Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms

 ,
,  and
and
Abstract
1. Introduction
2. Summary of Literature
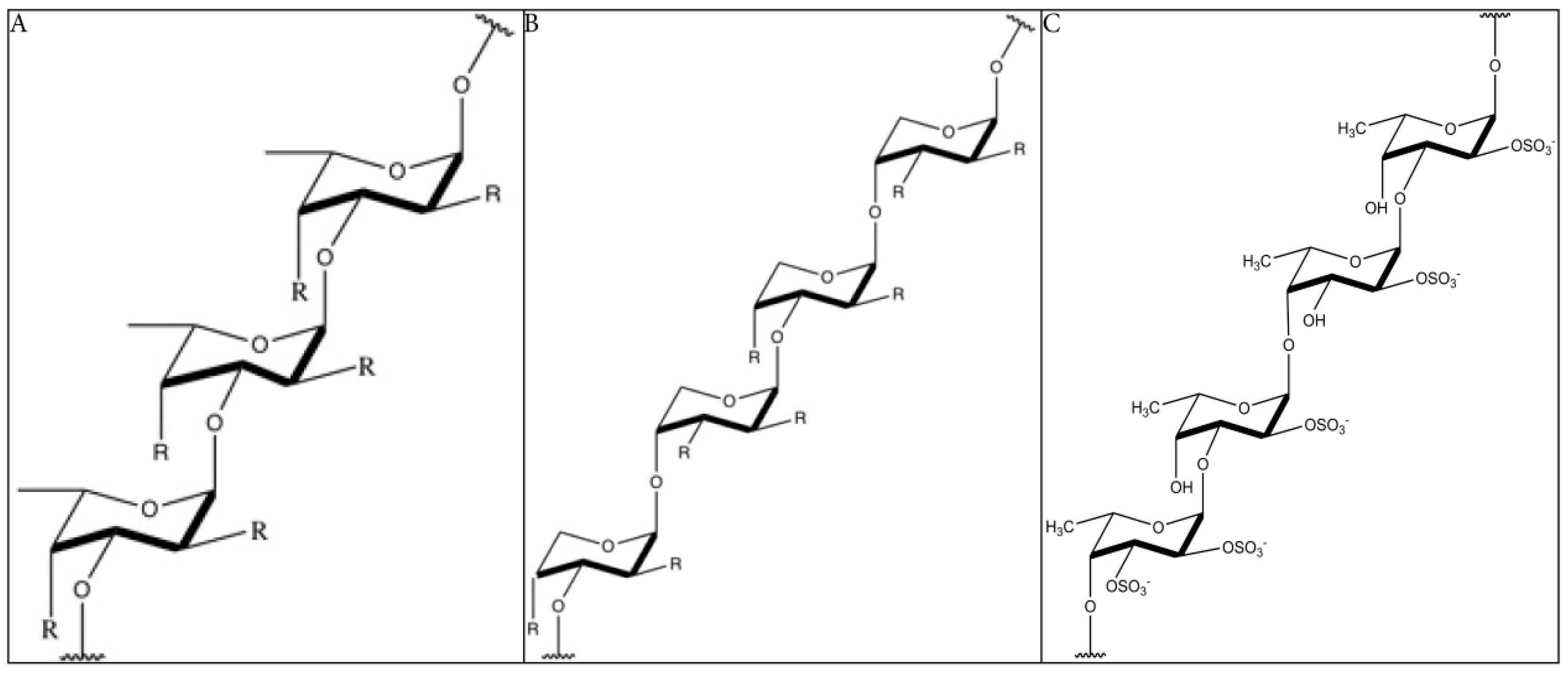
2.1. Structure of Fucoidan
2.2. Molecular Weight
2.3. Sulfate Groups
2.4. Pharmacokinetics
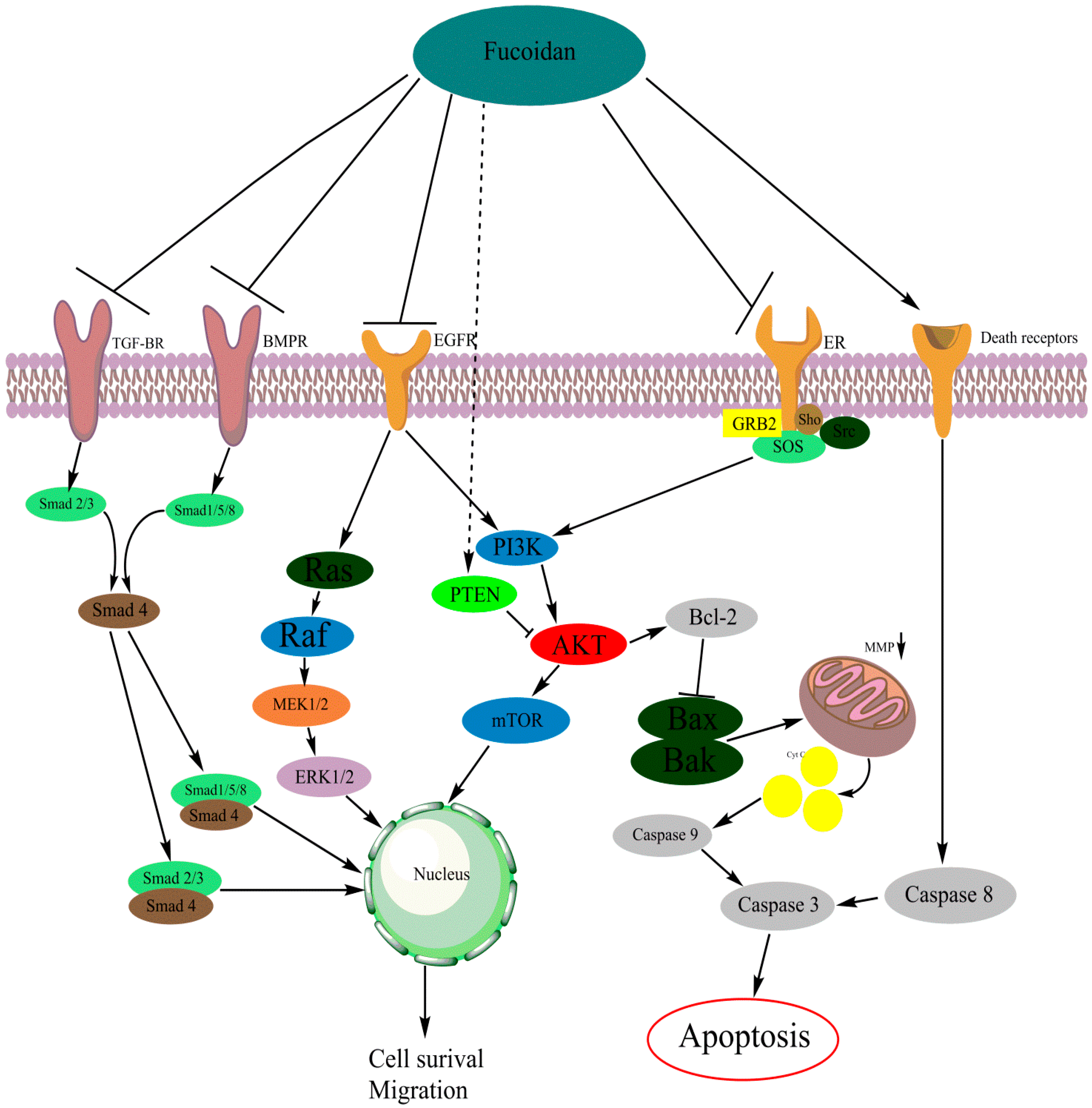
2.5. Cellular Mechanisms in Relation to Fucoidan Anti-Cancer Activity
2.5.1. The PI3K/AKT Pathway
2.5.2. MAPK Signaling Pathway
2.5.3. The Caspase Pathway
2.6. Growth-Involved Receptors
2.6.1. Transforming Growth Factor Beta
2.6.2. Bone Morphogenetic Protein
2.6.3. Vascular Endothelial Growth Factor
2.6.4. Estrogen Receptor
3. Discussion
3.1. Structure and Bioactivity
3.2. Cellular Mechanisms in Relation to Fucoidan Anti-Cancer Activity
3.3. Receptors in Relation to Fucoidan Anti-Cancer Activity
3.4. Clinical Significance
3.5. Future Perspectives
4. Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADME | Absorption, distribution, metabolism, and excretion |
| AKT | Protein Kinase B |
| Ang-2 | Angiopoietin-2 |
| ASK1 | Apoptosis Signal-regulating Kinase 1 |
| Bcl-2 | B-cell lymphoma 2 |
| BMP | Bone morphogenetic protein |
| Da | Dalton |
| E2 | Estradiol |
| ECM | Extracellular Matrix |
| EGF | Epidermal Growth Factor |
| ELISA | Enzyme-linked Immuno Sorbent Assay |
| EMT | Extracellular Matrix Kinase |
| ER | Estrogen Receptor |
| ERK | Extracellular Signal-Regulated Kinase |
| FGF | Fibroblast Growth Factor |
| FGFR | Fibroblast Growth Factor Receptor |
| FVE | Fucus vesiculosus Extract |
| GTP | Guanosine-5′-triphosphate |
| HIF-1a | Hypoxia-Inducible Factor 1-alpha |
| HMEC-1 | Human Microvascular Endothelial Cell line-1 |
| HMWF | High Molecular Weight Fucoidan |
| JNK | c-Jun N-terminal Kinase |
| LMWF | Low Molecular Weight Fucoidan |
| MAPK | Mitogen-Activated Protein Kinase |
| MMP | Matrix Metalloproteinase |
| MMWF | Medium Molecular Weight Fucoidan |
| mTOR | mammalian Target of Rapamycin |
| NF-kB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NRP1 | Neuropilin 1 |
| PKC | Protein Kinase C |
| PTEN | Phosphatase and Tensin Homolog |
| ROS | Reactive Oxygen Species |
| SDF-1 | Stromal cell Derived Factor-1 |
| TGF-β | Transforming Growth Factor-Béta |
| TGF-R | Transforming Growth Factor-Receptor |
| UPS | Undaria pinnatifida purified fraction |
| VEGF | Vascular Endothelial Growth Factor |
