Effect of Ishophloroglucin A, A Component of Ishige okamurae, on Glucose Homeostasis in the Pancreas and Muscle of High Fat Diet-Fed Mice

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
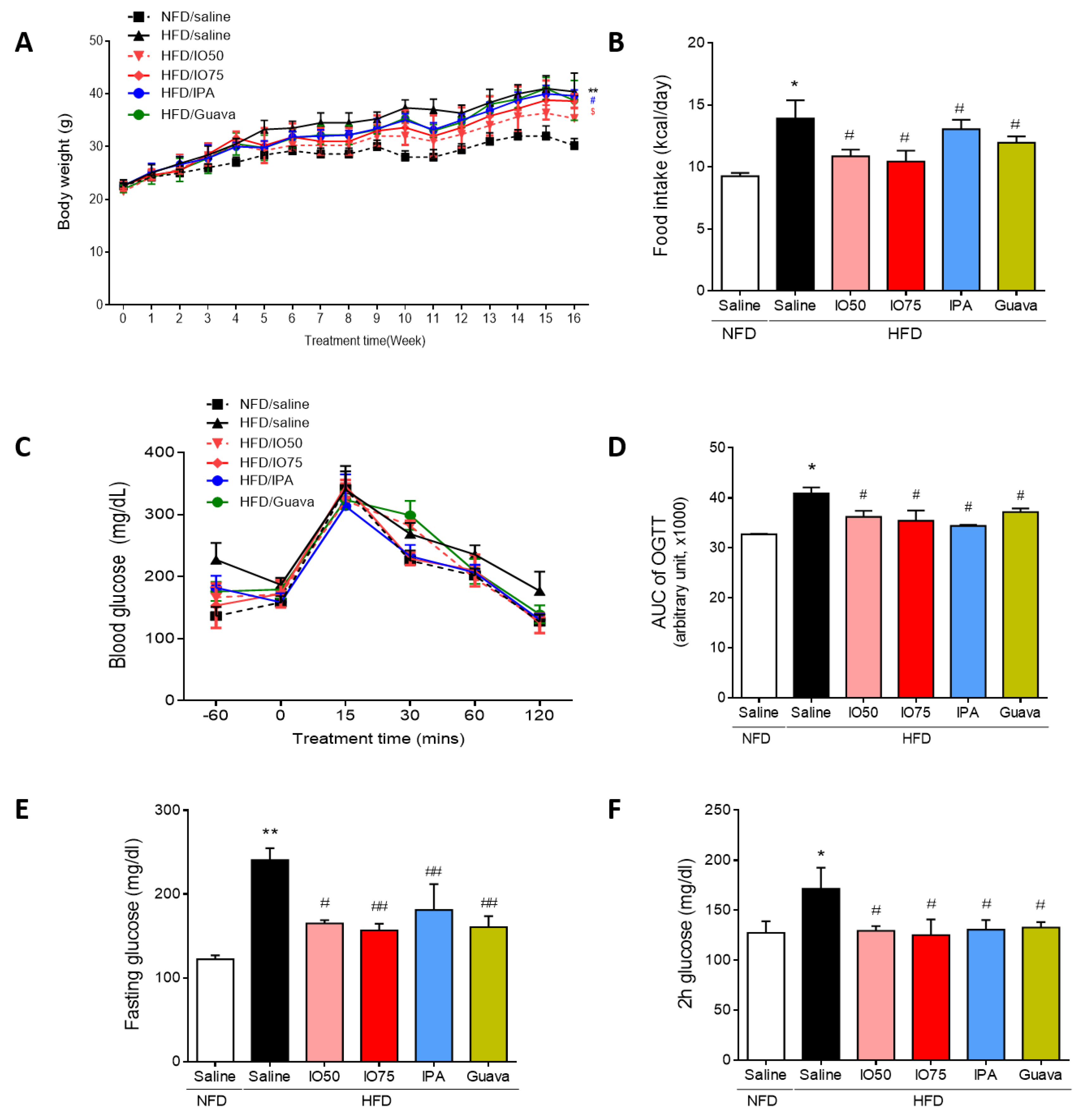
2.1. Improvement in Glucose Tolerance by IPA in HFD Mice
2.2. Regulation of Glucose Homeostatic Imbalance by IPA in HFD Mice
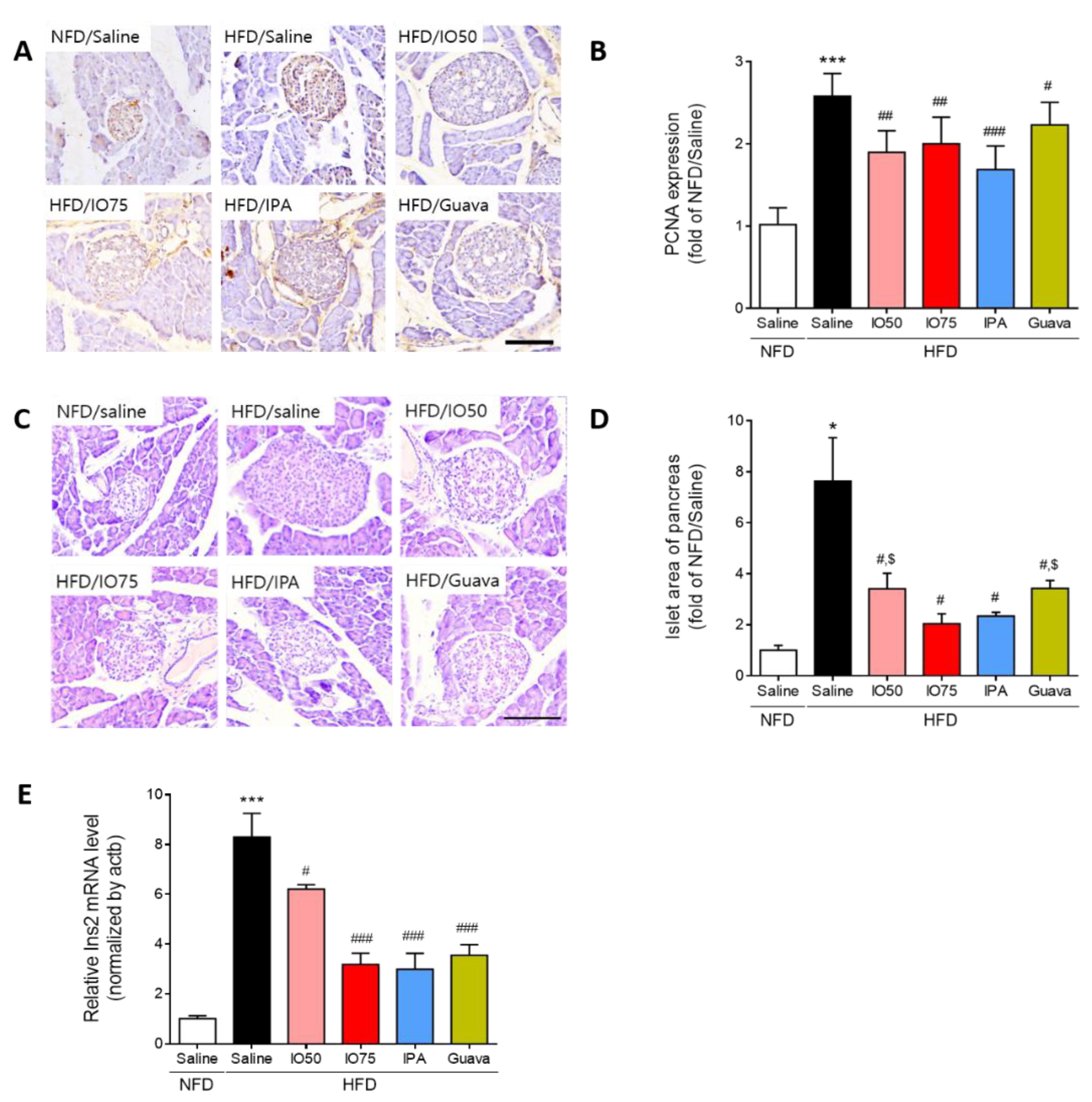
2.3. Protective Effect of IPA on Pancreatic Dysfunction in HFD Mice
2.4. Effect of IPA on Glucose Transport in the Pancreas and Skeletal Muscle of Mice
3. Discussion
4. Materials and Methods
4.1. Preparation of IO and IPA
4.2. Animals
4.3. Insulin Measurement
4.4. C-peptide Measurement
4.5. Sample Preparation
4.5.1. RNA Isolation and cDNA Synthesis
4.5.2. Tissue Preparation
4.6. Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
4.7. Immunohistochemistry: 3, 3 -Diaminobenzidine (DAB) Staining
4.8. Immunohistochemistry: Fluorescence Staining
4.9. Histology: Hematoxylin and Eosin Staining
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Kiuru, P.; D’Auria, M.V.; Muller, C.D.; Tammela, P.; Vuorela, H.; Yli-Kauhaluoma, J. Exploring marine resources for bioactive compounds. Planta Med. 2014, 80, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.A.; Lee, W.W.; Kim, J.-I.; Jeon, Y.-J. Exploiting biological activities of brown seaweed Ishige okamurae Yendo for potential industrial applications: A review. J. Appl. Phycol. 2017, 29, 3109–3119. [Google Scholar] [CrossRef]
- Ryu, B.; Jiang, Y.; Kim, H.-S.; Hyun, J.-M.; Lim, S.-B.; Li, Y.; Jeon, Y.-J. Ishophloroglucin A, a novel phlorotannin for standardizing the anti-α-glucosidase activity of Ishige okamurae. Mar. Drugs 2018, 16, 436. [Google Scholar] [CrossRef]
- Rudkowska, I. Functional foods for health: Focus on diabetes. Maturitas 2009, 62, 263–269. [Google Scholar] [CrossRef]
- Koolman, J.; Röhm, K.-H.; Wirth, J.; Robertson, M. Color Atlas of Biochemistry; Thieme: Stuttgart, Germany, 2005; Volume 2. [Google Scholar]
- Shepherd, P.R.; Kahn, B.B. Glucose transporters and insulin action—Implications for insulin resistance and diabetes mellitus. N. Engl. J. Med. 1999, 341, 248–257. [Google Scholar] [CrossRef]
