Anti-Inflammatory Effects of 5α,8α-Epidioxycholest-6-en-3β-ol, a Steroidal Endoperoxide Isolated from Aplysia depilans, Based on Bioguided Fractionation and NMR Analysis

 ,
,  ,
,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Effect of Non-Polar Fraction of A. depilans Extract
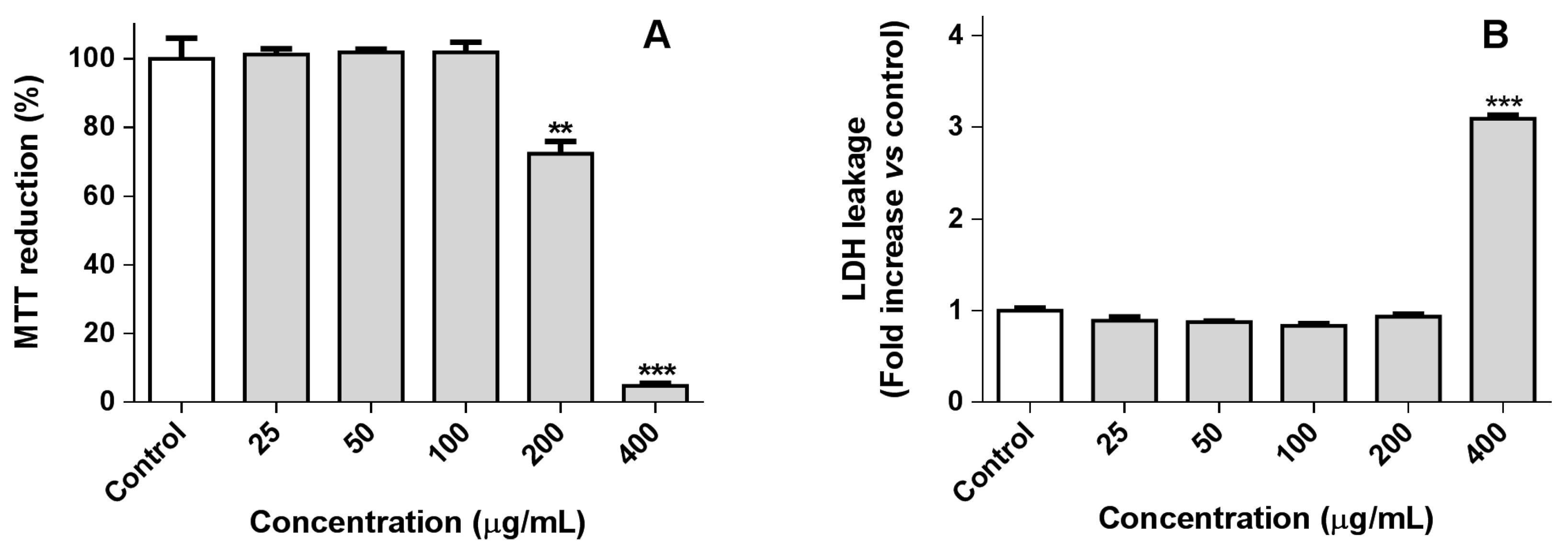
2.1.1. RAW 264.7 Macrophage Viability
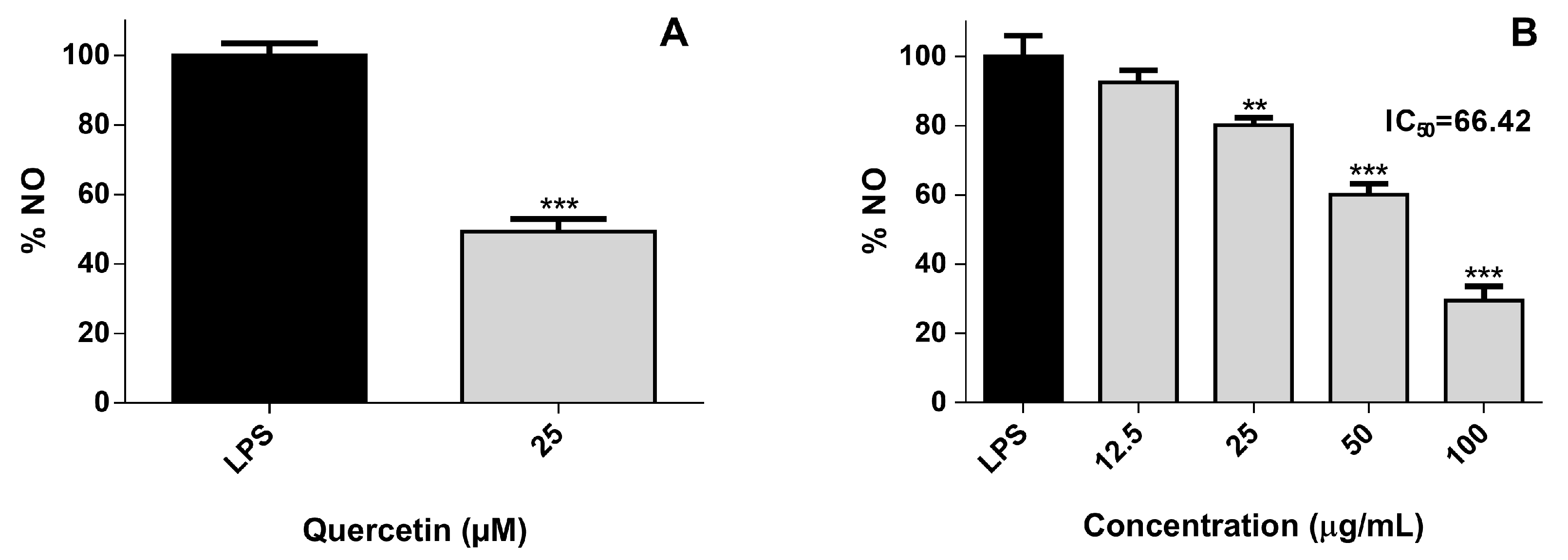
2.1.2. Cellular Nitric Oxide Levels
2.2. Effect of A. depilans Non-Polar Extract Sub-Fractions
2.3. Anti-Inflammatory Effect of EnP(5,8)
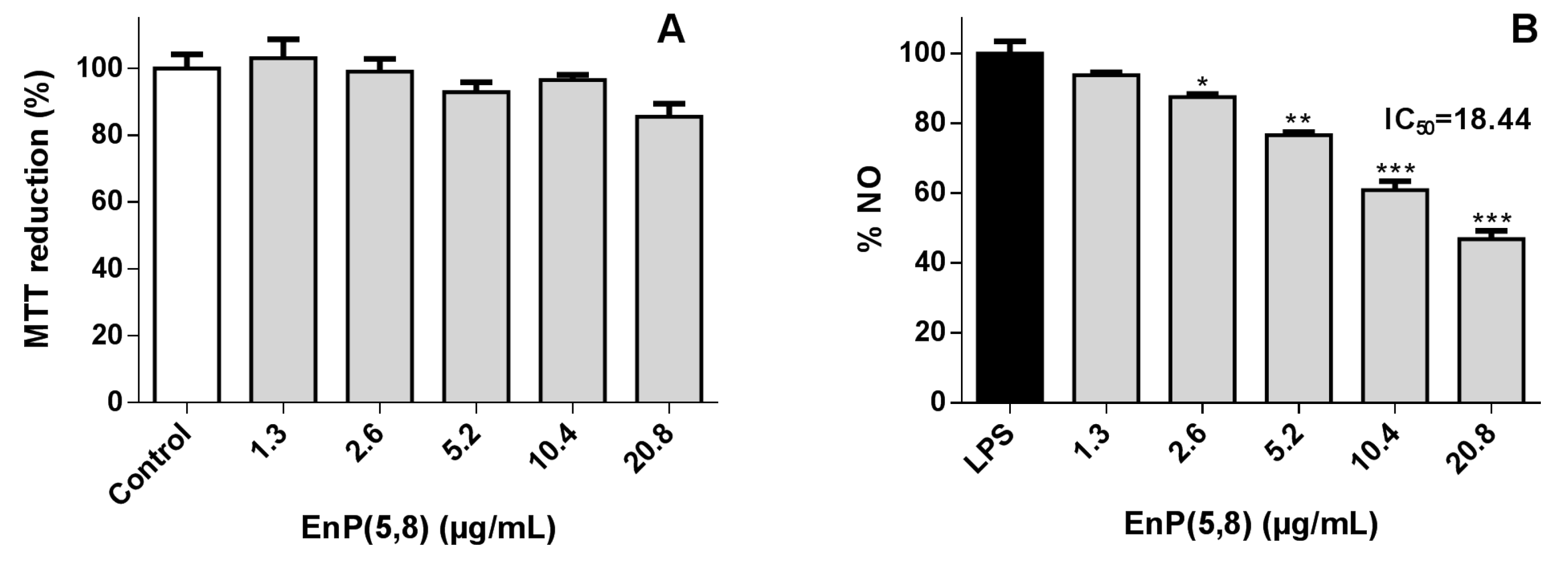
2.3.1. RAW 264.7 Macrophages Viability
2.3.2. Nitric Oxide Radical (●NO) Scavenging Assay
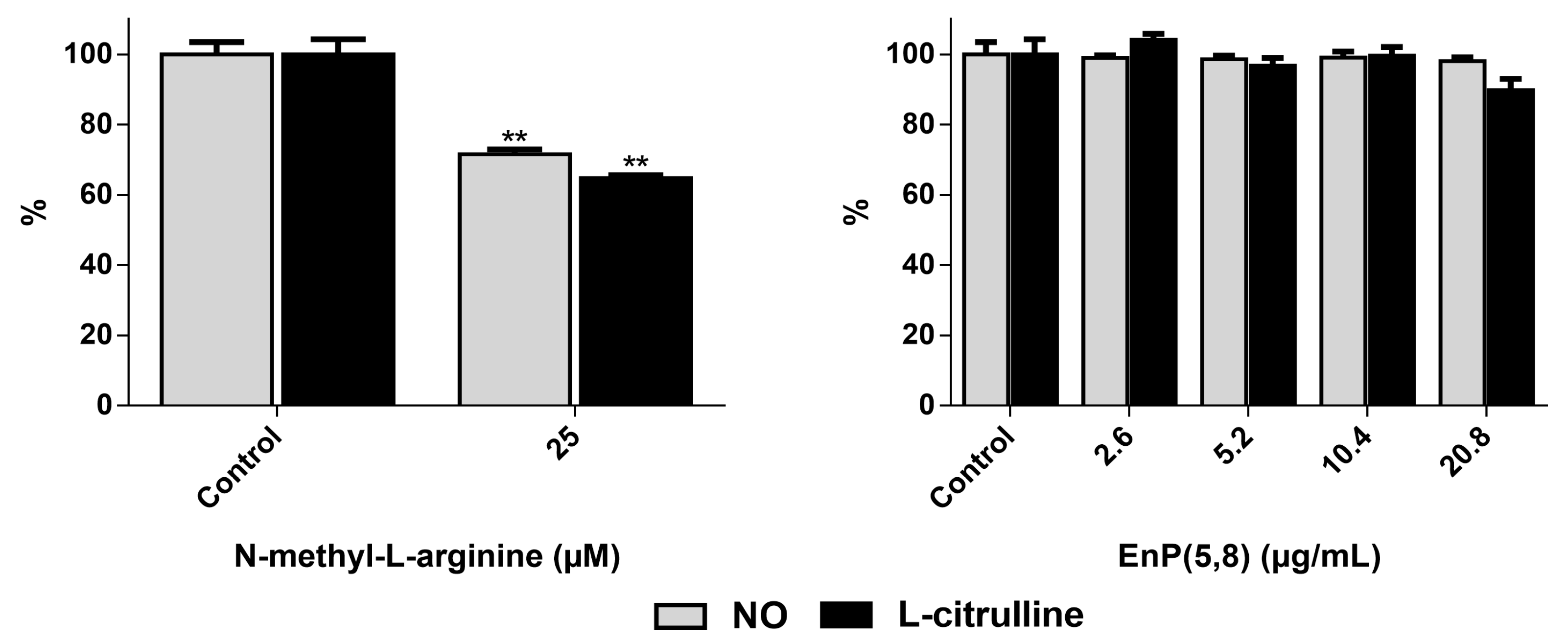
2.3.3. iNOS Direct Inhibition
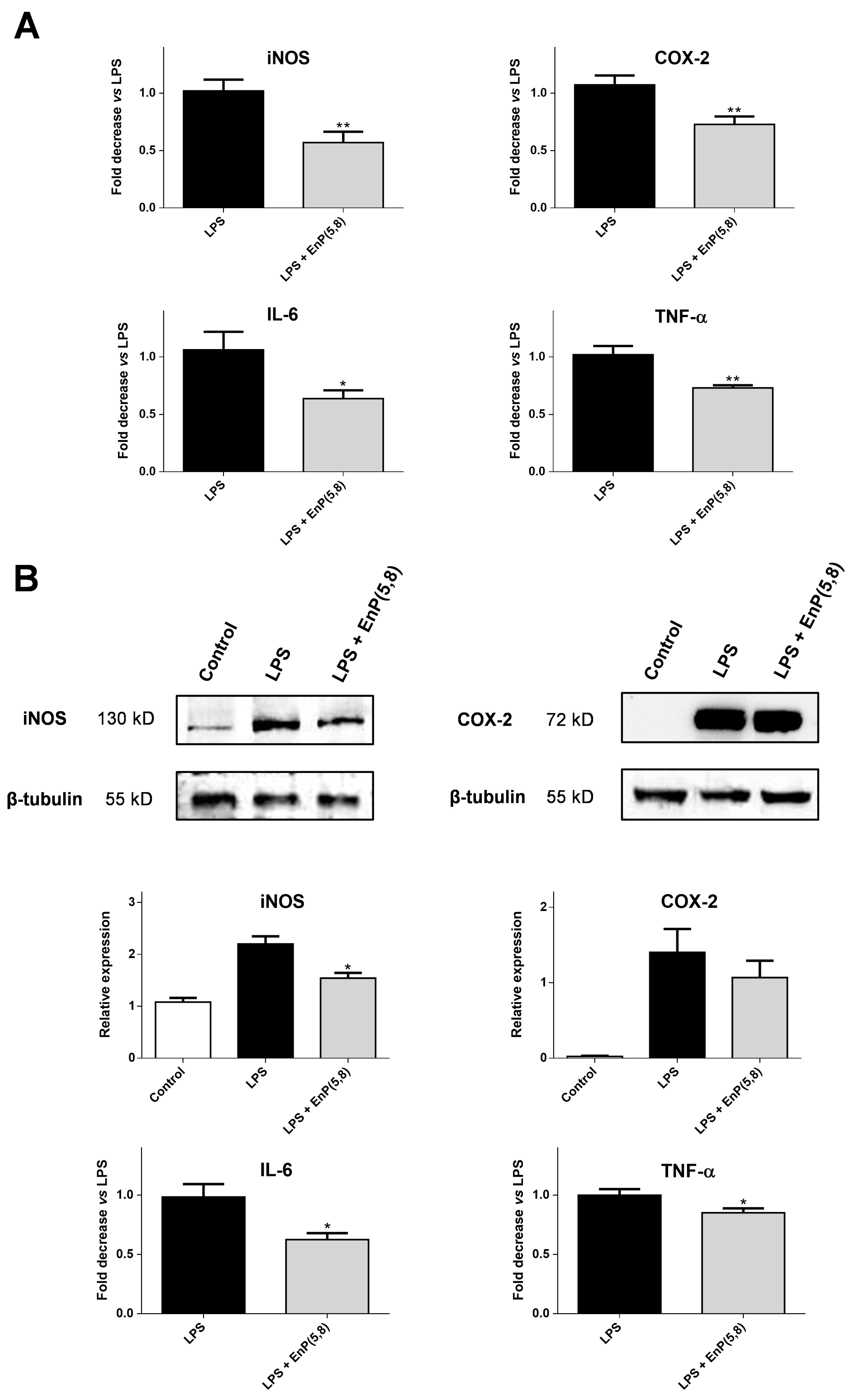
2.3.4. iNOS, COX-2, Interleukin 6 (IL-6), and Tumor Necrosis Factor Alpha (TNF-α) mRNA and Protein Expression
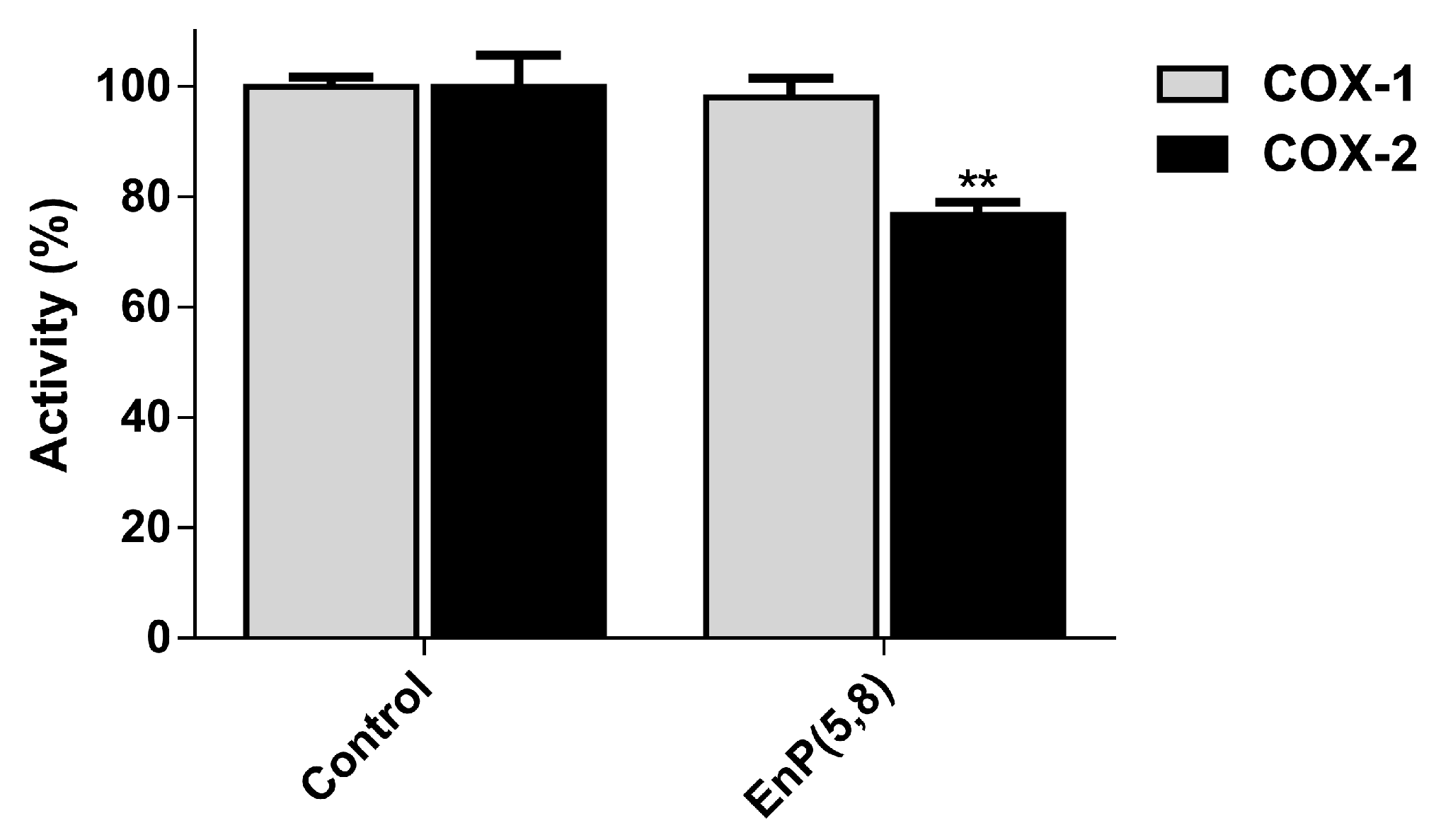
2.3.5. Arachidonic Acid Pathway Enzymes
3. Materials and Methods
3.1. Reagents and Standards
3.2. Sample
3.3. Extraction
3.4. Extract Fractionation
3.5. Cell Culture
3.6. MTT Reduction Assay
3.7. Membrane Integrity Assay
3.8. Determination of NO Levels
3.9. ●NO Scavenging Assay
3.10. iNOS Inhibition Assay
3.11. RNA Extraction, Quantification, Integrity, and Conversion
3.12. qPCR Analysis
3.13. Western Blot Analysis
3.14. Enzyme-Linked Immunosorbent Assay (ELISA)
3.15. PLA2 Inhibition Assay
3.16. 5-LOX Inhibition Assay
3.17. COX-1 and COX-2 Inhibition Assay
