Activation Studies of the γ-Carbonic Anhydrases from the Antarctic Marine Bacteria Pseudoalteromonas haloplanktis and Colwellia psychrerythraea with Amino Acids and Amines

 , , ,
, , ,  and
and 
Abstract
1. Introduction
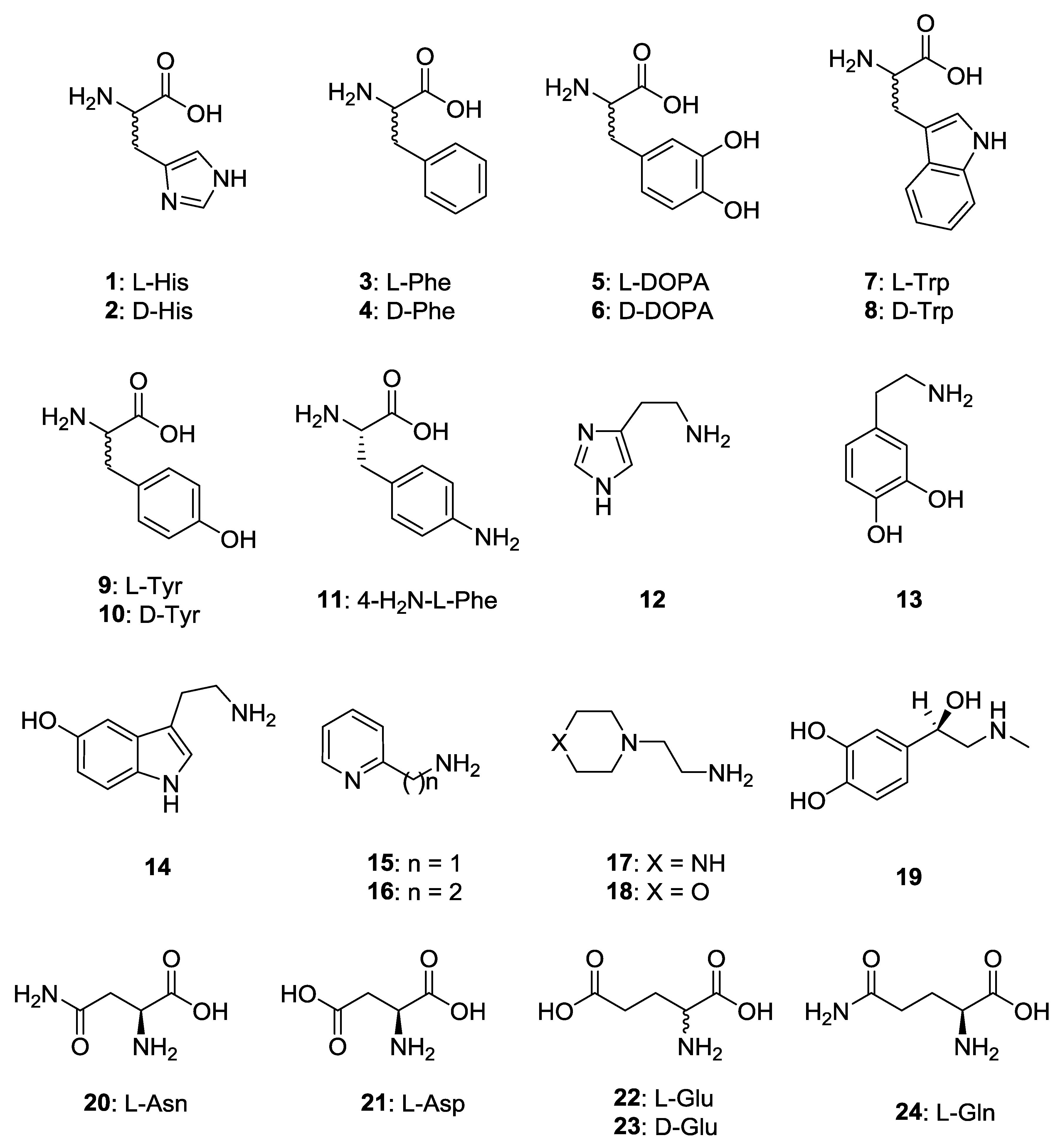
2. Results and Discussion
- (i)
- Firstly, PhaCA is much more sensitive to activation with amines and amino acids 1–24, compared to CpsCA, although both enzymes are prokaryotic, belonging to the same class. Indeed, the activation constants of these amines ranged between 0.72 and 32.4 µM for PhaCA, whereas they were only in the range of 4.79 to 100 µM for CpsCA.
