Screening of Chaetomorpha linum Lipidic Extract as a New Potential Source of Bioactive Compounds

 ,
,  ,
,  ,
, 

Abstract
1. Introduction
2. Results
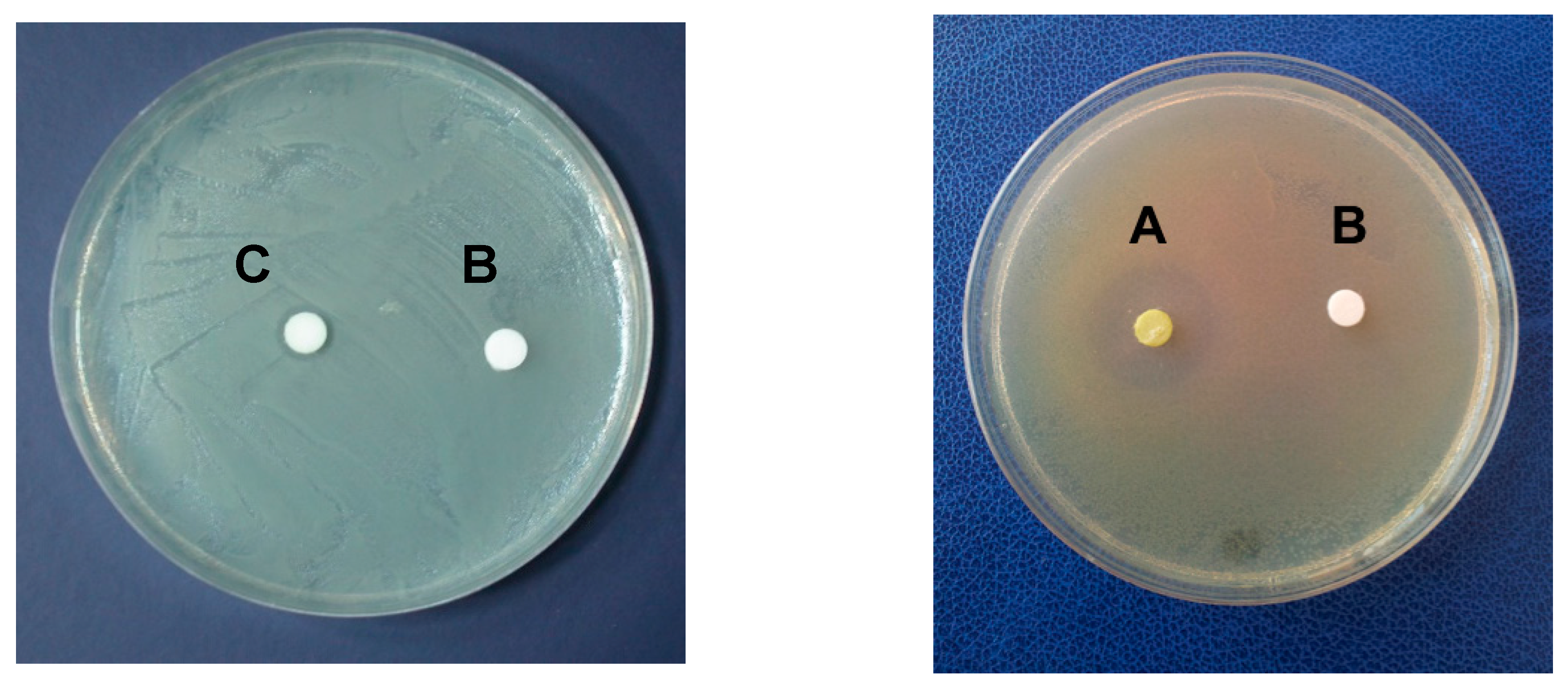
2.1. Antimicrobial Activity
2.2. Antioxidant Activity
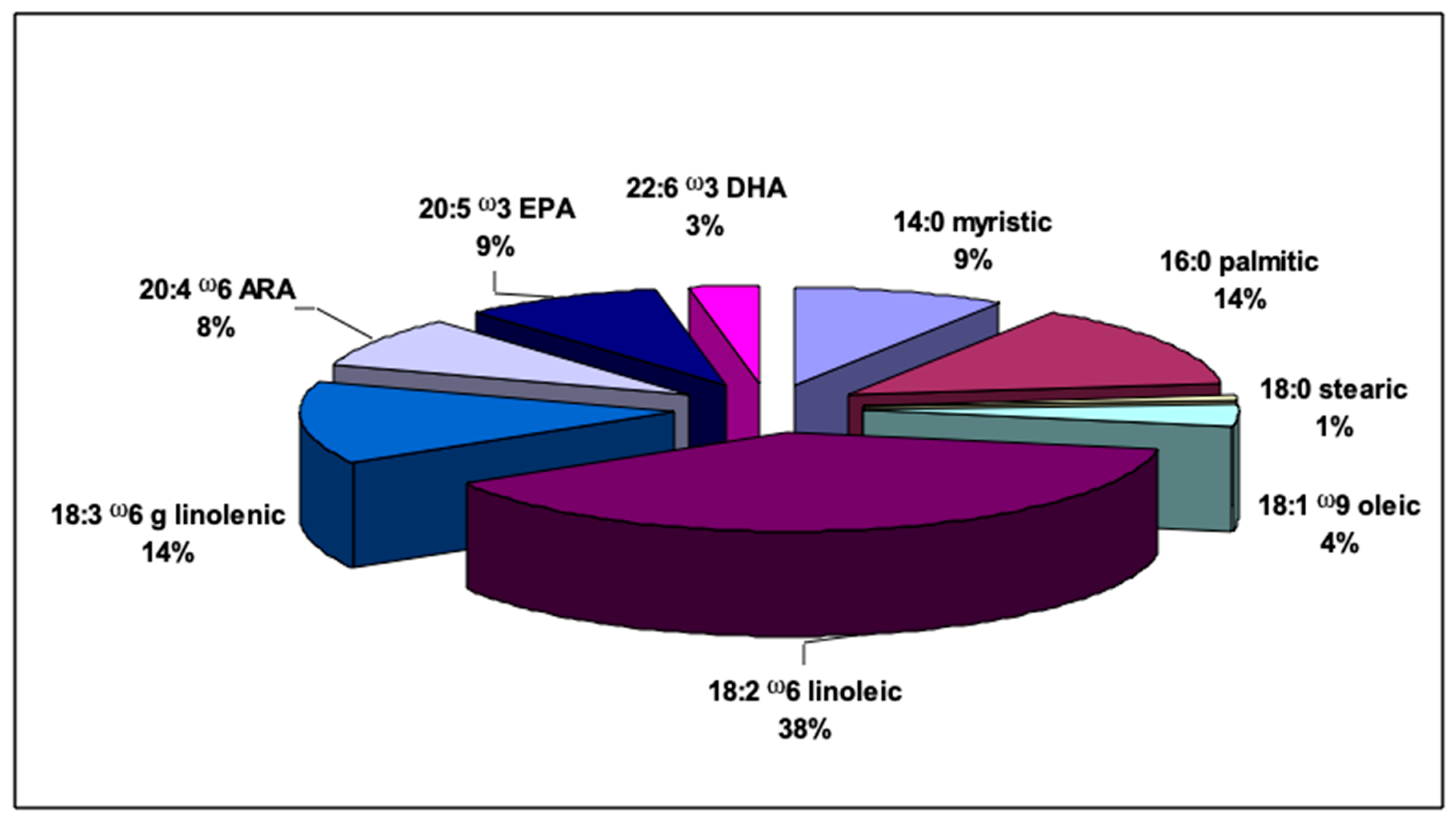
2.3. Fatty Acid Profile
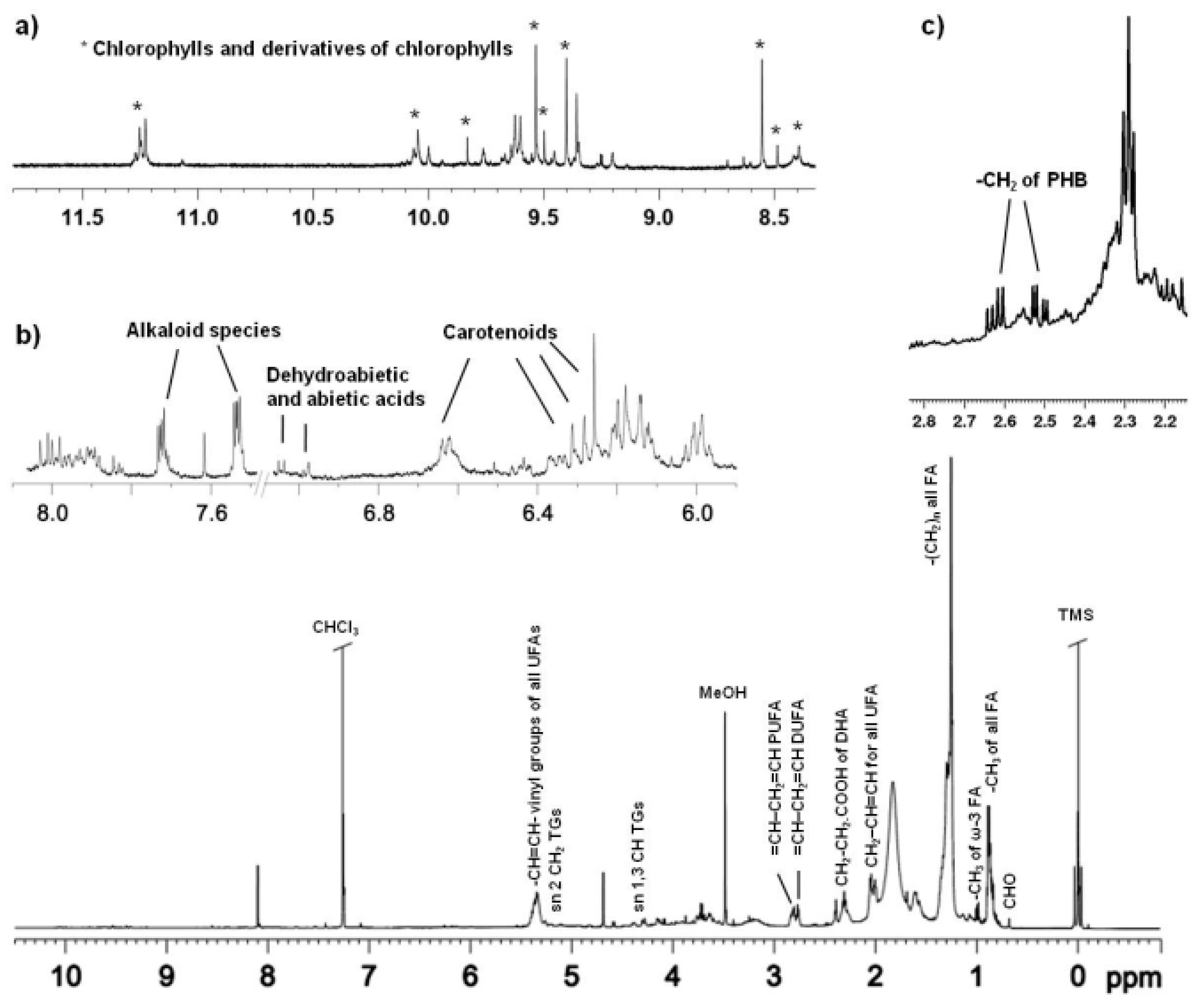
2.4. NMR Spectroscopy
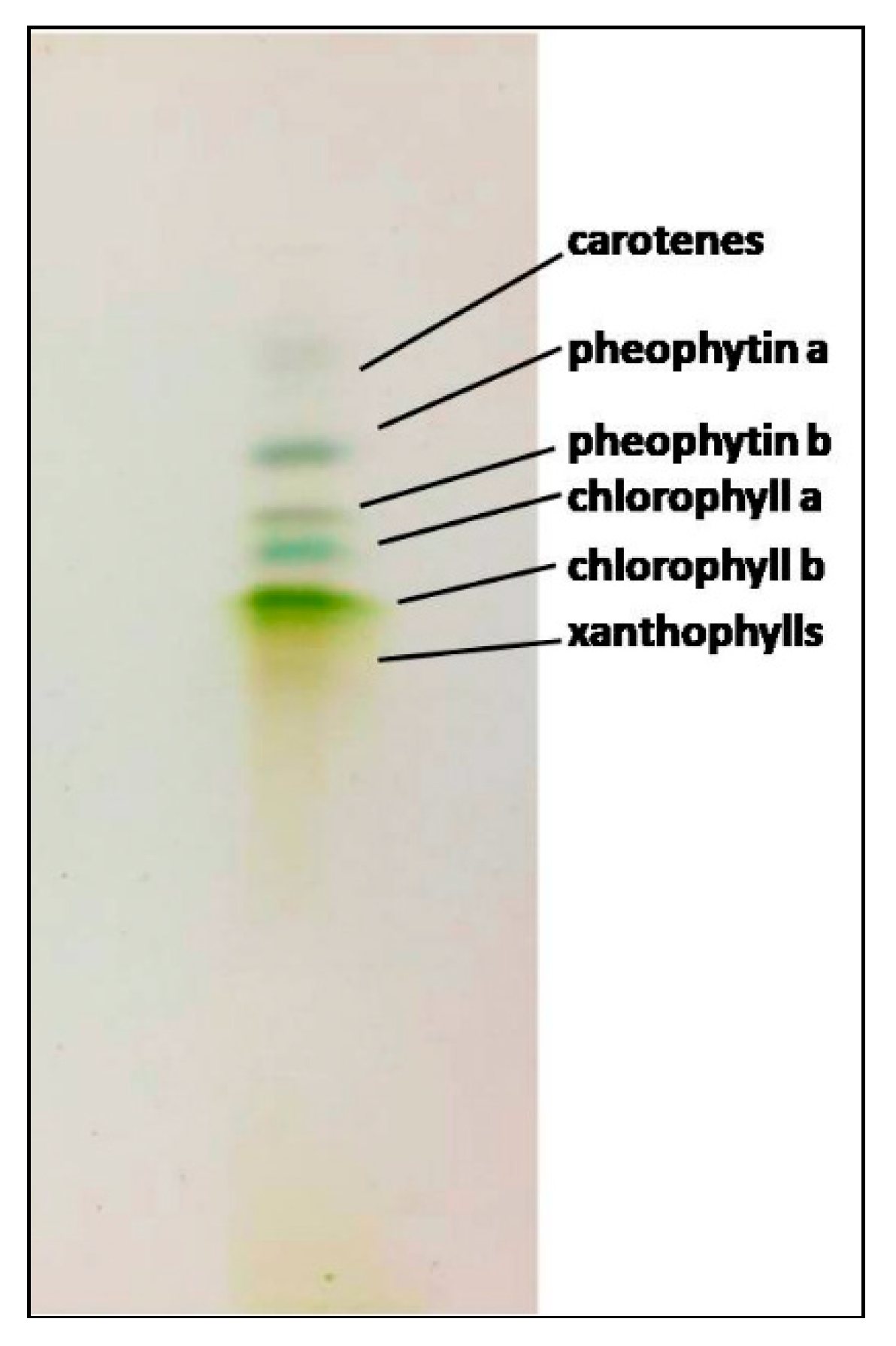
2.5. Thin-Layer Chromatography
2.6. Chaetomorpha Linum Cultivation Trials
3. Discussion
- Vibrio ordalii, previously designated as V. anguillarum biotype 2, causes worldwide serious haemorrhagic septicaemia inducing mortality in fish including cultured Atlantic salmon, Pacific salmon, rainbow trout, rockfish and gilthead sea bream [54,55,56,57]. Vibrio vulnificus is an opportunistic human pathogen causing three distinct syndromes: Primary septicemia, wound infection and gastroenteritis in susceptible individuals [58]. This species is highly lethal and is also responsible for several seafood-related deaths [59]. Similarly to other seafood-borne bacteria, it accumulates in the tissues of filter-feeders invertebrates such as oysters, clams and mussels. Infections are often acquired from eating raw oysters [60];
- The production of fish in aquaculture plants is becoming more and more necessary due to the increasing demand of fish food for a growing world population, especially in developing countries [63];
- The increase in production caused an increase in the use of antibiotics to contend with fish diseases. The main outcome of this growth was surely the development of an antibiotic resistance in the same fish and also in human consumers. Moreover, since antibiotics are served to fish through medicated feed, the excessive release in the environment of feed residues caused accumulation of antibiotics also into the sediments and the resultant selection of resistant microorganisms [64].