3.18. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pereira, R.B.; Andrade, P.B.; Valentão, P. Chemical diversity and biological properties of secondary metabolites from sea hares of Aplysia genus. Mar. Drugs 2016, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K. Molluscan biological and chemical diversity: Secondary metabolites and medicinal resources produced by marine molluscs. Biol. Rev. Camb. Philos. Soc. 2010, 85, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Pennings, S.C.; Paul, V.J. Sequestration of dietary secondary metabolites by three species of sea hares: Location, specificity and dynamics. Mar. Biol. 1993, 117, 535–546. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Lobo-da-Cunha, A.; Taveira, M.; Ferreira, M.; Valentão, P.; Andrade, P.B. Digestive gland from Aplysia depilans Gmelin: Leads for inflammation treatment. Molecules 2015, 20, 15766–15780. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.; Quiñoá, E.; Castedo, L.; Riguera, R. Epidioxy sterols from the tunicates Dendrodoa grossularia and Ascidiella aspersa and the gastropoda Aplysia depilans and Aplysia punctata. J. Nat. Prod. 1986, 49, 905–909. [Google Scholar] [CrossRef]
- Bu, M.; Yang, B.B.; Hu, L. Natural bioactive sterol 5α,8α-endoperoxides as drug lead compounds. Med. Chem. 2014, 4, 709–716. [Google Scholar] [CrossRef]
- Wu, S.J.; Lu, T.M.; Lai, M.N.; Ng, L.T. Immunomodulatory activities of medicinal mushroom Grifola frondosa extract and its bioactive constituent. Am. J. Chin. Med. 2013, 41, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Honda, M.; Sugiyama, S.; Higuchi, R.; Komori, T. Studies on the constituents of marine opisthobranchia III. Isolation and structure of two 5,8α-epidioxysterols and a cholesteryl ester mixture from the albumen gland of Aplysia juliana. Liebigs Annalen der Chemie 1988, 1988, 589–592. [Google Scholar] [CrossRef]
