudsen, N.; Meyer, A.S.; Lange, L. Proteomic enzyme analysis of the marine fungus Paradendryphiella salina reveals alginate lyase as a minimal adaptation strategy for brown algae degradation. Sci. Rep. 2019, 9, 12338. [Google Scholar] [CrossRef]
- Orsi, W.D.; Richards, T.A.; Francis, W.R. Predicted microbial secretomes and their target substrates in marine sediment. Nat. Microbiol. 2018, 3, 32–37. [Google Scholar] [CrossRef]
- Ben Ali, W.; Chaduli, D.; Navarro, D.; Lechat, C.; Turbé-Doan, A.; Bertrand, E.; Faulds, C.B.; Sciara, G.; Lesage-Meessen, L.; Record, E.; et al. Screening of five marine-derived fungal strains for their potential to produce oxidases with laccase activities suitable for biotechnological applications. BMC Biotechnol. 2020, 20, 27. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef]
- Davis, L.L.; Bartnicki-Garcia, S. Chitosan synthesis by the tandem action of chitin synthetase and chitin deacetylase from Mucor rouxii. Biochemistry 1984, 23, 1065–1073. [Google Scholar] [CrossRef]
- Volk, H.; Marton, K.; Flajšman, M.; Radišek, S.; Tian, H.; Hein, I.; Podlipnik, Č.; Thomma, B.P.; Košmelj, K.; Javornik, B. Chitin-binding protein of Verticillium nonalfalfae disguises fungus from plant chitinases and suppresses chitin-triggered host immunity. Mol. Plant. Microbe Interact. 2019, 32, 1378–1390. [Google Scholar] [CrossRef]
- Mewis, K.; Lenfant, N.; Lombard, V.; Henrissat, B. Dividing the large glycoside hydrolase family 43 into subfamilies: A motivation for detailed enzyme characterization. Appl. Environ. Microb. 2016, 82, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Sakka, M.; Yamada, K.; Kitamura, T.; Kunitake, E.; Kimura, T.; Sakka, K. The modular arabinanolytic enzyme Abf43A-Abf43B-Abf43C from Ruminiclostridium josui consists of three GH43 modules classified in different subfamilies. Enzyme Microb. Tech. 2019, 124, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, M.M.; Kersten, P.J.; Cullen, D.; Whittaker, J.W. Identification of catalytic residues in glyoxal oxidase by targeted mutagenesis. J. Biol. Chem. 1999, 274, 36226–36232. [Google Scholar] [CrossRef] [PubMed]
- Daou, M.; Piumi, F.; Cullen, D.; Record, E.; Faulds, C.B. Heterologous production and characterization of two glyoxal oxidases from Pycnoporus cinnabarinus. Appl. Environ. Microb. 2016, 82, 4867–4875. [Google Scholar] [CrossRef]
- Whittaker, M.M.; Kersten, P.J.; Nakamura, N.; San ders-Loehr, J.; Schweizer, E.S.; Whittaker, J.W. Glyoxal oxidase from Phanerochaete chrysosporium is a new radical-copper oxidase. J. Biol. Chem. 1996, 271, 681–687. [Google Scholar] [CrossRef]
- Wymelenberg, A.V.; Sabat, G.; Mozuch, M.; Kersten, P.J.; Cullen, D.; Blanchette, R.A. Structure, organization, and transcriptional regulation of a family of copper radical oxidase genes in the lignin-degrading basidiomycete Phanerochaete chrysosporium. Appl. Environ. Microb. 2006, 72, 4871–4877. [Google Scholar] [CrossRef]