4. Materials and Methods
4.1. Species Description and Collection
4.2. Preparation of Lipidic Extracts from the Macroalga
4.3. Test Microorganisms
4.4. Antimicrobial Activity
4.5. Antioxidant Activity
4.5.1. Oxygen Radical Absorbance Capacity Assay (ORAC)
4.5.2. Trolox Equivalent Antioxidant Capacity Assay (TEAC)
4.5.3. Folin–Ciocalteu (F–C) Assay
4.6. Gas Chromatographic Analysis of Fatty Acid
4.7. NMR Spectroscopy
4.8. Thin-Layer Chromatography
4.9. Chaetomorpha Linum Cultivation Trials
Author Contributions
Funding
Conflicts of Interest
References
- Valli, M.; Pivatto, M.; Danuello, A.; Castro-Gamboa, I.; Silva, D.H.S.; Cavalheiro, A.J.; Araújo, Â.R.; Furlan, M.; Lopes, M.N.; da Silva Bolzani, V. Tropical biodiversity: Has it been a potential source of secondary metabolites useful for medicinal chemistry? Quím. Nova 2012, 35, 2278–2287. [Google Scholar] [CrossRef]
- Duarte, K.; Justino, C.I.L.; Pereira, R.; Freitas, A.C.; Gomes, A.M.; Duarte, A.C.; Rocha-Santos, T.A.P. Green analytical methodologies for the discovery of bioactive compounds from marine sources. Trends Environ. Anal. Chem. 2014, 3–4, 43–52. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Petrocelli, A.; Cecere, E. The lipidic extract of the seaweed Gracilariopsis longissima (Rhodophyta, Gracilariales): A potential resource for biotechnological purposes? New Biotechnol. 2012, 29, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Cecere, E.; Petrocelli, A. Biotechnological potential of the seaweed Cladophora rupestris (Chlorophyta, Cladophorales) lipidic extract. New Biotechnol. 2014, 31, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine bioactive compounds and health promoting perspectives; innovation pathways for drug discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar] [CrossRef]
- Rasmussen, R.S.; Morrissey, M.T. Marine biotechnology for production of food ingredients. Adv. Food Nutr. Res. 2007, 52, 237–292. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Antibacterial derivatives of marine algae: An overview of pharmacological mechanisms and applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Zerrifi, S.; El Khalloufi, F.; Oudra, B.; Vasconcelos, V. Seaweed bioactive compounds against pathogens and microalgae: Potential uses on pharmacology and harmful algae bloom control. Mar. Drugs 2018, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Kelman, D.; Posner, E.K.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef] [PubMed]
- De Nys, R.; Guenther, J.; Uriz, M.J. Natural control of fouling. In Biofouling; Durr, S., Thomason, J.C., Eds.; Wiley–Blackwell: Oxford, UK, 2010; pp. 109–116. [Google Scholar]
- Hay, M.E. Seaweed chemical ecology: Its importance to individuals, populations, and communities. J. Phycol. 2001, 37, 22–23. [Google Scholar] [CrossRef]
- Williamson, J.E.; Carson, D.G.; de Nys, R.; Steinberg, P.D. Demographic consequences of an ontogenetic shift by a sea urchin in response to host plant chemistry. Ecology 2004, 85, 1355–1371. [Google Scholar] [CrossRef]
- Hay, M.E. Marine chemical ecology: Chemical signals and cues structure marine populations, communities, and ecosystems. Annu. Rev. Mar. Sci. 2009, 1, 193–212. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine pharmacology in 2009–2011: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef] [PubMed]
- Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Scheld, M.; Bartlett, J.G. Bad bugs need drugs: An update on the development pipeline from the antibacterial availability task force of the IDSA. Clin. Infect. Dis. 2006, 42, 657–668. [Google Scholar] [CrossRef]
- Manivasagan, P.; Kang, K.H.; Sivakumar, K.; Li-Chan, E.C.; Oh, H.M.; Kim, S.K. Marine actinobacteria: An important source of bioactive natural products. Environ. Toxicol. Pharmacol. 2014, 38, 172–188. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Ventola, C.L. The antibiotic resistance crisis: Part 2: Management strategies and new agents. Pharm. Ther. 2015, 40, 344–352. [Google Scholar]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef]
- Baltz, R.H. Marcel faber roundtable: Is our antibiotic pipeline unproductive because of starvation, constipation or lack of inspiration? J. Ind. Microbiol. Biotechnol. 2006, 33, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Peláez, F. The historical delivery of antibiotics from microbial natural products-can history repeat? Biochem. Pharmacol. 2006, 71, 981–990. [Google Scholar] [CrossRef]
- Smit, A.J. Bioactivity and pharmacology of seaweed natural products. In World Seaweed Resources: An Authoritative Reference System; Critchley, A.T., Ohno, M., Largo, D., Eds.; UNESCO: Paris, France, 2006; pp. 1–20. [Google Scholar]