- Mou, X.-F.; Bian, W.-T.; Wang, C.-Y.; Shao, C.-L. Secondary metabolites isolated from the sea hare Aplysia pulmonica from the South China Sea. Chem. Nat. Compd. 2016, 52, 758–760. [Google Scholar] [CrossRef]
- Gunatilaka, A.A.L.; Gopichand, Y.; Schmitz, F.J.; Djerassi, C. Minor and trace sterols in marine invertebrates. 26. Isolation and structure elucidation of nine new 5.alpha.,8.alpha.-epidoxy sterols from four marine organisms. J. Org. Chem. 1981, 46, 3860–3866. [Google Scholar] [CrossRef]
- Aknin, M.; Viracaoundin, I.; Faure, R.; Gaydou, E.M. 5α,8α-Epidioxycholest-6-en-3-β-ol from three cone snails of the Indian ocean. J. Am. Oil Chem. Soc. 1998, 75, 1679–1681. [Google Scholar] [CrossRef]
- Ioannou, E.; Abdel-Razik, A.F.; Zervou, M.; Christofidis, D.; Alexi, X.; Vagias, C.; Alexis, M.N.; Roussis, V. 5α,8α-Epidioxysterols from the gorgonian Eunicella cavolini and the ascidian Trididemnum inarmatum: Isolation and evaluation of their antiproliferative activity. Steroids 2009, 74, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Minh, C.V.; Kiem, P.V.; Huong le, M.; Kim, Y.H. Cytotoxic constituents of Diadema setosum. Arch. Pharm. Res. 2004, 27, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.E.; Capper, A.; Della Togna, G.; Paul, V.J.; Romero, L.I.; Johns, T.; Cubilla-Rios, L.; Capson, T.L. Ecology- and bioassay-guided drug discovery for treatments of tropical parasitic disease: 5alpha, 8alpha-epidioxycholest-6-en-3beta-ol isolated from the mollusk Dolabrifera dolabrifera shows significant activity against Leishmania donovani. Nat. Prod. Commun. 