Temperature Differentially Affects Gene Expression in Antarctic Thraustochytrid Oblongichytrium sp. RT2316-13

 , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Sequence Analysis and Transcriptome Annotation
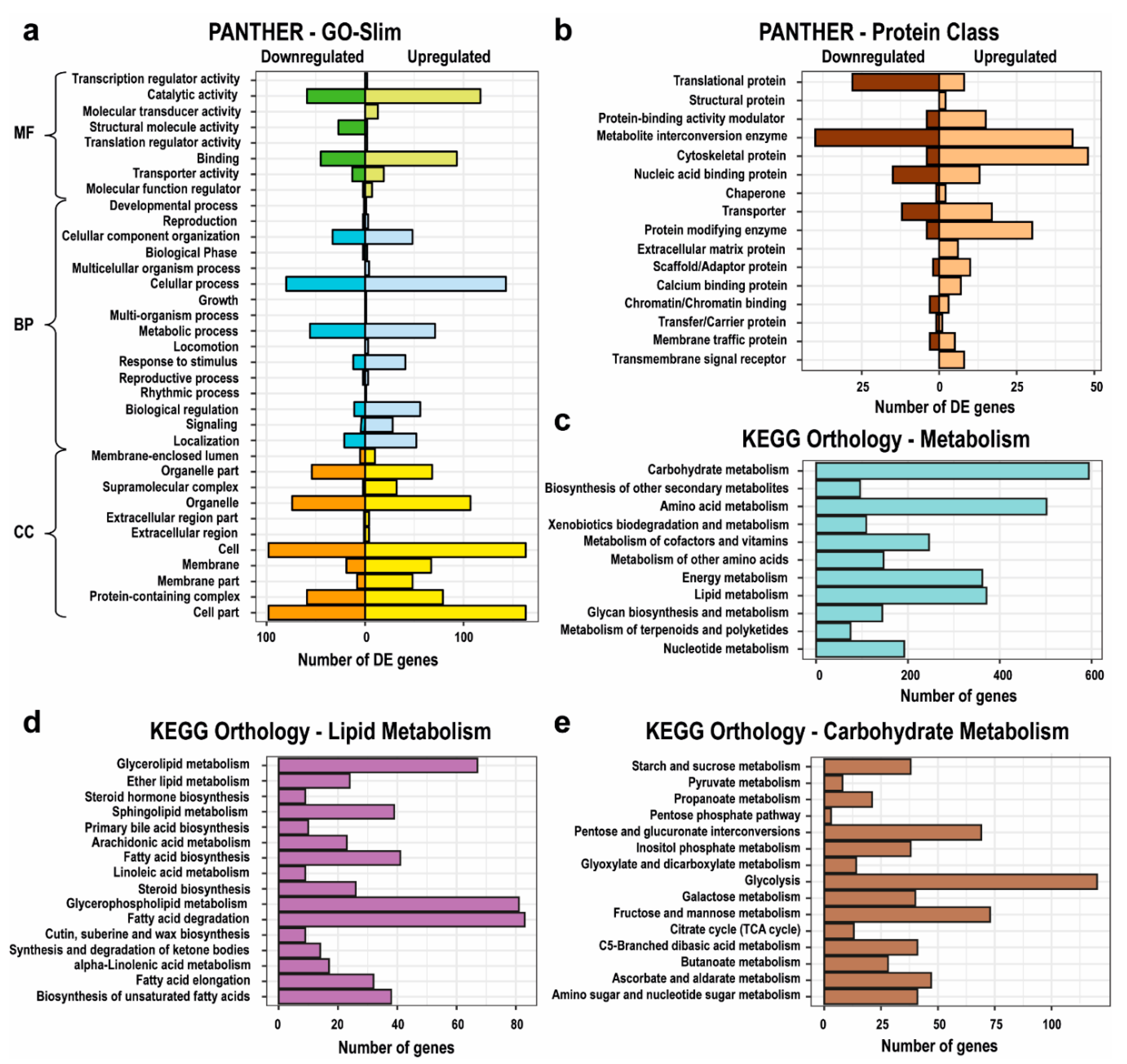
2.2. Differentially Expressed Genes
2.2.1. Cytoskeletal Proteins
2.2.2. Inositol Phosphate Metabolism
2.2.3. Glycolysis, Pentose Phosphate Pathway, and Tricarboxylic Acid Cycle
2.2.4. Respiratory Chain and Oxidative Phosphorylation
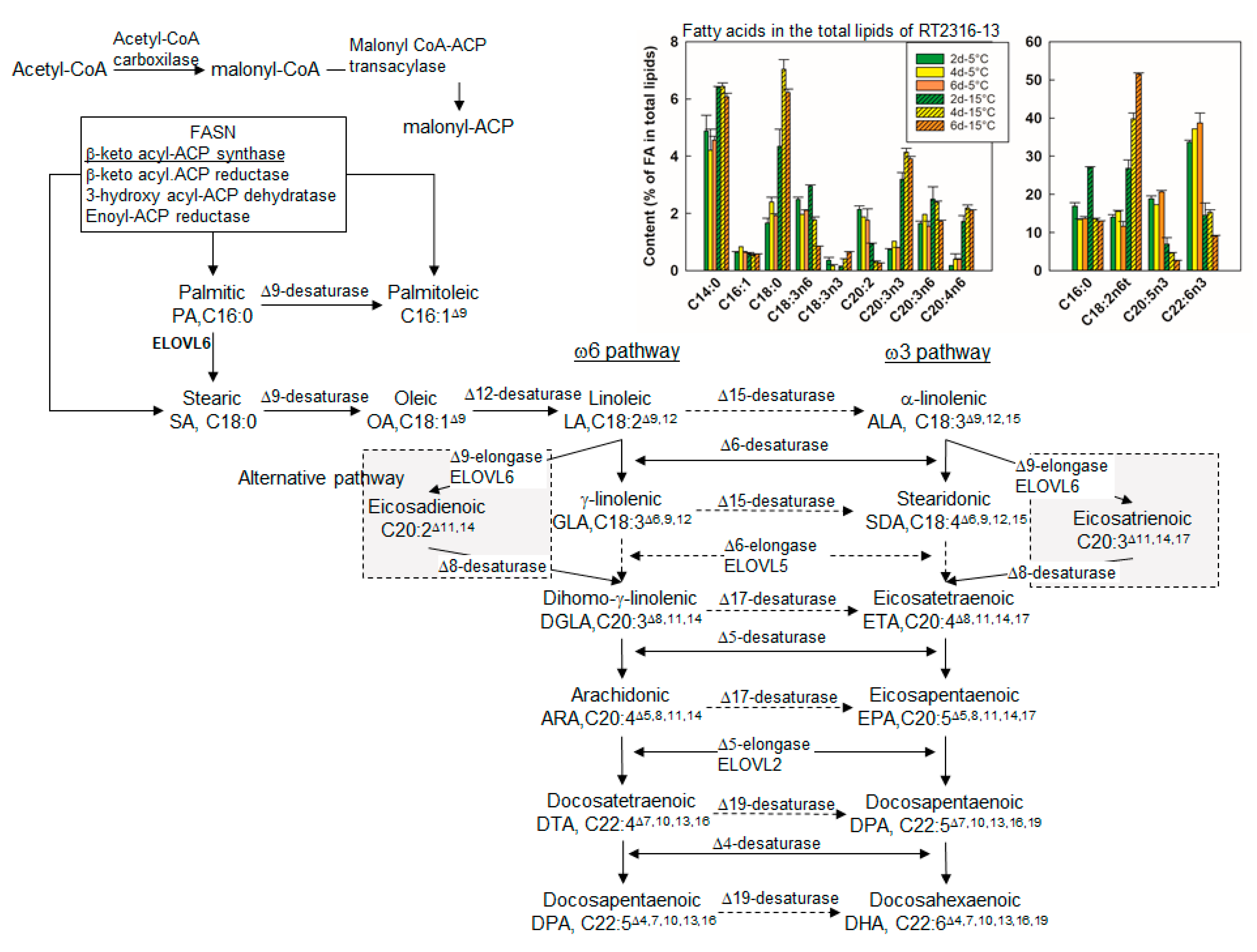
2.2.5. Lipid Metabolism
2.3. Validation of Gene Expression Profiles by qRT-PCR
3. Materials and Methods
3.1. Microorganism and Culture Conditions for RNA Extraction
3.2. RNA Extraction
3.3. Conversion of RNA to cDNA
3.4. Sequencing and Quality Control of RT2316-13 Transcriptome
3.5. Differential Gene Expression Analysis
3.6. Transcriptome Annotation and Enrichment Analysis
3.7. Validation of the Transcriptome Results
3.8. Data Availability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuratko, C.N.; Salem, N., Jr. Docosahexaenoic acid from algal oil. Eur. J. Lipid Sci. Technol. 2013, 115, 965–976. [Google Scholar] [CrossRef]
- Marchan, L.F.; Chang, K.J.L.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A Review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Narayan, B.; Miyashita, K.; Hosakawa, M. Physiological effects of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)—A Review. Food Rev. Int. 2006, 22, 291–307. [Google Scholar] [CrossRef]
- Allen, E.E.; Facciotti, D.; Bartlett, D.H. Monounsaturated but not polyunsaturated fatty acids are required for growth of the deep-sea bacterium Photobacterium profundum SS9 at high pressure and low temperature. Appl. Environ. Microbiol. 1999, 65, 1710–1720. [Google Scholar] [CrossRef] [PubMed]
- Kato, C.; Nogi, Y. Correlation between phylogenetic structure and function: Examples from deep-sea Shewanella. FEMS Microbiol. Ecol. 2001, 35, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, J.; Kurihara, T.; Yamamoto, K.; Nagayasu, M.; Tani, Y.; Mihara, H.; Hosokawa, M.; Baba, T.; Sato, S.B.; Esaki, N. Eicosapentaenoic acid plays a beneficial role in membrane organization and cell division of a cold-adapted bacterium, Shewanella livingstonensis Ac10. J. Bacteriol. 2009, 191, 632–640. [Google Scholar] [CrossRef]