- Cohen, N.; Shaw, J. Diabetes: Advances in treatment. Intern. Med. J. 2007, 37, 383–388. [Google Scholar] [CrossRef]
- Nathan, D.M. Diabetes: Advances in diagnosis and treatment. JAMA 2015, 314, 1052–1062. [Google Scholar] [CrossRef]
- Kazeem, M.I.; Davies, T.C. Anti-diabetic functional foods as sources of insulin secreting, insulin sensitizing and insulin mimetic agents. J. Funct. Foods 2016, 20, 122–138. [Google Scholar] [CrossRef]
- Xing, X.-H.; Zhang, Z.-M.; Hu, X.-Z.; Wu, R.-Q.; Xu, C. Antidiabetic effects of Artemisia sphaerocephala Krasch. gum, a novel food additive in China, on streptozotocin-induced type 2 diabetic rats. J. Ethnopharmacol. 2009, 125, 410–416. [Google Scholar] [CrossRef]
- Latha, R.C.R.; Daisy, P. Insulin-secretagogue, antihyperlipidemic and other protective effects of gallic acid isolated from Terminalia bellerica Roxb. in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2011, 189, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Egede, L.E.; Ye, X.; Zheng, D.; Silverstein, M.D. The prevalence and pattern of complementary and alternative medicine use in individuals with diabetes. Diabetes Care 2002, 25, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Tindle, H.A.; Davis, R.B.; Phillips, R.S.; Eisenberg, D.M. Trends in use of complementary and alternative medicine by US adults: 1997–2002. Altern. Ther. Health Med. 2005, 11, 42. [Google Scholar] [PubMed]
- Buettner, R.; Parhofer, K.; Woenckhaus, M.; Wrede, C.; Kunz-Schughart, L.; Scholmerich, J.; Bollheimer, L. Defining high-fat-diet rat models: Metabolic and molecular effects of different fat types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Chrysochou, P. Food health branding: The role of marketing mix elements and public discourse in conveying a healthy brand image. J. Mark. Commun. 2010, 16, 69–85. [Google Scholar] [CrossRef]
- Goetzke, B.; Nitzko, S.; Spiller, A. Consumption of organic and functional food. A matter of well-being and health? Appetite 2014, 77, 96–105. [Google Scholar] [CrossRef]