2013, 8, 1537–1540. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.B.; Taveira, M.; Valentao, P.; Sousa, C.; Andrade, P.B. Fatty acids from edible sea hares: anti-inflammatory capacity in LPS-stimulated RAW 264.7 cells involves iNOS modulation. RSC Adv. 2015, 5, 8981–8987. [Google Scholar] [CrossRef]
- Pereira, R.B.; Pinto, D.C.G.A.; Pereira, D.M.; Gomes, N.G.M.; Silva, A.M.S.; Andrade, P.B.; Valentão, P. UHPLC-MS/MS profiling of Aplysia depilans and assessment of its potential therapeutic use: Interference on iNOS expression in LPS-stimulated RAW 264.7 macrophages and caspase-mediated pro-apoptotic effect on SH-SY5Y cells. J. Funct. Foods 2017, 37, 164–175. [Google Scholar] [CrossRef]
- Pereira, R.B.; Almeida, A.A.; Pereira, D.M.; Silva, O.; Andrade, P.B.; Pinto, E.; Valentão, P. Trace elements in wild edible Aplysia species: Relationship with the desaturation–elongation indexes of fatty acids. Chemosphere 2018, 208, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Chang, C.-W.; Tseng, Y.-J.; Lee, J.; Sung, P.-J.; Su, J.-H.; Hwang, T.-L.; Dai, C.-F.; Wang, H.-C.; Sheu, J.-H. Bioactive steroids from the formosan soft coral Umbellulifera petasites. Mar. Drugs 2016, 14, 180. [Google Scholar] [CrossRef]