| VEGFR | Vascular Endothelial Growth Factor Receptor |
Appendix A
Appendix A.1. Web of Science
- TOPIC:(fucoidan) ANDTOPIC: (structure)
- TOPIC: (sulfated polysaccharides) ANDTOPIC:(structure)
- TOPIC: (sulfated polysaccharides) ANDTOPIC:(pharmacokinetics)
- TOPIC: (sulfated polysaccharides) ANDTOPIC:(absorption)
- TOPIC:(fucoidan) ANDTOPIC: (absorption)
- TOPIC:(fucoidan) ANDTOPIC:(pharmacokinetics)
- TOPIC:(fucoidan) ANDTOPIC: (degradation)
- TOPIC:(fucoidan) ANDTOPIC: (preparation) NOTTOPIC: (nanoparticles)
- TOPIC:(fucoidan) ANDTOPIC: (sulfate)
- TOPIC:(fucoidan) ANDTOPIC: (species)
- TOPIC:(fucoidan) ANDTOPIC: (fucus vesiculosus)
- TOPIC: (fucus vesiculosus) ANDTOPIC: (structure)
- TOPIC: (Undaria pinnatifida) ANDTOPIC: (structure)
- TOPIC:(fucoidan) ANDTOPIC: (Undaria pinnatifida)
- TOPIC:(fucoidan) ANDTOPIC: (cladosiphon okamuranus)
- TOPIC:(fucoidan) ANDTOPIC: (ascophyllum nodosum)
- TOPIC:(fucoidan) ANDTOPIC: (fucales)
- TOPIC:(fucoidan) ANDTOPIC: (brown algae)
- TOPIC:(fucoidan) ANDTOPIC: (cancer)
- TOPIC:(fucoidan) ANDTOPIC: (tumor)
- TOPIC: (Undaria pinnatifida) ANDTOPIC: (cancer)
- TOPIC: (fucus vesiculosus) ANDTOPIC: (tumor)
- TOPIC:(fucoidan) ANDTOPIC: (endometrial cancer)
- TOPIC:(fucoidan) ANDTOPIC: (breast cancer)
- TOPIC:(fucoidan) ANDTOPIC: (colon cancer)
- TOPIC:(fucoidan) ANDTOPIC: (lung cancer)
- TOPIC:(fucoidan) ANDTOPIC: (PI3K/AKT)
- TOPIC:(fucoidan) ANDTOPIC: (MAPK)
- TOPIC:(fucoidan) ANDTOPIC: (transforming growth factor beta)
- TOPIC:(fucoidan) ANDTOPIC: (PTEN)
- TOPIC: (Low molecular weight fucoidan) ANDTOPIC: (structure)
- TOPIC:(Low molecular weight fucoidan)
- TOPIC:(fucoidan) ANDTOPIC: (VEGF)
- TOPIC:(fucoidan) ANDTOPIC: (Bone morphogenetic protein)
- TOPIC:(fucoidan) ANDTOPIC: (estrogen)
- TOPIC:(fucoidan) ANDTOPIC: (migration)
- TOPIC:(fucoidan) ANDTOPIC: (apoptosis)
- TOPIC: (fucoidan) AND TOPIC: (caspase)
- TOPIC: (fucoidan) AND TOPIC: (caspase pathway)
- TOPIC: (fucoidan) AND TOPIC: (extrinsic apoptosis)
- TOPIC: (fucoidan) AND TOPIC: (intrinsic apoptosis)
Appendix A.2. Criteria for Considering Studies for This Review
Appendix A.3. Search Methods
References
- Svejda, B.; Aguiriano-Moser, V.; Sturm, S.; Hoger, H.; Ingolic, E.; Siegl, V.; Stuppner, H.; Pfragner, R. Anticancer Activity of Novel Plant Extracts from Trailliaedoxa gracilis (W. W. Smith & Forrest) in Human Carcinoid KRJ-I Cells. Anticancer Res. 2010, 30, 55–64. [Google Scholar] [PubMed]
