Novel Efficient Bioprocessing of Marine Chitins into Active Anticancer Prodigiosin

 , ,
, ,  ,
, 
 ,
, 
Abstract
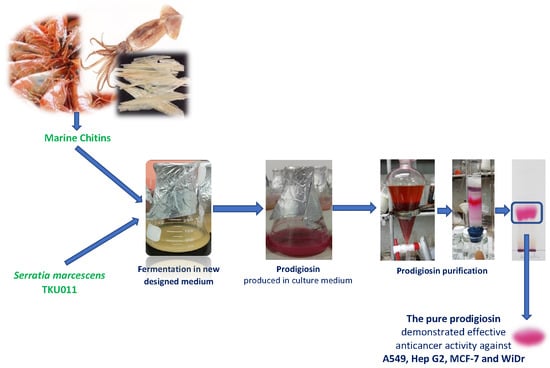
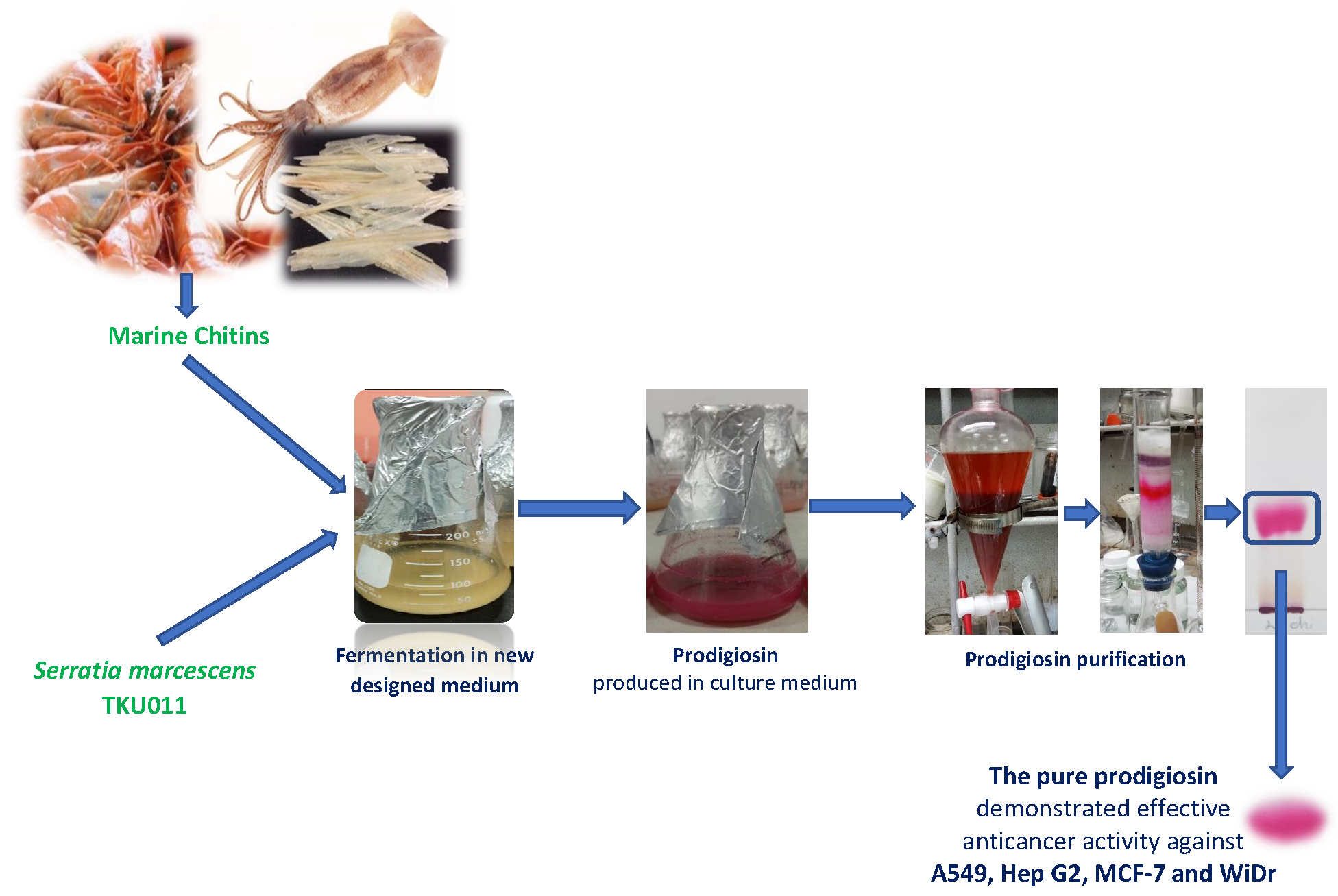
1. Introduction
2. Results and Discussion
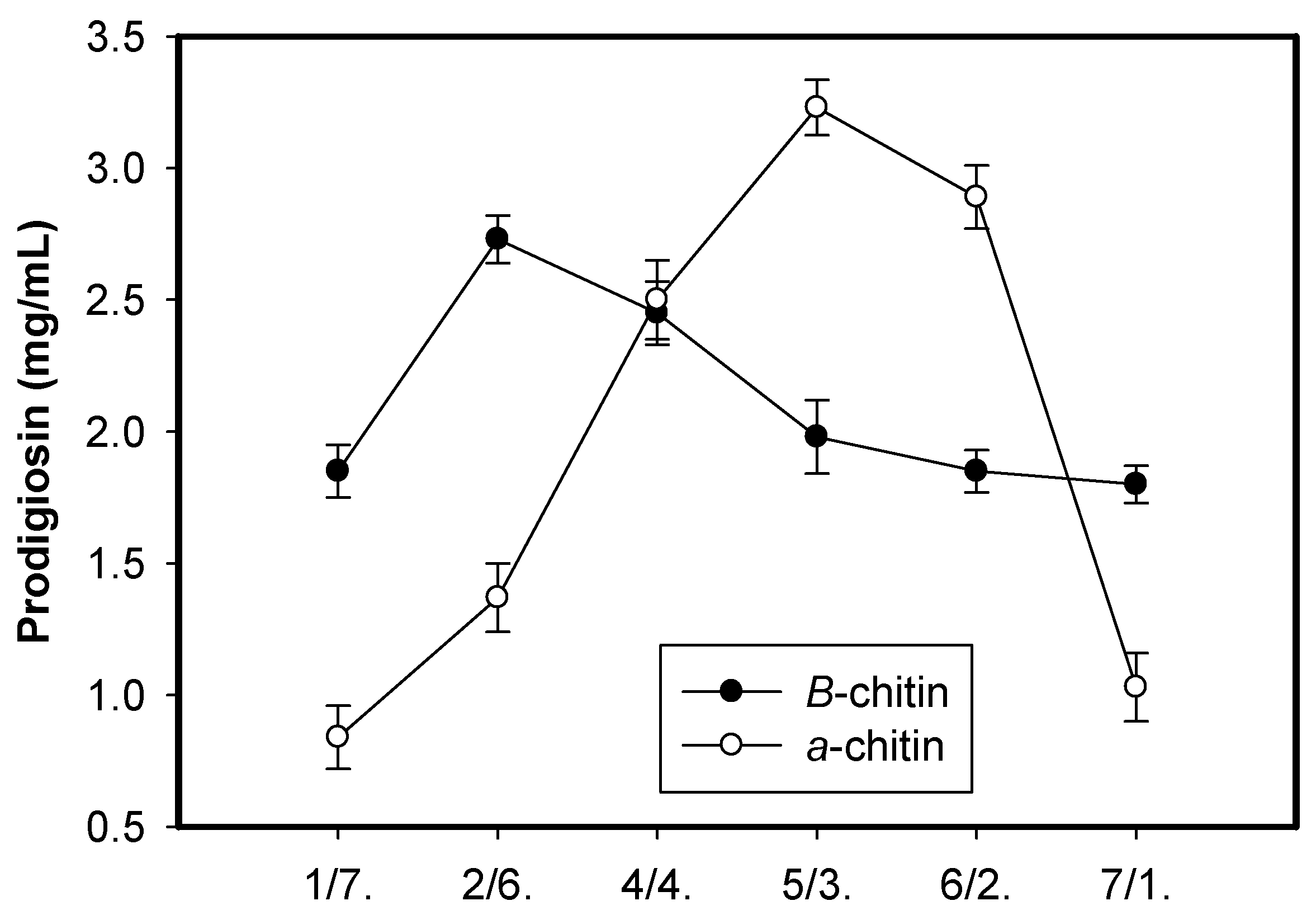
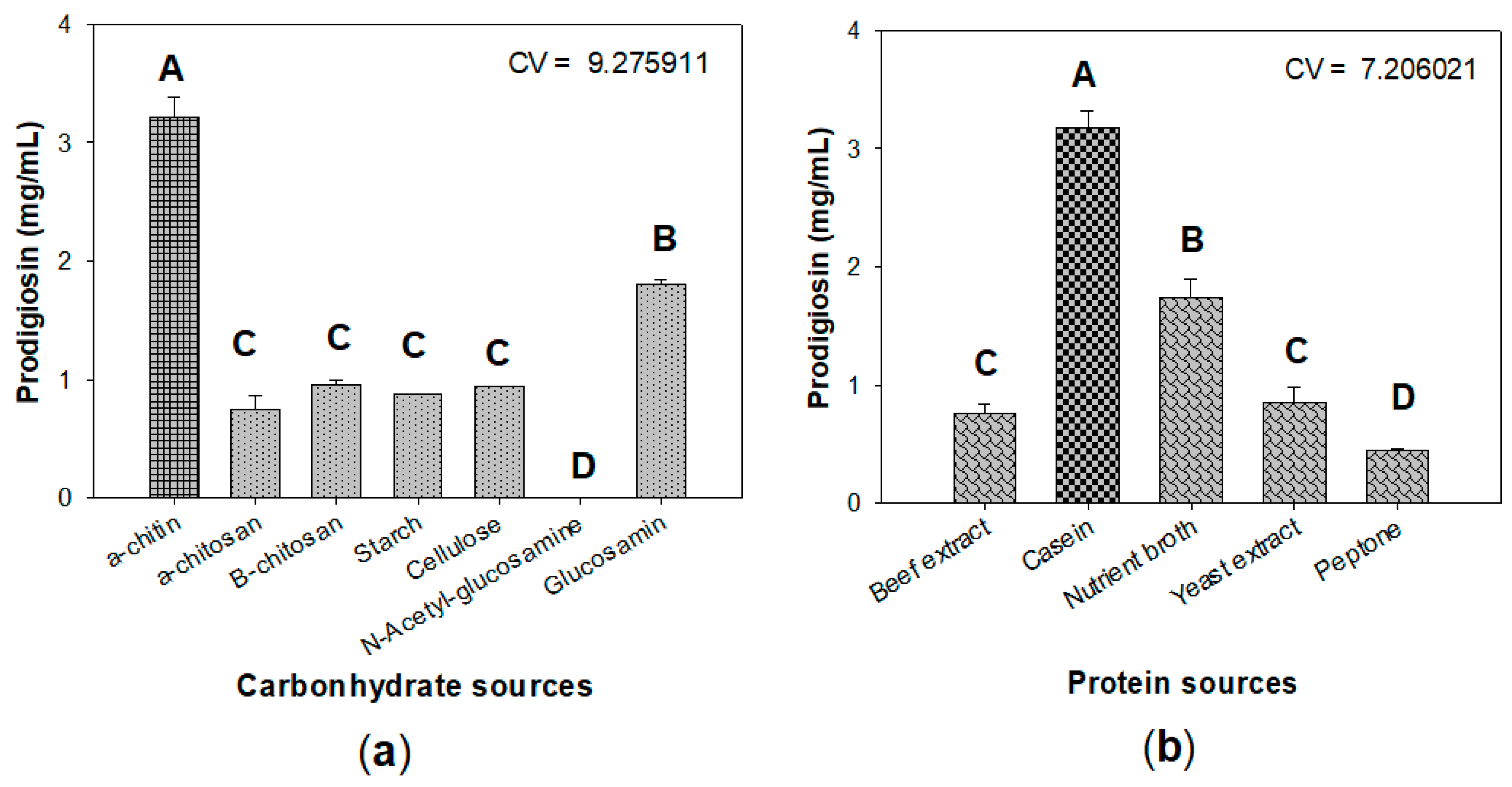
2.1. New Records of Marine α-Chitin as the Potential Carbon Source for Prodigiosin Synthesis by Serratia marcescens
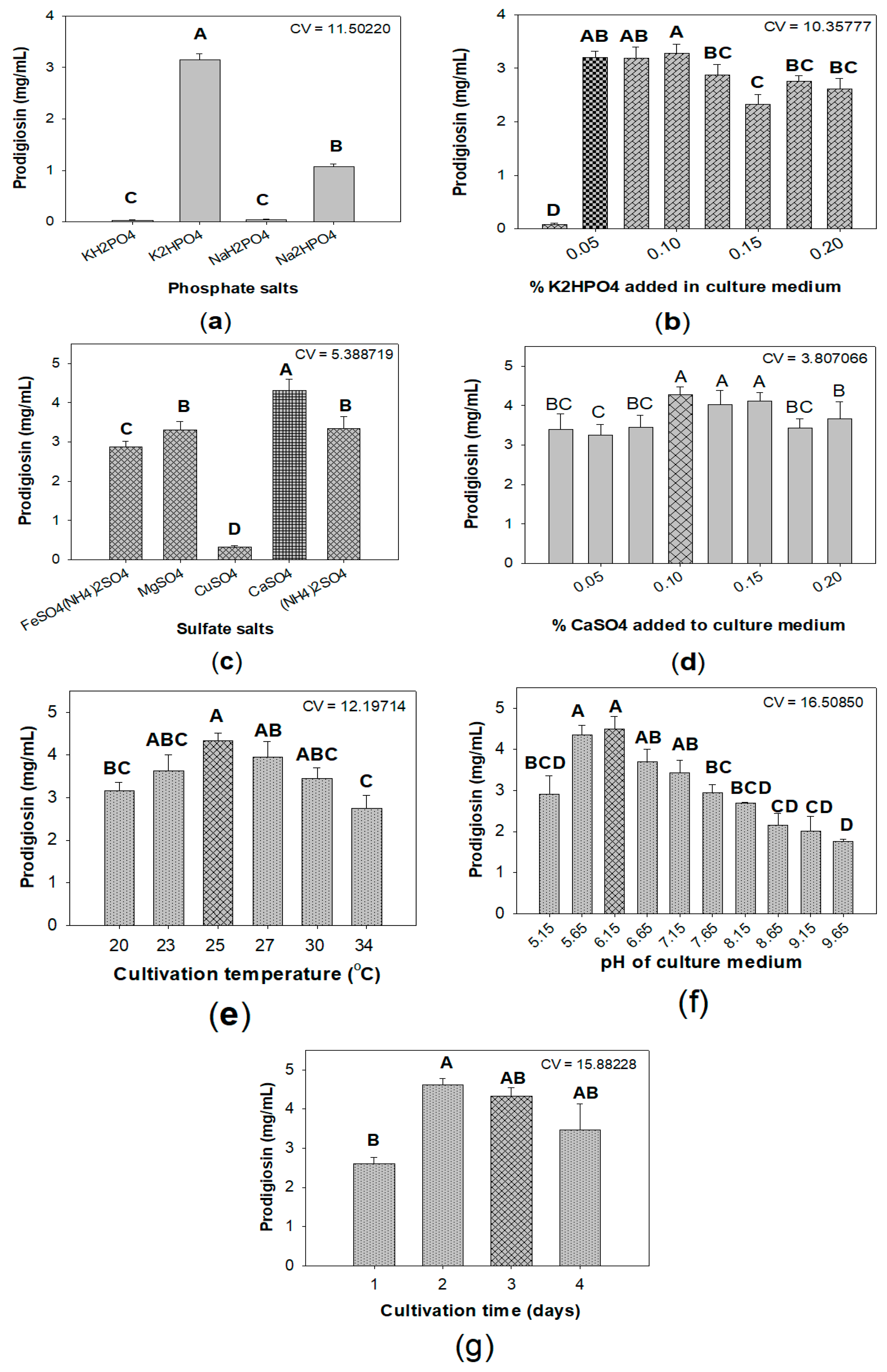
2.2. Optimization of Culture Conditions for Enhancement of Prodigiosin Production by Serratia marcescens
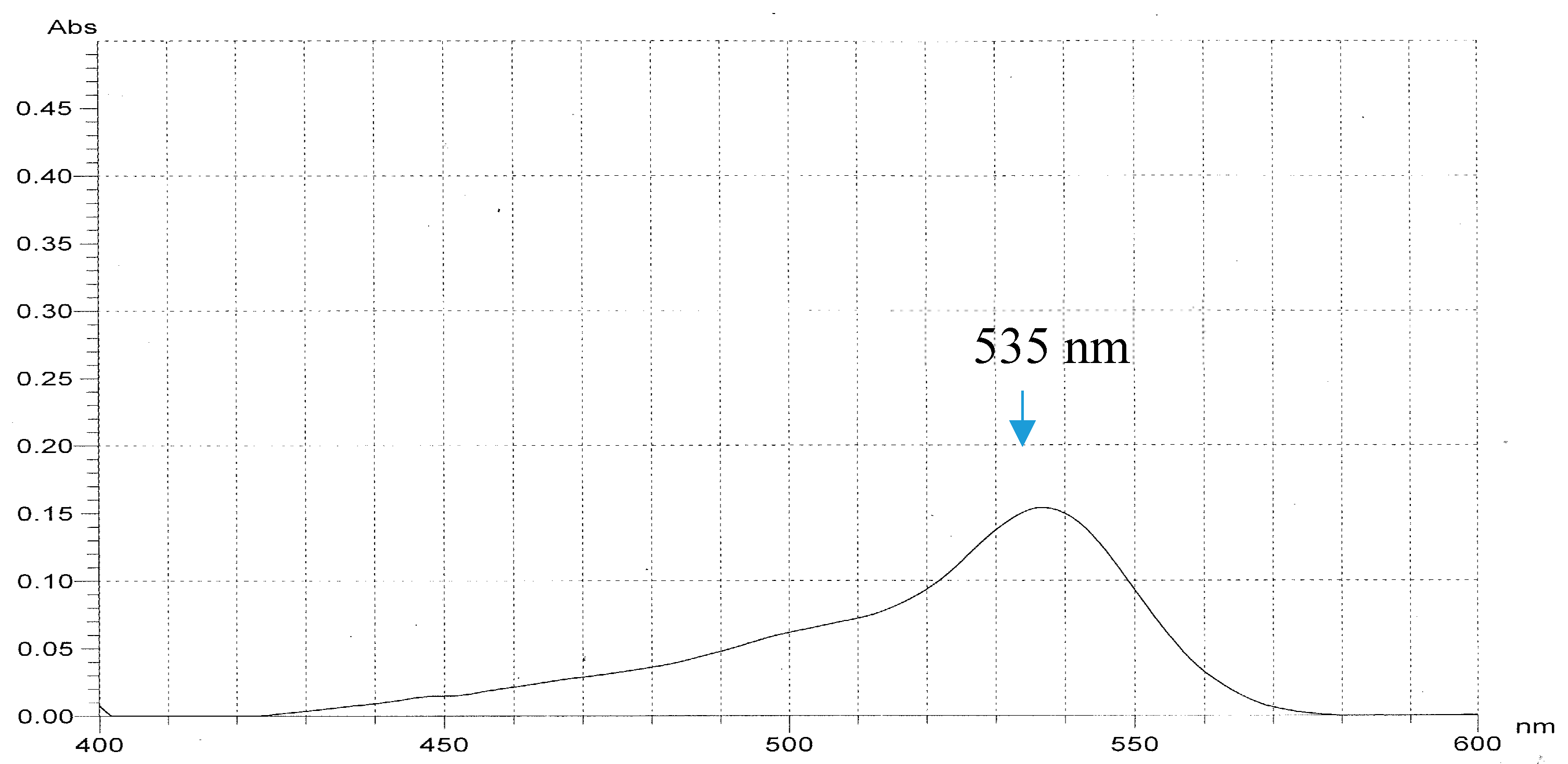
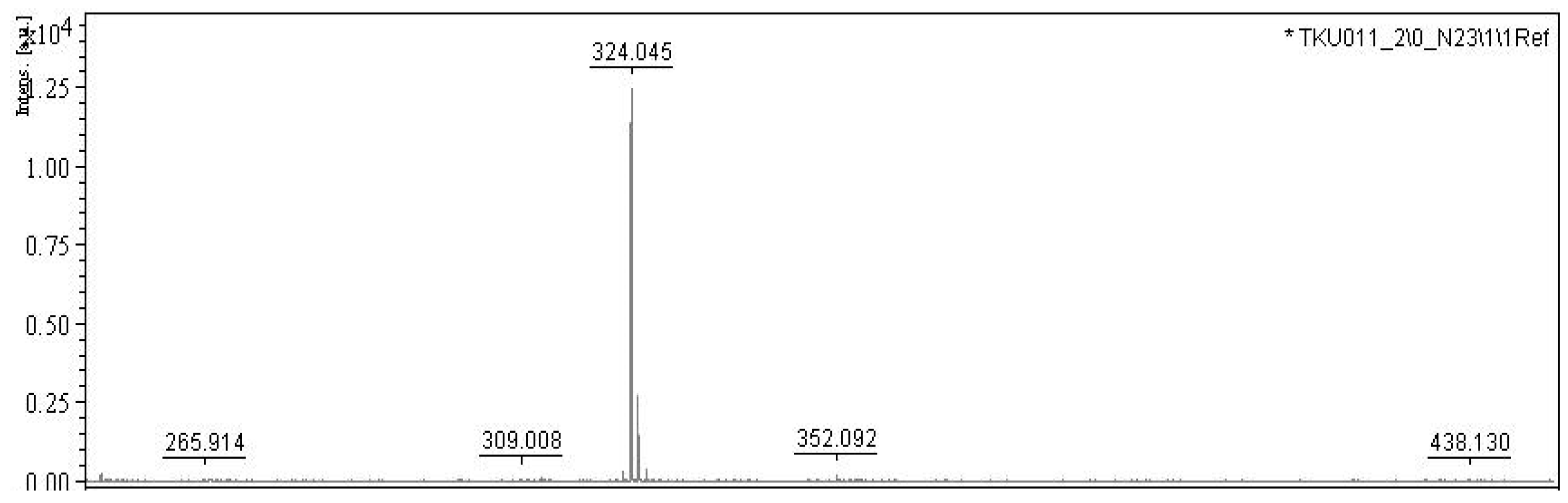
2.3. Purification and Qualification of Prodigiosin from Fermented Medium Containing α-Chitin
2.4. Evaluation of Inhibitory Effect of Prodigiosin against Cancerous Cell Lines Models
3. Materials and Methods
3.1. Materials
3.2. Fermentation for Prodigiosin Biosynthesis by S. marcescens TKU011
3.3. Prodigiosin Quilification and Purification
3.4. Biological Activity Assays
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nguyen, V.B.; Wang, S.L. Reclamation of marine chitinous materials for the production of α-glucosidase inhibitors via microbial conversion. Mar. Drugs 2017, 15, 350. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Tseng, S.C.; Wang, S.L. Production and characterization of antioxidant properties of exopolysaccharides from Paenibacillus mucilaginosus TKU032. Mar. Drugs 2016, 14, 40. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Wang, S.L. Recent advances in exopolysaccharides from Paenibacillus spp.: Production, isolation, structure, and bioactivities. Mar. Drugs 2015, 13, 1847–1863. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Wu, C.C.; Cheng, W.T.; Chen, Y.C.; Wang, C.L.; Wang, I.L.; Wang, S.L. Exopolysaccharides and antimicrobial biosurfactants produced by Paenibacillus macerans TKU029. Appl. Biochem. Biotechnol. 