- (ii)
- The most effective PhaCA activators were d-Phe, l-/d-DOPA, l-Tyr and amine 15, which showed KA values ranging between 0.72 and 3.27 µM. Compounds such as d-His, l-Trp, d-Tyr, histamine, dopamine, serotonin and 20–23 were also effective activators, with KA values ranging between 6.48 and 9.85 µM. Medium potency activators were l-His, l-Phe, d-Trp, amines 16–19 and l-Gln, which showed KA values ranging between 10.1 and 32.4 µM (Table 1). Thus, the SAR was rather complex, but generally the d-amino acids were better activators than their l-enantiomers (except for the enantiomer pairs l-/d-Trp and l-/d-Tyr). In some cases, the amines (histamine) were more effective activators than the structurally related amino acids (l- and d-His), whereas in other cases (l-/d-DOPA) the amino acids were more effective CAAs compared to the structurally related amine (dopamine). No major differences were observed between carboxylate/carboxamide derivatives in some cases (l-Asp/l-Asn), whereas for l-Glu, the carboxamide (l-Gln) was more than three times less effective as an activator.
- (iii)
- For CpsCA, l-Phe and l-Gln were devoid of activating effects up to a 100 µM concentration of compound in the assay system. Weak activators were also l-/d-His, d-Trp, dopamine, serotonin, amines 16–19, as well as l-Asn and l-Asp (KA values ranging between 27.3 and 79.8 µM). On the other hand, better activators of CpsCA were d-Phe, d-DOPA, l-Trp, l-/d-Tyr, 4-amino-l-Phe, histamine, 2-pyridyl-methylamine and l-/d-Glu, with KA values ranging between 11.2 and 24.4 µM. The most effective CpsCA activator was l-DOPA, with a KA of 4.79 µM. It is obvious that very small structural changes in the molecule of the activator have drastic effects on the enzyme activating effects. For example, the two enantiomers of DOPA have values of KA which differ by a factor of 2.33, with the l-enantiomer being the most active. However, the amino acid in which one of the two OH moieties of DOPA is missing, l-Tyr, was 4 times less effective compared to DOPA, whereas l-Phe was more than 20 times less effective as an activator of this isoform (the compound lacking both phenolic OH groups present in DOPA).
- (iv)
- As no X-ray crystal structure for adducts of activators with γ-CAs are available so far, we cannot rationalize our SAR data in detail. However, all the observations reported above concur with the fact that these compounds bind within the enzyme active site and facilitate the generation of the nucleophilic zinc hydroxide species of the enzyme.