- Stillwell, W.; Wassall, S.R. Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 2003, 126, 1–27. [Google Scholar] [CrossRef]
- Cullen, J.; Phillips, M.C.; Shipley, G.G. The effects of temperature on the composition and physical properties of the lipids of Pseudomonas fluorescens. Biochem. J. 1971, 125, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, M.C.; Cybulski, L.E.; Albanesi, D.; de Mendoza, D. Control of membrane lipid fluidity by molecular thermosensors. J. Bacteriol. 2004, 186, 6681–6688. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Murata, N. Membrane lipids in Cyanobacteria. In Lipids in Photosynthesis: Structure, Function and Genetics; Siegenthaler, P.-A., Murata, N., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherland, 1998; pp. 65–81. [Google Scholar] [CrossRef]
- Boelen, P.; van Dijk, R.; Damsté, J.S.S.; Rijpstra, W.I.C.; Buma, A.G.J. On the potential application of polar and temperate marine microalgae for EPA and DHA production. AMB Express 2013, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.W.; Jiang, Y.; Faan, Y.W.; Chen, F. Lipid characterization of mangrove thraustochytrid-Schizochytrium mangrovei. J. Agric. Food Chem. 2007, 55, 2906–2910. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, H.; Orikasa, Y.; Nishida, T. In Vivo conversion of triacylglycerol to docosahexaenoic acid-containing phospholipids in a thraustochytrid-like microorganism, strain 12B. Biotechnol. Lett. 2007, 29, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Shene, C.; Paredes, P.; Flores, L.; Leyton, A.; Asenjo, J.A.; Chisti, Y. Dynamic flux balance analysis of biomass and lipid production by Antarctic thraustochytrid Oblongichytrium sp. RT2316-13. Biotechnol. Bioeng. 2020, 117, 3006–3017. [Google Scholar] [CrossRef] [PubMed]
- Shene, C.; Paredes, P.; Vergara, D.; Leyton, A.; Garcés, M.; Flores, L.; Rubilar, M.; Bustamante, M.; Armenta, R. Antarctic thraustochytrids: Producers of long chain omega-3 polyunsaturated fatty acids. Microbiologyopen 2019, 9, e950. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Moss, S.T. (Ed.) Biology and phylogeny of the Labyrinthulales and Thraustochytriales. In Biology of Marine Fungi; Cambridge University Press: Cambridge, UK, 1986; pp. 105–130. [Google Scholar]
