Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed


Abstract
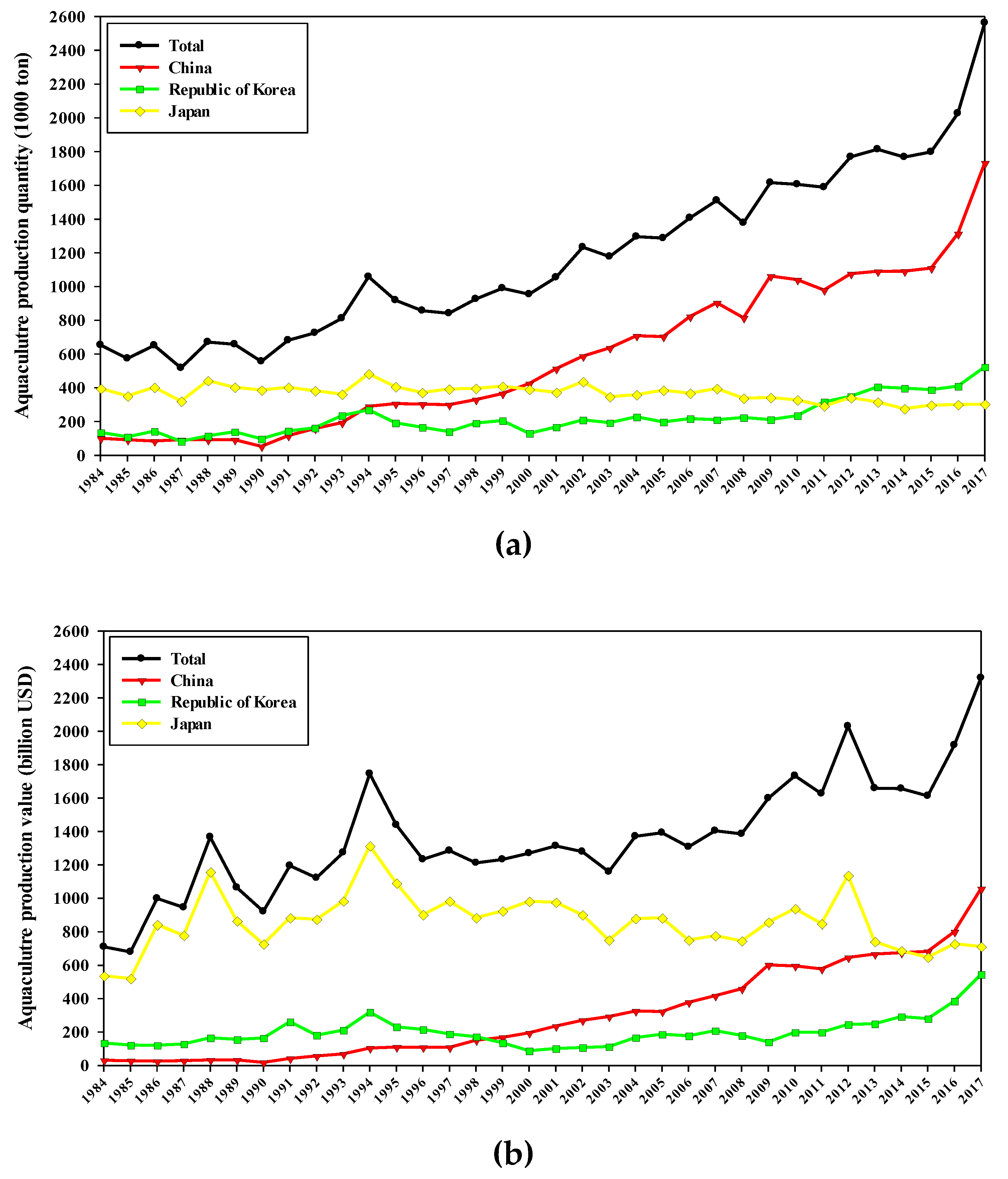
1. Introduction
2. Food Products Containing Lavers
2.1. Raw Lavers
2.2. Processed Laver Products
3. Lavers as Functional Foods: Unique Health Benefits of Laver
4. Future Perspectives on Technical Advances in Laver Utilization
4.1. Management of Current Issues: Control of Potential Risks from Farming to Processing of Lavers
4.1.1. Control of Chemical Risks
4.1.2. Control of Microbiological Risks
4.2. Future Issues: Identifying the Health-Promoting Properties of Laver
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sutherland, J.E.; Lindstrom, S.C.; Nelson, W.A.; Brodie, J.; Lynch, M.D.; Hwang, M.S.; Choi, H.G.; Miyata, M.; Kikuchi, N.; Oliveira, M.C. A new look at an ancient order: Generic revision of the Bangiales (Rhodophyta). J. Phycol. 2011, 47, 1131–1151. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.P.; Uzunov, B.A. An ethnobiological glance on globalization impact on the traditional use of algae and fungi as food in Bulgaria. J. Nutr. Food Sci. 2015, 5, 1. [Google Scholar]
- Pereira, L. A review of the nutrient composition of selected edible seaweeds. Seaweed Ecol. Nutr. Compos. Med. Uses 2011, 7, 15–47. [Google Scholar]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- FAO. Fishery and Aquaculture Statistics. Global Aquaculture Production. In FAO Fisheries and Aquaculture Department; FAO: Rome, Italy, 2019; Available online: http://www.fao.org/fishery/ (accessed on 18 September 2019).
- Levine, I.A.; Sahoo, D. Porphyra: Harvesting Gold from the Sea; IK International Pvt Ltd.: New Delhi, India, 2009. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. Algaebase, World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2019. [Google Scholar]
- Admassu, H.; Abera, T.; Abraha, B.; Yang, R.; Zhao, W. Proximate, mineral and amino acid composition of dried laver (Porphyra spp.) seaweed. J. Acad. Ind. Res. 2018, 6, 149. [Google Scholar]
- Cao, J.; Wang, J.; Wang, S.; Xu, X. Porphyra species: A mini-review of its pharmacological and nutritional properties. J. Med. Food 2016, 19, 111–119. [Google Scholar] [CrossRef]
- Bito, T.; Teng, F.; Watanabe, F. Bioactive compounds of edible purple laver Porphyra sp.(Nori). J. Agric. Food Chem. 2017, 65, 10685–10692. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Cornish, M.L.; Garbary, D.J. Antioxidants from macroalgae: Potential applications in human health and nutrition. Algae 2010, 25, 155–171. [Google Scholar] [CrossRef]
- Kasimala, M.B.; Mebrahtu, L.; Magoha, P.P.; Asgedom, G.; Kasimala, M.B. A review on biochemical composition and nutritional aspects of seaweeds. Carib J. Scitech 2015, 3, 789–797. [Google Scholar]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Abowei, J.F.N.; Ezekiel, E.N. The potentials and utilization of Seaweeds. Scientia 2013, 4, 58–66. [Google Scholar]
- Jesmi, D.; Viji, P.; Rao, B.M. Seaweeds: A promising functional food ingredient. In Training Manual on Value Addition of Seafood; Visakhapatnam Research Centre of ICAR-Central Institute of Fisheries Technology: Visakhapatnam, India, 2018. [Google Scholar]
- Rajapakse, N.; Kim, S.K. Nutritional and digestive health benefits of seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar] [PubMed]
- Madhusudan, C.; Manoj, S.; Rahul, K.; Rishi, C.M. Seaweeds: A diet with nutritional, medicinal and industrial value. Res. J. Med. Plant 2011, 5, 153–157. [Google Scholar] [CrossRef]
- Mok, J.S.; Lee, T.S.; Son, K.T.; Song, K.C.; Kwon, J.Y.; Lee, K.J.; Kim, J.H. Proximate composition and mineral content of laver Porphyra yezoensis from the Korean coast. Korean J. Fish Aquat. Sci. 2011, 44, 554–559. [Google Scholar] [CrossRef]
- Patarra, R.F.; Paiva, L.; Neto, A.I.; Lima, E.; Baptista, J. Nutritional value of selected macroalgae. J. Appl. Phycol. 2011, 23, 205–208. [Google Scholar] [CrossRef]
- Kavale, M.G.; Kazi, M.A.; Bagal, P.U.; Singh, V.; Behera, D.P. Food value of Pyropia vietnamensis (Bangiales, Rhodophyta) from India. Indian J. Geo Mar. Sci. 2018, 47, 402–408. [Google Scholar]
- Cian, R.E.; Fajardo, M.A.; Alaiz, M.; Vioque, J.; González, R.J.; Drago, S.R. Chemical composition, nutritional and antioxidant properties of the red edible seaweed Porphyra columbina. Int. J. Food Sci. Nutr. 2014, 65, 299–305. [Google Scholar] [CrossRef]
- Taboada, M.C.; Millán, R.; Miguez, M.I. Nutritional value of the marine algae wakame (Undaria pinnatifida) and nori (Porphyra purpurea) as food supplements. J. Appl. Phycol. 2013, 25, 1271–1276. [Google Scholar] [CrossRef]
- Taboada, C.; Millan, R.; Miguez, I. Evaluation of marine algae Undaria pinnatifida and Porphyra purpurea as a food supplement: Composition, nutritional value and effect of intake on intestinal, hepatic and renal enzyme activities in rats. J. Sci. Food Agric. 2013, 93, 1863–1868. [Google Scholar] [CrossRef]
- Jung, S.M.; Kang, S.G.; Kim, K.T.; Lee, H.J.; Kim, A.; Shin, H.W. The analysis of proximate composition, minerals and amino acid content of red alga Pyropia dentata by cultivation sites. Korean J. Environ. Ecol. 2015, 29, 1–6. [Google Scholar] [CrossRef]
- Kavale, M.G.; Kazi, M.A.; Sreenadhan, N.; Murgan, P. Nutritional profiling of Pyropia acanthophora var. robusta (Bangiales, Rhodophyta) from Indian waters. J. Appl. Phycol. 2017, 29, 2013–2020. [Google Scholar] [CrossRef]
- Jung, S.M.; Kang, S.G.; Son, J.S.; Jeon, J.H.; Lee, H.J.; Shin, H.W. Temporal and spatial variations in the proximate composition, amino acid, and mineral content of Pyropia yezoensis. J. Appl. Phycol. 2016, 28, 3459–3467. [Google Scholar] [CrossRef]
- CODEX. Regional standard for laver products (CXS 323R-2017). In Codex Alimentarius International Food Standards; CODEX: Montreal, QC, Canada, 2017. [Google Scholar]
- Sanjeewa, K.A.; Lee, W.; Jeon, Y.J. Nutrients and bioactive potentials of edible green and red seaweed in Korea. Fish Aquat. Sci. 2018, 21, 19. [Google Scholar] [CrossRef]
- Oh, S.; Kim, J.; Kim, H.; Son, S.; Choe, E. Composition and antioxidant activity of dried laver, Dolgim. Korean J. Food Sci. Technol. 2013, 45, 403–408. [Google Scholar] [CrossRef]
- Kim, K.W.; Hwang, J.H.; Oh, M.J.; Kim, M.Y.; Choi, M.R.; Park, W.M. Studies on the major nutritional components of commercial dried lavers (Porphyra yezoensis) cultivated in Korea. Korean J. Food Preserv. 2014, 21, 702–709. [Google Scholar] [CrossRef]
- Hwang, E.S.; Ki, K.N.; Chung, H.Y. Proximate composition, amino acid, mineral, and heavy metal content of dried laver. Prev. Nutr. Food Sci. 2013, 18, 139. [Google Scholar] [CrossRef]
- Lee, J.A. Comparative study on the physicochemical character of commercial dried laver in Korea. Culi. Sci. Hos. Res. 2018, 24, 92–99. [Google Scholar]
- Park, W.M.; Kang, D.S.; Bae, T.J. Studies on organic acid, vitamin and free sugar contents of commercial dried lavers (Porphyra yezoensis) cultivated in Korea. J. Korean Soc. Food Sci. Nutr. 2014, 43, 172–177. [Google Scholar] [CrossRef][Green Version]
- Park, W.M.; Kim, K.W.; Kang, D.S.; Bae, T.J. Studies on anion, element, chromaticity and antioxidant activities of commercial dried lavers (Porphyra yezoensis) cultivated in Korea. J. Korean Soc. Food Sci. Nutr. 2014, 43, 323–327. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; Heredia, V.; Pastén, A.; Di Scala, K. An edible red seaweed (Pyropia orbicularis): Influence of vacuum drying on physicochemical composition, bioactive compounds, antioxidant capacity, and pigments. J. Appl. Phycol. 2018, 30, 673–683. [Google Scholar] [CrossRef]
