Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors

 , and
, and
Abstract
1. Introduction
2. Results
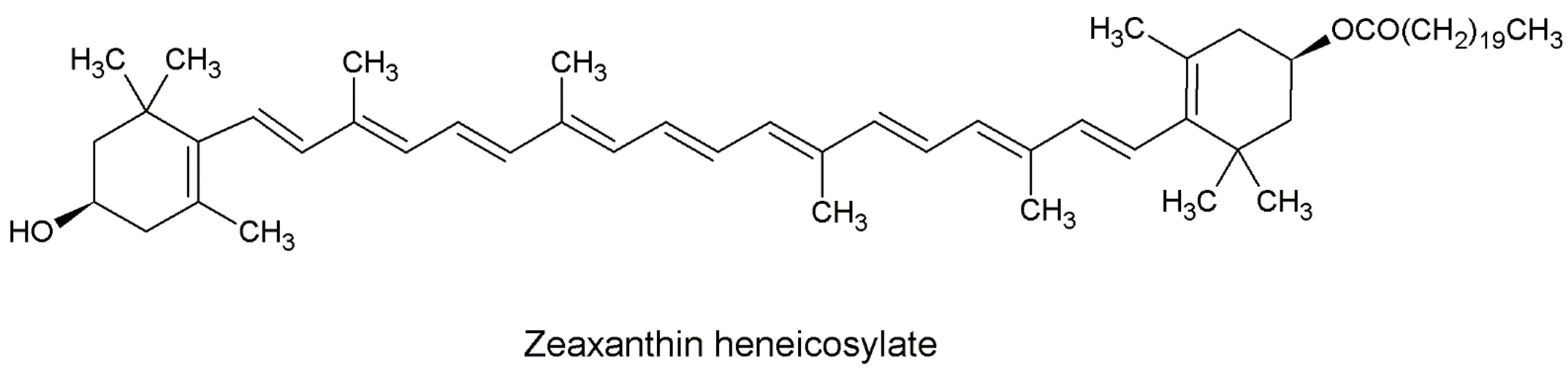
2.1. Isolation and Identification of ZH
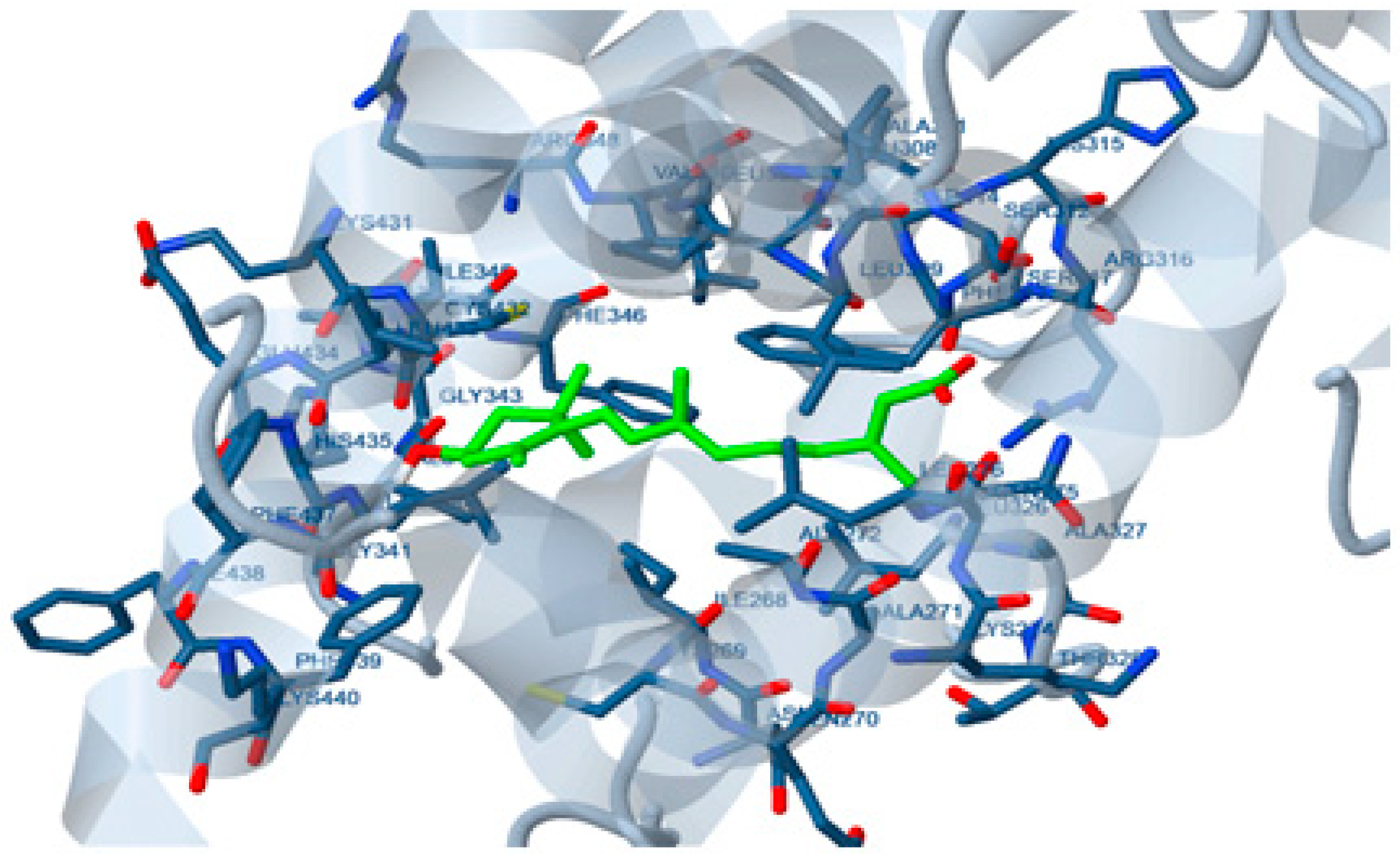
2.2. Molecular Docking Study
2.3. Pharmacological Study
2.3.1. Acute Toxicity Study on Mice
2.3.2. Effect of ZH on Age-Associated Cardiac Dysfunction in Rats
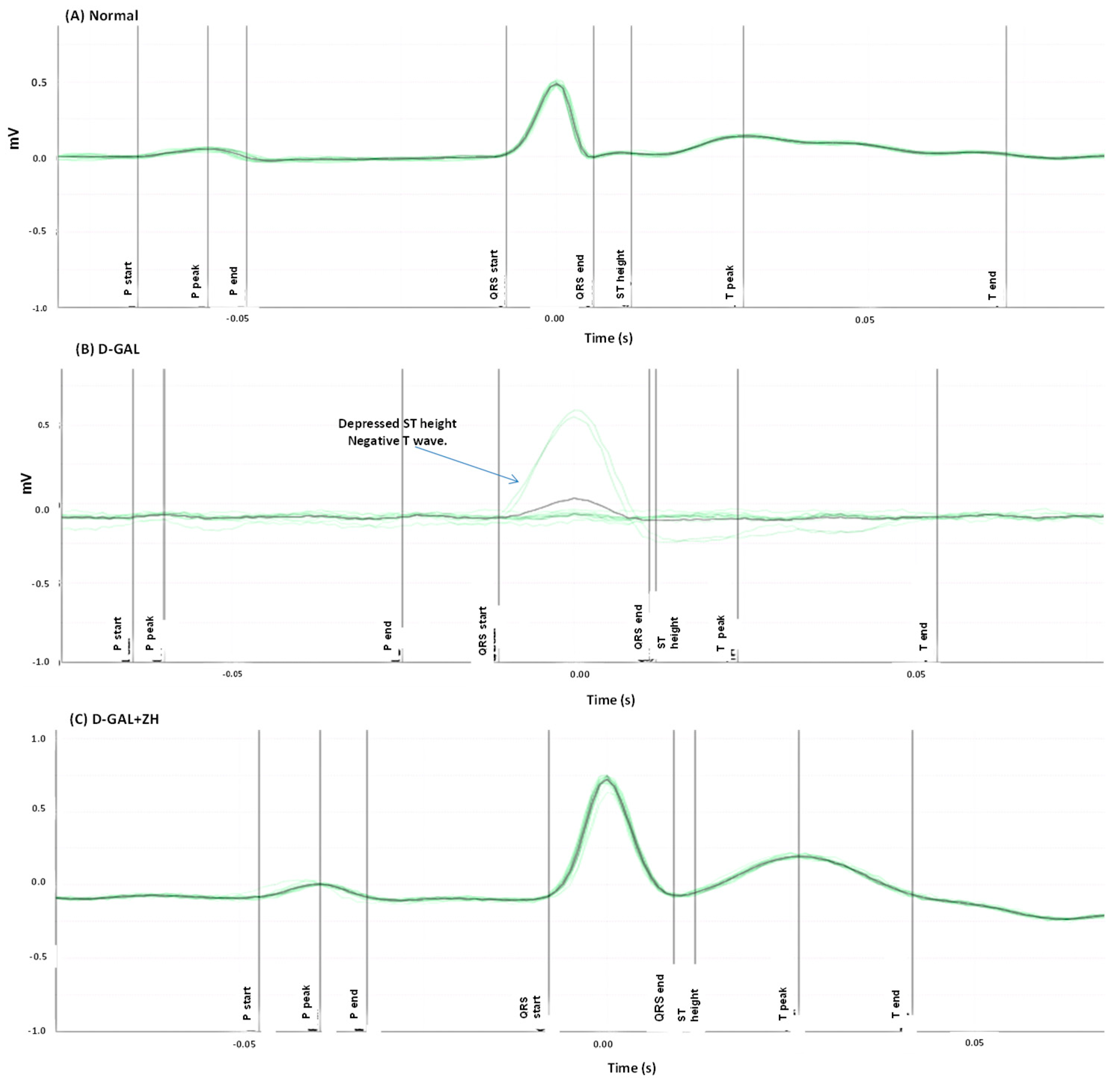
Effect of ZH on the Electrocardiographic (ECG) Measurements
Effect of ZH on Biomarkers Measured in the Serum
Effect of ZH on Biomarkers Measured in the Cardiac Tissue
Effect of ZH on RAR-α Expression
Effect of ZH on the Histopathological Changes
2.3.3. Biochemical and Behavioral Safety Assessment
3. Discussion
4. Materials and Methods
4.1. Cultivation of Dunaliella salina
4.2. Preparation of Algal Extract
4.3. Isolation Purification and Identification ZH
4.4. Docking Study
4.5. Pharmacological Study
4.5.1. Animals
4.5.2. Chemicals
4.5.3. Acute Toxicity Study
4.5.4. Age-associated Cardiac Dysfunction Study
Experimental Design
Electrocardiographic (ECG) Measurements
Serum Collection and Tissue Preparation
Serum Biochemical Analysis
Cardiac Biochemical Analysis
Assessment of Retinoic Acid Receptor-α Expression
Preparation of Sections for Histopathological Examination
4.5.5. Biochemical and Behavioral Safety Study
4.5.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paiva, S.A.; Matsubara, L.S.; Matsubara, B.B.; Minicucci, M.F.; Azevedo, P.S.; Campana, A.O.; Zornoff, L.A. Retinoic acid supplementation attenuates ventricular remodeling after myocardial infarction in rats. J. Nutr. 2005, 135, 2326–2328. [Google Scholar] [CrossRef] [PubMed]
- Bushue, N.; Wan, Y.J. Retinoid pathway and cancer therapeutics. Adv. Drug Deliv. Rev. 2010, 62, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Guleria, R.S.; Nizamutdinova, I.T.; Baker, K.M.; Pan, J. High glucose-induced repression of RAR/RXR in cardiomyocytes is mediated through oxidative stress/JNK signaling. J. Cell. Physiol. 2012, 227, 2632–2644. [Google Scholar] [CrossRef] [PubMed]
- Das, B.C.; Thapa, P.; Karki, R.; Das, S.; Mahapatra, S.; Liu, T.C.; Torregroza, I.; Wallace, D.P.; Kambhampati, S.; Van Veldhuizen, P.; et al. Retinoic acid signaling pathways in development and diseases. Bioorg. Med. Chem. 2014, 22, 673–683. [Google Scholar] [CrossRef]