- Hao, L.; Scholey, J.M. Intraflagellar transport at a glance. J. Cell Sci. 2009, 122, 889–892. [Google Scholar] [CrossRef]
- Martin, T.F.J. Phosphoinositide lipids as signaling molecules: Common themes for signal transduction, cytoskeletal regulation, and membrane trafficking. Annu. Rev. Cell Dev. Biol. 1998, 14, 231–264. [Google Scholar] [CrossRef]
- Stolz, L.E.; Kuo, W.J.; Longchamps, J.; Sekhon, M.K.; York, J.D. INP51, a yeast inositol polyphosphate 5-phosphatase required for phosphatidylinositol 4,5-bisphosphate homeostasis and whose absence confers a cold-resistant phenotype. J. Biol. Chem. 1998, 273, 11852–11861. [Google Scholar] [CrossRef]
- Ruelland, E.; Cantrel, C.; Gawer, M.; Kader, J.C.; Zachowski, A. Activation of phospholipases C and D is an early response to a cold exposure in Arabidopsis suspension cells. Plant Physiol. 2002, 130, 999–1007. [Google Scholar] [CrossRef]
- Schade, A.; Walliser, C.; Wist, M.; Haas, J.; Vatter, P.; Kraus, J.M.; Filingeri, D.; Havenith, G.; Kestler, H.A.; Milner, J.D.; et al. Cool-temperature-mediated activation of phospholipase C-γ2 in the human hereditary disease PLAID. Cell. Signal. 2016, 28, 1237–1251. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. R. 2005, 69, 262–291. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.A.; Kornberg, H.L. Net formation of phosphoenolpyruvate from pyruvate by Escherichia coli. Biochim. Biophys. Acta 1965, 104, 618–620. [Google Scholar] [CrossRef]
- Xu, Y.F.; Létisse, F.; Absalan, F.; Lu, W.; Kuznetsova, E.; Brown, G.; Caudy, A.A.; Yakunin, A.F.; Broach, J.R.; Rabinowitz, J.D. Nucleotide degradation and ribose salvage in yeast. Mol. Syst. Biol. 2013, 9, 665. [Google Scholar] [CrossRef]
- Mager, W.H.; Planta, R.J. Coordinate expression of ribosomal protein genes in yeast as a function of cellular growth rate. Mol. Cell. Biochem. 1991, 104, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, M.G.; Camici, M.; Mascia, L.; Sgarrella, F.; Ipata, P.L. Pentose phosphates in nucleoside interconversion and catabolism. FEBS J. 2006, 273, 1089–1101. [Google Scholar] [CrossRef]