- Yamada; Yamada; Fukuda; Yamada. Bioavailability of dried asakusanori (Porphyra tenera) as a source of cobalamin (vitamin B12). Int. J. Vitam. Nutr. Res. 1999, 69, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, S.; Sugiyama, S.; Ebara, S.; Miyamoto, E.; Abe, K.; Tamura, Y.; Watanabe, F.; Tsuyama, S.; Nakano, Y. Feeding dried purple laver (nori) to vitamin B12-deficient rats significantly improves vitamin B12 status. Br. J. Nutr. 2001, 85, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Han, J.S.; Lee, Y.J.; Yoon, M.R. Changes of chromaticity and mineral contents of laver dishes using various cooking methods. J. East Asian Soc. Diet. Life 2003, 13, 326. [Google Scholar]
- Hwang, E.S. Composition of amino acids, minerals, and heavy metals in differently cooked laver (Porphyra tenera). J. Korean Soc. Food Sci. Nutr. 2013, 42, 1270–1276. [Google Scholar] [CrossRef]
- Miyamoto, E.; Yabuta, Y.; Kwak, C.S.; Enomoto, T.; Watanabe, F. Characterization of vitamin B12 compounds from Korean purple laver (Porphyra sp.) products. J. Agric. Food Chem. 2009, 57, 2793–2796. [Google Scholar] [CrossRef]
- Son, K.T.; Lach, T.; Jung, Y.; Kang, S.K.; Eom, S.H.; Lee, D.S.; Lee, M.S.; Kim, Y.M. Food hazard analysis during dried-laver processing. Fish Aquat. Sci. 2014, 17, 197–201. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosyn. Res. 2015, 123, 227–239. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef]
- Kwon, M.J.; Nam, T.J. Porphyran induces apoptosis related signal pathway in AGS gastric cancer cell lines. Life Sci. 2006, 79, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhou, C.; Yang, H.; Huang, X.; Ma, H.; Qin, X.; Hu, J. Effect of ultrasonic treatment on the degradation and inhibition cancer cell lines of polysaccharides from Porphyra yezoensis. Carbohydr. Polym. 2015, 117, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Noda, H.; Amano, H.; Arashima, K.; Nisizawa, K. Antitumor activity of marine algae. Hydrobiologia 1990, 204, 577–584. [Google Scholar] [CrossRef]
- Yamamoto, I.; Maruyama, H. Effect of dietary seaweed preparations on 1, 2-dimethylhydrazine-induced intestinal carcinogenesis in rats. Cancer Lett. 1985, 26, 241–251. [Google Scholar] [CrossRef]
- Kazłowska, K.; Lin, H.T.V.; Chang, S.; Tsai, G. In vitro and in vivo anticancer effects of sterol fraction from red algae Porphyra dentata. Evid. Based Complement. Alternat. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Ichihara, T.; Wanibuchi, H.; Taniyama, T.; Okai, Y.; Yano, Y.; Otani, S.; Imaoka, S.; Funae, Y.; Fukushima, S. Inhibition of liver glutathione S-transferase placental form-positive foci development in the rat hepatocarcinogenesis by Porphyra tenera (Asakusa-nori). Cancer Lett. 1999, 141, 211–218. [Google Scholar] [CrossRef]
- Park, S.J.; Ryu, J.; Kim, I.H.; Choi, Y.H.; Nam, T.J. Activation of the mTOR signaling pathway in breast cancer MCF-7 cells by a peptide derived from Porphyra yezoensis. Oncol. Rep. 2015, 33, 19–24. [Google Scholar] [CrossRef]
- He, D.; Wu, S.; Yan, L.; Zuo, J.; Cheng, Y.; Wang, H.; Liu, J.; Zhang, X.; Wu, M.; Choi, J.I.; et al. Antitumor bioactivity of porphyran extracted from Pyropia yezoensis Chonsoo2 on human cancer cell lines. J. Sci. Food Agric. 2019, 99, 6722–6730. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Xue, Y.-T. Optimization of microwave assisted extraction, chemical characterization and antitumor activities of polysaccharides from porphyra haitanensis. Carbohydr. Polym. 2019, 206, 179–186. [Google Scholar] [CrossRef]
- Abe, S.; Kaneda, T. Occurrence of gamma-butyrobetaine in a red alga, Porphyra yezoensis. Bull. Jpn. Soc. Sci. Fish. 1973, 39, 239. [Google Scholar] [CrossRef][Green Version]
- Noda, H. Health benefits and nutritional properties of nori. J. Appl. Phycol. 1993, 5, 255–258. [Google Scholar] [CrossRef]
- Tsuge, K.; Okabe, M.; Yoshimura, T.; Sumi, T.; Tachibana, H.; Yamada, K. Dietary effects of porphyran from Porphyra yezoensis on growth and lipid metabolism of Sprague-Dawley rats. Food Sci. Technol. Res. 2007, 10, 147–151. [Google Scholar] [CrossRef]
- Inoue, N.; Yamano, N.; Sakata, K.; Nagao, K.; Hama, Y.; Yanagita, T. The sulfated polysaccharide porphyran reduces apolipoprotein B100 secretion and lipid synthesis in HepG2 cells. Biosci. Biotechnol. Biochem. 2009, 73, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Escrig, A.; Sánchez-Muniz, F. Dietary fibre from edible seaweeds: Chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr. Res. 2000, 20, 585–598. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Yamanaka, Y.; TsujiI, K.; IchikahaE, T.; Nakaguwa, A.; Kawamura, M. Effect of dietary taurine on cholesterol gallstone formation and tissue cholesterol contents in mice. J. Nutr. Sci. Vitaminol. 1985, 31, 225–232. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Wang, J.; Zhang, H.; Niu, X.; Li, P. Preparation of the different derivatives of the low-molecular-weight porphyran from Porphyra haitanensis and their antioxidant activities in vitro. Int. J. Biol. Macromol. 2009, 45, 22–26. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Q.; Qi, H.; Zhang, H.; Niu, X.; Xu, Z.; Li, Z. Degradation of porphyran from Porphyra haitanensis and the antioxidant activities of the degraded porphyrans with different molecular weight. Int. J. Biol. Macromol. 2006, 38, 45–50. [Google Scholar] [CrossRef]
- Shin, E.S.; Hwang, H.J.; Kim, I.H.; Nam, T.J. A glycoprotein from Porphyra yezoensis produces anti-inflammatory effects in liposaccharide-stimulated macrophages via the TLR4 signaling pathway. Int. J. Mol. Med. 2011, 28, 809–815. [Google Scholar]
- Choe, E.; Oh, S. Effects of water activity on the lipid oxidation and antioxidants of dried laver (Porphyra) during storage in the dark. J. Food Sci. 2013, 78, C1144–C1151. [Google Scholar] [CrossRef]
- Gong, G.; Zhao, J.; Wang, C.; Wei, M.; Dang, T.; Deng, Y.; Sun, J.; Song, S.; Huang, L.; Wang, Z. Structural characterization and antioxidant activities of the degradation products from Porphyra haitanensis polysaccharides. Process Biochem. 2018, 74, 185–193. [Google Scholar] [CrossRef]
- Cermeno, M.; Stack, J.; Tobin, P.; O‘Keeffe, M.B.; Harnedy, P.A.; Stengel, D.; FitzGerald, R.J. Peptide identification from a Porphyra dioica protein hydrolysate with antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activities. Food Funct. 2019, 10, 3421–3429. [Google Scholar] [CrossRef]
- Bhatia, S.; Rathee, P.; Sharma, K.; Chaugule, B.; Kar, N.; Bera, T. Immuno-modulation effect of sulphated polysaccharide (porphyran) from Porphyra vietnamensis. Int. J. Biol. Macromol. 2013, 57, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-M.; Xu, S.-S.; Li, L.; Pan, T.-M.; Shi, C.-L.; Liu, H.; Cao, M.-J.; Su, W.-J.; Liu, G.-M. In vitro and in vivo immunomodulatory activity of sulfated polysaccharide from Porphyra haitanensis. Carbohydr. Polym. 2017, 165, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Kim, I.H.; Nam, T.J. Bioactive peptide from Pyropia yezoensis and its anti-inflammatory activities. Int. J. Mol. Med. 2015, 36, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Kwon, M.J.; Kim, I.H.; Kim, Y.M.; Lee, M.K.; Nam, T.J. Pyropia yezoensis glycoprotein promotes the M1 to M2 macrophage phenotypic switch via the STAT3 and STAT6 transcription factors. Int. J. Mol. Med. 2016, 38, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Yanagido, A.; Ueno, M.; Jiang, Z.; Cho, K.; Yamaguchi, K.; Kim, D.; Oda, T. Increase in anti-inflammatory activities of radical-degraded porphyrans isolated from discolored nori (Pyropia yezoensis). Int. J. Biol. Macromol. 2018, 117, 78–86. [Google Scholar] [CrossRef]