- Phadwal, K.; Singh, P.K. Isolation and characterization of an indigenous isolate of Dunaliella sp. for beta-carotene and glycerol production from a hypersaline lake in India. J. Basic Microbiol. 2003, 43, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Talero, E.; Garcia-Maurino, S.; Avila-Roman, J.; Rodriguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive Compounds Isolated from Microalgae in Chronic Inflammation and Cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Jaleel, G.; Saleh, D.; Hussein, R. Protective and therapeutic potentials of Dunaliella salina on aging-associated cardiac dysfunction in rats. Asian Pac. J. Trop. Biomed. 2018, 8, 403–410. [Google Scholar]
- El-Baz, F.K.; Hussein, R.A.; Mahmoud, K.; Abdo, S.M. Cytotoxic activity of carotenoid rich fractions from Haematococcus pluvialis and Dunaliella salina microalgae and the identification of the phytoconstituents using LC-DAD/ESI-MS. Phytother. Res. 2018, 32, 298–304. [Google Scholar] [CrossRef]
- El-Baz, F.K.; Aly, H.F.; Fayed, D.B. Dunaliella salina improved obesity-associated inflammation and oxidative damage in animals’ rodent. Asian J. Pharm. Clin. Res. 2018, 11, 240–264. [Google Scholar] [CrossRef]
- Khachik, F.; Englert, G.; Daitch, C.E.; Beecher, G.R.; Tonucci, L.H.; Lusby, W.R. Isolation and structural elucidation of the geometrical isomers of lutein and zeaxanthin in extracts from human plasma. J. Chromatogr. 1992, 582, 153–166. [Google Scholar] [CrossRef]
- Madhavan, J.; Chandrasekharan, S.; Priya, M.K.; Godavarthi, A. Modulatory Effect of Carotenoid Supplement Constituting Lutein and Zeaxanthin (10:1) on Anti-oxidant Enzymes and Macular Pigments Level in Rats. Pharmacog. Mag. 2018, 14, 268–274. [Google Scholar] [CrossRef]
- Bestetti, R.B.; Oliveira, J.S. The surface electrocardiogram: A simple and reliable method for detecting overt and latent heart disease in rats. Br. J. Med. Biol. Res. 1990, 23, 1213–1222. [Google Scholar]
- Rossi, S.; Baruffi, S.; Bertuzzi, A.; Miragoli, M.; Corradi, D.; Maestri, R.; Alinovi, R.; Mutti, A.; Musso, E.; Sgoifo, A.; et al. Ventricular activation is impaired in aged rat hearts. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2336–H2347. [Google Scholar] [CrossRef][Green Version]