- Mathieu, Y.; Piumi, F.; Valli, R.; Aramburu, J.C.; Ferreira, P.; Faulds, C.B.; Record, E. Activities of secreted aryl alcohol quinone oxidoreductases from Pycnoporus cinnabarinus provide insights into fungal degradation of plant biomass. Appl. Environ. Microb. 2016, 82, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- Turbe-Doan, A.; Record, E.; Lombard, V.; Kumar, R.; Levasseur, A.; Henrissat, B.; Garron, M.-L. Trichoderma reesei dehydrogenase, a pyrroloquinoline quinone-dependent member of auxiliary activity family 12 of the carbohydrate-active Enzymes database: Functional and structural characterization. Appl. Environ. Microb. 2019, 85, e00964-19. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Ishida, T.; Yoshida, M.; Samejima, M.; Ohno, H.; Igarashi, K.; Nakamura, N. Crystal structure of the catalytic and cytochrome b domains in a eukaryotic pyrroloquinoline quinone-dependent dehydrogenase. Appl. Environ. Microb. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Várnai, A.; Umezawa, K.; Yoshida, M.; Eijsink, V.G. The pyrroloquinoline-quinone-dependent pyranose dehydrogenase from Coprinopsis cinerea drives lytic polysaccharide monooxygenase action. Appl. Environ. Microb. 2018, 84, e00156-18. [Google Scholar] [CrossRef]
- Netea, M.G.; Joosten, L.A.B.; Latz, E.; Mills, K.H.G.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.J.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, aaf1098. [Google Scholar] [CrossRef] [PubMed]
- Garajova, S.; Mathieu, Y.; Beccia, M.R.; Bennati-Granier, C.; Biaso, F.; Fanuel, M.; Ropartz, D.; Guigliarelli, B.; Record, E.; Rogniaux, H. Single-domain flavoenzymes trigger lytic polysaccharide monooxygenases for oxidative degradation of cellulose. Sci. Rep. 2016, 6, 28276. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, G.R.; Henrissat, B.; Davies, G.J.; Walton, P.H. Discovery and characterization of a new family of lytic polysaccharide monooxygenases. Nat. Chem. Biol. 2014, 10, 122–126. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, B.; Yang, Z.; Chen, Z.; Chen, B.; Deng, Y.; Jiang, Y.; van Peer, A.F. Identification and expression analysis of a new glycoside hydrolase family 55 exo-β-1,3-glucanase-encoding gene in Volvariella volvacea suggests a role in fruiting body development. Gene 2013, 527, 154–160. [Google Scholar] [CrossRef]
- Mouyna, I.; Hartl, L.; Latgé, J.-P. β-1,3-glucan modifying enzymes in Aspergillus fumigatus. Front. Microbiol. 2013, 4, 81. [Google Scholar] [CrossRef]
- Jiang, C.; Song, J.; Cong, H.; Zhang, J.; Yang, Q. Expression and characterization of a novel antifungal Exo-β-1,3-glucanase from Chaetomium cupreum. Appl. Biochem. Biotech. 2017, 182, 261–275. [Google Scholar] [CrossRef]
- Becker, S.; Tebben, J.; Coffinet, S.; Wiltshire, K.; Iversen, M.H.; Harder, T.; Hinrichs, K.-U.; Hehemann, J.-H. Laminarin is a major molecule in the marine carbon cycle. Proc. Natl. Acad. Sci. USA 2020, 117, 6599–6607. [Google Scholar] [CrossRef]
- Burtseva, Y.V.; Sova, V.; Mikhail, P.; Zvyagintseva, T. Enzymes of carbohydrate metabolism of mycelial fungi from marine environments. β-l,3-glucanase of the marine fungus Chaetomium indicum. Biochemistry 2000, 65, 1175–1183. [Google Scholar] [PubMed]
- Suriya, J.; Bharathiraja, S.; Krishnan, M.; Manivasagan, P.; Kim, S.-K. Chapter Eight—Marine Microbial Amylases: Properties and Applications. Adv. Food Nutr. Res. 2016, 79, 161–177. [Google Scholar] [PubMed]
- Wang, Y.; Barth, D.; Tamminen, A.; Wiebe, M.G. Growth of marine fungi on polymeric substrates. BMC Biotechnol. 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, M.N.; Ozdemir, S.C.; Uzel, A. Xylanase production from marine derived Trichoderma pleuroticola 08ÇK001 strain isolated from mediterranean coastal sediments. J. Basic. Microb. 2017, 57, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Ling, J.; Ahmad, M.; Lin, L.; Zhang, Y.; Wang, C. Culturable fungal diversity and cellulase production by mixed culture Aspergillus fungi from Sanya mangrove. J. Gen. Appl. Microbiol. 2018, 64, 293–298. [Google Scholar] [CrossRef]