- Ma, Z.; Tan, Y.; Cui, G.; Feng, Y.; Cui, Q.; Song, X. Transcriptome and gene expression analysis of DHA producer Aurantiochytrium under low temperature conditions. Sci. Rep. 2015, 5, 14446. [Google Scholar] [CrossRef]
- Yazawa, K. Production of eicosapentaenoic acid from marine bacteria. Lipids 1996, 31, S297–S300. [Google Scholar] [CrossRef]
- Tanaka, M.; Ueno, A.; Kawasaki, K.; Yumoto, I.; Ohgiya, S.; Hoshino, T.; Ishizaki, K.; Okuyama, H.; Morita, N. Isolation of clustered genes that are notably homologous to the eicosapentaenoic acid biosynthesis gene cluster from the docosahexaenoic acid-producing bacterium Vibrio marinus strain MP-1. Biotechnol. Lett. 1999, 21, 939–945. [Google Scholar] [CrossRef]
- Allen, E.E.; Bartlett, D.H. Structure and regulation of the omega-3 polyunsaturated fatty acid synthase genes from the deep-sea bacterium Photobacterium profundum strain SS9. Microbiology 2002, 148, 1903–1913. [Google Scholar] [CrossRef]
- Metz, J.G.; Roessler, P.; Facciotti, D.; Levering, C.; Dittrich, F.; Lassner, M.; Valentine, R.; Lardizabal, K.; Domergue, F.; Yamada, A.; et al. Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science 2001, 293, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, S.; Liu, L.; Li, S.; Luo, Y.; Lv, C.; Wang, B.; Cheng, C.H.K.; Chen, H.; Yang, X. Genome sequencing and analysis of Thraustochytriidae sp. SZU445 provides novel insights into the polyunsaturated fatty acid biosynthesis pathway. Mar. Drugs 2020, 18, 118. [Google Scholar] [CrossRef] [PubMed]
- Damude, H.G.; Zhang, H.; Farrall, L.; Ripp, K.G.; Tomb, J.F.; Hollerbach, D.; Yadav, N.S. Identification of bifunctional Δ12/ω3 fatty acid desaturases for improving the ratio of ω3 to ω6 fatty acids in microbes and plants. Proc. Natl. Acad. Sci. USA 2006, 103, 9446–9451. [Google Scholar] [CrossRef] [PubMed]
- Garwin, J.L.; Klages, A.L.; Cronan, J.E., Jr. β-Ketoacyl-acyl carrier protein synthase II of Escherichia coli. Evidence for function in the thermal regulation of fatty acid synthesis. J. Biol. Chem. 1980, 255, 3263–3265. [Google Scholar]
- Kihara, A. Very long-chain fatty acids: Elongation, physiology and related disorders. J. Biochem. 2012, 152, 387–395. [Google Scholar] [CrossRef]