2014, 172, 933–950. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, W.T.; Lin, Z.H.; Kuo, Y.H.; Nguyen, A.D.; Pan, P.S.; Wang, S.L. An amphiprotic novel chitosanase from Bacillus mycoides and its application in the production of chitooligomers with their antioxidant and anti-inflammatory evaluation. Int. J. Mol. Sci. 2016, 17, 1302. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Liang, T.W.; Liu, K.C.; Hsu, Y.W.; Hsu, H.C.; Wang, S.L. Isolation and identification of a novel antioxidant with antitumor activity from Serratia ureilytica using squid pen as fermentation substrate. Mar. Biotechnol. 2011, 13, 451–461. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, T.H.; Doan, C.T.; Tran, T.N.; Nguyen, A.D.; Kuo, Y.-H.; Wang, S.-L. Production and bioactivity-guided isolation of antioxidants with α-glucosidase inhibitory and anti-NO properties from marine chitinous materials. Molecules 2018, 23, 1124. [Google Scholar] [CrossRef]
- Wang, S.L.; Huang, T.Y.; Wang, C.Y.; Liang, T.W.; Yen, Y.H.; Sakata, Y. Bioconversion of squid pen by Lactobacillus paracasei subsp paracasei TKU010 for the production of proteases and lettuce enhancing biofertilizers. Bioresour. Technol. 2008, 99, 5436–5443. [Google Scholar] [CrossRef]
- Wang, S.L.; Wang, C.Y.; Yen, Y.H.; Liang, T.W.; Chen, S.Y.; Chen, C.H. Enhanced production of insecticidal prodigiosin from Serratia marcescens TKU011 in media containing squid pen. Process Biochem. 2012, 47, 1684–1690. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, C.H.; Wang, S.L. Production of insecticidal materials from Pseudomonas tamsuii. Res. Chem. Intermed. 2015, 41, 7965–7971. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, S.Y.; Yen, Y.H.; Liang, T.W. Utilization of chitinous materials in pigment adsorption. Food Chem. 2012, 135, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Lo, B.C.; Wang, S.L. Chitinolytic bacteria-assisted conversion of squid pen and its effect on dyes and adsorption. Mar. Drugs 2015, 13, 4576–4593. