Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva rigida, Gracilaria sp. and Fucus vesiculosus




Abstract
1. Introduction
2. Results and Discussion
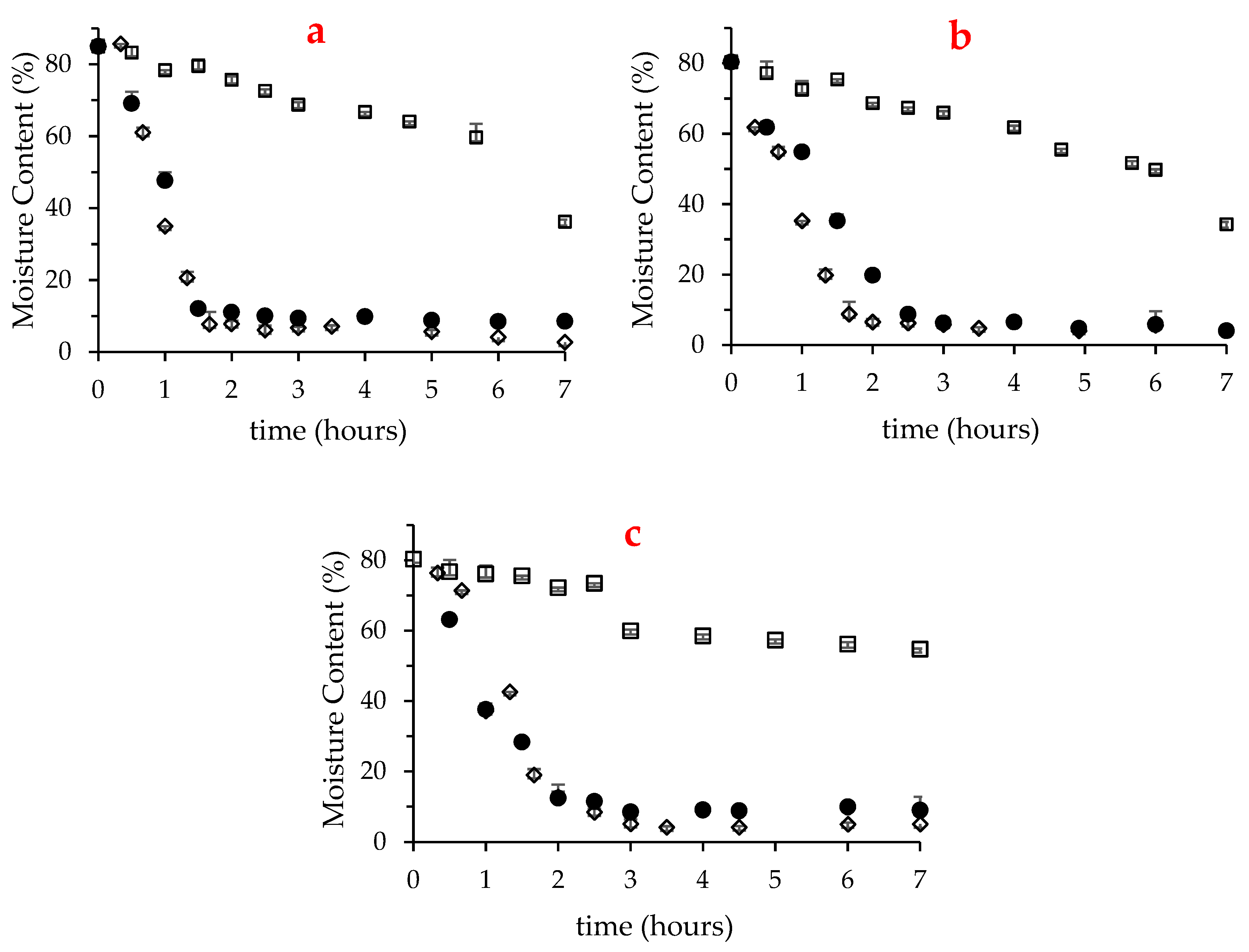
2.1. Moisture Content
2.2. Surface Color
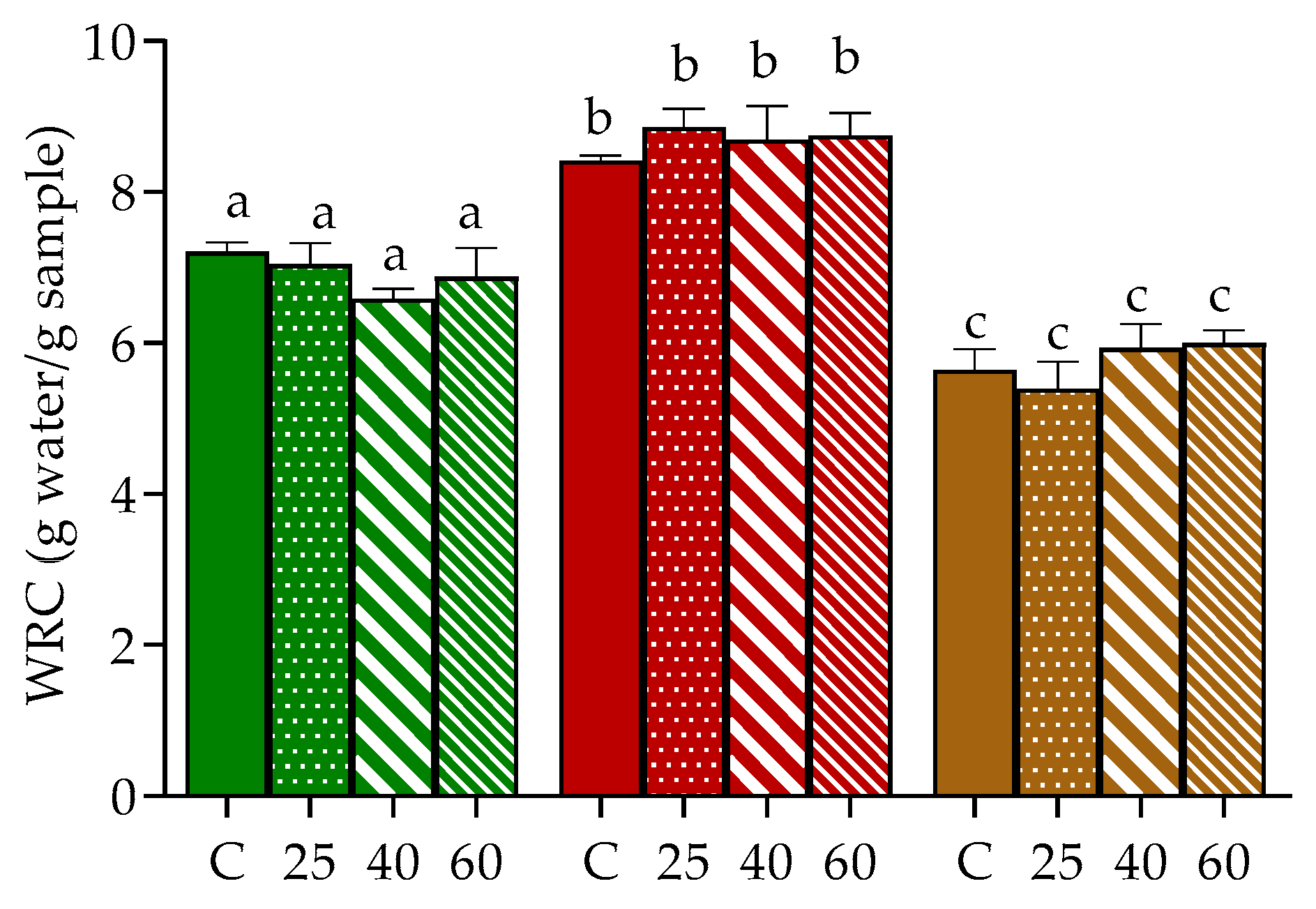
2.3. Water Retention Capacity (WRC)
2.4. Extractability of Valuable Compounds
2.4.1. Pigments
2.4.2. Phenolic Compounds and Antioxidant Activity
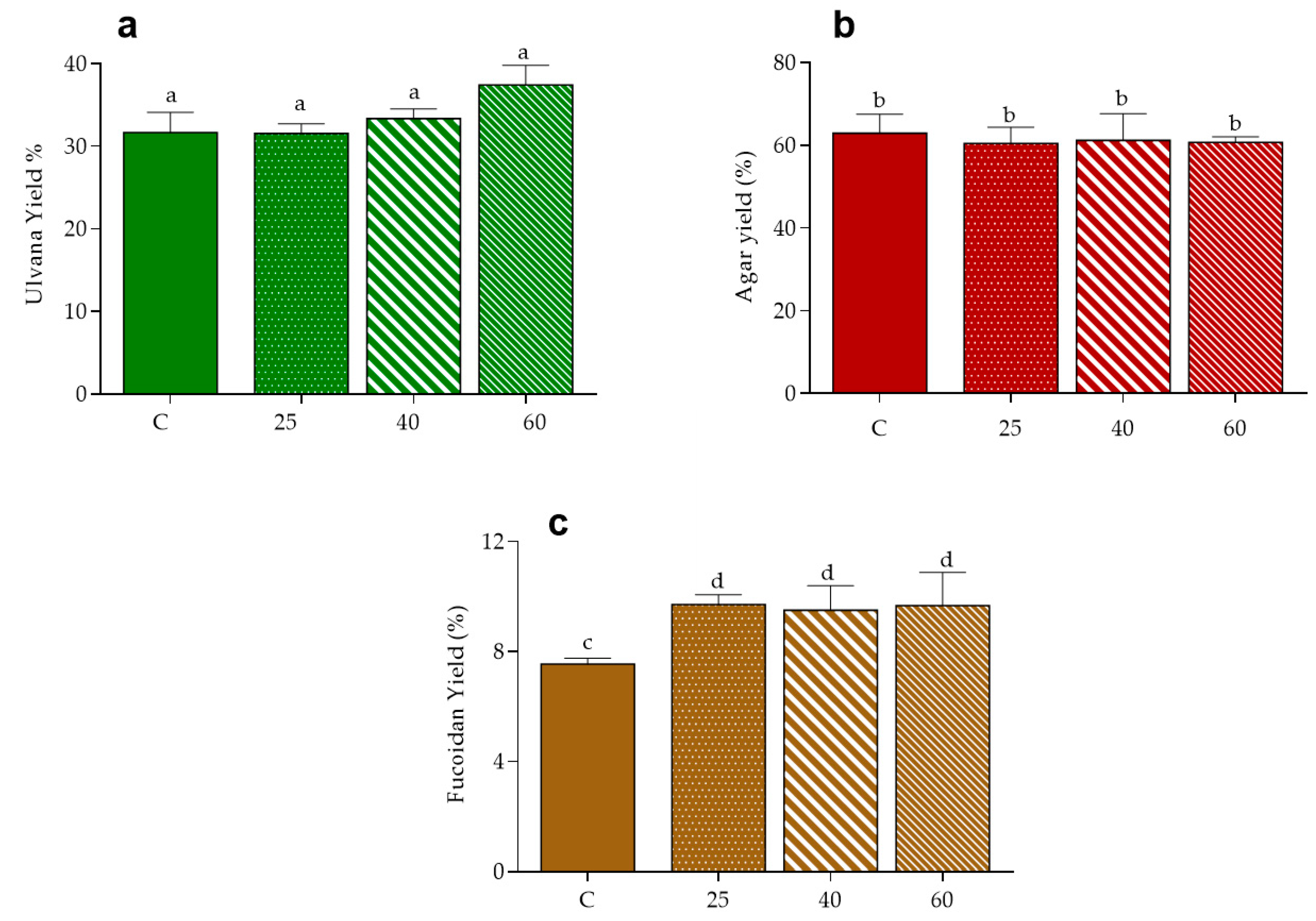
2.5. Polysaccharides
3. Materials and Methods
3.1. Chemicals
3.2. Sample Collection and Drying Processing
3.3. Surface Color and Browning Index
3.4. Water Retention Capacity (WRC)
3.5. Extration and Quantification of Pigments
3.5.1. Chlorophylls, Carotenoides and Fucoxantin
3.5.2. Phycobiliproteins
3.6. Obtaining of Methanolic Extracts
3.6.1. Total Phenolics Compounds (TPC)
3.6.2. Antioxidant Activity
3.7. Polysaccharides Extraction
3.7.1. Ulvans
3.7.2. Agar
3.7.3. Fucoidan
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Delaney, A.; Frangoudes, K.; Ii, S.-A. Society and Seaweed. In Seaweed in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2016; Volume 2, pp. 7–40. ISBN 9780128027936. [Google Scholar]
- Cardoso, S.; Carvalho, L.; Silva, P.; Rodrigues, M.; Pereira, O.; Pereira, L. Bioproducts from Seaweeds: A Review with Special Focus on the Iberian Peninsula. Curr. Org. Chem. 2014, 18, 896–917. [Google Scholar] [CrossRef]
- Kraan, S. Algal Polysaccharides, Novel Applications and Outlook. In Carbohydrates—Comprehensive Studies on Glycobiology and Glycotechnology; InTech: London, UK, 2012; Volume 2, p. 64. ISBN 9789537619992. [Google Scholar]
- Higashimura, Y.; Naito, Y.; Takagi, T.; Mizushima, K.; Hirai, Y.; Harusato, A.; Ohnogi, H.; Yamaji, R.; Inui, H.; Nakano, Y.; et al. Oligosaccharides from agar inhibit murine intestinal inflammation through the induction of heme oxygenase-1 expression. J. Gastroenterol. 