- Fannin, J.; Rice, K.M.; Thulluri, S.; Dornon, L.; Arvapalli, R.K.; Wehner, P.; Blough, E.R. Age-associated alterations of cardiac structure and function in the female F344xBN rat heart. Age 2014, 36, 9684. [Google Scholar] [CrossRef]
- Jansen, J.A.; Noorman, M.; Musa, H.; Stein, M.; de Jong, S.; van der Nagel, R.; Hund, T.J.; Mohler, P.J.; Vos, M.A.; van Veen, T.A.; et al. Reduced heterogeneous expression of Cx43 results in decreased Nav1.5 expression and reduced sodium current that accounts for arrhythmia vulnerability in conditional Cx43 knockout mice. Heart Rhythm 2012, 9, 600–607. [Google Scholar] [CrossRef]
- Hamilton, C.A.; Brosnan, M.J.; McIntyre, M.; Graham, D.; Dominiczak, A.F. Superoxide excess in hypertension and aging: A common cause of endothelial dysfunction. Hypertension 2001, 37, 529–534. [Google Scholar] [CrossRef]
- Meador, B.M.; Krzyszton, C.P.; Johnson, R.W.; Huey, K.A. Effects of IL-10 and age on IL-6, IL-1beta, and TNF-alpha responses in mouse skeletal and cardiac muscle to an acute inflammatory insult. J. Appl. Physiol. 2008, 104, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Shah, A.M. Age-associated pro-inflammatory remodeling and functional phenotype in the heart and large arteries. J. Mol. Cell. Cardiol. 2015, 83, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Q.; Yang, C.; Wang, Y.Y.; Wang, P.; Chen, H.L.; Zhang, X.D.; Liu, R.; Li, W.L.; Qin, X.J.; Liang, X.; et al. RAGE upregulation and nuclear factor-kappaB activation associated with ageing rat cardiomyocyte dysfunction. Gen. Physiol. Biophys. 2008, 27, 152–158. [Google Scholar] [PubMed]
- Gupta, S.; Young, D.; Maitra, R.K.; Gupta, A.; Popovic, Z.B.; Yong, S.L.; Mahajan, A.; Wang, Q.; Sen, S. Prevention of cardiac hypertrophy and heart failure by silencing of NF-kappaB. J. Mol. Biol. 2008, 375, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Wang, J.; Xing, F.; Han, T.; Jiao, R.; Liong, E.C.; Fung, M.L.; So, K.F.; Tipoe, G.L. Zeaxanthin dipalmitate therapeutically improves hepatic functions in an alcoholic fatty liver disease model through modulating MAPK pathway. PLoS ONE 2014, 9, e95214. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.A. Zeaxanthin: Review of Toxicological Data and Acceptable Daily Intake. J. Ophthalmol. 2016, 2016, 15. [Google Scholar] [CrossRef]