- Cuong, H.D.; Thuy, T.T.T.; Huong, T.T.; Ly, B.M.; Van, T.T.T. Structure and hypolipidaemic activity of fucoidan extracted from brown seaweed Sargassum henslowianum. Nat. Prod. Res. 2015, 29, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Kinghorn, A.D.; Chin, Y.W.; Swanson, S.M. Discovery of natural product anticancer agents from biodiverse organisms. Curr. Opin. Drug Discov. Dev. 2009, 12, 189–196. [Google Scholar]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Kim, W.J.; Kim, S.M.; Pohl, R.; Kvasnicka, F.; Copikova, J.; Park, Y.I. Structure and antitumour activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Cock, J.M.; Peters, A.F.; Coelho, S.M. Brown algae. Curr. Biol. 2011, 21, R573–R575. [Google Scholar] [CrossRef] [PubMed]
- Cunha, L.; Grenha, A. Sulfated Seaweed Polysaccharides as Multifunctional Materials in Drug Delivery Applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Kim, E.A.; Son, K.T.; Jeon, Y.J. Bioactive properties and potentials cosmeceutical applications of phlorotannins isolated from brown seaweeds: A review. J. Photochem. Photobiol. B-Biol. 2016, 162, 100–105. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global Cancer Statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- Leal, M.C.; Munro, M.H.G.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Avila, L.M.; Morales-Martínez, M.E.; Ricque-Marie, D.; Cruz-Suarez, L.E.; Zapata-Benavides, P.; Morán-Santibañez, K.; Rodríguez-Padilla, C. In vitro anti-canine distemper virus activity of fucoidan extracted from the brown alga Cladosiphon okamuranus. VirusDisease 2014, 25, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Besednova, N.N.; Zaporozhets, T.S.; Somova, L.M.; Kuznetsova, T.A. Review: Prospects for the Use of Extracts and Polysaccharides from Marine Algae to Prevent and Treat the Diseases Caused by Helicobacter Pylori. Helicobacter 2015, 20, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan Extracts Ameliorate Acute Colitis. PLoS ONE 2015, 10, e0128453. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.; Lu, J.; Roscilli, G.; Aurisicchio, L.; Cappelletti, M.; Pavoni, E.; White, W.L.; Bedogni, B. The natural compound fucoidan from New Zealand Undaria pinnatifida synergizes with the ERBB inhibitor lapatinib enhancing melanoma growth inhibition. Oncotarget 2017, 8, 17887–17896. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, F.; Wei, X.J.; Zhao, R.X. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Lee, J.S.; Kim, W.S.; Jeon, Y.J. The potential of brown-algae polysaccharides for the development of anticancer agents: An update on anticancer effects reported for fucoidan and laminaran. Carbohydr. Polym. 2017, 177, 451–459. [Google Scholar] [CrossRef]
- Chevolot, L.; Mulloy, B.; Ratiskol, J.; Foucault, A.; Colliec-Jouault, S. A disaccharide repeat unit is the major structure in fucoidans from two species of brown algae. Carbohydr. Res. 2001, 330, 529–535. [Google Scholar] [CrossRef]
- Wu, L.; Sun, J.; Su, X.T.; Yu, Q.L.; Yu, Q.Y.; Zhang, P. A review about the development of fucoidan in antitumor activity: Progress and challenges. Carbohydr. Polym. 2016, 154, 96–111. [Google Scholar] [CrossRef]
- Nishino, T.; Nishioka, C.; Ura, H.; Nagumo, T. Isolation and partial characterization of a novel amino sugar-containing fucan sulfate from commercial Fucus-vesiculosus Fucoidan. Carbohydr. Res. 1994, 255, 213–224. [Google Scholar] [CrossRef]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus evanescens C.Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A highly regular fraction of a fucoidan from the brown seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus serratus L. Carbohydr. Res. 2006, 341, 238–245. [Google Scholar] [CrossRef]
- Adhikari, U.; Mateii, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structure and antiviral activity of sulfated fucans from Stoechospermum marginatum. Phytochemistry 2006, 67, 2474–2482. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.A.; Szegezdi, E.; Mulloy, B.; Samali, A.; Tuohy, M.G. An Unfractionated Fucoidan from Ascophyllum nodosum: Extraction, Characterization, and Apoptotic Effects in Vitro (vol 74, pg 1851, 2011). J. Nat. Prod. 2012, 75, 1674. [Google Scholar] [CrossRef]
- Descamps, V.; Colin, S.; Lahaye, M.; Jam, M.; Richard, C.; Potin, P.; Barbeyron, T.; Yvin, J.C.; Kloareg, B. Isolation and culture of a marine bacterium degrading the sulfated fucans from marine brown algae. Mar. Biotechnol. 2006, 8, 27–39. [Google Scholar] [CrossRef]