2013, 48, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Sousa, R.A.; Reis, R.L. A practical perspective on ulvan extracted from green algae. J. Appl. Phycol. 2013, 25, 407–424. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, M.; Inizan, F.; Vigoureux, J. NMR analysis of the chemical structure of ulvan and of ulvan-boron complex formation. Carbohydr. Polym. 1998, 36, 239–249. [Google Scholar] [CrossRef]
- De Agostini, A.I.; Dong, J.-C.; de Vantéry Arrighi, C.; Ramus, M.-A.; Dentand-Quadri, I.; Thalmann, S.; Ventura, P.; Ibecheole, V.; Monge, F.; Fischer, A.-M.; et al. Human Follicular Fluid Heparan Sulfate Contains Abundant 3-O-Sulfated Chains with Anticoagulant Activity. J. Biol. Chem. 2008, 283, 28115–28124. [Google Scholar] [CrossRef]
- Nagaoka, M.; Shibata, H.; Kimura, I.; Hashimoto, S. Oligosaccharide Derivates and Process for Producing the Same. US Patent 6645940, 11 November 2003. [Google Scholar]
- Ma, L.; Lin, X.-M. Effects of lutein and zeaxanthin on aspects of eye health. J. Sci. Food Agric. 2010, 90, 2–12. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Biochim. Biophys. Acta Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef]
- Lorbeer, A.J.; Tham, R.; Zhang, W. Potential products from the highly diverse and endemic macroalgae of Southern Australia and pathways for their sustainable production. J. Appl. Phycol. 2013, 25, 717–732. [Google Scholar] [CrossRef]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Fucaceae: A Source of Bioactive Phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.C.C.; Cheung, P.C.K.; Ang, P.O. Comparative studies on the effect of three drying methods on the nutritional composition of seaweed Sargassum hemiphyllum (Turn) C Ag. J. Agric. Food Chem. 1997, 45, 3056–3059. [Google Scholar] [CrossRef]
- Abreu, M.; Pereira, R.; Sassi, J. Marine Algae and the Global Food Industry. In Marine Algae; CRC Press: Boca Raton, FL, USA, 2014; pp. 300–319. ISBN 978-1-4665-8167-8. [Google Scholar]