- Berg, J.; Dingbo, L. Lutein and Zeaxanthin: An overview of metabolism and eye health. J. Human Nutr. Food Sci. 2014, 2, 1048. [Google Scholar]
- Bernstein, P.S.; Khachik, F.; Carvalho, L.S.; Muir, G.J.; Zhao, D.Y.; Katz, N.B. Identification and quantitation of carotenoids and their metabolites in the tissues of the human eye. Exp. Eye Res. 2001, 72, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Palm-Leis, A.; Singh, U.S.; Herbelin, B.S.; Olsovsky, G.D.; Baker, K.M.; Pan, J. Mitogen-activated protein kinases and mitogen-activated protein kinase phosphatases mediate the inhibitory effects of all-trans retinoic acid on the hypertrophic growth of cardiomyocytes. J. Biol. Chem. 2004, 279, 54905–54917. [Google Scholar] [CrossRef] [PubMed]
- Guleria, R.S.; Choudhary, R.; Tanaka, T.; Baker, K.M.; Pan, J. Retinoic acid receptor-mediated signaling protects cardiomyocytes from hyperglycemia induced apoptosis: Role of the renin-angiotensin system. J. Cell. Physiol. 2011, 226, 1292–1307. [Google Scholar] [CrossRef]
- Nizamutdinova, I.T.; Guleria, R.S.; Singh, A.B.; Kendall, J.A., Jr.; Baker, K.M.; Pan, J. Retinoic acid protects cardiomyocytes from high glucose-induced apoptosis through inhibition of NF-kappaB signaling pathway. J. Cell. Physiol. 2013, 228, 380–392. [Google Scholar] [CrossRef]
- Wang, H.J.; Zhu, Y.C.; Yao, T. Effects of all-trans retinoic acid on angiotensin II-induced myocyte hypertrophy. J. Appl. Physiol. 2002, 92, 2162–2168. [Google Scholar] [CrossRef]
- Zhu, S.; Guleria, R.S.; Thomas, C.M.; Roth, A.; Gerilechaogetu, F.; Kumar, R.; Dostal, D.E.; Baker, K.M.; Pan, J. Loss of myocardial retinoic acid receptor alpha induces diastolic dysfunction by promoting intracellular oxidative stress and calcium mishandling in adult mice. J. Mol. Cell. Cardiol. 2016, 99, 100–112. [Google Scholar] [CrossRef]
- Iribarren, C.; Folsom, A.R.; Jacobs, D.R., Jr.; Gross, M.D.; Belcher, J.D.; Eckfeldt, J.H. Association of serum vitamin levels, LDL susceptibility to oxidation, and autoantibodies against MDA-LDL with carotid atherosclerosis. A case-control study. The ARIC Study Investigators. Atherosclerosis Risk in Communities. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T.H. Carotenoids and cardiovascular health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar] [CrossRef]