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Utilization of fishery processing by-product squid pens for α-glucosidase inhibitors production by Paenibacillus sp. Mar. Drugs 2017, 15, 274. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Anti-α-glucosidase activity by a protease from Bacillus licheniformis. Molecules 2019, 24, 691. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Su, Y.C.; Nguyen, V.B.; Nguyen, A.D. Reclamation of shrimp heads for the production of α-glucosidase inhibitors by Staphylococcus sp. TKU043. Res. Chem. Intermed. 2018, 44, 4929–4937. [Google Scholar] [CrossRef]
- Hsu, C.H.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Conversion of shrimp heads to α-glucosidase inhibitors via co-culture of Bacillus mycoides TKU040 and Rhizobium sp. TKU041. Res. Chem. Intermed. 2017, 44, 4597–4607. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, A.D.; Kuo, Y.-H.; Wang, S.-L. Biosynthesis of α-glucosidase inhibitors by a newly isolated bacterium, Paenibacillus sp. TKU042 and Its effect on reducing plasma glucose in a mouse model. Int. J. Mol. Sci. 2017, 18, 700. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L. Production of potent antidiabetic compounds from shrimp head powder via Paenibacillus conversion. Process Biochem. 2019, 76, 18–24. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, S.Y.; Chen, Y.C.; Chen, Y.C.; Yen, Y.H.; Wang, S.L. Enhancement of prodigiosin production by Serratia marcescens TKU011 and its insecticidal activity relative to food colourants. J. Food Sci. 2013, 78, 1743–1751. [Google Scholar] [CrossRef]
- Furstner, A. Chemistry and biology of roseophilin and the prodigiosin alkaloids: A survey of the last 2500 years. Chem. Int. Ed. Engl. 2003, 42, 3582–3603. [Google Scholar] [CrossRef]