- Jangam, S.V.; Law, C.L.; Mujumdar, A.S. Classification and Selection of Dryers for Foods; Singapore Contents: Singapore, 2010; ISBN 9789810867591. [Google Scholar]
- Chen, X.D. Food drying fundamentals. In Drying Technologies in Food Processing; Blackwell: Hoboken, NJ, USA, 2008; p. 350. ISBN 978-1-4051-5763-6. [Google Scholar]
- Robic, A.; Sassi, J.-F.; Lahaye, M. Impact of stabilization treatments of the green seaweed Ulva rotundata (Chlorophyta) on the extraction yield, the physico-chemical and rheological properties of ulvan. Carbohydr. Polym. 2008, 74, 344–352. [Google Scholar] [CrossRef]
- Rodrigues, S.; Afonso, C.; Mouga, T.; Nunes, P. Effects of different drying conditions on the rehydration ratio and water holding capacity properties in three different species of algae Ulva lactuca, Codium vermilara and Codium tomentosum. Front. Mar. Sci. 2014, 1. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; García, V.; Pastén, A.; López, J.; Goñi, G. Effect of different drying methods on phytochemical content and amino acid and fatty acid profiles of the green seaweed, Ulva spp. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef]
- Tello-Ireland, C.; Lemus-Mondaca, R.; Vega-Gálvez, A.; López, J.; Di Scala, K. Influence of hot-air temperature on drying kinetics, functional properties, colour, phycobiliproteins, antioxidant capacity, texture and agar yield of alga Gracilaria chilensis. LWT Food Sci. Technol. 2011, 44, 2112–2118. [Google Scholar] [CrossRef]
- Moreira, R.; Chenlo, F.; Sineiro, J.; Arufe, S.; Sexto, S. Drying temperature effect on powder physical properties and aqueous extract characteristics of Fucus vesiculosus. J. Appl. Phycol. 2016, 28, 2485–2494. [Google Scholar] [CrossRef]
- Gupta, S.; Cox, S.; Abu-Ghannam, N. Effect of different drying temperatures on the moisture and phytochemical constituents of edible Irish brown seaweed. LWT Food Sci. Technol. 2011, 44, 1266–1272. [Google Scholar] [CrossRef]