- Kim, R.J.; Kim, H.J.; Shim, D.; Suh, M.C. Molecular and biochemical characterizations of the monoacylglycerol lipase gene family of Arabidopsis thaliana. Plant J. 2016, 85, 758–771. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K.; Raghu, G.; Sharma, Y.V.R.K.; Biju, A.R.; Rajasekharan, M.V.; Shivaji, S. Increase in oxidative stress at low temperature in an Antarctic bacterium. Curr. Microbiol. 2011, 62, 544–546. [Google Scholar] [CrossRef]
- De Maayer, P.; Anderson, D.; Cary, C.; Cowan, D.A. Some like it cold: Understanding the survival strategies of psychrophiles. EMBO Rep. 2014, 15, 508–517. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; De Benedictis, V.; Ruggiero, F.M.; Petrosillo, G. Functional role of cardiolipin in mitochondrial bioenergetics. Biochim. Biophys. Acta 2014, 1837, 408–417. [Google Scholar] [CrossRef]
- Demel, R.A.; Kruyff, B. The function of sterols in membranes. Biochim. Biophys. Acta 1976, 457, 109–132. [Google Scholar] [CrossRef]
- Hemmi, H.; Julmanop, C.; Hirata, D.; Tsuchiya, E.; Takemoto, J.Y.; Miyakawa, T. The physiological roles of membrane ergosterol as revealed by the phenotypes of syr1/erg3 null mutant of Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 1995, 59, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Quilodrán, B.; Hinzpeter, I.; Hormazabal, E.; Quiroz, A.; Shene, C. Docosahexaenoic acid (C22:6n-3, DHA) and astaxanthin production by Thraustochytriidae sp. AS4-A1 a native strain with high similitude to Ulkenia sp.: Evaluation of liquid residues from food industry as nutrient sources. Enzyme Microb. Technol. 2010, 47, 24–30. [Google Scholar] [CrossRef]
- Shene, C.; Leyton, A.; Rubilar, M.; Pinelo, M.; Acevedo, F.; Morales, E. Production of lipids and docosahexaenoic acid by a native Thraustochytrium strain. Eur. J. Lipid Sci. Technol. 2013, 115, 890–900. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, e323. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Huang, J.X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol update for large-scale genome and gene function analysis with the PANTHER classification system (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Metric | Value |
|---|---|
| Total raw reads (bp) | 174,570,852 |
| Total high quality reads (bp) | 173,659,258 |