- Wang, Y.; Hwang, J.Y.; Park, H.B.; Yadav, D.; Oda, T.; Jin, J.O. Porphyran isolated from Pyropia yezoensis inhibits lipopolysaccharide-induced activation of dendritic cells in mice. Carbohydr. Polym. 2020, 229, 115457. [Google Scholar] [CrossRef]
- Choi, J.W.; Kim, Y.M.; Park, S.J.; Kim, I.H.; Nam, T.J. Protective effect of Porphyra yezoensis glycoprotein on D-galactosamine-induced cytotoxicity in Hepa 1c1c7 cells. Mol. Med. Rep. 2015, 11, 3914–3919. [Google Scholar] [CrossRef]
- Choi, J.W.; Kim, I.H.; Kim, Y.M.; Lee, M.K.; Choi, Y.H.; Nam, T.J. Protective effect of Pyropia yezoensis glycoprotein on chronic ethanol consumption-induced hepatotoxicity in rats. Mol. Med. Rep. 2016, 14, 4881–4886. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Bhuiyan, M.M.H.; Hannan, M.A.; Getachew, P.; Hong, Y.K.; Choi, J.S.; Choi, I.S.; Moon, I.S. The edible red alga Porphyra yezoensis promotes neuronal survival and cytoarchitecture in primary hippocampal neurons. Cell. Mol. Neurobiol. 2016, 36, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Deng, Z.; Geng, L.; Wang, J.; Zhang, Q. In vitro evaluation of the neuroprotective effect of oligo-porphyran from Porphyra yezoensis in PC12 cells. J. Appl. Phycol. 2019, 31, 2559–2571. [Google Scholar] [CrossRef]
- Ueno, M.; Cho, K.; Isaka, S.; Nishiguchi, T.; Yamaguchi, K.; Kim, D.; Oda, T. Inhibitory effect of sulphated polysaccharide porphyran (isolated from Porphyra yezoensis) on RANKL-induced differentiation of RAW264. 7 cells into osteoclasts. Phytother. Res. 2018, 32, 452–458. [Google Scholar] [CrossRef]
- Kitano, Y.; Murazumi, K.; Duan, J.; Kurose, K.; Kobayashi, S.; Sugawara, T.; Hirata, T. Effect of dietary porphyran from the red alga, Porphyra yezoensis, on glucose metabolism in diabetic KK-Ay mice. J. Nutr. Sci. Vitaminol. 2012, 58, 14–19. [Google Scholar] [CrossRef]
- Chao, P.C.; Hsu, C.C.; Liu, W.H. Renal protective effects of Porphyra dentate aqueous extract in diabetic mice. Biomedicine (Taipei) 2014, 4, 18. [Google Scholar] [CrossRef]
- Cao, C.; Chen, M.; Liang, B.; Xu, J.; Ye, T.; Xia, Z. Hypoglycemic effect of abandoned Porphyra haitanensis polysaccharides in alloxan-induced diabetic mice. Bioact. Carbohydr. Diet. Fibre 2016, 8, 1–6. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.; Yang, R.; Zhao, W. Identification of bioactive peptides with α-amylase inhibitory potential from enzymatic protein hydrolysates of red seaweed (Porphyra spp). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef]
- Usov, A.I. Polysaccharides of the red algae. In Advances in Carbohydrate Chemistry and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2011; Volume 65, pp. 115–217. [Google Scholar]
- Ren, D.; Noda, H.; Amano, H.; Nishino, T.; Nishizawa, K. Study on antihypertensive and antihyperlipidemic effects of marine algae. Fish Sci. 1994, 60, 83–88. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Watanabe, F.; Yabuta, Y.; Bito, T.; Teng, F. Vitamin B12-containing plant food sources for vegetarians. Nutrients 2014, 6, 1861–1873. [Google Scholar] [CrossRef]
- Škrovánková, S. Seaweed vitamins as nutraceuticals. Adv. Food Nutr. Res. 2011, 64, 357–369. [Google Scholar] [PubMed]
- Rauma, A.L.; Törrönen, R.; Hänninen, O.; Mykkänen, H. Vitamin B-12 status of long-term adherents of a strict uncooked vegan diet (“living food diet”) is compromised. J. Nutr. 1995, 125, 2511–2515. [Google Scholar] [PubMed]
- van den Berg, H.; Brandsen, L.; Sinkeldam, B.J. Vitamin B-12 content and bioavailability of spirulina and nori in rats. J. Nutr. Biochem. 1991, 2, 314–318. [Google Scholar] [CrossRef]
- Kawasaki, A.; Ono, A.; Mizuta, S.; Kamiya, M.; Takenaga, T.; Murakami, S. The taurine content of Japanese seaweed. In Taurine 10; Springer: New York, NY, USA, 2017; pp. 1105–1112. [Google Scholar]
- Kwak, E.S. Combined Effect of Citric Acid and Drying for the Improvement of Microbiological Quality in Dried Laver. Master’s Thesis, Korea university, Seoul, Korea, 2016. [Google Scholar]
- Kang, S.J.; Yang, S.Y.; Lee, J.W.; Lee, K.W. Polycyclic aromatic hydrocarbons in seasoned-roasted laver and their reduction according to the mixing ratio of seasoning oil and heat treatment in a model system. Food Sci. Biotechnol. 2019, 28, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.I.; Kim, J.G.; Kwon, S.C. A study on physical risk and chemical risk analaysis of seasoned laver. J. Korea Acad. Industr. Coop. Soc. 2017, 18, 620–626. [Google Scholar]
- Bouga, M.; Combet, E. Emergence of seaweed and seaweed-containing foods in the UK: Focus on labeling, iodine content, toxicity and nutrition. Foods 2015, 4, 240–253. [Google Scholar] [CrossRef]
- Yeh, T.S.; Hung, N.H.; Lin, T.C. Analysis of iodine content in seaweed by GC-ECD and estimation of iodine intake. J. Food Drug Anal. 2014, 22, 189–196. [Google Scholar] [CrossRef]
- Jeon, M.J.; Kim, W.G.; Kwon, H.; Kim, M.; Park, S.; Oh, H.-S.; Han, M.; Kim, T.Y.; Shong, Y.K.; Kim, W.B. Excessive iodine intake and thyrotropin reference interval: Data from the Korean National Health and Nutrition Examination Survey. Thyroid 2017, 27, 967–972. [Google Scholar] [CrossRef]
- Bulow Pedersen, I.; Knudsen, N.; Jørgensen, T.; Perrild, H.; Ovesen, L.; Laurberg, P. Large differences in incidences of overt hyper-and hypothyroidism associated with a small difference in iodine intake: A prospective comparative register-based population survey. J. Clin. Endocrinol. 2002, 87, 4462–4469. [Google Scholar] [CrossRef]
- Li, Y.; Teng, D.; Shan, Z.; Teng, X.; Guan, H.; Yu, X.; Fan, C.; Chong, W.; Yang, F.; Dai, H. Antithyroperoxidase and antithyroglobulin antibodies in a five-year follow-up survey of populations with different iodine intakes. J. Clin. Endocrinol. 2008, 93, 1751–1757. [Google Scholar] [CrossRef]
- Gao, Y.; Yue, J. Dietary fiber and human health. In Cereals and Pulses: Nutraceutical Properties and Health Benefits; Wiley: Hoboken, NJ, USA, 2012; pp. 261–271. [Google Scholar]
- Yang, W.H.; Lee, H.J.; Lee, S.Y.; Kim, S.G.; Kim, G.B. Heavy metal contents and food safety assessment of processed seaweeds and cultured lavers. J. Korean Soc. Mar. 2016, 19, 203–210. [Google Scholar]
- Pérez, A.A.; Farías, S.S.; Strobl, A.M.; Pérez, L.B.; López, C.M.; Piñeiro, A.; Roses, O.; Fajardo, M.A. Levels of essential and toxic elements in Porphyra columbina and Ulva sp. from San Jorge Gulf, Patagonia Argentina. Sci. Total Environ. 2007, 376, 51–59. [Google Scholar]
- Hwang, Y.O.; Park, S.G.; Park, G.Y.; Choi, S.M.; Kim, M.Y. Total arsenic, mercury, lead, and cadmium contents in edible dried seaweed in Korea. Food Addit. Contam. B Surveill. 2010, 3, 7–13. [Google Scholar] [CrossRef] [PubMed]
- JECFA Database for the Evaluations of Flavours, Food Additives, Contaminants, Toxicants and Veterinary Drugs. Available online: https://apps.who.int/food-additives-contaminants-jecfa-database/search.aspx (accessed on 16 December 2019).