- Perera, C. Selected Quality Attributes of Dried Foods. Dry. Technol. 2005, 23, 717–730. [Google Scholar] [CrossRef]
- Bonazzi, C.; Dumoulin, E. Modern Drying Technology; Tsotsas, E., Mujumdar, A.S., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2011; ISBN 9783527631667. [Google Scholar]
- Kumar, V.; Fotedar, R. Agar extraction process for Gracilaria cliftonii (Withell, Millar, & Kraft, 1994). Carbohydr. Polym. 2009, 78, 813–819. [Google Scholar] [CrossRef]
- Kursar, T.A; van der Meer, J.; Alberte, R.S. Light-Harvesting System of the Red Alga Gracilaria tikvahiae: II. Phycobilisome Characteristics of Pigment Mutants. Plant Physiol. 1983, 73, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Masson, M.L. Instrumental color and sensory acceptance of soy-based emulsions: A response surface approach. Ciência Tecnol. Aliment. 2010, 30, 1090–1096. [Google Scholar] [CrossRef]
- Larrosa, A.P.Q.; Comitre, A.A.; Vaz, L.B.; Pinto, L.A.A. Influence of air temperature on characterisistics and bioactive compounds in vaccum drying of Arthrospira spirulina. J. Food Process Eng. 2017, 1–9. [Google Scholar] [CrossRef]
- Assawarachan, R.; Nookong, M.; Chailungka, N.; Amornlerdpison, D. Effects of microwave power on the drying characteristics, color and phenolic content of Spirogyra sp. J. Food Agric. Environ. 2013, 11, 15–18. [Google Scholar] [CrossRef]
- Pathare, P.B.; Opara, U.L.; Al-Said, F.A.J. Colour Measurement and Analysis in Fresh and Processed Foods: A Review. Food Bioprocess Technol. 2013, 6, 36–60. [Google Scholar] [CrossRef]
- Benjama, O.; Masniyom, P. Nutritional composition and physicochemical properties of two green seaweeds (Ulva pertusa and U. intestinalis) from the Pattani Bay in Southern Thailand. Songklanakarin J. Sci. Technol. 2011, 33, 575–583. [Google Scholar]
- Garau, M.C.; Simal, S.; Femenia, A.; Rosselló, C. Drying of orange skin: Drying kinetics modelling and functional properties. J. Food Eng. 2006, 75, 288–295. [Google Scholar] [CrossRef]