3. Materials and Methods
CA Activity/Activation Measurements
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mocali, S.; Chiellini, C.; Fabiani, A.; Decuzzi, S.; de Pascale, D.; Parrilli, E.; Tutino, M.L.; Perrin, E.; Bosi, E.; Fondi, M.; et al. Ecology of cold environments: New insights of bacterial metabolic adaptation through an integrated genomic-phenomic approach. Sci. Rep. 2017, 7, 839. [Google Scholar] [CrossRef] [PubMed]
- Fondi, M.; Bosi, E.; Presta, L.; Natoli, D.; Fani, R. Modelling microbial metabolic rewiring during growth in a complex medium. BMC Genom. 2016, 17, 970. [Google Scholar] [CrossRef]
- De Luca, V.; Vullo, D.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, characterization and anion inhibition studies of a new γ-carbonic anhydrase from the Antarctic bacterium Pseudoalteromonas haloplanktis. Bioorg. Med. Chem. 2015, 23, 4405–4409. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the γ-carbonic anhydrase from the Antarctic bacterium Pseudoalteromonas haloplanktis. Bioorg. Med. Chem. Lett. 2015, 25, 3550–3555. [Google Scholar] [CrossRef] [PubMed]
- Czajka, J.J.; Abernathy, M.H.; Benites, V.T.; Baidoo, E.E.K.; Deming, J.W.; Tang, Y.J. Model metabolic strategy for heterotrophic bacteria in the cold ocean based on Colwellia psychrerythraea 34H. Proc. Natl. Acad. Sci. USA 2018, 115, 12507–12512. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the γ-carbonic anhydrase from the Antarctic bacterium Colwellia psychrerythraea. Bioorg. Med. Chem. Lett. 2016, 26, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Vullo, D.; Del Prete, S.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, characterization and anion inhibition studies of a γ-carbonic anhydrase from the Antarctic bacterium Colwellia psychrerythraea. Bioorg. Med. Chem. 2016, 24, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic Anhydrases and Metabolism. Metabolites 2018, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta-and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzyme Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Bua, S.; Haapanen, S.; Kuuslahti, M.; Parkkila, S.; Supuran, C.T. Sulfonamide Inhibition Studies of a New β-Carbonic Anhydrase from the Pathogenic Protozoan Entamoebahistolytica. Int. J. Mol. Sci. 2018, 19, 3946. [Google Scholar] [CrossRef]
- Haapanen, S.; Bua, S.; Kuuslahti, M.; Parkkila, S.; Supuran, C.T. Cloning, Characterization and Anion Inhibition Studies of a β-Carbonic Anhydrase from the Pathogenic Protozoan Entamoeba histolytica. Molecules 2018, 23, 3112. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. An Overview of the Bacterial Carbonic Anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef]
- Neri, D.; Supuran, C.T. Interfering with pH regulation in tumours as a therapeutic strategy. Nat. Rev. Drug Discov. 2011, 10, 767–777. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. New light on bacterial carbonic anhydrases phylogeny based on the analysis of signal peptide sequences. J. Enzyme Inhib. Med. Chem. 2016, 31, 1254–1260. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. Biomedical applications of prokaryotic carbonic anhydrases. Expert Opin. Ther. Pat. 2018, 28, 745–754. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic Anhydrase Inhibition and the Management of Hypoxic Tumors. Metabolites 2017, 7, 48. [Google Scholar] [CrossRef]