- Castaneda, O.A.; Lee, S.C.; Ho, C.T.; Huang, T.C. Macrophages in oxidative stress and models to evaluate the antioxidant function of dietary natural compounds. J. Food Drug Anal. 2017, 25, 111–118. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.J.; Reum Jang, A.; Jang, H.J.; Yang, K.S. Inhibition of nitric oxide production, iNOS and COX-2 expression of ergosterol derivatives from Phellinus pini. Natural Product Sciences 2012, 18, 147–152. [Google Scholar]
- Lu, Y.; Lin, Y.-C.; Wen, Z.-H.; Su, J.-H.; Sung, P.-J.; Hsu, C.-H.; Kuo, Y.-H.; Chiang, M.Y.; Dai, C.-F.; Sheu, J.-H. Steroid and cembranoids from the Dongsha atoll soft coral Lobophytum sarcophytoides. Tetrahedron 2010, 66, 7129–7135. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, J.; Ma, W.; Fang, W.; Chen, Z.; Yang, B.; Liu, Y. Bioactivities of six sterols isolated from marine invertebrates. Pharm. Biol. 2014, 52, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Barnig, C.; Frossard, N.; Levy, B.D. Towards targeting resolution pathways of airway inflammation in asthma. Pharmacol. Ther. 2018, 186, 98–113. [Google Scholar] [CrossRef]
- Prasher, P.; Mudila, H.; Sharma, M.; Khati, B. Developmental perspectives of the drugs targeting enzyme-instigated inflammation: A mini review. Med. Chem. Res. 2019, 28, 417–449. [Google Scholar] [CrossRef]
- Pairet, M.; Engelhardt, G. Distinct isoforms (COX-1 and COX-2) of cyclooxygenase: Possible physiological and therapeutic implications. Fundam. Clin. Pharmacol. 1996, 10, 1–15. [Google Scholar] [CrossRef]
- Kam, P.C.A.; See, A.U.L. Cyclo-oxygenase isoenzymes: Physiological and pharmacological role. Anaesthesia 2000, 55, 442–449. [Google Scholar] [CrossRef]