| High quality content (%) | 99.47 |
| Number of transcripts | 118,595 |
| Number of genes | 69,220 |
| Total size (Mbp) | 147.5 |
| N50 (bp) | 2518 |
| Average length (bp) | 1243 |
| Median length (bp) | 554 |
| Library § | Quantity | Percent of Total (%) |
|---|---|---|
| Complete BUSCOs | 223 | 87.4 |
| Single-copy BUSCOs | 125 | 49.0 |
| Duplicated BUSCOs | 98 | 38.4 |
| Fragmented BUSCOs | 12 | 4.7 |
| Missing BUSCOs | 20 | 7.9 |
| Total BUSCO genes | 255 | 100 |
| Enzyme (Reaction) | EC Number | Swiss Prot ID |
|---|---|---|
| Fatty acid biosynthesis | ||
| Acetyl-CoA carboxylase | 6.4.1.2 | ACACB_HUMAN; ACAC_YEAST; ACAC_SCHPO; ACACA_MOUSE |
| Fatty acid synthase, FASN | 2.3.1.85 | FAS_HUMAN |
| Fatty acid synthase subunit β | 2.3.1.86 | FAS1_CANAX |
| Fatty acid synthase subunit α | 2.3.1.86 | FAS2_PENPA |
| Fatty acid synthase β subunit aflB | ATNM_EMENI | |
| Malonyl CoA-acyl carrier protein transacylase | 2.3.1.39 | FABD_BACSU |
| 3-Oxoacyl-[acyl-carrier-protein] reductase FabG | 1.1.1.100 | FABG_THEMA; FABG_RICPR; FABG_VIBCH |
| 3-Oxoacyl-[acyl-carrier-protein] synthase * | 2.3.1.179 | KASM_ARATH |
| Hydroxyacyl-thioester dehydratase type 2 | 4.2.1. | HTD2_HUMAN |
| Fatty acyl-CoA synthetase A | 6.2.1.3 | FCSA_DICDI |
| Very long-chain acyl-CoA synthetase | 6.2.1.3 | S27A2_MOUSE; S27A2_HUMAN |
| Enoyl-CoA hydratase ACTT6 § | 5.3.3.14 | ACTT6_ALTAL |
| Fatty acid elongation | ||
| 3-Ketoacyl-CoA thiolase | 2.3.1.16 | THIKB_RAT |
| Trifunctional enzyme subunit β | 2.3.1.16 | ECHB_MACFA; ECHB_BOVIN |
| Trifunctional enzyme subunit α | 2.3.1.16 | ECHA_HUMAN; ECHA_PIG |
| 3-Hydroxyacyl-CoA dehydrogenase | 1.1.1.35 | HCD2_DROME; HCDH2_CAEEL; HCDH1_CAEEL |
| Enoyl-CoA hydratase | 4.2.1.17 | ECHM_BOVIN; ECHM_DICDI; ECHM_RAT |
| Enoyl-[acyl-carrier-protein] reductase 1 | 1.3.1.; 1.3.1.38 | ETR1_DEBHA; MECR_DICDI |
| Lysosomal thioesterase PPT2-A | 3.1.2.22 | PPT2A_XENLA |
| Palmitoyl-protein thioesterase 1 | 3.1.2.22 | PPT1_MACFA |
| Elongation of very long chain fatty acids protein 2 (ELOVL2) ¥ | 2.3.1.199 | ELOV2_HUMAN |
| Elongation of very long chain fatty acids protein 4 (ELOVL4) £ | 2.3.1.199 | ELOV4_HUMAN; ELOV4_MACMU |
| Elongation of very long chain fatty acids protein 6 (ELOVL6) ‡ | 2.3.1.199 | ELO6_CAEEL; ELOV6_DANRE; ELOV6_MOUSE |