- Theerachat, M.; Guieysse, D.; Morel, S.; Remaud-Siméon, M.; Chulalaksananukul, W. Laccases from marine organisms and their applications in the biodegradation of toxic and environmental pollutants: A review. Appl. Biochem. Biotech. 2019, 187, 583–611. [Google Scholar] [CrossRef]
- Bonugli-santos, R.C.; Durrant, L.R.; Sette, L.D. Laccase activity and putative laccase genes in marine-derived basidiomycetes. Fungal. Biol. 2010, 114, 863–872. [Google Scholar] [CrossRef]
- Bonugli-Santos, R.C.; Durrant, L.R.; da Silva, M.; Sette, L.D. Production of laccase, manganese peroxidase and lignin peroxidase by Brazilian marine-derived fungi. Enzym. Microb. Tech. 2010, 46, 32–37. [Google Scholar] [CrossRef]
- Atalla, M.M.; Zeinab, H.K.; Eman, R.H.; Amani, A.Y.; Abeer, A.A. Characterization and kinetic properties of the purified Trematosphaeria mangrovei laccase enzyme. Saudi J. Biol. Sci. 2013, 20, 373–381. [Google Scholar] [CrossRef]
- Wikee, S.; Hatton, J.; Turbé-Doan, A.; Mathieu, Y.; Daou, M.; Lomascolo, A.; Kumar, A.; Lumyong, S.; Sciara, G.; Faulds, C.B.; et al. Characterization and Dye Decolorization Potential of Two Laccases from the Marine-Derived Fungus Pestalotiopsis sp. Int. J. Mol. Sci. 2019, 20, 1864. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Hanse, B.; van Leeuwen, G.C.M.; Groenewald, J.Z.; Crous, P.W. Stemphylium revisited. Stud. Mycol. 2017, 87, 77–103. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2013, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Kern, M.; McGeehan, J.E.; Streeter, S.D.; Martin, R.N.A.; Besser, K.; Elias, L.; Eborall, W.; Malyon, G.P.; Payne, C.M.; Himmel, M.E.; et al. Structural characterization of a unique marine animal family 7 cellobiohydrolase suggests a mechanism of cellulase salt tolerance. Proc. Natl. Acad. Sci. USA 2013, 110, 10189–10194. [Google Scholar] [CrossRef] [PubMed]
- Patel, I.; Kracher, D.; Ma, S.; Garajova, S.; Haon, M.; Faulds, C.B.; Berrin, J.-G.; Ludwig, R.; Record, E. Salt-responsive lytic polysaccharide monooxygenases from the mangrove fungus Pestalotiopsis sp. NCi6. Biotechnol. Biofuels 2016, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Poidevin, L.; Berrin, J.-G.; Bennati-Granier, C.; Levasseur, A.; Herpoël-Gimbert, I.; Chevret, D.; Coutinho, P.M.; Henrissat, B.; Heiss-Blanquet, S.; Record, E. Comparative analyses of Podospora anserina secretomes reveal a large array of lignocellulose-active enzymes. Appl. Microbiol. Biot. 2014, 98, 7457–7469. [Google Scholar] [CrossRef] [PubMed]
- Ravalason, H.; Grisel, S.; Chevret, D.; Favel, A.; Berrin, J.-G.; Sigoillot, J.-C.; Herpoël-Gimbert, I. Fusarium verticillioides secretome as a source of auxiliary enzymes to enhance saccharification of wheat straw. Bioresour. Technol. 2012, 114, 589–596. [Google Scholar] [CrossRef]
- Ravalason, H.; Jan, G.; Mollé, D.; Pasco, M.; Coutinho, P.M.; Lapierre, C.; Pollet, B.; Bertaud, F.; Petit-Conil, M.; Grisel, S.; et al. Secretome analysis of Phanerochaete chrysosporium strain CIRM-BRFM41 grown on softwood. Appl. Microbiol. Biot. 2008, 80, 719. [Google Scholar] [CrossRef]