- Smit, A.J. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Biological activities and potential industrial applications of fucose rich sulphated polysaccharides and fucoidans isolated from brown seaweeds: A review. Carbohydr. Polym. 2012, 88, 13–20. [Google Scholar] [CrossRef]
- Stabili, L.; Fraschetti, S.; Acquaviva, M.I.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Gerardi, C.; Narracci, M.; Rizzo, L. The potential exploitation of the Mediterranean invasive alga Caulerpa cylindracea: Can the invasion be transformed into a gain? Mar. Drugs 2016, 14, 210. [Google Scholar] [CrossRef]
- Bansemir, A.; Blume, M.; Schröder, S.; Lindequist, U. Screening of cultivated seaweeds for antibacterial activity against fish pathogenic bacteria. Aquaculture 2006, 252, 79–84. [Google Scholar] [CrossRef]
- Ganeshamurthy, R.; Kumar, T.T.A.; Dhayanithi, N.B. Effect of secondary metabolites of the seaweed (Halimeda micronesia) at Lakshadweep Islands against aquatic pathogens. Int. J. Pharm. Biol. Sci. 2012, 3, B213–B220. [Google Scholar]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar] [CrossRef]
- Boonchum, W.; Peerapornpisal, Y.; Kanjanapothi, D.; Pekkoh, J.; Pumas, C.; Jamjai, U.; Amornlerdpison, D.; Noiraksar, T.; Vacharapiyasophon, P. Antioxidant activity of some seaweeds from the Gulf of Thailand. Int. J. Agric. Biol. 2011, 13, 95–99. [Google Scholar] [CrossRef]
- Sutour, S.; Xu, T.; Casabianca, H.; Paoli, M.; De Rocca-Serra, D.; Tomi, F.; Garrido, M.; Pasqualini, M.; Aiello, A.; Castola, V.; et al. Chemical composition of extracts from Chaetomorpha linum (Miller) Kütz. A potential use in the cosmetic industry. Int. J. Phytocosmet. Nat. Ingred. 2015, 2, 5. [Google Scholar] [CrossRef]
- Rindi, F.; Verbruggen, H. Taxonomic reshuffling of the Cladophoraceae. J. Phycol. 2016, 52, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 14 December 2018).
- Novaczek, I. A Guide to the Common Edible and Medicinal Sea Plants of the Pacific Islands; The University of the South Pacific: Suva, Fiji, 2001; p. 6. ISBN 982-203-835-6. [Google Scholar]
- Farasat, M.; Khavari-Nejad, R.A.; Nabavi, S.M.B.; Namjooyan, F. Antioxidant properties of some filamentous green algae (Chaetomorpha Genus). Braz. Arch. Biol. Technol. 2013, 56, 921–927. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef]
- Patarra, R.F.; Leite, J.; Pereira, R.; Baptista, J.; Neto, A.I. Fatty acid composition of selected macrophytes. Nat. Prod. Res. 2013, 27, 665–669. [Google Scholar] [CrossRef]
- Sfriso, A.; Wolf, M.A.; Moro, I.; Sfriso, A.A.; Buosi, A.; Verlaque, M. Rediscovery of a Forgotten Mediterranean Chaetomorpha Species in the Venice Lagoon (North Adriatic Sea): Chaetomorpha stricta Schiffner (Cladophorales, Chlorophyta). Cryptogam. Algol. 2018, 39, 293–312. [Google Scholar] [CrossRef]
- Menéndez, M. Effect of nutrient pulses on photosynthesis of Chaetomorpha linum from a shallow Mediterranean coastal lagoon. Aquat. Bot. 2005, 82, 181–192. [Google Scholar] [CrossRef]
- Cecere, E.; Portacci, G.; Petrocelli, A. Fragmentation and ball-like forms of Chaetomorpha linum (Cladophorales, Chlorophyta) in the Mar Piccolo of Taranto. Biol. Mar. Mediterr. 2009, 16, 252–253. [Google Scholar]
- Minicante, S.A.; Michelet, S.; Bruno, F.; Castelli, G.; Vitale, F.; Sfriso, A.; Morabito, M.; Genovese, G. Bioactivity of phycocolloids against the mediterranean protozoan Leishmania infantum: An inceptive study. Sustainability 2016, 8, 1131. [Google Scholar] [CrossRef]
- Petrocelli, A.; Cecere, E.; Rubino, F. Successions of phytobenthos species in a Mediterranean transitional water system: The importance of long term observations. Nat. Conserv. 2019, 34, 217–246. [Google Scholar] [CrossRef]
- Mannina, L.; Sobolev, A.P.; Capitani, D.; Iaffaldano, N.; Rosato, M.P.; Ragni, P.; Reale, A.; Sorrentino, E.; D’Amico, I.; Coppola, R. NMR metabolic profiling of organic and aqueous sea bass extracts: Implications in the discrimination of wild and cultured sea bass. Talanta 2008, 77, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Alexandri, E.; Ahmed, R.; Siddiqui, H.; Choudhary, M.; Tsiafoulis, C.; Gerothanassis, I. High resolution NMR spectroscopy as a structural and analytical tool for unsaturated lipids in solution. Molecules 2017, 22, 1663. [Google Scholar] [CrossRef]
- Kumar, R.; Bansal, V.; Patel, M.B.; Sarpal, A.S. Compositional analysis of algal biomass in a Nuclear Magnetic Resonance (NMR) tube. J. Algal Biomass Util. 2014, 5, 36–45. [Google Scholar]
- Costa, M.S.; Rego, A.; Ramos, V.; Afonso, T.B.; Freitas, S.; Preto, M.; Lopes, V.; Vasconcelos, V.; Magalhães, C.; Leão, P.N. The conifer biomarkers dehydroabietic and abietic acids are widespread in Cyanobacteria. Sci. Rep. 2016, 6, 23436. [Google Scholar] [CrossRef] [PubMed]