- Neto, R.; Marçal, C.; Queirós, A.; Abreu, H.; Silva, A.; Cardoso, S. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef] [PubMed]
- Saati, E.A. The Solvent Effectiveness on Extraction Process. Makara J. Technol. 2011, 15, 5–8. [Google Scholar]
- Henriques, M.; Silva, A.; Rocha, J. Extraction and quantification of pigments from a marine microalga: A simple and reproducible method. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. 2007, 586–593. [Google Scholar]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Tiwari, B.K.; Troy, D. Seaweed minor constituents. Seaweed Sustain. Food Non-Food Appl. 2015, 193–242. [Google Scholar] [CrossRef]
- Patel, A.; Pawar, R.; Mishra, S.; Sonawane, S.; Ghosh, P.K. Kinetic studies on thermal denaturation of C-phycocyanin. Indian J. Biochem. Biophys. 2004, 41, 254–257. [Google Scholar] [PubMed]
- Shafiur Rahman, M. Drying of Fish and Seafood. In Handbook of Industrial Drying, Third Edition; CRC Press: Boca Raton, FL, USA, 2006; ISBN 978-1-57444-668-5. [Google Scholar]
- Chinnadurai, S.; Karthik, G.; Chermapandi, P.; Hemalatha, A.; Anantharaman. Estimation of Major Pigment Content in Seaweeds Collected From Pondicherry Coast. Experiment 2013, 9, 522–525. [Google Scholar]
- Stévant, P.; Indergård, E.; Ólafsdóttir, A.; Marfaing, H.; Larssen, W.E.; Fleurence, J.; Roleda, M.Y.; Rustad, T.; Slizyte, R.; Nordtvedt, T.S. Effects of drying on the nutrient content and physico-chemical and sensory characteristics of the edible kelp Saccharina latissima. J. Appl. Phycol. 2018, 30, 2587–2599. [Google Scholar] [CrossRef]
- Ismail, N.; Abdullah, A.; Rowi, S.; Zaidi Jusoh, A. Effect of drying temperature on the content of fucoxanthin, phenolic and antioxidant activity of Malaysian brown seaweed, Sargassum sp. J. Trop. Agric. Food Sci. 2017, 45, 25–36. [Google Scholar]