- Vogel, H.J. A convenient growth medium for Neurospora (medium N). Microb. Genet. Bull. 1956, 13, 42–43. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assembled Contig Statistics | |
|---|---|
| Total length of contigs | 110,739,726 bp |
| Total number of contigs | 350,657 |
| N25 stats | 25% of total contig length is contained in the 5614 contigs > = 2983 bp |
| N50 stats | 50% of total contig length is contained in the 26,298 contigs > = 504 bp |
| N75 stats | 75% of total contig length is contained in the 142,056 contigs > = 176 bp |
| Total GC count | 59,818,129 bp |
| % GC | 54.02 |
| Accession Number | Number of Total Spectra | Number of Unique Peptides | Predicted Protein Functions | Predicted Family (Subfamily) * | Induction | ||
|---|---|---|---|---|---|---|---|
| W/Ws | S/Ss | W/Ws | S/Ss | ||||
| APMZ2_prot13181 | 222/101 | 26/13 | 75/43 | 16/10 | Glucoamylase | GH15-CBM20 | S/Ss W/Ws |
| APMZ2_prot912 | 110/53 | 17/6 | 41/30 | 14/6 | Copper radical oxidase | AA5_1 | S/Ss W/Ws |
| APMZ2_prot15973 | 92/56 | 15/10 | 33/25 | 9/8 | β-1,3-glucanase | GH55 | S/Ss W/Ws |
| APMZ2_prot2472 | 58/16 | 10/7 | 24/13 | 9/7 | α-amylase | GH13_1 | S/Ss W/Ws |
| APMZ2_prot14594 | 50/0 | 3/0 | 31/0 | 3/0 | β-1,4-xylanase | GH10 | S W |
| APMZ2_prot3103 | 36/8 | 7/0 | 15/7 | 6/0 | α-mannosidase | GH47 | S W/Ws |
| APMZ2_prot1893 | 34/5 | 2/0 | 28/5 | 2/0 | α-l-rhamnosidase | GH78 | S W/Ws |
| APMZ2_prot27112 | 33/16 | 0/0 | 23/15 | 0/0 | Glucose/methanol/choline oxidoreductase (GMC) | AA3_2 | W/Ws |
| APMZ2_prot4629 | 29/18 | 5/8 | 20/13 | 5/6 | Pyrroloquinoline quinone-dependent oxidoreductase | AA12 | S/Ss W/Ws |
| APMZ2_prot29015 | 28/0 | 0/0 | 22/0 | 0/0 | α-glucosidase | GH31 | W |
| APMZ2_prot1520 | 20/0 | 2/0 | 18/0 | 2/0 | β-1,4-glucosidase | GH3 | S W |
| APMZ2_prot21697 | 15/2 | 0/0 | 12/2 | 0/0 | β-d-glucosaminidase | GH20 | W/Ws |
| APMZ2_prot29106 | 15/0 | 0/0 | 12/0 | 0/0 | α-glucosidase | GH31 | W |
| APMZ2_prot275 | 13/0 | 3/0 | 10/0 | 3/0 | Chitin deacetylase | CBM18-CE4-CBM18-CBM18 | S W |
| APMZ2_prot24323 | 12/0 | 2/0 | 11/0 | 2/0 | Glucose/methanol/choline oxidoreductase (GMC) | AA3_2 | S W |
| APMZ2_prot2902 | 12/5 | 0/0 | 4/0 | 0/0 | α,α-trehalase | GH37 | W/Ws |
| APMZ2_prot3532 | 11/0 | 0/0 | 9/0 | 0/0 | β-1,4-glucosidase | GH3 | W |
| APMZ2_prot3750 | 11/0 | 0/0 | 9/0 | 0/0 | β-1,4-glucosidase | GH3 | W |
| APMZ2_prot15280 | 11/0 | 0/0 | 10/0 | 0/0 | β-1,4-glucosidase | GH3 | W |
| APMZ2_prot8136 | 10/3 | 2/0 | 8/3 | 2/0 | Rhamnogalacturonyl hydrolase | GH105 | S W/Ws |
| APMZ2_prot26178 | 9/0 | 2/0 | 8/0 | 2/0 | Endo/Exo-β-1,4-glucanase | GH55 | W/S |
| APMZ2_prot15779 | 9/0 | 0/0 | 5/0 | 0/0 | α-mannosidase | GH47 | W |
| APMZ2_prot27204 | 8/8 | 0/2 | 7/7 | 0/2 | Glucoamylase | GH15-CBM20 | Ss W/Ws |
| APMZ2_prot7619 | 8/0 | 2/0 | 8/0 | 2/0 | Feruloyl esterase | CE1 | S W |
| APMZ2_prot18739 | 8/0 | 0/0 | 6/0 | 0/0 | Carbohydrate Binding | CBM50-CBM50-CBM50 | W |
| APMZ2_prot8482 | 7/4 | 0/0 | 7/4 | 0/0 | β-1,3-1,4-glucan endo-1,3- β-glucosidase | GH17 | W/Ws |
| APMZ2_prot17304 | 7/0 | 3/0 | 6/0 | 3/0 | β-1,3-galactanase | GH43_24-CBM35 | S W |
| APMZ2_prot11667 | 7/4 | 0/0 | 7/4 | 0/0 | β-glucuronidase/heparanase | GH79 | W/Ws |
| APMZ2_prot17211 | 7/0 | 0/0 | 7/0 | 0/0 | β-l-arabinofuranosidase | GH142 | W |