- Zhan, Z.; Wang, W.; Zhang, G. Risk analysis of aluminium contamination in kelp and laver. J. Food Saf. Food Qual. 2016, 7, 1330–1334. [Google Scholar]
- Zhang, Q.; Ma, J.; Ju, M.; Lu, Q.; Shen, H. Analysis of inorganic arsenic in Pyropia yezoensis and determination of total arsenic in aquaculture areas. Shanghai Hai Yang Da Xue Xue Bao 2014, 23, 518–522. [Google Scholar]
- Zhu, Y.; Shao, B.; Ding, H.; Chen, D.; Shi, Y.; Ji, G.; Fan, B. Contaminations of arsenic, aluminum, lead and cadmium in dried streak laver in Nantong area. J. Food Saf. Food Qual. 2018, 9, 1950–1954. [Google Scholar]
- Wei, C.; Li, W.; Zhang, C.; Van Hulle, M.; Cornelis, R.; Zhang, X. Safety evaluation of organoarsenical species in edible Porphyra from the China Sea. J. Agric. Food Chem. 2003, 51, 5176–5182. [Google Scholar] [CrossRef]
- Taylor, V.F.; Li, Z.; Sayarath, V.; Palys, T.J.; Morse, K.R.; Scholz-Bright, R.A.; Karagas, M.R. Distinct arsenic metabolites following seaweed consumption in humans. Sci. Rep. 2017, 7, 3920. [Google Scholar] [CrossRef]
- Smith, J.L.; Summers, G.; Wong, R. Nutrient and heavy metal content of edible seaweeds in New Zealand. N. Z. J. Crop Hortic. Sci. 2010, 38, 19–28. [Google Scholar] [CrossRef]
- Khan, N.; Ryu, K.Y.; Choi, J.Y.; Nho, E.Y.; Habte, G.; Choi, H.; Kim, M.H.; Park, K.S.; Kim, K.S. Determination of toxic heavy metals and speciation of arsenic in seaweeds from South Korea. Food Chem. 2015, 169, 464–470. [Google Scholar] [CrossRef]
- Mok, J.S.; Son, K.T.; Lee, T.S.; Lee, K.J.; Jung, Y.J.; Kim, J.H. Removal of hazardous heavy metals (Cd, Cr, and Pb) from laver Pyropia sp. with acid treatment. Korean J. Fish Aquat. Sci. 2016, 49, 556–563. [Google Scholar] [CrossRef][Green Version]
- Oya-Ohta, Y.; Kaise, T.; Ochi, T. Induction of chromosomal aberrations in cultured human fibroblasts by inorganic and organic arsenic compounds and the different roles of glutathione in such induction. Mutat. Res. 1996, 357, 123–129. [Google Scholar] [CrossRef]
- SAC (The Standardization Administration of China), GB 2733-2005. In Hygienic Standard for Fresh and Frozen Marine Products of Animal Origin; SAC: Beijing, China, 2005.
- Harakeh, S.; Yassine, H.; El-Fadel, M. Antimicrobial-resistant patterns of Escherichia coli and Salmonella strains in the aquatic Lebanese environments. Environ. Pollut. 2006, 143, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Ayers, T.; Mahon, B.E.; Swerdlow, D.L. Epidemiology of seafood-associated infections in the United States. Clin. Microbiol. Rev. 2010, 23, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.S.; Kim, N.H.; Kim, H.W.; Kim, S.A.; Jo, J.I.; Kim, S.H.; Lee, S.H.; Ha, S.D.; Rhee, M.S. Microbiological quality of seasoned roasted laver and potential hazard control in a real processing line. J. Food Prot. 2014, 77, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Röhr, A.; Lüddecke, K.; Drusch, S.; Müller, M.J.; Alvensleben, R.V. Food quality and safety––consumer perception and public health concern. Food Control 2005, 16, 649–655. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, J.; Mu, W.; Fu, Z.; Zhang, X. Consumers’ perception toward quality and safety of fishery products, Beijing, China. Food Control 2009, 20, 918–922. [Google Scholar] [CrossRef]
- Kim, N.H.; Yun, A.R.; Rhee, M.S. Prevalence and classification of toxigenic Staphylococcus aureus isolated from refrigerated ready-to-eat foods (sushi, kimbab and California rolls) in Korea. J. Appl. Microbiol. 2011, 111, 1456–1464. [Google Scholar] [CrossRef]
- Chung, H.J.; Lee, N.Y.; Jo, C.; Shin, D.H.; Byun, M.W. Use of gamma irradiation for inactivation of pathogens inoculated into Kimbab, steamed rice rolled by dried laver. Food Control 2007, 18, 108–112. [Google Scholar] [CrossRef]
- Ahn, H.J.; Yook, H.S.; Kim, D.H.; Kim, S.; Byun, M.W. Identification of radiation-resistant bacterium isolated from dried laver (Porphyra tenera). J. Korean Soc. Food Sci. Nutr. 2001, 30, 193–195. [Google Scholar]
- Costello, M.; Rhee, M.S.; Bates, M.P.; Clark, S.; Luedecke, L.O.; Kang, D.H. Eleven-year trends of microbiological quality in bulk tank milk. Food Prot. Trends 2003, 23, 393–400. [Google Scholar]
- Lee, T.S.; Lee, H.J.; Byun, H.S.; Kim, J.H.; Park, M.J.; Park, H.Y.; Jung, K.J. Effect of heat treatment in dried lavers and modified processing. Korean J. Fish Aquat. Sci. 2000, 33, 529–532. [Google Scholar]
- Choi, Y.M.; Park, H.J.; Jang, H.I.; Kim, S.A.; Imm, J.Y.; Hwang, I.G.; Rhee, M.S. Changes in microbial contamination levels of porcine carcasses and fresh pork in slaughterhouses, processing lines, retail outlets, and local markets by commercial distribution. Res. Vet. Sci. 2013, 94, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Noh, B.Y.; Hwang, S.H.; Cho, Y.S. Microbial contamination levels in Porphyra sp. distributed in Korea. Korean J. Fish Aquat. Sci. 2019, 52, 180–184. [Google Scholar]
- Lee, E.J.; Kim, G.R.; Lee, H.J.; Kwon, J.H. Monitoring microbiological contamination, pre-decontamination, and irradiation status of commercial dried laver (Porphyra sp.) products. Korean J. Food Sci. Technol. 2017, 49, 20–27. [Google Scholar] [CrossRef]
- Lee, N.Y.; Jo, C.H.; Chung, H.J.; Kang, H.J.; Kim, J.K.; Kim, H.J.; Byun, M.W. The prediction of the origin of microbial contamination in Kimbab and improvement of microbiological safety by gamma irradiation. Korean J. Food Sci. Technol. 2005, 37, 279–286. [Google Scholar]
- Jang, H.G.; Kim, N.H.; Choi, Y.M.; Rhee, M.S. Microbiological quality and risk factors related to sandwiches served in bakeries, cafés, and sandwich bars in South Korea. J. Food Prot. 2013, 76, 231–238. [Google Scholar] [CrossRef]
- Rhee, M.S.; Kang, D.H. Rapid and simple estimation of microbiological quality of raw milk using chromogenic Limulus amoebocyte lysate endpoint assay. J. Food Prot. 2002, 65, 1447–1451. [Google Scholar] [CrossRef]
- Kim, D.H.; Song, H.P.; Kim, J.K.; Byun, M.W.; Kim, J.O.; Lee, H.J. Determination of microbial contamination in the process of rice roled in dried laver and improvement of shelf-life by gamma irradiation. J. Korean Soc. Food Sci. Nutr. 2003, 32, 991–996. [Google Scholar]
- Kim, J.S. Microflora Analysis and Pathogen Control of Agricultural Products and Ready-to-Eat Foods. Master’s Thesis, Kyungwon University, Busan, Korea, 2008. [Google Scholar]
- Kang, M.J.; Lee, H.T.; Kim, J.Y. Hazard analysis, determination of critical control points, and establishment of critical limits for seasoned laver. Culi. Sci. Hos. Res. 2015, 21, 1–10. [Google Scholar]
- Kim, K.Y.; Yoon, S.Y. A study on microbiological risk assessment for the HACCP system construction of seasoned laver. Korean J. Environ. Health Sci. 2013, 39, 268–278. [Google Scholar] [CrossRef]
- Park, S.Y.; Song, H.H.; Ha, S.D. Synergistic effects of NaOCl and ultrasound combination on the reduction of Escherichia coli and Bacillus cereus in raw laver. Foodborne Pathog. Dis. 2014, 11, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Puligundla, P.; Kim, J.W.; Mok, C. Effect of low-pressure air plasma on the microbial load and physicochemical characteristics of dried laver. LWT Food Sci. Technol. 2015, 63, 966–971. [Google Scholar] [CrossRef]
- Kim, Y.J.; Oh, H.S.; Kim, M.J.; Kim, J.H.; Goh, J.B.; Choi, I.Y.; Park, M.K. Identification of electron beam-resistant bacteria in the microbial reduction of dried laver (Porphyra tenera) subjected to electron beam treatment. Korean J. Food Preserv. 2016, 23, 139–143. [Google Scholar] [CrossRef]