- Bonds, D.E.; Harrington, M.; Worrall, B.B.; Bertoni, A.G.; Eaton, C.B.; Hsia, J.; Robinson, J.; Clemons, T.E.; Fine, L.J.; Chew, E.Y. Effect of Long-Chain ω-3 Fatty Acids and Lutein + Zeaxanthin Supplements on Cardiovascular Outcomes. JAMA Int. Med. 2014, 174, 763. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar]
- Bikadi, Z.; Hazai, E. Application of the PM6 semi-empirical method to modeling proteins enhances docking accuracy of AutoDock. J. Cheminf. 2009, 1, 1–15. [Google Scholar] [CrossRef]
- Morris, G.; Goodsell, D.; Halliday, R.; Huuey, R.; Belew, R.; Olsen, A. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Solis, F.; Wets, R. Minimization by Random Search Techniques. Math. Oper. Res. 1981, 6, 19–30. [Google Scholar] [CrossRef]
- Liu, J.-P.; Chen, K.-J. Guideline Development Team. Methodology guideline for clinical studies investigating traditional Chinese medicine and integrative medicine: Executive summary. Complement. Ther. Med. 2015, 23, 751–756. [Google Scholar] [PubMed]
- Wurzburg, U.; Hennrich, N.; Lang, H.; Prellwitz, W.; Neumeier, D.; Knedel, M. Determination of creatine kinase-MB in serum using inhibiting antibodies (author’s transl). Klinische Wochenschrift 1976, 54, 357–360. [Google Scholar] [PubMed]
- Buhl, S.N.; Jackson, K.Y. Optimal conditions and comparison of lactate dehydrogenase catalysis of the lactate-to-pyruvate and pyruvate-to-lactate reactions in human serum at 25, 30, and 37 degrees C. Clin. Chem. 1978, 24, 828–831. [Google Scholar]
- Ali, F.K.; El-Shafai, S.A.; Samhan, F.A.; Khalil, W.K.B. Effect of water pollution on expression of immune response genes of Solea aegyptiaca in Lake Qarun. Af. J. Biotechnol. 2008, 7, 1418–1425. [Google Scholar]
- Khalil, W.K.; Booles, H.F. Protective role of selenium against over-expression of cancer-related apoptotic genes induced by O-cresol in rats. Arh. Hig. Rada Toksikol. 2011, 62, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Larsen, K. Creatinine assay by a reaction-kinetic principle. Clin. Chim. Acta 1972, 41, 209–217. [Google Scholar] [CrossRef]