- Cruces, E.; Rojas-Lillo, Y.; Ramirez-Kushel, E.; Atala, E.; López-Alarcón, C.; Lissi, E.; Gómez, I. Comparison of different techniques for the preservation and extraction of phlorotannins in the kelp Lessonia spicata (Phaeophyceae): Assays of DPPH, ORAC-PGR, and ORAC-FL as testing methods. J. Appl. Phycol. 2016, 28, 573–580. [Google Scholar] [CrossRef]
- Hernández-Carmona, G.; Freile-Pelegrín, Y.; Hernández-Garibay, E. Conventional and alternative technologies for the extraction of algal polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Boca Raton, FL, USA, 2013; pp. 475–516. ISBN 9780857098689. [Google Scholar]
- Lahaye, M.; Robic, A. Structure and function properties of Ulvan, a polysaccharide from green seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef]
- Villanueva, R.D.; Sousa, A.M.M.; Gonçalves, M.P.; Nilsson, M.; Hilliou, L. Production and properties of agar from the invasive marine alga, Gracilaria vermiculophylla (Gracilariales, Rhodophyta). J. Appl. Phycol. 2010, 22, 211–220. [Google Scholar] [CrossRef]
- Cefola, M.; D’Antuono, I.; Pace, B.; Calabrese, N.; Carito, A.; Linsalata, V.; Cardinali, A. Biochemical relationships and browning index for assessing the storage suitability of artichoke genotypes. Food Res. Int. 2012, 48, 397–403. [Google Scholar] [CrossRef]
- Martins, M.; Vieira, F.A.; Correia, I.; Ferreira, R.A.S.; Abreu, H.; Coutinho, J.A.P.; Ventura, S.P.M. Recovery of phycobiliproteins from the red macroalga Gracilaria sp. using ionic liquid aqueous solutions. Green Chem. 2016, 18, 4287–4296. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef] [PubMed]
- Saada, M.; Falleh, H.; Catarino, M.; Cardoso, S.; Ksouri, R. Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb. Molecules 2018, 23, 2177. [Google Scholar] [CrossRef] [PubMed]
- Pankiewicz, R.; Łęska, B.; Messyasz, B.; Fabrowska, J.; Sołoducha, M.; Pikosz, M. First isolation of polysaccharidic ulvans from the cell walls of freshwater algae. Algal Res. 2016, 19, 348–354. [Google Scholar] [CrossRef]