- Ramnath, M.G.; Thirugnanasampandan, R.; Sadasivam, M.; Mohan, P.S. Antioxidant, antibacterial and antiacetylcholinesterase activities of abietic acid from Isodon wightii (Bentham) H. Hara. Free Rad. Antiox. 2015, 5, 1–5. [Google Scholar] [CrossRef]
- Sobolev, A.P.; Brosio, E.; Gianferri, R.; Segre, A.L. Metabolic profile of lettuce leaves by high-field NMR spectra. Magn. Reson. Chem. 2005, 43, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Valverde, J.; This, H. 1H NMR quantitative determination of photosynthetic pigments from green beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2008, 56, 314–320. [Google Scholar] [CrossRef]
- Guadagno, C.R.; Greca, M.D.; De Santo, A.V.; D’Ambrosio, N. NMR (1H) analysis of crude extracts detects light stress in Beta vulgaris and Spinacia oleracea leaves. Photosynth. Res. 2013, 115, 115–122. [Google Scholar] [CrossRef]
- De Pascali, S.A.; Del Coco, L.; Felline, S.; Mollo, E.; Terlizzi, A.; Fanizzi, F.P. 1H NMR spectroscopy and MVA analysis of Diplodus sargus eating the exotic pest Caulerpa cylindracea. Mar. Drugs 2015, 13, 3550–3566. [Google Scholar] [CrossRef]
- Sarjito, S.; Radjasa, O.K.; Sabdono, A.; Prayitno, S.B.; Hutabarat, S. Phylogenetic diversity of the causative agents of vibriosis associated with groupers fish from Karimunjawa Islands, Indonesia. Curr. Res. Bacteriol. 2009, 2, 14–21. [Google Scholar]
- Novriadi, R. Vibriosis in aquaculture. OmniAkuatika 2016, 12, 1–12. [Google Scholar] [CrossRef]
- Muroga, K.; Yasuhiko, J.; Masumura, K. Vibrio ordalii isolated from diseased ayu (Plecoglossus altivelis) and rockfish (Sebastes schlegeli). Fish Pathol. 1986, 21, 239–243. [Google Scholar] [CrossRef]
- Colquhoun, D.J.; Aase, I.L.; Wallace, C.; Baklien, Å.; Gravningen, K. First description of Vibrio ordalii from Chile. Bull. Eur. Assoc. Fish Pathol. 2004, 24, 185–188. [Google Scholar]
- Silva-Rubio, A.; Acevedo, C.; Magariños, B.; Jaureguiberry, B.; Toranzo, A.E.; Avendaño-Herrera, R. Antigenic and molecular characterization of Vibrio ordalii strains isolated from Atlantic salmon Salmo salar in Chile. Dis. Aquat. Org. 2008, 79, 27–35. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akayli, T.; Timur, G.; Albayrak, G.; Aydemir, B. Identification and genotyping of Vibrio ordalii: A comparison of different methods. Isr. J. Aquac. Bamigdeh 2010, 62, 9–18. [Google Scholar]
- Strom, M.S.; Paranjpye, R.N. Epidemiology and pathogenesis of Vibrio vulnificus. Microbes Infect. 2000, 2, 177–188. [Google Scholar] [CrossRef]
- Tao, Z.; Larsen, A.M.; Bullard, S.A.; Wright, A.C.; Arias, C.R. Prevalence and population structure of Vibrio vulnificus on fishes from the northern Gulf of Mexico. Appl. Environ. Microbiol. 2012, 78, 7611–7618. [Google Scholar] [CrossRef] [PubMed]
- Daniels, N.A. Vibrio vulnificus oysters: Pearls and Perils. Clin. Infect. Dis. 2011, 52, 788–792. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H. Major bacterial diseases in aquaculture and their vaccine development. Anim. Sci. Rev. 2012, 7, 141–156. [Google Scholar] [CrossRef]
- Genschick, S.; Marinda, P.; Tembo, G.; Kaminski, A.M.; Thilsted, S.H. Fish consumption in urban Lusaka: The need for aquaculture to improve targeting of the poor. Aquaculture 2018, 492, 280–289. [Google Scholar] [CrossRef]
- Miranda, C.D.; Godoy, F.A.; Lee, M. Current status of the use of antibiotics and their antimicrobial resistance in the Chilean salmon farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.; Ripol, A.; Afonso, C.; Freire, M.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Bandarra, N. Fatty acid profiles of the main lipid classes of green seaweeds from fish pond aquaculture. Food Sci. Nutr. 2017, 5, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Ktari, L.; Romdhane, Y.B.R.; Aoun, B.; Sadok, S.; Boudabous, A.; El Bour, M. Antimicrobial fatty acids from green alga Ulva rigida (Chlorophyta). BioMed Res. Int. 2018, 2018, 3069595. [Google Scholar] [CrossRef]
- Floreto, E.A.T.; Hirata, H.; Ando, S.; Yamasaki, S. Fatty acid composition of Ulva pertusa Kjellman (Chlorophyta) and Gracilaria incurvata Okamura (Rhodophyta) in Japanese coastal waters. Bot. Mar. 1993, 36, 217–222. [Google Scholar] [CrossRef]
- Banaimoon, S.A. Fatty acids in marine macroalgae from southern Yemen (Hadramout) including occurrence of eicosatetraenoic (20:4) and eicosapentaenoic (20:5) acids. Bot. Mar. 1992, 35, 165–168. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Levchenko, E.V. Lipids of the red alga Gracilaria verrucosa (Huds.) Papenf. Bot. Mar. 1997, 40, 541–546. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Vaskovsky, V.E.; Titlyanova, T.V. Fatty acids of marine algae from the Pacific coast of North California. Bot. Mar. 2002, 45, 17–22. [Google Scholar] [CrossRef]