| APMZ2_prot21080 | 6/5 | 0/0 | 5/5 | 0/0 | Lytic polysaccharide mono-oxygenase (LPMO) | AA11 | W/Ws |
| APMZ2_prot9402 | 6/0 | 0/0 | 6/0 | 0/0 | α-l-arabinofuranosidase | GH51 | W |
| APMZ2_prot13544 | 6/0 | 0/0 | 3/0 | 0/0 | α-mannosidase | GH47 | W |
| APMZ2_prot21512 | 5/2 | 0/0 | 4/0 | 0/0 | β-d-glucosaminidase | GH20 | W |
| APMZ2_prot23159 | 5/0 | 0/0 | 5/0 | 0/0 | Carbohydrate-binding | CBM50 | W |
| APMZ2_prot27520 | 5/0 | 0/0 | 5/0 | 0/0 | β-1,4-glucosidase | GH1 | W |
| APMZ2_prot7701 | 4/0 | 2/2 | 4/0 | 2/2 | Chitin deacetylase | CE4-CBM18-CBM18 | S/Ss W |
| APMZ2_prot6391 | 4/0 | 0/0 | 4/0 | 0/0 | Lacto-N-biosidase | GH136 | W |
| APMZ2_prot20074 | 4/0 | 0/0 | 4/0 | 0/0 | Lacto-N-biosidase | GH136 | W |
| APMZ2_prot9251 | 4/0 | 0/0 | 4/0 | 0/0 | Chitinase | GH18 | W |
| APMZ2_prot29026 | 4/0 | 0/0 | 4/0 | 0/0 | Carbohydrate esterase | CE15 | W |
| APMZ2_prot1535 | 4/0 | 0/0 | 3/0 | 0/0 | β-1,3-1,4-glucan endo-1,3-β-glucosidase | GH17 | W |
| APMZ2_prot16320 | 3/0 | 3/0 | 3/0 | 3/0 | α-glucosidase | GH31 | S W |
| APMZ2_prot5721 | 3/0 | 0/0 | 3/0 | 0/0 | α-l-rhamnosidase | GH78 | W |
| APMZ2_prot21603 | 3/0 | 0/0 | 3/0 | 0/0 | α-l-arabinofuranosidase | GH43_26 | W |
| APMZ2_prot411 | 3/0 | 0/0 | 3/0 | 0/0 | β-1,4-xylanase | CBM1-GH10 | W |
| APMZ2_prot2229 | 3/0 | 0/0 | 3/0 | 0/0 | Glucose/methanol/choline oxidoreductase (GMC) | AA3_2 | W |
| APMZ2_prot19857 | 2/0 | 0/0 | 2/0 | 0/0 | Dextranase/isopullulanase | GH49 | W |
| APMZ2_prot23037 | 2/0 | 0/0 | 2/0 | 0/0 | α-l-arabinofuranosidase/β-d-xylosidase/α-l-arabinanase/β-d-galactofuranosidase | GH43_22-GH43_34 | W |
| APMZ2_prot3876 | 2/0 | 0/0 | 2/0 | 0/0 | β-d-glucosaminidase | GH20 | W |
| APMZ2_prot11475 | 0/6 | 0/0 | 0/6 | 0/0 | Glucooligosaccharide oxidase | AA7 | Ws |
| APMZ2_prot21161 | 0/3 | 0/3 | 0/3 | 0/3 | Alginate lyase | PL7_4 | Ss Ws |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Ali, W.; Navarro, D.; Kumar, A.; Drula, E.; Turbé-Doan, A.; Correia, L.O.; Baumberger, S.; Bertrand, E.; Faulds, C.B.; Henrissat, B.; et al. Characterization of the CAZy Repertoire from the Marine-Derived Fungus Stemphylium lucomagnoense in Relation to Saline Conditions. Mar. Drugs 2020, 18, 461. https://doi.org/10.3390/md18090461
Ben Ali W, Navarro D, Kumar A, Drula E, Turbé-Doan A, Correia LO, Baumberger S, Bertrand E, Faulds CB, Henrissat B, et al. Characterization of the CAZy Repertoire from the Marine-Derived Fungus Stemphylium lucomagnoense in Relation to Saline Conditions. Marine Drugs. 2020; 18(9):461. https://doi.org/10.3390/md18090461
Chicago/Turabian StyleBen Ali, Wissal, David Navarro, Abhishek Kumar, Elodie Drula, Annick Turbé-Doan, Lydie Oliveira Correia, Stéphanie Baumberger, Emmanuel Bertrand, Craig B. Faulds, Bernard Henrissat, and et al. 2020. "Characterization of the CAZy Repertoire from the Marine-Derived Fungus Stemphylium lucomagnoense in Relation to Saline Conditions" Marine Drugs 18, no. 9: 461. https://doi.org/10.3390/md18090461
APA StyleBen Ali, W., Navarro, D., Kumar, A., Drula, E., Turbé-Doan, A., Correia, L. O., Baumberger, S., Bertrand, E., Faulds, C. B., Henrissat, B., Sciara, G., Mechichi, T., & Record, E. (2020). Characterization of the CAZy Repertoire from the Marine-Derived Fungus Stemphylium lucomagnoense in Relation to Saline Conditions. Marine Drugs, 18(9), 461. https://doi.org/10.3390/md18090461