- Cerdeno, A.M.; Bibb, M.J.; Challis, G.L. Analysis of the prodiginine biosynthesis gene cluster of Streptomyces coelicolor A3(2): New mechanisms for chain initiation and termination in modular multienzymes. Chem. Biol. 2001, 8, 817–829. [Google Scholar] [CrossRef]
- Samrot, A.V.; Chandana, K.; Senthilkumar, P.; Narendra, K.G. Optimization of prodigiosin production by Serratia marcescens SU-10 and evaluation of its bioactivity. Int. Res. J. Biotechnol. 2011, 2, 128–133. [Google Scholar]
- de Casullo Araújo, H.W.; Fukushima, K.; Campos Takaki, G.M. Prodigiosin production by Serratia marcescens UCP 1549 using renewable-resources as a low-cost substrate. Molecules 2010, 15, 6931–6940. [Google Scholar] [CrossRef] [PubMed]
- Gulani, C.; Bhattacharya, S.; Das, A. Assessment of process parameters influencing the enhanced production of prodigiosin from Serratia marcescens and evaluation of its antimicrobial, antioxidant and dyeing potentials. Malays. J. Microbiol. 2012, 8, 116–122. [Google Scholar]
- Nakamura, K.; Kitamura, K. Process for Preparation of Prodigiosin. U.S. Patent 4,266,028A, 5 May 1981. [Google Scholar]
- Giri, A.V.; Anandkumar, N.; Muthukumaran, G.; Pennathur, G. A novel medium for the enhanced cell growth and production of prodigiosin from Serratia marcescens isolated from soil. BMC Microbiol. 2004, 4, 11–18. [Google Scholar] [CrossRef]
- Shahitha, S.; Poornima, K. Enhanced production of prodigiosin production in Serratia marcescens. J. Appl. Pharm. Sci. 2012, 2, 138–140. [Google Scholar] [CrossRef]
- Wang, S.L.; Kao, D.Y.; Wang, C.L.; Yen, Y.H.; Chern, M.K.; Chen, Y.H. A solvent stable metalloprotease produced by Bacillus sp. TKU004 and its application in the deproteinization of squid pen for beta-chitin preparation. Enzyme Microb. Technol. 2006, 39, 724–731. [Google Scholar] [CrossRef]
- Wei, Y.H.; Chen, W.C. Enhanced production of prodigiosin-like pigment from Serratia marcescens SMdeltaR by medium improvement and oil-supplementation strategies. J. Biosci. Bioeng. 2005, 99, 616–622. [Google Scholar] [CrossRef]