- Altgeld, T.; Gene, R.; Glaeske, G.; Koli, P.; Rosenbrock, R.; Trojan, A. Prevention and Health Promotion. In A Program for Improved Health and Social Policy; Bonner Universitaet Sdruckerei: Bonn, Germany, 2006. [Google Scholar]
- Ballali, S.; Lanciai, F. Functional food and diabetes: A natural way in diabetes prevention? Int. J. Food Sci. Nutr. 2012, 63, 51–61. [Google Scholar] [CrossRef]
- Bech-Larsen, T.; Grunert, K.G. The perceived healthiness of functional foods: A conjoint study of Danish, Finnish and American consumers’ perception of functional foods. Appetite 2003, 40, 9–14. [Google Scholar] [CrossRef]
- Deguchi, Y.; Miyazaki, K. Anti-hyperglycemic and anti-hyperlipidemic effects of guava leaf extract. Nutr. Metab. 2010, 7, 9. [Google Scholar] [CrossRef]
- Rolls, B.J.; Shide, D.J. The influence of dietary fat on food intake and body weight. Nutr. Rev. 1992, 50, 283–290. [Google Scholar] [CrossRef]
- Tremblay, A.; Lavallee, N.; Alméras, N.; Allard, L.; Després, J.-P.; Bouchard, C. Nutritional determinants of the increase in energy intake associated with a high-fat diet. Am. J. Clin. Nutr. 1991, 53, 1134–1137. [Google Scholar] [CrossRef] [PubMed]
- Bringhenti, I.; Moraes-Teixeira, J.A.; Cunha, M.R.; Ornellas, F.; Mandarim-de-Lacerda, C.A.; Aguila, M.B. Maternal obesity during the preconception and early life periods alters pancreatic development in early and adult life in male mouse offspring. PLoS ONE 2013, 8, e55711. [Google Scholar] [CrossRef] [PubMed]
- Bonny, C.; Roduit, R.; Gremlich, S.; Nicod, P.; Thorens, B.; Waeber, G. The loss of GLUT2 expression in the pancreatic β-cells of diabetic db/db mice is associated with an impaired DNA-binding activity of islet-specific trans-acting factors. Mol. Cell. Endocrinol. 1997, 135, 59–65. [Google Scholar] [CrossRef]
- Tremblay, F.; Lavigne, C.; Jacques, H.; Marette, A. Defective insulin-induced GLUT4 translocation in skeletal muscle of high fat–fed rats is associated with alterations in both Akt/protein kinase B and atypical protein kinase C (ζ/λ) activities. Diabetes 2001, 50, 1901–1910. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Cho, S.-Y.; Bhuiyan, M.J.H.; Kim, K.H.; Lee, S.-J. Anti-diabetic effects of lemon balm (Melissa officinalis) essential oil on glucose-and lipid-regulating enzymes in type 2 diabetic mice. Br. J. Nutr. 2010, 104, 180–188. [Google Scholar] [CrossRef]