- Van Ginneken, V.J.T.; Helsper, J.P.F.G.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef]
- Ibarguren, M.; López, D.J.; Escribá, P.V. The effect of natural and synthetic fatty acids on membrane structure, microdomain organization, cellular functions and human health. Biochim. Biophys. Acta Biomembr. 2014, 1838, 1518–1528. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Harel, M.; Koven, W.; Lein, I.; Bar, Y.; Behrens, P.; Stubblefield, J.; Zohar, Y.; Place, A.R. Advanced DHA, EPA and ArA enrichment materials for marine aquaculture using single cell heterotrophs. Aquaculture 2002, 213, 347–362. [Google Scholar] [CrossRef]
- Chin, H.J.; Shen, T.F.; Su, H.P.; Ding, S.T. Schizochytrium limacinum SR-21 as a source of docosahexaenoic acid: Optimal growth and use as a dietary supplement for laying hens. Aust. J. Agric. Res. 2006, 57, 13–20. [Google Scholar] [CrossRef]
- Shi, Q.; Rong, H.; Hao, M.; Zhu, D.; Aweya, J.J.; Li, S.; Wen, X. Effects of dietary Sargassum horneri on growth performance, serum biochemical parameters, hepatic antioxidant status, and immune responses of juvenile black sea bream Acanthopagrus schlegelii. J. Appl. Phycol. 2019, 31, 771–778. [Google Scholar] [CrossRef]
- Gonzalez, M.A.; Perez-Guaita, D.; Correa-Royero, J.; Zapata, B.; Agudelo, L.; Mesa-Arango, A.; Betancur-Galvis, L. Synthesis and biological evaluation of dehydroabietic acid derivatives. Eur. J. Med. Chem. 2010, 45, 811–816. [Google Scholar] [CrossRef]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Cerón, M.C.; García-Malea, M.C.; Rivas, J.; Acien, F.G.; Fernandez, J.M.; Del Río, E.; Guerrero, M.G.; Molina, E. Antioxidant activity of Haematococcus pluvialis cells grown in continuous culture as a function of their carotenoid and fatty acid content. Appl. Microbiol. Biotechnol. 2007, 74, 1112–1119. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Chao, P.Y.; Hu, S.P.; Yang, C.M. The antioxidant and free radical scavenging activities of chlorophylls and pheophytins. Food Nutr. Sci. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Plaza, M.; Cifuentes, A.; Ibáñez, E. In the search of new functional ingredients from algae. Trends Food Sci. Technol. 2008, 19, 31–39. [Google Scholar] [CrossRef]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Zaheer, M.R.; Kuddus, M. PHB (poly-b-hydroxybutyrate) and its enzymatic degradation. Polym. Adv. Technol. 2018, 29, 30–40. [Google Scholar] [CrossRef]
- Abbing, M.R. Plastic Soup an Atlas of Ocean Pollution; Island Press: London, UK, 2019; pp. 1–124. [Google Scholar]
- Pachekoski, W.M.; Agnelli, J.A.M.; Belem, L.P. Thermal, mechanical and morphological properties of poly(hydrohybutyrate) and polypropylene blends after processing. Mater. Res. 2009, 12, 159–164. [Google Scholar] [CrossRef]
- Karan, H.; Funk, C.; Grabert, M.; Oey, M.; Hankamer, B. Green bioplastics as part of a circular bioeconomy. Trends Plant Sci. 2019, 24, 237–249. [Google Scholar] [CrossRef]
- Noreen, A.; Zia, K.M.; Zuber, M.; Ali, M.; Mujahid, M. A critical review of algal biomass: A versatile platform of bio-based polyesters from renewable resources. Int. J. Biol. Macromol. 2016, 86, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Kientz, B.; Thabard, M.; Cragg, S.M.; Pope, J.; Hellio, C. A new method for removing microflora from macroalgal surfaces: An important step for natural product discovery. Bot. Mar. 2011, 54, 457–469. [Google Scholar] [CrossRef]
- Cavallo, R.A.; Stabili, L. Presence of vibrios in seawater and Mytilus galloprovincialis (Lam.) from the Mar Piccolo of Taranto (Ionian Sea). Water Res. 2002, 36, 3719–3726. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Cavallo, R.A. Mytilus galloprovincialis filter feeding on the bacterial community in a Mediterranean coastal area (Northern Ionian Sea, Italy). Water Res. 2005, 39, 469–477. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Cavallo, R.; Acquaviva, M.; Stabili, L.; Cecere, E.; Petrocelli, A.; Narracci, M. Antibacterial activity of marine macroalgae against fish pathogenic Vibrio species. Cent. Eur. J. Biol. 2013, 8, 646–653. [Google Scholar] [CrossRef]
- Ely, R.; Supriya, T.; Naik, C.G. Antimicrobial activity of marine organisms collected off the coast of South East India. J. Exp. Mar. Biol. Ecol. 2004, 309, 121–127. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC-fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improvement ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Magalhaes, L.M.; Santos, F.; Segundo, M.A.; Reis, S.; Lima, J.L.F.C. Rapid microplate high-throughput methodology for assessment of Folin-Ciocalteu reducing capacity. Talanta 2010, 83, 441–447. [Google Scholar] [CrossRef]