- Jabbar Mian, A.; Percival, E. Carbohydrates of the brown seaweeds himanthalia lorea and bifurcaria bifurcata: Part II. structural studies of the “fucans”. Carbohydr. Res. 1973, 26, 147–161. [Google Scholar] [CrossRef]
- Chevolot, L.; Foucault, A.; Chaubet, F.; Kervarec, N.; Sinquin, C.; Fisher, A.M.; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res. 1999, 319, 154–165. [Google Scholar] [CrossRef]
- Nagaoka, M.; Shibata, H.; Kimura-Takagi, I.; Hashimoto, S.; Kimura, K.; Makino, T.; Aiyama, R.; Ueyama, S.; Yokokura, T. Structural study of fucoidan from Cladosiphon okamuranus Tokida. Glycoconjug. J. 1999, 16, 19–26. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Pujol, C.A.; Damonte, E.B.; Flores, M.L.; Stortz, C.A. Fucoidans from the brown seaweed Adenocystis utricularis: Extraction methods, antiviral activity and structural studies. Carbohydr. Res. 2003, 338, 153–165. [Google Scholar] [CrossRef]
- Marais, M.F.; Joseleau, J.P. A fucoidan fraction from Ascophyllum nodosum. Carbohydr. Res. 2001, 336, 155–159. [Google Scholar] [CrossRef]
- Duarte, M.E.R.; Cardoso, M.A.; Noseda, M.D.; Cerezo, A.S. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Designed optimization of a single-step extraction of fucose-containing sulfated polysaccharides from Sargassum sp. J. Appl. Phycol. 2012, 24, 715–723. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wei, X.J.; Sun, J.L.; Xu, S.Y. Structural investigation of a fucoidan containing a fucose-free core from the brown seaweed, Hizikia fusiforme. Carbohydr. Res. 2006, 341, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Chizhov, A.O.; Dell, A.; Morris, H.R.; Haslam, S.M.; McDowell, R.A.; Shashkov, A.S.; Nifant’ev, N.E.; Khatuntseva, E.A.; Usov, A.I. A study of fucoidan from the brown seaweed Chorda filum. Carbohydr. Res. 1999, 320, 108–119. [Google Scholar] [CrossRef]
- Hemmingson, J.A.; Falshaw, R.; Furneaux, R.H.; Thompson, K. Structure and antiviral activity of the galactofucan sulfates extracted from Undaria Pinnatifida (Phaeophyta). J. Appl. Phycol. 2006, 18, 185–193. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhang, Q.B.; Wang, J.; Shi, X.L.; Zhang, Z.S. Analysis of the monosaccharide composition of fucoidan by precolumn derivation HPLC. Chin. J. Oceanol. Limnol. 2009, 27, 578–582. [Google Scholar] [CrossRef]