- Winzell, M.S.; Ahrén, B. The high-fat diet–fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53, S215–S219. [Google Scholar] [CrossRef]
- Srinivasan, K.; Viswanad, B.; Asrat, L.; Kaul, C.; Ramarao, P. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res. 2005, 52, 313–320. [Google Scholar] [CrossRef]
- Heding, L.G.; Larsen, U.; Markussen, J.; Jørgensen, K.; Hallund, O. Radioimmunoassays for human, pork and ox C-peptides and related substances. In Radioimmunoassays for Insulin, C-Peptide and Proinsulin; Springer: Berlin/Heidelberg, Germany, 1988; pp. 43–47. [Google Scholar]
- Sima, A.A.; Kamiya, H.; Li, Z.G. Insulin, C-peptide, hyperglycemia, and central nervous system complications in diabetes. Eur. J. Pharmacol. 2004, 490, 187–197. [Google Scholar] [CrossRef]
- Grunberger, G.; Sima, A.A. The C-peptide signaling. J. Diabetes Res. 2004, 5, 25–36. [Google Scholar] [CrossRef]
- Kasuga, M. Insulin resistance and pancreatic β cell failure. J. Clin. Investig. 2006, 116, 1756–1760. [Google Scholar] [CrossRef]
- Matveyenko, A.V.; Gurlo, T.; Daval, M.; Butler, A.E.; Butler, P.C. Successful versus failed adaptation to high-fat diet–induced insulin resistance: The role of IAPP-induced β-cell endoplasmic reticulum stress. Diabetes 2009, 58, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Janson, J.; Soeller, W.C.; Butler, P.C. Increased β-cell apoptosis prevents adaptive increase in β-cell mass in mouse model of type 2 diabetes: Evidence for role of islet amyloid formation rather than direct action of amyloid. Diabetes 2003, 52, 2304–2314. [Google Scholar] [CrossRef] [PubMed]
- Rahier, J.; Guiot, Y.; Goebbels, R.; Sempoux, C.; Henquin, J.-C. Pancreatic β-cell mass in European subjects with type 2 diabetes. Diabetes Obes. Metab. 2008, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Poitout, V.; Robertson, R.P. An integrated view of β-cell dysfunction in type-II diabetes. Annu. Rev. Med. 1996, 47, 69–83. [Google Scholar] [CrossRef]