- Folch, J.; Less, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–508. [Google Scholar]
- Budge, S.M.; Parrish, C.C. FA determination in cold water marine samples. Lipids 2003, 38, 781–791. [Google Scholar] [CrossRef]
- Pavia, D.L.; Lampman, G.M.; Kriz, G.S.; Engel, R.G. Isolation of chlorophyll and carotenoid pigments from spinach. In Introduction to Organic Laboratory Techniques: A Microscale Approach, 3rd ed.; Saunders College Publishing, Ed.; Brooks Cole: Pacific Grove, CA, USA, 1999; pp. 1–7. [Google Scholar]
- Rebecca, L.J.; Sharmila, S.; Das, M.P.; Seshiah, C. Extraction and purification of carotenoids from vegetables. J. Chem. Pharm. Res. 2014, 6, 594–598. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Strain | Diameter of Growth Inhibition (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 μL | 5 μL | 10 μL | 20 μL | 30 μL | 40 μL | 60 μL | 80 μL | 100 μL | |
| Candida albicans | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Candida famata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Candida glabrata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Enterococcus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pseudomonas sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Staphylococcus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Streptococcus agalactiae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio alginolyticus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio harveyi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio mediterranei | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio ordalii | 0 | 8 | 12 | 12 | 12 | 12 | 12 | 12 | 12 |
| Vibrio parahaemolyticus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio salmonicida | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio vulnificus | 0 | 8 | 12 | 12 | 12 | 12 | 12 | 12 | 12 |
| TEAC (μmolTE/g Extract) | ORAC (μmolTE/g Extract) | Folin–Ciocalteu (mgGAE/g Extract) |
|---|---|---|
| 30.554 ± 2.297 | 170.960 ± 16.830 | 5.867 ± 0.136 |
| Compound | Assignment | δ1H (ppm, Multiplicity) | δ13C (ppm) |
|---|---|---|---|
| CHO | –CH3-18 | 0.68 (s) | 11.67 |
| –CH3-26 | 0.86 | 22.45–22.38 | |
| –CH3-21 | 0.92 (d) | 18.56 | |
| –CH3-19 | 1.01 (s) | 19.06 | |
| All FA (SFA, UFA) | –CH3 | 0.97–1.02 | 14.18 |
| All FA | –(CH2)– COOCH2CH2 | 1.22–1.33 (m) 1.46–1.68 (m) | |
| UFA | CH2=CH2–CH2 | 1.98–2.07 | 27.2 |
| All FA | CH2–C=O | 2.32–2.38 | |
| ARA | CH2–COOH | 2.38 | |
| DHA | CH2–CH2–COOH | 2.38–2.42 | 22.59 |
| 34.07 | |||
| PHB | CH3 | 1.26 | |
| CH | 5.23 | ||
| CH2 | 2.48 (dd) | 40.75 | |
| CH2 | 2.58(dd) | 169.9 | |
| DUFA | CH2 | 2.73–2.78 | 25.6 |
| PUFA ω-3 (DHA, linolenic acid) | CH2 | 2.78–2.86 | |
| MAGs | CHOCO | 3.63 | 70.5 |
| DAGs | OH–CH2–CH | 3.73 (m) | |
| 2’CHOCO | 5.08 (m) | ||
| TGs | CH2 (sn1,3) | 4.15 | 62.01 |
| CH2 (sn1,3) | 4.28 | ||
| CH (sn2) | 5.26 | ||
| All UFAs | CH=CH | 5.30–5.42 (m) | 68.6 |
| Dehydroabietic and abietic acids | CH | 6.88 | 125.57 |
| CH | 7.00 | 127.28 | |
| CH | 7.16 | ||
| Alkaloid species | 7.53 | 130.88 | |
| 7.72 | 128.90 | ||
| Chlorophylls a | CH-20 | 8.55 | |
| CH-5 | 9.54 | ||
| Chlorophylls b | 9.83 | ||
| CH-5 | 9.99 | ||
| 10.04 | |||
| CHO-7 | 11.23 | ||
| 11.25 | |||
| Pheophytin a | CH | 9.35 | |
| 9.40 | |||
| Pheophytin b | CH | 9.60 | |
| 9.62 | |||
| Lutein | CH=CH | 6.13–6.32 | |
| β-carotene | CH=CH | 6.57–6.69 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stabili, L.; Acquaviva, M.I.; Angilè, F.; Cavallo, R.A.; Cecere, E.; Del Coco, L.; Fanizzi, F.P.; Gerardi, C.; Narracci, M.; Petrocelli, A. Screening of Chaetomorpha linum Lipidic Extract as a New Potential Source of Bioactive Compounds. Mar. Drugs 2019, 17, 313. https://doi.org/10.3390/md17060313
Stabili L, Acquaviva MI, Angilè F, Cavallo RA, Cecere E, Del Coco L, Fanizzi FP, Gerardi C, Narracci M, Petrocelli A. Screening of Chaetomorpha linum Lipidic Extract as a New Potential Source of Bioactive Compounds. Marine Drugs. 2019; 17(6):313. https://doi.org/10.3390/md17060313
Chicago/Turabian StyleStabili, Loredana, Maria Immacolata Acquaviva, Federica Angilè, Rosa Anna Cavallo, Ester Cecere, Laura Del Coco, Francesco Paolo Fanizzi, Carmela Gerardi, Marcella Narracci, and Antonella Petrocelli. 2019. "Screening of Chaetomorpha linum Lipidic Extract as a New Potential Source of Bioactive Compounds" Marine Drugs 17, no. 6: 313. https://doi.org/10.3390/md17060313
APA StyleStabili, L., Acquaviva, M. I., Angilè, F., Cavallo, R. A., Cecere, E., Del Coco, L., Fanizzi, F. P., Gerardi, C., Narracci, M., & Petrocelli, A. (2019). Screening of Chaetomorpha linum Lipidic Extract as a New Potential Source of Bioactive Compounds. Marine Drugs, 17(6), 313. https://doi.org/10.3390/md17060313