- Wei, Y.H.; Yu, W.J.; Chen, W.C. Enhanced undecylprodigiosin production from Serratia marcescens SS-1 by medium formulation and amino-acid supplementation. J. Biosci. Bioeng. 2005, 100, 466–471. [Google Scholar] [CrossRef]
- Wang, X.; Tao, J.; Wei, D.; Shen, Y.; Tong, W. Development of an adsorption procedure for the direct separation and purification of prodigiosin from culture broth. Biotechnol. Appl. Biochem. 2004, 40, 277–280. [Google Scholar]
- Solé, M.; Rius, N.; Francia, A.; Lorén, J.G. The effect of pH on prodigiosin production by non-proliferating cells of Serratia marcescens. Lett. Appl. Microbiol. 1994, 19, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Jia, X.; Fang, Y.; Chen, L.; Zhang, H.; Lin, R.; Chen, J. Enhanced production of prodigiosin by Serratia marcescens FZSF02 in the form of pigment pellets. Electron. J. Biotechnol. 2019, 40, 58–64. [Google Scholar] [CrossRef]
- Sumathi, C.; MohanaPriya, D.; Swarnalatha, S.; Dinesh, M.G.; Sekaran, G. Production of prodigiosin using tannery fleshing and evaluating its pharmacological effects. SCI World J. 2014, 2014, 290327. [Google Scholar] [CrossRef] [PubMed]
- Holly, S.; Matthew, C.; Lee, E.; George, P.C.S. Phosphate availability regulates biosynthesis of two antibiotics, prodigiosin and carbapenem, in Serratia via both quorum-sensing-dependent and -independent pathways. Mol. Microbiol. 2003, 47, 303–320. [Google Scholar]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Reclamation of marine chitinous materials for chitosanase production via microbial conversion by Paenibacillus macerans. Mar. Drugs 2018, 16, 429. [Google Scholar] [CrossRef]
- Arivizhivendhan, K.V.; Mahesh, M.; Boopathy, R.; Swarnalatha, S.; Regina Mary, R.; Sekaran, G. Antioxidant and antimicrobial activity of bioactive prodigiosin produces from Serratia marcescens using agricultural waste as a substrate. J. Food Sci. Technol. 2018, 55, 2661. [Google Scholar] [CrossRef]