- Fawcett, J.K.; Scott, J.E. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Heart Rate Beat/Min | PR Interval (s) | QRS Interval (s) | ST Height (mV) |
|---|---|---|---|---|
| Normal | 336.0 ± 10.26 | 0.0487 ± 0.0016 | 0.0128 ± 0.0002 | 0.0241 ± 0.002 |
| d-GAL | 422.4 ± 16.56 * | 0.0576 ± 0.0028 * | 0.0353 ± 0.0012 * | −0.0838 ± 0.010 * |
| d-GAL+ ZH | 293.8 ± 14.47 a | 0.0448 ± 0.0020 *,a | 0.0216 ± 0.0021 *,a | −0.0097 ± 0.014 *,a |
| Groups | Serum Homocysteine (U/L) | Serum CK-MB (U/L) | Serum LDH (U/L) | Cardiac GLUT-4 |
|---|---|---|---|---|
| Normal | 3.95 ± 0.34 | 88.18 ± 4.72 | 184.52 ± 143.9 | 10.65 ± 0.37 |
| d-GAL | 10.85 ± 0.16 * | 797.06 ± 53.90 * | 465.42 ± 34.79 * | 4.13 ± 0.35 * |
| d-GAL + ZH | 5.10 ± 0.35 a | 291.40 ± 22.38 a | 267.41 ± 20.63 a | 7.02 ± 0.74 a |
| Groups | Serum ALT (U/mL) | Serum AST (U/mL) | Urea (mg/dL) | Creatinine (mg/dL) |
|---|---|---|---|---|
| Normal | 48.33 ± 0.62 | 88.68 ± 4.08 | 41.74 ± 1.01 | 0.54 ± 0.08 |
| d-GAL | 121.37 ± 10.66 * | 140.7 ± 6.09 * | 54.25 ± 4.4 * | 0.98 ± 0.02 * |
| d-GAL + ZH | 56.21 ± 5.22 a | 91.48 ± 6.88 a | 43.67 ± 0.92 a | 0.59 ± 0.02 a |
| Gene | Forward | Reference (GenBank) |
|---|---|---|
| RAR-α | F: aca act tcc cgc ttt tca cc | BC099830.1 |
| R: ctt tcg tac atc ttg ccc cg | ||
| GAPDH | F:cgg cta cta gcg gtt tta cg | AY340484.1 |
| R:aag aag atg cgg ctg act gt |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Baz, F.K.; Hussein, R.A.; Saleh, D.O.; Abdel Jaleel, G.A.R. Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors. Mar. Drugs 2019, 17, 290. https://doi.org/10.3390/md17050290
El-Baz FK, Hussein RA, Saleh DO, Abdel Jaleel GAR. Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors. Marine Drugs. 2019; 17(5):290. https://doi.org/10.3390/md17050290
Chicago/Turabian StyleEl-Baz, Farouk Kamel, Rehab Ali Hussein, Dalia Osama Saleh, and Gehad Abdel Raheem Abdel Jaleel. 2019. "Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors" Marine Drugs 17, no. 5: 290. https://doi.org/10.3390/md17050290
APA StyleEl-Baz, F. K., Hussein, R. A., Saleh, D. O., & Abdel Jaleel, G. A. R. (2019). Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors. Marine Drugs, 17(5), 290. https://doi.org/10.3390/md17050290