- Gao, J.-M.; Wang, M.; Liu, L.-P.; Wei, G.-H.; Zhang, A.-L.; Draghici, C.; Konishi, Y. Ergosterol peroxides as phospholipase A2 inhibitors from the fungus Lactarius hatsudake. Phytomedicine 2007, 14, 821–824. [Google Scholar] [CrossRef]
- Khayat, M.T.; Ibrahim, S.R.M.; Mohamed, G.A.; Abdallah, H.M. Anti-inflammatory metabolites from endophytic fungus Fusarium sp. Phytochem. Lett. 2019, 29, 104–109. [Google Scholar] [CrossRef]
- Chang, M.C.; Uang, B.J.; Tsai, C.Y.; Wu, H.L.; Lin, B.R.; Lee, C.S.; Chen, Y.J.; Chang, C.H.; Tsai, Y.L.; Kao, C.J.; et al. Hydroxychavicol, a novel betel leaf component, inhibits platelet aggregation by suppression of cyclooxygenase, thromboxane production and calcium mobilization. Br. J. Pharmacol. 2007, 152, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Loza-Mejía, M.; Salazar, J. Sterols and triterpenoids as potential anti-inflammatories: Molecular docking studies for binding to some enzymes involved in inflammatory pathways. J. Mol. Graph. Model. 2015, 62, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mills, G.L.; Nair, M.G. Cyclooxygenase inhibitory and antioxidant compounds from the fruiting body of an edible mushroom, Agrocybe aegerita. Phytomedicine 2003, 10, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Ebada, S.S.; Edrada, R.A.; Lin, W.; Proksch, P. Methods for isolation, purification and structural elucidation of bioactive secondary metabolites from marine invertebrates. Nat. Protoc. 2008, 3, 1820–1831. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.T.; Andrade, P.B.; Paiva-Martins, F.; Valentão, P.; Pereira, D.M. In Vitro anti-inflammatory and cytotoxic effects of aqueous extracts from the edible sea anemones Anemonia sulcata and Actinia equina. Int. J. Mol. Sci. 2017, 18, 653. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.M.; Correia-da-Silva, G.; Valentão, P.; Teixeira, N.; Andrade, P.B. Anti-inflammatory effect of unsaturated fatty acids and ergosta-7,22-dien-3-ol from Marthasterias glacialis: Prevention of CHOP-mediated ER-stress and NF-κB activation. PLoS ONE 2014, 9, e88341. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jiménez, M.; Cabanes, J.; Gandía-Herrero, F.; Escribano, J.; García-Carmona, F.; Pérez-Gilabert, M. A continuous spectrophotometric assay for phospholipase A2 activity. Anal. Biochem. 2003, 319, 131–137. [Google Scholar] [CrossRef]
- Kachmar, M.R.; Oliveira, A.P.; Valentão, P.; Gil-Izquierdo, A.; Domínguez-Perles, R.; Ouahbi, A.; Badaoui, K.E.; Andrade, P.B.; Ferreres, F. HPLC-DAD-ESI/MSn phenolic profile and in vitro biological potential of Centaurium erythraea Rafn aqueous extract. Food Chem. 2019, 278, 424–433. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primers | Annealing Temperature (°C) | Amplicon Length (bp) |
|---|---|---|---|---|
| Nos2 (iNOS) | NM_001313921.1 | F: CCGCCGCTCTAATACTTA R: TTCATCAAGGAATTATACA | 58.0 | 121 |
| Ptgs2 (COX-2) | NM_011198.4 | F: TGAGTACCGCAAACGCTTCT R: CAGCCATTTCCTTCTCTCCTGT | 60.0 | 74 |
| Il6 (IL-6) | NM_001314054.1 | F: AGACAAAGCCAGAGTCCTTCAG R: TGACTCCAGCTTATCTCTTGGT | 59.0 | 75 |
| Tnf (TNF-α) | NM_001278601.1 | F: ACTGAACTTCGGGGTGATCG R: GTGGTTTGTGAGTGTGAGGGT | 59.0 | 100 |