- Yenkejeh, R.A.; Sam, M.R.; Esmaeillou, M. Targeting survivin with prodigiosin isolated from cell wall of Serratia marcescens induces apoptosis in hepatocellular carcinoma cells. Hum. Exp. Toxicol. 2017, 36, 402–411. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Liu, F.; Hou, D.-X.; Xu, J.; Zhao, X.; Yang, F.; Feng, X. Prodigiosin promotes Nrf2 activation to inhibit oxidative stress induced by microcystin-LR in HepG2 cells. Toxins 2019, 11, 403. [Google Scholar] [CrossRef]
- Orlikova, B.; Legrand, N.; Panning, J.; Dicato, M.; Diederich, M. Anti-inflammatory and anticancer drugs from nature. Cancer Res. Treat. 2014, 159, 123–143. [Google Scholar]
- Li, D.; Liu, J.; Wang, X.; Kong, D.; Du, W.; Li, H.; Hse, C.-Y.; Shupe, T.; Zhou, D.; Zhao, K. Biological potential and mechanism of prodigiosin from Serratia marcescens subsp. lawsoniana in human choriocarcinoma and prostate cancer cell lines. Int. J. Mol. Sci. 2018, 19, 3465. [Google Scholar]
- Dalili, D.; Fouladdel, S.; Rastkari, N.; Samadi, N.; Ahmadkhaniha, R.; Ardavan, A.; Azizi, E. Prodigiosin, the red pigment of Serratia marcescens, shows cytotoxic effects and apoptosis induction in HT-29 and T47D cancer cell lines. Nat. Prod. Res. 2011, 26, 2078–2083. [Google Scholar] [PubMed]
- Perez-Tomas, R.; Montaner, B.; Llagostera, E.; Soto-Cerrato, V. The prodigiosins, proapoptotic drugs with anticancer properties. Biochem. Pharmacol. 2003, 66, 1447–1452. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L.; Nguyen, T.H.; Nguyen, T.H.; Trinh, T.H.T.; Nong, T.T.; Nguyen, T.U.; Nguyen, V.N.; Nguyen, A.D. Reclamation of rhizobacteria newly isolated from black pepper plant roots as potential biocontrol agents of root-knot nematodes. Res. Chem. Intermed. 2019. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PG – Producing Strains | C/N Source | |
|---|---|---|