| Putative elongation of fatty acids protein | 2.3.1.199 | Y2012_DICDI |
| Very-long-chain 3-oxoacyl-CoA reductase | 1.1.1.330 | MKAR_LACBS; KCR1_ARATH |
| Very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase | 4.2.1.134 | HACD_CAEEL |
| Very-long-chain enoyl-CoA reductase | 1.3.1.93 | TECR_ARATH; TECR_DICDI |
| Cytosolic acyl coenzyme A thioester hydrolase | 3.1.2.2 | BACH_HUMAN; BACH_RAT |
| Biosynthesis of unsaturated fatty acids | ||
| Acyl-CoA desaturase (Δ9 desaturase) | 1.14.19.1 | ACOD2_MOUSE |
| Acyl-CoA desaturase 1 | 1.14.19.1 | SCD1_TACFU |
| Acyl-CoA 6-desaturase | 1.14.19.3 | LLCD_SYNY3; FADS2_PONAB |
| Delta(12) fatty acid desaturase FAD2 | 1.14.19.6 | FAD2_CALOF |
| Acyl-lipid (7-3)-desaturase (Δ4 desaturase) | 1.14.19.31 | D4FAD_EUGGR; D4FAD_THRSP |
| Sphingolipid delta(4)-desaturase | 1.14.19.17; 1.14.18.5 | DEGS_KOMPG; DEGS_CANAL |
| Delta(8)-fatty-acid desaturase | 1.14.19.3 | SLD1_EUGGR ⁑; SLD2_ARATH |
| Acyl-CoA thioesterase 2 | 3.1.2. | TESB_ECOLI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paredes, P.; Larama, G.; Flores, L.; Leyton, A.; Ili, C.G.; Asenjo, J.A.; Chisti, Y.; Shene, C. Temperature Differentially Affects Gene Expression in Antarctic Thraustochytrid Oblongichytrium sp. RT2316-13. Mar. Drugs 2020, 18, 563. https://doi.org/10.3390/md18110563
Paredes P, Larama G, Flores L, Leyton A, Ili CG, Asenjo JA, Chisti Y, Shene C. Temperature Differentially Affects Gene Expression in Antarctic Thraustochytrid Oblongichytrium sp. RT2316-13. Marine Drugs. 2020; 18(11):563. https://doi.org/10.3390/md18110563
Chicago/Turabian StyleParedes, Paris, Giovanni Larama, Liset Flores, Allison Leyton, Carmen Gloria Ili, Juan A. Asenjo, Yusuf Chisti, and Carolina Shene. 2020. "Temperature Differentially Affects Gene Expression in Antarctic Thraustochytrid Oblongichytrium sp. RT2316-13" Marine Drugs 18, no. 11: 563. https://doi.org/10.3390/md18110563
APA StyleParedes, P., Larama, G., Flores, L., Leyton, A., Ili, C. G., Asenjo, J. A., Chisti, Y., & Shene, C. (2020). Temperature Differentially Affects Gene Expression in Antarctic Thraustochytrid Oblongichytrium sp. RT2316-13. Marine Drugs, 18(11), 563. https://doi.org/10.3390/md18110563