- Supuran, C.T. Advances in structure-based drug discovery of carbonic anhydrase inhibitors. Expert Opin. Drug Discov. 2017, 12, 61–88. [Google Scholar] [CrossRef]
- Nishimori, I.; Onishi, S.; Takeuchi, H.; Supuran, C.T. The α and β-Classes Carbonic Anhydrases from Helicobacter pylori as Novel Drug Targets. Curr. Pharm. Des. 2008, 14, 622–630. [Google Scholar]
- da Silva Cardoso, V.; Vermelho, A.B.; Ricci Junior, E.; Almeida Rodrigues, I.; Mazotto, A.M.; Supuran, C.T. Antileishmanial activity of sulphonamidenanoemulsions targeting the β-carbonic anhydrase from Leishmania species. J. Enzyme Inhib. Med. Chem. 2018, 33, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Vermelho, A.B.; da Silva Cardoso, V.; Ricci Junior, E.; Dos Santos, E.P.; Supuran, C.T. Nanoemulsions of sulfonamide carbonic anhydrase inhibitors strongly inhibit the growth of Trypanosomacruzi. J. Enzyme Inhib. Med. Chem. 2018, 33, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. How many carbonic anhydrase inhibition mechanisms exist? J. Enzyme Inhib. Med. Chem. 2016, 31, 345–360. [Google Scholar] [CrossRef]
- Murray, A.B.; Aggarwal, M.; Pinard, M.; Vullo, D.; Patrauchan, M.; Supuran, C.T.; McKenna, R. Structural Mapping of Anion Inhibitors to β-Carbonic Anhydrase psCA3 from Pseudomonas aeruginosa. Chem. Med. Chem. 2018, 13, 2024–2029. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.A.; Ferry, J.G.; Supuran, C.T. Inhibition of the archaeal beta-class (Cab) and gamma-class (Cam) carbonic anhydrases. Curr. Top. Med. Chem. 2007, 7, 901–908. [Google Scholar] [CrossRef]
- Nocentini, A.; Vullo, D.; Del Prete, S.; Osman, S.M.; Alasmary, F.A.S.; AlOthman, Z.; Capasso, C.; Carta, F.; Gratteri, P.; Supuran, C.T. Inhibition of the β-carbonic anhydrase from the dandruff-producing fungus Malasseziaglobosa with monothiocarbamates. J. Enzyme Inhib. Med. Chem. 2017, 32, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; Supuran, C.T. (In)organic anions as carbonic anhydrase inhibitors. J. Inorg. Biochem. 2012, 111, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, A.; Capasso, C.; De Luca, V.; Monti, S.M.; Carginale, V.; Supuran, C.T.; Scozzafava, A.; Pedone, C.; Rossi, M.; De Simone, G. X-ray structure of the first “extremo-α-carbonic anhydrase”, a dimeric enzyme from the thermophilic bacterium Sulfurihydrogenibium yellowstonense YO3AOP1. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, A.; Vullo, D.; De Luca, V.; Scozzafava, A.; Carginale, V.; Rossi, M.; Supuran, C.T.; Capasso, C. The extremo-α-carbonic anhydrase (CA) from Sulfurihydrogenibium azorense, the fastest CA known, is highly activated by amino acids and amines. Bioorg. Med. Chem. Lett. 2013, 23, 1087–1090. [Google Scholar] [CrossRef]
- Vullo, D.; De Luca, V.; Scozzafava, A.; Carginale, V.; Rossi, M.; Supuran, C.T.; Capasso, C. The first activation study of a bacterial carbonic anhydrase (CA). The thermostable α-CA from Sulfurihydrogenibium yellowstonense YO3AOP1 is highly activated by amino acids and amines. Bioorg. Med. Chem. Lett. 2012, 22, 6324–6327. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Applications of carbonic anhydrases inhibitors in renal and central nervous system diseases. Expert Opin. Ther. Pat. 2018, 28, 713–721. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrase inhibitors and their potential in a range of therapeutic areas. Expert Opin. Ther. Pat. 2018, 28, 709–712. [Google Scholar] [CrossRef]