| 0.94% α-Chitin + 0.56% Casein | 1.5% Squid Pens | |
| S. marcescens TKU011 | 335 ± 14.4 a | 243 ± 24.8 b |
| S. marcescens CC17 | 227 ± 2.93 b | 150 ± 5.77 c |
| S. marcescens TNU01 | 329 ± 16.7 a | 240 ± 17.3 b |
| S. marcescens TNU02 | 325 ± 14.4 a | 236 ± 20.8 b |
| No bacteria | - | - |
| PG – Producing Strains | C/N Source | Prodigiosin (mg/100mL) | Reference |
|---|---|---|---|
| S. marcescens TKU011 | 0.94% α-chitin + 0.56% Casein | 335 | This study |
| S. marcescens TKU011 | 1.5% squid pens | 97.8 | [9] |
| S. marcescens TKU011 | 1.5% peanut powder | 116.8 | [9] |
| S. marcescens TKU011 | 1.0% shrimp shells powders | 19 | [9] |
| S. marcescens TKU011 | 1.0% crab shells powders | 11 | [9] |
| S. marcescens TKU011 | 1.0% shrimp heads powders | 3 | [9] |
| S. marcescens TKU011 | 1.5% squid pens | 248 | [28] |
| S. marcescens | 2.0% peanut seed | 387.5 | [26] |
| S. marcescens | 2.0% peanut oil | 289 | [26] |
| S. marcescens | 2.0% sesame seed | 1668 | [26] |
| S. marcescens | 2.0% sesame oil | 100.6 | [26] |
| S. marcescens | 2.0% copra seed | 194 | [26] |
| S. marcescens | 2.0% coconut oil | 142 | [26] |
| S. marcescens SMΔR | Modified Luria-Bertani broth, 6.0% sunflower oil | 79 | [29] |
| S. marcescens SS-1 | 5 g/L yeast extract as sole N/C source | 69 | [30] |
| S. marcescens Nima | 2% tryptone/glycerol (1/1) | 12.5 | [31] |
| S. marcescens Nima | 100 mM 3-[N-morpholino]-ethanesulphonic acid | 47.5 | [32] |
| S. marcescens FZSF02 | 6.97 g/L of peanut powder, 11.29 mL/L of olive oil and 16.02 g/L of beef extract | 1362.2 | [33] |
| S. marcescens FZSF02 | 1% Soya peptone | 117.4 | [33] |
| S. marcescens FZSF02 | 1% Tryptone | 35.3 | [33] |
| S. marcescens FZSF02 | 1% Yeast extract | 38.02 | [33] |
| S. marcescens FZSF02 | 1% Fish meal | 0 | [33] |
| S. marcescens FZSF02 | 1% Soybean powder | 0 | [33] |
| S. marcescens FZSF02 | 1% Corn steep liquor | 0 | [33] |
| Max Inhibition Against Cancerous Cell Lines (%) | ||||