- Lee, E.J.; Ameer, K.; Kim, G.R.; Chung, M.S.; Kwon, J.H. Effects of approved dose of e-beam irradiation on microbiological and physicochemical qualities of dried laver products and detection of their irradiation status. Food Sci. Biotechnol. 2018, 27, 233–240. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, G.R.; Ameer, K.; Kyung, H.K.; Kwon, J.H. Application of electron beam irradiation for improving the microbial quality of processed laver products and luminescence detection of irradiated lavers. Appl. Biol. Chem. 2018, 61, 79–89. [Google Scholar] [CrossRef]
- Lee, E.J.; Ameer, K.; Jo, Y.; Kim, G.R.; Kyung, H.K.; Kwon, J.H. Effects of heat-assisted irradiation treatment on microbial and physicochemical qualities of dried laver (Porphyra spp.) and optimization by response surface methodology. Aquac. Res. 2019, 50, 464–473. [Google Scholar] [CrossRef]
- Zichightech. Apparatus for Sterilization of Watery. Laver. Patent 1,020,180,018,019, 21 February 2018.
- Shirako, K.K. Apparatus for Producing Dried Laver and Dried. Laver. Patent 2,012,034,664, 23 February 2012. [Google Scholar]
- Gachon University. Food Surface Sterilization Method Using Non-Thermal. Plasma. Patent 1,020,140,002,357, 8 January 2014. [Google Scholar]
- Koasa Corporation. Method for Producing Roasted. Nori. Patent WO/2017/077738 (PCT/JP2016/070306), 11 May 2017.
- Reith, M. Complete uncleotide sequence of the Porphyra purpurea chloroplast genome. Plant Mol. Biol. Rep. 1995, 13, 327–332. [Google Scholar] [CrossRef]
- Burger, G.; Saint-Louis, D.; Gray, M.W.; Lang, B.F. Complete sequence of the mitochondrial DNA of the red alga Porphyra purpurea: Cyanobacterial introns and shared ancestry of red and green algae. Plant Cell 1999, 11, 1675–1694. [Google Scholar] [CrossRef]
- Hughey, J.R.; Gabrielson, P.W.; Rohmer, L.; Tortolani, J.; Silva, M.; Miller, K.A.; Young, J.D.; Martell, C.; Ruediger, E. Minimally destructive sampling of type specimens of Pyropia (Bangiales, Rhodophyta) recovers complete plastid and mitochondrial genomes. Sci. Rep. 2014, 4, 5113. [Google Scholar] [CrossRef]
- Brawley, S.H.; Blouin, N.A.; Ficko-Blean, E.; Wheeler, G.L.; Lohr, M.; Goodson, H.V.; Jenkins, J.W.; Blaby-Haas, C.E.; Helliwell, K.E.; Chan, C.X. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. USA 2017, 114, E6361–E6370. [Google Scholar] [CrossRef]
- Harden, L.K.; Morales, K.M.; Hughey, J.R. Identification of a new marine algal species Pyropia nitida sp. nov.(Bangiales: Rhodophyta) from Monterey, California. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 3058–3062. [Google Scholar] [PubMed]
- Li, X.; Xu, J.; He, Y.; Shen, S.; Zhu, J.; Shen, Z. The complete nuclear ribosomal DNA (nrDNA) cistron sequence of Pyropia yezoensis (Bangiales, Rhodophyta). J. Appl. Phycol. 2016, 28, 663–669. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, B.; Kong, F.; Wang, L. The complete mitochondrial genome of Pyropia haitanensis Chang et Zheng. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2012, 23, 344–346. [Google Scholar] [CrossRef]
- Kong, F.; Sun, P.; Cao, M.; Wang, L.; Mao, Y. Complete mitochondrial genome of Pyropia yezoensis: Reasserting the revision of genus Porphyra. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2014, 25, 335–336. [Google Scholar] [CrossRef]
- Hwang, M.S.; Kim, S.O.; Ha, D.S.; Lee, J.E.; Lee, S.R. Complete mitochondrial genome sequence of Pyropia yezoensis (Bangiales, Rhodophyta) from Korea. Plant Biotechnol. Rep. 2014, 8, 221–227. [Google Scholar] [CrossRef]
- Wang, L.; Mao, Y.; Kong, F.; Li, G.; Ma, F.; Zhang, B.; Sun, P.; Bi, G.; Zhang, F.; Xue, H. Complete sequence and analysis of plastid genomes of two economically important red algae: Pyropia haitanensis and Pyropia yezoensis. PLoS ONE 2013, 8, e65902. [Google Scholar] [CrossRef]
- Nakamura, Y.; Sasaki, N.; Kobayashi, M.; Ojima, N.; Yasuike, M.; Shigenobu, Y.; Satomi, M.; Fukuma, Y.; Shiwaku, K.; Tsujimoto, A. The first symbiont-free genome sequence of marine red alga, Susabi-nori (Pyropia yezoensis). PLoS ONE 2013, 8, e57122. [Google Scholar] [CrossRef]
- Miranda, L.N.; Hutchison, K.; Grossman, A.R.; Brawley, S.H. Diversity and abundance of the bacterial community of the red macroalga Porphyra umbilicalis: Did bacterial farmers produce macroalgae? PLoS ONE 2013, 8, e58269. [Google Scholar] [CrossRef]
- Li, C.; Kong, F.; Sun, P.; Bi, G.; Li, N.; Mao, Y.; Sun, M. Genome-wide identification and expression pattern analysis under abiotic stress of mitogen-activated protein kinase genes in Pyropia yezoensis. J. Appl. Phycol. 2018, 30, 2561–2572. [Google Scholar] [CrossRef]
- Li, B.; Chen, C.; Xu, Y.; Ji, D.; Xie, C. Validation of housekeeping genes as internal controls for studying the gene expression in Pyropia haitanensis (Bangiales, Rhodophyta) by quantitative real-time PCR. Acta Oceanol. Sin. 2014, 33, 152–159. [Google Scholar] [CrossRef]
- Mikami, K.; Li, C.; Irie, R.; Hama, Y. A unique life cycle transition in the red seaweed Pyropia yezoensis depends on apospory. Commun. Biol. 2019, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.X.; Blouin, N.A.; Zhuang, Y.; Zäuner, S.; Prochnik, S.E.; Lindquist, E.; Lin, S.; Benning, C.; Lohr, M.; Yarish, C. Porphyra (Bangiophyceae) transcriptomes provide insights into red algal development and metabolism. J. Phycol. 2012, 48, 1328–1342. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Li, B.; Xu, Y.; Ji, D.; Chen, C. Characterization of the global transcriptome for Pyropia haitanensis (Bangiales, Rhodophyta) and development of cSSR markers. BMC Genom. 2013, 14, 107. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, R.; Sekine, N.; Mizuta, H.; Uji, T. Transcriptomic analysis under ethylene precursor treatment uncovers the regulation of gene expression linked to sexual reproduction in the dioecious red alga Pyropia pseudolinearis. J. Appl. Phycol. 2019, 31, 3317–3329. [Google Scholar] [CrossRef]
- Gao, D.; Kong, F.; Sun, P.; Bi, G.; Mao, Y. Transcriptome-wide identification of optimal reference genes for expression analysis of Pyropia yezoensis responses to abiotic stress. BMC Genom. 2018, 19, 251. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chang, J.; Zheng, H.; Ji, D.; Xu, Y.; Chen, C.; Xie, C. Full-length transcriptome sequences obtained by a combination of sequencing platforms applied to heat shock proteins and polyunsaturated fatty acids biosynthesis in Pyropia haitanensis. J. Appl. Phycol. 2019, 31, 1483–1492. [Google Scholar] [CrossRef]
- Choi, S.; Hwang, M.S.; Im, S.; Kim, N.; Jeong, W.J.; Park, E.J.; Gong, Y.G.; Choi, D.W. Transcriptome sequencing and comparative analysis of the gametophyte thalli of Pyropia tenera under normal and high temperature conditions. J. Appl. Phycol. 2013, 25, 1237–1246. [Google Scholar] [CrossRef]
- Sun, P.; Mao, Y.; Li, G.; Cao, M.; Kong, F.; Wang, L.; Bi, G. Comparative transcriptome profiling of Pyropia yezoensis (Ueda) MS Hwang & HG Choi in response to temperature stresses. BMC Genom. 2015, 16, 463. [Google Scholar]
- Wang, W.; Teng, F.; Lin, Y.; Ji, D.; Xu, Y.; Chen, C.; Xie, C. Transcriptomic study to understand thermal adaptation in a high temperature-tolerant strain of Pyropia haitanensis. PLoS ONE 2018, 13, e0195842. [Google Scholar] [CrossRef]