- Nocentini, A.; Supuran, C.T. Carbonic anhydrase inhibitors as antitumor/antimetastaticagents: A patent review (2008–2018). Expert Opin. Ther. Pat. 2018, 28, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Canto de Souza, L.; Provensi, G.; Vullo, D.; Carta, F.; Scozzafava, A.; Costa, A.; Schmidt, S.D.; Passani, M.B.; Supuran, C.T.; Blandina, P. Carbonic anhydrase activation enhances object recognition memory in mice through phosphorylation of the extracellular signal-regulated kinase in the cortex and the hippocampus. Neuropharmacology 2017, 118, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrase activators. Future Med. Chem. 2018, 10, 561–573. [Google Scholar] [CrossRef]
- Angeli, A.; Kuuslahti, M.; Parkkila, S.; Supuran, C.T. Activation studies with amines and amino acids of the α-carbonic anhydrase from the pathogenic protozoan Trypanosoma cruzi. Bioorg. Med. Chem. 2018, 26, 4187–4190. [Google Scholar] [CrossRef]
- Angeli, A.; Del Prete, S.; Alasmary, F.A.S.; Alqahtani, L.S.; AlOthman, Z.; Donald, W.A.; Capasso, C.; Supuran, C.T. The first activation studies of the η-carbonic anhydrase from the malaria parasite Plasmodium falciparum with amines and amino acids. Bioorg. Chem. 2018, 80, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Stefanucci, A.; Angeli, A.; Dimmito, M.P.; Luisi, G.; Del Prete, S.; Capasso, C.; Donald, W.A.; Mollica, A.; Supuran, C.T. Activation of β- and γ-carbonic anhydrases from pathogenic bacteria with tripeptides. J. Enzyme Inhib. Med. Chem. 2018, 33, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Angeli, A.; Alasmary, F.A.S.; Del Prete, S.; Osman, S.M.; AlOthman, Z.; Donald, W.A.; Capasso, C.; Supuran, C.T. The first activation study of a δ-carbonic anhydrase: TweCAδ from the diatom Thalassiosiraweissflogii is effectively activated by amines and amino acids. J. Enzyme Inhib. Med. Chem. 2018, 33, 680–685. [Google Scholar] [CrossRef]
- Briganti, F.; Mangani, S.; Orioli, P.; Scozzafava, A.; Vernaglione, G.; Supuran, C.T. Carbonic anhydrase activators: X-ray crystallographic and spectroscopic investigations for the interaction of isozymes I and II with histamine. Biochemistry 1997, 36, 10384–10392. [Google Scholar] [CrossRef]
- Clare, B.W.; Supuran, C.T. Carbonic anhydrase activators. 3: Structure-activity correlations for a series of isozyme II activators. J. Pharm. Sci. 1994, 83, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [PubMed]
- Temperini, C.; Scozzafava, A.; Vullo, D.; Supuran, C.T. Carbonic anhydrase activators. Activation of isoforms I, II, IV, VA, VII, and XIV with l- and d-phenylalanine and crystallographic analysis of their adducts with isozymeII: Stereospecific recognition within the active site of an enzyme and its consequences for the drug design. J. Med. Chem. 2006, 49, 3019–3027. [Google Scholar] [PubMed]