- Durning, S.P.; Flanagan-Steet, H.; Prasad, N.; Wells, L. O-linked β-N-acetylglucosamine (O-GlcNAc) acts as a glucose sensor to epigenetically regulate the insulin gene in pancreatic beta cells. J. Biol. Chem. 2016, 291, 2107–2118. [Google Scholar] [CrossRef]
- Nishiumi, S.; Bessyo, H.; Kubo, M.; Aoki, Y.; Tanaka, A.; Yoshida, K.-I.; Ashida, H. Green and black tea suppress hyperglycemia and insulin resistance by retaining the expression of glucose transporter 4 in muscle of high-fat diet-fed C57BL/6J mice. J. Agric. Food Chem. 2010, 58, 12916–12923. [Google Scholar] [CrossRef]
- Herman, M.A.; Kahn, B.B. Glucose transport and sensing in the maintenance of glucose homeostasis and metabolic harmony. J. Clin. Investig. 2006, 116, 1767–1775. [Google Scholar] [CrossRef]
- Shan, W.-F.; Chen, B.-Q.; Zhu, S.-J.; Jiang, L.; Zhou, Y.-F. Effects of GLUT4 expression on insulin resistance in patients with advanced liver cirrhosis. J. Zhejiang Univ. Sci. B 2011, 12, 677. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.-W.; Son, M.; Choi, J.; Oh, S.; Jeon, Y.-J.; Byun, K.; Ryu, B. Effect of Ishophloroglucin A, A Component of Ishige okamurae, on Glucose Homeostasis in the Pancreas and Muscle of High Fat Diet-Fed Mice. Mar. Drugs 2019, 17, 608. https://doi.org/10.3390/md17110608
Yang H-W, Son M, Choi J, Oh S, Jeon Y-J, Byun K, Ryu B. Effect of Ishophloroglucin A, A Component of Ishige okamurae, on Glucose Homeostasis in the Pancreas and Muscle of High Fat Diet-Fed Mice. Marine Drugs. 2019; 17(11):608. https://doi.org/10.3390/md17110608
Chicago/Turabian StyleYang, Hye-Won, Myeongjoo Son, Junwon Choi, Seyeon Oh, You-Jin Jeon, Kyunghee Byun, and BoMi Ryu. 2019. "Effect of Ishophloroglucin A, A Component of Ishige okamurae, on Glucose Homeostasis in the Pancreas and Muscle of High Fat Diet-Fed Mice" Marine Drugs 17, no. 11: 608. https://doi.org/10.3390/md17110608
APA StyleYang, H.-W., Son, M., Choi, J., Oh, S., Jeon, Y.-J., Byun, K., & Ryu, B. (2019). Effect of Ishophloroglucin A, A Component of Ishige okamurae, on Glucose Homeostasis in the Pancreas and Muscle of High Fat Diet-Fed Mice. Marine Drugs, 17(11), 608. https://doi.org/10.3390/md17110608