- Contreras-Porcia, L.; López-Cristoffanini, C.; Lovazzano, C.; Flores-Molina, M.R.; Thomas, D.; Núñez, A.; Fierro, C.; Guajardo, E.; Correa, J.A.; Kube, M. Expresión diferencial de genes en Pyropia columbina (Bangiales, Rhodophyta) bajo hidratación y desecación natural. Lat. Am. J. Aquat. Res. 2013, 41, 933–958. [Google Scholar] [CrossRef]
- Im, S.; Jung, H.S.; Lee, H.N.; Park, E.J.; Hwang, M.S.; Jeong, W.J.; Choi, D.W. Transcriptome-based identification of the desiccation response genes in the marine red alga Pyropia tenera (Rhodophyta) and enhancement of abiotic stress tolerance by PtDRG2 in chlamydomonas. Phycologia 2017, 56, 83. [Google Scholar]
- Wang, W.; Xu, Y.; Chen, T.; Xing, L.; Xu, K.; Ji, D.; Chen, C.; Xie, C. Regulatory mechanisms underlying the maintenance of homeostasis in Pyropia haitanensis under hypersaline stress conditions. Sci. Total Environ. 2019, 662, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.; Kumar, V.; Nappi, J.; Gardiner, M. Microbial diversity and symbiotic interactions with macroalgae. In Algal and Cyanobacteria Symbioses; World Scientific: Singapore, 2017; pp. 493–546. [Google Scholar]
- Quigley, C.T.; Morrison, H.G.; Mendonça, I.R.; Brawley, S.H. A common garden experiment with Porphyra umbilicalis (Rhodophyta) evaluates methods to study spatial differences in the macroalgal microbiome. J. Phycol. 2018, 54, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Chemical interactions between marine macroalgae and bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–299. [Google Scholar] [CrossRef]
- Harley, C.D.; Anderson, K.M.; Demes, K.W.; Jorve, J.P.; Kordas, R.L.; Coyle, T.A.; Graham, M.H. Effects of climate change on global seaweed communities. J. Phycol. 2012, 48, 1064–1078. [Google Scholar] [CrossRef]
- Guan, X.; Zhou, W.; Hu, C.; Zhu, M.; Ding, Y.; Gai, S.; Zheng, X.; Zhu, J.; Lu, Q. Bacterial community temporal dynamics and disease-related variations in the seawater of Pyropia (laver) seedling pools. J. Appl. Phycol. 2018, 30, 1217–1224. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, C.; Ji, D.; Hang, N.; Xie, C. Proteomic profile analysis of Pyropia haitanensis in response to high-temperature stress. J. Appl. Phycol. 2014, 26, 607–618. [Google Scholar] [CrossRef]
- Shi, J.; Chen, Y.; Xu, Y.; Ji, D.; Chen, C.; Xie, C. Differential proteomic analysis by iTRAQ reveals the mechanism of Pyropia haitanensis responding to high temperature stress. Sci. Rep. 2017, 7, 44734. [Google Scholar] [CrossRef]
- López-Cristoffanini, C.; Zapata, J.; Gaillard, F.; Potin, P.; Correa, J.A.; Contreras-Porcia, L. Identification of proteins involved in desiccation tolerance in the red seaweed Pyropia orbicularis (Rhodophyta, Bangiales). Proteomics 2015, 15, 3954–3968. [Google Scholar] [CrossRef]
- Xu, K.; Xu, Y.; Ji, D.; Xie, J.; Chen, C.; Xie, C. Proteomic analysis of the economic seaweed Pyropia haitanensis in response to desiccation. Algal Res. 2016, 19, 198–206. [Google Scholar] [CrossRef]
- Khan, S.; Mao, Y.; Gao, D.; Riaz, S.; Niaz, Z.; Tang, L.; Khan, S.; Wang, D. Identification of proteins responding to pathogen-infection in the red alga Pyropia yezoensis using iTRAQ quantitative proteomics. BMC Genom. 2018, 19, 842. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Choi, J.I. Identification, characterization, and proteomic studies of an aldehyde dehydognease (ALDH) from Pyropia yezoensis (Bangiales, Rhodophyta). J. Appl. Phycol. 2018, 30, 2117–2127. [Google Scholar] [CrossRef]
- Lee, H.J.; Choi, J.I. Isolation and characterization of a high-growth-rate strain in Pyropia yezoensis induced by ethyl methane sulfonate. J. Appl. Phycol. 2018, 30, 2513–2522. [Google Scholar] [CrossRef]
- Lee, H.J.; Choi, J. Proteomic analysis of Pyropia yezoensis mutant induced by gamma irradiation. Phycologia 2017, 56, 113. [Google Scholar]
- Kumari, P. Seaweed lipidomics in the era of ‘omics’ biology: A contemporary perspective. In Systems Biology of Marine Ecosystems; Springer: New York, NY, USA, 2017; pp. 49–97. [Google Scholar]
- Chen, J.; Li, M.; Yang, R.; Luo, Q.; Xu, J.; Ye, Y.; Yan, X. Profiling lipidome changes of Pyropia haitanensis in short-term response to high-temperature stress. J. Appl. Phycol. 2016, 28, 1903–1913. [Google Scholar] [CrossRef]
- da Costa, E.; Azevedo, V.; Melo, T.; Rego, A.; V Evtuguin, D.; Domingues, P.; Calado, R.; Pereira, R.; Abreu, M.; Domingues, M. High-resolution lipidomics of the early life stages of the red seaweed Porphyra dioica. Molecules 2018, 23, 187. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Chen, D.; Lou, Y.; Ye, Y.; Yang, R. Metabolomic profile characteristics of Pyropia haitanensis as affected by harvest time. Food Sci. Technol. Res. 2016, 22, 529–536. [Google Scholar] [CrossRef]
- Ye, Y.; Yang, R.; Lou, Y.; Chen, J.; Yan, X.; Tang, H. Effects of food processing on the nutrient composition of Pyropia yezoensis products revealed by NMR-based metabolomic analysis. J. Food Nutr. Reslov.Iissn 2014, 2, 749–756. [Google Scholar] [CrossRef]
- Hamid, S.S.; Wakayama, M.; Ichihara, K.; Sakurai, K.; Ashino, Y.; Kadowaki, R.; Soga, T.; Tomita, M. Metabolome profiling of various seaweed species discriminates between brown, red, and green algae. Planta 2019, 249, 1921–1947. [Google Scholar] [CrossRef]
- Mao, Y.; Chen, N.; Cao, M.; Chen, R.; Guan, X.; Wang, D. Functional characterization and evolutionary analysis of glycine-betaine biosynthesis pathway in red seaweed Pyropia yezoensis. Mar. Drugs 2019, 17, 70. [Google Scholar] [CrossRef]
- Yan, Y.; Yang, H.; Tang, L.; Li, J.; Mao, Y.; Mo, Z. Compositional shifts of bacterial communities associated with Pyropia yezoensis and surrounding seawater co-occurring with red rot disease. Front. Microbiol. 2019, 10, 1666. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Park, E.J.; Choi, J.I. Isolation, morphological characteristics and proteomic profile analysis of thermo-tolerant Pyropia yezoensis mutant in response to high-temperature stress. Ocean Sci. J. 2019, 54, 65–78. [Google Scholar] [CrossRef]


{kind=link}
| Genus | Species 1 |
|---|---|
| Porphyra | P. acanthophora, P. columbina, P. dentata, P. dioica, P. fucicola, P. haitanensis, P. kanakaensis, P. perforata, P. pseudolinearis, P. purpurea, P. sanjuanensis P. seriata, P. tenera, P. umbilicalis, P. vietnamensis, P. yezoensis |
| Pyropia2 | P. acanthophora, P. columbina, P. dentata, P. fucicola, P. haitanensis, P. kanakaensis, P. nitida, P. orbicularis, P. perforata, P. pseudolinearis P. seriata, P. tenera, P. vietnamensis, P. yezoensis |
| Product Type | Raw Material (Species) | Nutritional Values from Proximate Analysis (w/w %) | Other Nutritional Substances | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|
| Carbohydrate | Dietary Fiber | Protein | Lipid | Ash | Moisture | ||||
| Raw wet laver | P. yezoensis | 1.2–2.7 | - 1 | 3.0–5.0 | 0.5 | 3.6–4.3 | 89.2–90.5 | mineral | [19] |
| Porphyra sp. | - | 43.1, 38.9 | 25.6, 26.0 | - | - | - | - | [20] | |
| Raw laver (dry weight) 2 | P. vietnamensis | 38.8–60.4 | - | 12.4–20.5 | 0.2–2.7 | 3.9–7.4 | 13.6–20.7 | fatty acids | [21] |
| P. dentata | 45.7–45.9 | - | 36.2–37.7 | 0.7–1.0 | 7.1–8.2 | 8.6–8.8 | mineral, amino acids | [25] | |
| P. purpurea | 21.7 3 | 22.9 | 33.2 | 1.0 | 21.3 | - | amino acids, fatty acids, sterol | [24] | |
| P. columbina | - | 48.0 | 24.6 | 0.3 | 6.5 | 12.8 | mineral, amino acids, fatty acids, antioxidants, phenolic compounds | [22] | |
| P. yezoensis | 51.2–57.9 | - | 36.2–39.2 | 2.3–3.1 | 3.8–7.3 | - | mineral, amino acids | [27] | |