- Angeli, A.; Donald, W.A.; Parkkila, S.; Supuran, C.T. Activation studies with amines and amino acids of the β-carbonic anhydrase from the pathogenic protozoan Leishmania donovanichagasi. Bioorg. Chem. 2018, 78, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Rami, M.; Winum, J.Y.; Supuran, C.T.; Melnyk, P.; Yous, S. (Hetero)aryl substituted thiazol-2,4-yl scaffold as human carbonic anhydrase I, II, VII and XIV activators. J. Enzyme Inhib. Med. Chem. 2019, 34, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.H.; Tran, T.N.; Podgorski, M.N.; Bell, S.G.; Supuran, C.T.; Donald, W.A. Nanoscale ion emitters in native mass spectrometry for measuring ligand-protein binding affinities. ACS Cent. Sci. 2019, 5, 308–318. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Isozyme | kcat 1 (s−1) | KM 1 (mM) | (kcat) l-Trp 2 (s−1) | KA 3 (μM) l-Trp |
|---|---|---|---|---|
| hCA I a | 2.0 × 105 | 4.0 | 3.4 × 105 | 44.0 |
| hCA II a | 1.4 × 106 | 9.3 | 4.9 × 106 | 27.0 |
| PhaCA b | 1.4 × 105 | 7.3 | 7.6 × 105 | 7.12 |
| CpsCA b | 6.0 × 105 | 12.7 | 9.9 × 105 | 21.3 |
| No. | Compound | KA (μM) * | |||
|---|---|---|---|---|---|
| hCA I a | hCA II a | PhaCA b | CpsCA b | ||
| 1 | l-His | 0.03 | 10.9 | 12.6 | 47.5 |
| 2 | d-His | 0.09 | 43 | 9.41 | 35.9 |
| 3 | l-Phe | 0.07 | 0.013 | 15.8 | >100 |
| 4 | d-Phe | 86 | 0.035 | 3.19 | 15.4 |
| 5 | l-DOPA | 3.1 | 11.4 | 1.08 | 4.79 |
| 6 | d-DOPA | 4.9 | 7.8 | 0.72 | 11.2 |
| 7 | l-Trp | 44 | 27 | 7.12 | 21.3 |
| 8 | d-Trp | 41 | 12 | 13.9 | 36.8 |
| 9 | l-Tyr | 0.02 | 0.011 | 1.02 | 19.5 |
| 10 | d-Tyr | 0.04 | 0.013 | 7.35 | 18.4 |
| 11 | 4-H2N-l-Phe | 0.24 | 0.15 | 3.27 | 17.2 |
| 12 | Histamine | 2.1 | 125 | 6.48 | 20.6 |
| 13 | Dopamine | 13.5 | 9.2 | 8.70 | 32.1 |
| 14 | Serotonin | 45 | 50 | 9.05 | 34.8 |
| 15 | 2-Pyridyl-methylamine | 26 | 34 | 2.39 | 21.5 |
| 16 | 2-(2-Aminoethyl)pyridine | 13 | 15 | 18.7 | 38.2 |
| 17 | 1-(2-Aminoethyl)-piperazine | 7.4 | 2.3 | 15.1 | 33.0 |
| 18 | 4-(2-Aminoethyl)-morpholine | 0.14 | 0.19 | 10.1 | 34.3 |
| 19 | l-Adrenaline | 0.09 | 96.0 | 17.5 | 79.8 |
| 20 | l-Asn | 11.3 | >100 | 9.80 | 27.9 |
| 21 | l-Asp | 5.20 | >100 | 9.85 | 27.3 |
| 22 | l-Glu | 6.43 | >100 | 9.01 | 24.4 |
| 23 | d-Glu | 10.7 | >100 | 4.72 | 12.0 |
| 24 | l-Gln | >100 | >50 | 32.4 | >100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angeli, A.; Del Prete, S.; Osman, S.M.; AlOthman, Z.; Donald, W.A.; Capasso, C.; Supuran, C.T. Activation Studies of the γ-Carbonic Anhydrases from the Antarctic Marine Bacteria Pseudoalteromonas haloplanktis and Colwellia psychrerythraea with Amino Acids and Amines. Mar. Drugs 2019, 17, 238. https://doi.org/10.3390/md17040238
Angeli A, Del Prete S, Osman SM, AlOthman Z, Donald WA, Capasso C, Supuran CT. Activation Studies of the γ-Carbonic Anhydrases from the Antarctic Marine Bacteria Pseudoalteromonas haloplanktis and Colwellia psychrerythraea with Amino Acids and Amines. Marine Drugs. 2019; 17(4):238. https://doi.org/10.3390/md17040238
Chicago/Turabian StyleAngeli, Andrea, Sonia Del Prete, Sameh M. Osman, Zeid AlOthman, William A. Donald, Clemente Capasso, and Claudiu T. Supuran. 2019. "Activation Studies of the γ-Carbonic Anhydrases from the Antarctic Marine Bacteria Pseudoalteromonas haloplanktis and Colwellia psychrerythraea with Amino Acids and Amines" Marine Drugs 17, no. 4: 238. https://doi.org/10.3390/md17040238
APA StyleAngeli, A., Del Prete, S., Osman, S. M., AlOthman, Z., Donald, W. A., Capasso, C., & Supuran, C. T. (2019). Activation Studies of the γ-Carbonic Anhydrases from the Antarctic Marine Bacteria Pseudoalteromonas haloplanktis and Colwellia psychrerythraea with Amino Acids and Amines. Marine Drugs, 17(4), 238. https://doi.org/10.3390/md17040238