- Wang, C.Y.; Chen, Y.C. Extraction and characterization of fucoidan from six brown macroalgae. J. Mar. Sci. Technol. 2016, 24, 319–328. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Drying Condition | CIELAB | U. rigida | Gracilaria sp. | F. vesiculosus |
|---|---|---|---|---|
| Fresh | a* | −10.18 ± 0.52 | 4.86 ± 0.47 d | 1.12 ± 0.33 f |
| b* | 43.65 ± 0.98 a | 3.77 ± 0.50 d | 2.56 ± 1.08 f | |
| L* | 72.18 ± 1.13 a | 14.32 ± 0.91 d | 21.00 ± 1.24 f | |
| BI | 65.17 | 70.94 | 18.65 | |
| ΔE* | - | - | - | |
| 25 °C | a* | −11.86 ± 0.37 a | 9.73 ± 0.80 e | 0.65 ± 0.25 f |
| b* | 44.89 ± 0.90 a | 5.16 ± 0.81 e | 2.27 ± 0.65 f | |
| L* | 72.07 ± 1.16ab | 18.52 ± 0.84 e | 21.46 ± 0.79 f | |
| BI | 65.11 | 96.20 | 14.25 | |
| ΔE* | 2.09 | 6.59 | 0.72 | |
| 40 °C | a* | −11.60 ± 0.25 a | 9.09 ± 1.71 e | 0.91 ± 0.40 f |
| b* | 47.20 ± 1.74 b | 4.03 ± 0.61 de | 3.99 ± 0.94 g | |
| L* | 70.20 ± 1.19 b | 18.65 ± 1.46 e | 23.15 ± 1.05 g | |
| BI | 75.16 | 81.21 | 23.04 | |
| ΔE* | 4.30 | 6.05 | 2.59 | |
| 60 °C | a* | −13.57 ± 0.34 b | 10.09 ± 0.71 f | 1.46 ± 0.50 f |
| b* | 50.61 ± 0.64 c | 4.72 ± 0.46 de | 7.24 ± 0.99 h | |
| L* | 67.28 ± 1.99 c | 18.49 ± 1.88 e | 25.41 ± 1.12 h | |
| BI | 87.15 | 94.60 | 39.76 | |
| ΔE* | 9.17 | 6.76 | 6.43 |
| Seaweed | Drying Condition | Chlorophylls | Carotenoids | |||
|---|---|---|---|---|---|---|
| Chl a | Chl b | Lut | ||||
| U. rigida | C | 2.66 ± 0.11 a | 0.88 ± 0.08 a | 0.42 ± 0.06 a | ||
| 25 | 4.55 ± 0.31 b | 1.21 ± 0.10 b | 1.20 ± 0.09 b | |||
| 40 | 2.58 ± 0.19 c | 0.68 ± 0.01 a | 0.66 ± 0.02 a | |||
| 60 | 2.67 ± 0.14 c | 0.83 ± 0.03 ab | 0.78 ± 0.06 a | |||
| Chl a | Phe a | Lut | βCart | Fucx | ||
| F. vesiculosus | C | 1.06 ± 0.07 d | 2.41 ± 0.11 d | 0.18 ± 0.02 d | 0.47 ± 0.04 d | 0.85 ± 0.06 d |
| 25 | 0.24 ± 0.02 e | 2.50 ± 0.28 d | 0.03 ± 0.00 d | 0.24 ± 0.01 d | 0.78 ± 0.06 d | |
| 40 | 0.71 ± 0.03 f | 2.96 ± 0.25 e | 0.15 ± 0.02 d | 0.55 ± 0.05 d | 1.79 ± 0.08 c | |