| P. acanthophora var. robusta | 35.5–61.0 | - | 14.1–18.4 | 1.7–2.6 | 4.2–6.8 | 12.5–21.5 | mineral, fatty acids, pigments, vitamin | [26] | |
| P. purpurea | - | - | - | - | - | - | mineral, vitamin | [23] | |
| Category | Product Type 1 (Species) 2 | Nutritional Values from Proximate Analysis (w/w %) | Other Nutritional Substances | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|
| Carbohydrate | Dietary Fiber | Protein | Lipid | Ash | Moisture | ||||
| Processed laver products | DL (Porphyra spp.) | 36.8 | 31.6 | 43.0 | 0.5 | 10.3 | 9.4 | mineral, amino acids | [8] |
| DL (P. dentata, P. seriata) | 47.6 | 40.4 | 37.3 | 0.3 | 7.6 | 7.3 | fatty acids, pigments, antioxidants | [30] | |
| DL | 43.8–46.2 | - 3 | 37.8–40.0 | 1.5–2.3 | 8.0–9.0 | 5.7–7.4 | mineral, amino acids, fatty acids, component sugar | [31] | |
| DL (P. tenera) | - | - | 36.9 | 2.3 | 9.1 | 3.7 | mineral, amino acids | [32] | |
| DL (P. haitanensis) | - | - | 32.16 | 1.96 | 8.78 | 6.74 | - | ||
| DL (P. yezoensis) | 45.4–50.0 | - | 29.3–35.0 | 1.8–2.0 | 8.1–9.9 | 8.2–9.8 | mineral | [19] | |
| DL | - | - | - | - | - | 8.4 | phenolic compounds | [33] | |
| DL | - | - | - | - | - | 7.6 | - | ||
| RL | - | - | - | - | - | 8.7 | - | ||
| DL | - | - | - | - | - | 8.7 | - | ||
| DL (P. yezoensis) | - | - | - | - | - | - | vitamin, organic acid, free sugar | [34] | |
| DL | - | - | - | - | - | - | phenolic compounds, anion, element | [35] | |
| DL (P. tenera, P. yezoensis + P. dentata, P. seriata) | 41.7 | 33.4 | 38.4 | 0.3 | 8.0 | 11.6 | - | [30] | |
| Processed laver products with other seaweeds as optional ingredients | DL combined with green laver (Ulva spp.) 4 | 43.7 | 36.6 | 35.0 | 0.8 | 9.1 | 11.4 | - | [30] |
| Health Functionality | Major Components Linked to Health Functionalities | References |
|---|---|---|
| Anti-cancer | polysaccharides (dietary fiber, porphyran), phospholipids, sterol, peptide | [17,46,47,48,49,50,51,52,53,54] |
| Prevention of cardiovascular disease (e.g., hypertension, atherosclerosis, ischemia) | betaine, dietary fiber, taurine, porphyran | [17,55,56,57,58,59,60,61] |
| Antioxidant effect (e.g., Anti-ageing) | porphyran, glycoprotein, polyphenols, tocopherols, peptide | [62,63,64,65,66,67] |
| Anti-inflammatory effect and immunomodulation | glycoprotein, porphyran | [64,68,69,70,71,72,73] |
| Alcohol metabolism | glycoprotein | [74,75] |
| Prevention of nervous diseases (e.g., Alzheimer’s diseases, methylmalonic acidemias) | taurine, porphyran | [38,76,77] |
| Prevention of bone disease (e.g., osteoporosis, rheumatoid arthritis) | porphyran, glycoprotein | [64,78] |
| Anti-diabetes mellitus | phenolic compounds (carotenoids, anthocyanins), polysaccharides (porphyran), peptide | [79,80,81,82] |
| Category | Product Type 1 | Target | Results (mg/kg or μg /g of dw 2) | References |
|---|---|---|---|---|
| Raw laver | - | arsenic (As) | 9.59–34.0 | [100] |
| 22.9–33.8 | [101] | |||
| 0.22–0.70 | [105] | |||
| 12.87 | [109] | |||
| cadmium (Cd) | 0.40–1.21 | [100] | ||
| 2.83–3.54 | [101] | |||
| chromium (Cr) | 0.32–0.86 | [100] | ||
| copper (Cu) | 7.92–16.9 | [100] | ||
| 1.94–6.94 | [101] | |||
| iron (Fe) | 290–723 | [100] | ||
| lead (Pb) | 0.78–1.30 | [100] | ||
| <LOD 3 | [101] | |||
| 0.98 | [109] | |||
| mercury (Hg) | 0.005–0.006 | [100] | ||
| 0.03 | [109] | |||
| nickel (Ni) | 0.69–1.04 | [100] | ||
| 0.74–1.51 | [101] | |||
| zinc (Zn) | 18.0–57.7 | [100] | ||
| 21.1–70.1 | [101] | |||
| Processed laver products | DL | aluminum (Al) | 388.6–623.4 | [104] |
| 66–511 | [106] | |||
| arsenic (As) | 13.5–32.8 | [100] | ||
| <LOD–29.850 | [102] | |||
| ND 4–0.303 | [106] | |||
| 30.18–39.05 | [42] | |||
| cadmium (Cd) | 0.69–4.73 | [100] | ||
| 0.108–3.11 | [106] | |||
| 0.076–0.318 | [42] | |||
| 0.501–2.421 | [102] | |||
| chromium (Cr) | 0.46–0.66 | [100] | ||
| copper (Cu) | 5.02–8.64 | [100] | ||
| iron (Fe) | 103–214 | [100] | ||
| lead (Pb) | ND–0.86 | [100] | ||
| ND | [42] | |||
| <LOD–2.362 | [102] | |||
| ND–0.208 | [106] | |||
| mercury (Hg) | 0.004–0.008 | [100] | ||
| 0.005–0.009 | [42] | |||
| 0.002–0.050 | [102] | |||
| nickel (Ni) | 0.17–1.49 | [100] | ||
| zinc (Zn) | 27.1–57.7 | [100] | ||
| DL, RL | arsenic (As) | 2.1–21.6 | [107] | |
| - 5 | aluminum (Al) | 15.50 6 | [110] | |
| arsenic (As) | 2.07 6 | [110] | ||
| cadmium (Cd) | 0.109 6 | [110] | ||
| lead (Pb) | 0.063 6 | [110] | ||
| mercury (Hg) | <LOD | [110] |
| Category | Target Microorganisms | Product Type 1 | Results | References 2 |
|---|---|---|---|---|
| Processed laver products | Mesophilic bacteria | Standard | 4.48 log CFU/g | [113] |
| DL | 6.5 log CFU/g | [121] | ||
| DL | 7.6 log CFU/g | [123] | ||
| RL | 7.5 log CFU/g | |||
| DL | 6.9 log CFU/g | [125] | ||
| RL | 3.4 log CFU/g | |||
| SL | 4.9 log CFU/g | |||
| DL | 5.6–7.2 log CFU/g | [126] | ||
| RL | 3.6 log CFU/g | |||
| SL | 4.3–6.0 log CFU/g | |||
| Coliforms | Standard | 30 MPN 3/100 g | [113] | |
| DL | 3.2 MPN/ 100 g | [123] | ||
| RL | 3.7 MPN/ 100 g | |||
| DL | 2.1 log CFU/g | [125] | ||
| RL | 1.6 log CFU/g | |||
| SL | 1.0 log CFU/g | |||
| DL | 1.9–2.2 log CFU/g | [126] | ||
| Yeast/mold | Standard | 2.48 log CFU/g | [113] | |
| DL | 4.3–4.9 log CFU/g | [126] | ||
| RL | 2.1 log CFU/g | |||
| SL | 2.1–4.7 log CFU/g | |||
| Bacillus cereus | DL | 2.3 log CFU/g | [126] | |
| Raw materials of food products using laver | Mesophilic bacteria | Standard | 4.48 log CFU/g | [113] |
| DL | 8.8 log CFU/g | [127] | ||
| DL | ca. 7.0 log CFU/g | [130] | ||
| RL | ca. 6.0 log CFU/g | |||
| DL | 5.3 log CFU/g | [131] | ||
| Coliforms | Standard | 30 MPN/100 g | [113] | |
| DL | ca. 3.0 log CFU/g | [130] | ||
| RL | ca.2.0 log CFU/g | |||
| DL | detection rate 6% | [131] | ||
| B. cereus | DL | detection rate 12% | [131] | |
| Clostridium perfringens | DL | detection rate 3% | [131] | |
| Work-in-process and end-products from manufacturing plants | Mesophilic bacteria | Standard | 4.48 log CFU/g | [113] |
| DL | 5.6–8.0 log CFU/g | [42] | ||
| DL | 4.4–7.8 log CFU/g | [116] | ||
| SL | 1.3–5.9 log CFU/g | |||
| DL | 4.7–4.8 log CFU/g | [132] | ||
| SL | ND 4–1.0 log CFU/g | |||
| DL | 3.4–3.6 log CFU/g | [133] | ||
| SL | 1.4–2.8 log CFU/g | |||
| Coliforms | Standard | 30 MPN/100 g | [113] | |
| DL | 54–27,600 MPN/100 g | [42] |
| Target Product | Treatment Methods | Target Microorganisms | Treatment Conditions | Microbial Reduction (log CFU/g) | References |
|---|---|---|---|---|---|
| Raw harvested laver | NaOCl + ultrasound | Escherichia coli Bacillus cereus | 200 ppm, 60 min | 2.6 3.2 | [134] |
| Kimbab | Gamma irradiation | Mesophilic bacteria | 1–3 kGy, 24 h | 1.0–2.0 | [130] |
| Kimbab | Gamma irradiation | Escherichia coli Salmonella Typhimurium Staphylococcus aureus Listeria monocytogenes | 1–3 kGy, 24 h | 1.3–ND 1 2.3–ND 3.4–ND 2.7–ND | [120] |
| DL 2 | UV | Mesophilic bacteria | 20 W, 20 min | 1.0 | [123] |
| DL | Gamma irradiation | Escherichia coli Salmonella Typhimurium Staphylococcus aureus Listeria ivanoviis | 1–3 kGy, 24 h | 2.7–ND 1.7–ND 2.0–ND 1.6–ND | [127] |
| DL | Corona discharge plasma | Mesophilic bacteria | 3312 rpm, 58Hz, 20 min | 2.0 | [135] |
| DL | Low–pressure air plasma | Mesophilic bacteria | 20 min | 1.5–2.0 | |
| DL | e-beam | Mesophilic bacteria | 4 kGy | 1.4 | [136] |
| DL | Heat-assisted e-beam irradiation | Mesophilic bacteria | 1.8–3.0 kGy, 154–170 °C, 10–18 s | > 2.0 3 | [139] |
| DL | Heat-assisted e-beam irradiation | Coliform | 4 kGy, dose rate as 2.1 kGy/h | > 1.5 4 | [137] |