|---|---|---|---|---|
| MCF-7 | A549 | Hep G2 | WiDr | |
| Crude sample | 91.6 ± 1.76 a,b | 89.2 ± 1.43 b | 93.2 ± 2.12 a,b | 79.4 ± 1.72 c |
| Purified Prodigiosin | 92.5 ± 1.4 a,b | 92.6 ± 1.9 a,b | 93.9 ± 2.0 a | 90.2 ± 1.12 a,b |
| Mitomycin C | 94.1 ± 1.61a | 93.3 ± 1.54 a,b | 91.7 ± 1.01 a,b | 92.6 ± 1.02 a,b |
| Inhibition Against Cancerous Cell Lines (IC50, µg/mL) | ||||
|---|---|---|---|---|
| MCF-7 | A549 | Hep G2 | WiDr | |
| Crude sample | 0.44 ± 0.09 c,b | 0.46 ± 0.01 b | 0.38 ± 0.01 c,b | 0.88 ± 0.05 a |
| Purified Prodigiosin | 0.04 ± 0.01 e | 0.06 ± 0.01 e | 0.04 ± 0.01 e | 0.20 ± 0.03 d |
| Mitomycin C | 0.11 ± 0.01 e | 0.10 ± 0.01 e | 0.13 ± 0.01 e | 0.10 ± 0.01 e |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, V.B.; Chen, S.-P.; Nguyen, T.H.; Nguyen, M.T.; Tran, T.T.T.; Doan, C.T.; Tran, T.N.; Nguyen, A.D.; Kuo, Y.-H.; Wang, S.-L. Novel Efficient Bioprocessing of Marine Chitins into Active Anticancer Prodigiosin. Mar. Drugs 2020, 18, 15. https://doi.org/10.3390/md18010015
Nguyen VB, Chen S-P, Nguyen TH, Nguyen MT, Tran TTT, Doan CT, Tran TN, Nguyen AD, Kuo Y-H, Wang S-L. Novel Efficient Bioprocessing of Marine Chitins into Active Anticancer Prodigiosin. Marine Drugs. 2020; 18(1):15. https://doi.org/10.3390/md18010015
Chicago/Turabian StyleNguyen, Van Bon, Shan-Ping Chen, Thi Hanh Nguyen, Minh Trung Nguyen, Thi Thanh Thao Tran, Chien Thang Doan, Thi Ngoc Tran, Anh Dzung Nguyen, Yao-Haur Kuo, and San-Lang Wang. 2020. "Novel Efficient Bioprocessing of Marine Chitins into Active Anticancer Prodigiosin" Marine Drugs 18, no. 1: 15. https://doi.org/10.3390/md18010015
APA StyleNguyen, V. B., Chen, S.-P., Nguyen, T. H., Nguyen, M. T., Tran, T. T. T., Doan, C. T., Tran, T. N., Nguyen, A. D., Kuo, Y.-H., & Wang, S.-L. (2020). Novel Efficient Bioprocessing of Marine Chitins into Active Anticancer Prodigiosin. Marine Drugs, 18(1), 15. https://doi.org/10.3390/md18010015