| 60 | 0.35 ± 0.25 ef | 2.99 ± 0.03 e | 0.07 ± 0.00 d | 0.35 ± 0.02 d | 0.91 ± 0.05 d | |
| Chlorophylls | Phycobiliprotein | |||||
| Chl a | PE | PC | APC | |||
| Gracilaria sp. | C | 0.34 ± 0.11 gh | 1.37 ± 0.09 g | 0.45 ± 0.02 g | 0.34 ± 0.02 g | |
| 25 | 0.82 ± 0.01 h | 2.37 ± 0.19 h | 0.69 ± 0.09 g | 0.45 ± 0.06 g | ||
| 40 | 0.14 ± 0.06 g | 1.47 ± 0.13 g | 0.42 ± 0.06 h | 0.46 ± 0.08 g | ||
| 60 | 0.30 ± 0.02 g | 1.19 ± 0.13 g | 0.51 ± 0.19 h | 0.32 ± 0.08 g | ||
| Drying Condition | U. rigida | Gracilaria sp. | F. vesiculosus | |||
|---|---|---|---|---|---|---|
| TPC | IC50 | TPC | IC50 | TPC | IC50 | |
| C | 0.93 ± 0.13 a | 0.47 ± 0.15 a | 1.39 ± 0.48 b | 0.80 ± 0.09 b | 11.0 ± 0.28 d | 0.06 ± 0.01 d |
| 25 | 1.24 ± 0.31 a | 0.34 ± 0.17 a | 1.46 ± 0.42 b | 0.79 ± 0.13 b | 6.72 ± 1.41 e | 0.09 ± 0.02 d |
| 40 | 0.96 ± 0.32 a | 0.43 ± 0.08 a | 1.24 ± 0.43 b | 1.70 ± 0.53 c | 7.28 ± 1.35 e | 0.09 ± 0.01 d |
| 60 | 0.74 ± 0.13 a | 0.31 ± 0.06 a | 1.11 ± 0.34 b | 1.89 ± 0.72 c | 7.20 ± 0.45 e | 0.09 ± 0.06 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.F.R.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva rigida, Gracilaria sp. and Fucus vesiculosus. Mar. Drugs 2019, 17, 90. https://doi.org/10.3390/md17020090
Silva AFR, Abreu H, Silva AMS, Cardoso SM. Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva rigida, Gracilaria sp. and Fucus vesiculosus. Marine Drugs. 2019; 17(2):90. https://doi.org/10.3390/md17020090
Chicago/Turabian StyleSilva, Andreia F.R., Helena Abreu, Artur M.S. Silva, and Susana M. Cardoso. 2019. "Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva rigida, Gracilaria sp. and Fucus vesiculosus" Marine Drugs 17, no. 2: 90. https://doi.org/10.3390/md17020090
APA StyleSilva, A. F. R., Abreu, H., Silva, A. M. S., & Cardoso, S. M. (2019). Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva rigida, Gracilaria sp. and Fucus vesiculosus. Marine Drugs, 17(2), 90. https://doi.org/10.3390/md17020090