| DL | Heat-assisted low-dosee-beam irradiation | Coliform | 1 kGy, dose rate as 2.1 kGy/hg | > 1.4 5 | [138] |
| Omics Technology | Topic | Species | Major Findings | References |
|---|---|---|---|---|
| Genome | Whole genome sequencing and genomic feature | P. umbilicalis | - Genome governing nutritional/functional values linked to the growth and survival strategy of laver under stressful condition of natural habitat (intertidal zone) | [147] |
| P. yezoensis | - First report on the genome sequence of nuclear ribosomal DNA (nrDNA) cistron | [149] | ||
| P. yezoensis | - Genome sequence and annotated functional genes from P. yezoensis - Identification of photosynthesis system and key genes governing color of laver | [154] | ||
| Genome-wide identification of functional genes | P. yezoensis | - Gene structure associated with mitogen-activated protein kinases from P. yezoensis (PyMAPKs) | [156] | |
| Comparative genomics | P. perforata P. sanjuanensis P. fucicola P. kanakaensis | - Reliable analytical method for the genomes of laver by the destructive sampling of type specimen | [146] | |
| P. nitida | - Recognition of new red algal species | [148] | ||
| P. haitanensis | - Supportive data for the phylogenic differences between Pyropia from Porphyra | [150] | ||
| P. yezoensis | - Supportive data for the phylogenic differences between Pyropia from Porphyra | [151] | ||
| P. yezoensis | - Different genomic structure of strains according to the regions of cultivars (Korea and China) | [152] | ||
| P. haitanensis, P. yezoensis | - Biodiversity and distinct phylogenies of laver compared with other red algae | [153] | ||
| Transcriptome | Analytical techniques | P. haitanensis | - Selection of housekeeping gene mostly adequate for the designation of internal control based on the stability under abiotic stresses | [157] |
| Unique life cycle | P. yezoensis | -Transition observed in the life cycle with apospory | [158] | |
| P. umbilicalis, P. purpurea | - Evolutionary analysis for the growth and development of laver | [159] | ||
| P. haitanensis | -Transcriptomic profile under different physiological conditions -Role of cSSR markers linked to the differences in the gene expressions among lifecycle stages of laver | [160] | ||
| P. pseudolinearis | - Impact of ethylene precursor treatment to the regulation of gene expression governing reproduction | [161] | ||
| Stress response | P. yezoensis | - Stress response of PyMAPK gene family | [156] | |
| P. yezoensis | - Identification of key response genes expressed under various abiotic stresses | [162] | ||
| P. haitanensis | - Role of heat shock proteins against the abiotic stresses | [163] | ||
| P. tenera | - Distinct transcriptional characteristics of gametophyte thalli by high-temperature stresses | [164] | ||
| P. yezoensis | - Transcriptomic profiles in response to stresses associated with temperature | [165] | ||
| P. haitanensis | - Identification of key response genes expressed under thermal stresses - Mechanisms on the adaptation of high-temperature tolerant strain | [166] | ||
| P. columbina | - Identification of mechanisms on resistance and key response genes expressed under stresses from desiccation-hydration cycles in natural habitat | [167] | ||
| P. tenera | - Identification of mechanisms on resistance and key response genes expressed under desiccation | [168] | ||
| P. haitanensis | - Identification of mechanisms on resistance and maintenance of homeostasis under stresses from hypersaline conditions | [169] | ||
| Biosynthesis | P. yezoensis | - Role of glycine-betaine (GB) capable of maintenance of osmotic balance in response to desiccation stresses - Identification of major enzymes involved in the biosynthesis of GB | [189] | |
| Microbiome | Diversity in the microbiota | P. umbilicalis | - Seasonal variation to the microbial community in laver - Identification of bacterial groups which are expected to contribute to the evolution and/or function of laver | [155] |
| P. yezoensis | - Seasonal variation and the effects of the yellow spot disease outbreaks to the microbial community in the seawater of laver seedling pools - Identification of disease-associated bacteria | [174] | ||
| Analytical techniques | P. umbilicalis | - Microbial communities affected by the sampling position of laver and the stabilization techniques applied for the microbiome analysis | [171] | |
| Influencing factor (Red rot disease) | P. yezoensis | - Alterations of bacterial community by red dot disease - Close association between health status of algal host (uninfected or infected) and bacterial community | [190] | |
| Proteome | Mechanism of stress-tolerance | P. haitanensis | - Investigation on the key metabolisms elucidating the mechanisms of resistance to high-temperature | [175,176] |
| P. orbicularis | - Investigation on the key metabolisms elucidating the mechanisms of resistance to desiccation | [177,178] | ||
| P. haitanensis | ||||
| Mechanism of infection resistance | P. yezoensis | - Investigation on the pathogen-responsive proteins elucidating the mechanisms of responses against the infection | [179] | |
| Identification of key functional protein | P. yezoensis | - Identification of major protein [Pyropia yezoensis aldehyde dehydognease (PyALDH)] which contributes to the resistance of laver against oxidative stress | [180] | |
| Mutation of laver strain | P. yezoensis | - Induction of high-growth-rate mutation by the exposure to ethyl methane sulfonate - Comparative analysis for the proteome of mutated strain with wild-type strain with the perspective to the enhanced growth | [181] | |
| P. yezoensis | - Induction of thermo-tolerance mutation by the exposure to gamma-irradiation - Isolation of protein from thermo-tolerant mutant which contributes to the resistance against elevated temperature | [182,191] | ||
| Lipidome | Lipidomic variations | P. haitanensis | - Identification of lipid biomarkers distinctly expressed under elevated temperatures | [184] |
| P. dioica | - Differences in composition of major lipid molecular species according to the life cycle stages between the blade and conchocelis | [185] | ||
| Metabolome | Metabolomic variations | P. haitanensis | - Changes in the nutrient composition according to the harvest time | [186] |
| P. yezoensis | - Changes in the nutrient composition which can determine the taste of laver by the food processing steps not only for seasoning but also washing, cutting, and roasting | [187] | ||
| Metabolite profile | P. pseudolinearis | - Distinctive characteristics of metabolites among species of edible seaweeds (brown, red, and green algae) and sorbitol as the major sugar metabolite in laver | [188] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, T.J.; Rhee, M.S. Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed. Mar. Drugs 2020, 18, 14. https://doi.org/10.3390/md18010014
Cho TJ, Rhee MS. Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed. Marine Drugs. 2020; 18(1):14. https://doi.org/10.3390/md18010014
Chicago/Turabian StyleCho, Tae Jin, and Min Suk Rhee. 2020. "Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed" Marine Drugs 18, no. 1: 14. https://doi.org/10.3390/md18010014
APA StyleCho, T. J., & Rhee, M. S. (2020). Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed. Marine Drugs, 18(1), 14. https://doi.org/10.3390/md18010014