| Actb (β-actin) | NM_007393.5 | F: TATAAAACCCGGCGGCGCA R: TCATCCATGGCGAACTGGTG | 61.5 | 117 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, R.B.; Pereira, D.M.; Jiménez, C.; Rodríguez, J.; Nieto, R.M.; Videira, R.A.; Silva, O.; Andrade, P.B.; Valentão, P. Anti-Inflammatory Effects of 5α,8α-Epidioxycholest-6-en-3β-ol, a Steroidal Endoperoxide Isolated from Aplysia depilans, Based on Bioguided Fractionation and NMR Analysis. Mar. Drugs 2019, 17, 330. https://doi.org/10.3390/md17060330
Pereira RB, Pereira DM, Jiménez C, Rodríguez J, Nieto RM, Videira RA, Silva O, Andrade PB, Valentão P. Anti-Inflammatory Effects of 5α,8α-Epidioxycholest-6-en-3β-ol, a Steroidal Endoperoxide Isolated from Aplysia depilans, Based on Bioguided Fractionation and NMR Analysis. Marine Drugs. 2019; 17(6):330. https://doi.org/10.3390/md17060330
Chicago/Turabian StylePereira, Renato B., David M. Pereira, Carlos Jiménez, Jaime Rodríguez, Rosa M. Nieto, Romeu A. Videira, Olga Silva, Paula B. Andrade, and Patrícia Valentão. 2019. "Anti-Inflammatory Effects of 5α,8α-Epidioxycholest-6-en-3β-ol, a Steroidal Endoperoxide Isolated from Aplysia depilans, Based on Bioguided Fractionation and NMR Analysis" Marine Drugs 17, no. 6: 330. https://doi.org/10.3390/md17060330
APA StylePereira, R. B., Pereira, D. M., Jiménez, C., Rodríguez, J., Nieto, R. M., Videira, R. A., Silva, O., Andrade, P. B., & Valentão, P. (2019). Anti-Inflammatory Effects of 5α,8α-Epidioxycholest-6-en-3β-ol, a Steroidal Endoperoxide Isolated from Aplysia depilans, Based on Bioguided Fractionation and NMR Analysis. Marine Drugs, 17(6), 330. https://doi.org/10.3390/md17060330