- Mak, W.; Hamid, N.; Liu, T.; Lu, J.; White, W.L. Fucoidan from New Zealand Undaria pinnatifida: Monthly variations and determination of antioxidant activities. Carbohydr. Polym. 2013, 95, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, H.R.; Biller, P.; Ross, A.B.; Adams, J.M.M. The seasonal variation of fucoidan within three species of brown macroalgae. Algal Res.-Biomass Biofuels Bioprod. 2017, 22, 79–86. [Google Scholar] [CrossRef]
- Senthilkumar, K.; Manivasagan, P.; Venkatesan, J.; Kim, S.K. Brown seaweed fucoidan: Biological activity and apoptosis, growth signaling mechanism in cancer. Int. J. Biol. Macromol. 2013, 60, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Teruya, K.; Eto, H.; Shirahata, S. Induction of Apoptosis by Low-Molecular-Weight Fucoidan through Calcium- and Caspase-Dependent Mitochondrial Pathways in MDA-MB-231 Breast Cancer Cells. Biosci. Biotechnol. Biochem. 2013, 77, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Hsu, W.L.; Hwang, P.A.; Chou, T.C. Low Molecular Weight Fucoidan Inhibits Tumor Angiogenesis through Downregulation of HIF-1/VEGF Signaling under Hypoxia. Mar. Drugs 2015, 13, 4436–4451. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Hsu, W.L.; Hwang, P.A.; Chen, Y.L.; Chou, T.C. Combined administration of fucoidan ameliorates tumor and chemotherapy-induced skeletal muscle atrophy in bladder cancer-bearing mice. Oncotarget 2016, 7, 51608–51618. [Google Scholar] [CrossRef] [PubMed]
- Matou, S.; Helley, D.; Chabut, D.; Bros, A.; Fischer, A.M. Effect of fucoidan on fibroblast growth factor-2-induced angiogenesis in vitro. Thromb. Res. 2002, 106, 213–221. [Google Scholar] [CrossRef]
- Chabut, D.; Fischer, A.M.; Hettey, D.; Colliec, S. Low molecular weight fucoidan promotes FGF-2-induced vascular tube formation by human endothelial cells, with decreased PAI-1 release and ICAM-1 downregulation. Thromb. Res. 2004, 113, 93–95. [Google Scholar] [CrossRef]
- Matsubara, K.; Xue, C.; Zhao, X.; Mori, M.; Sugawara, T.; Hirata, T. Effects of middle molecular weight fucoidans on in vitro and ex vivo angiogenesis of endothelial cells. Int. J. Mol. Med. 2005, 15, 695–699. [Google Scholar] [CrossRef]
- Hwang, P.A.; Yan, M.D.; Kuo, K.L.; Phan, N.N.; Lin, Y.C. A mechanism of low molecular weight fucoidans degraded by enzymatic and acidic hydrolysis for the prevention of UVB damage. J. Appl. Phycol. 2017, 29, 521–529. [Google Scholar] [CrossRef]
- Haroun-Bouhedja, F.; Ellouali, M.; Sinquin, C.; Boisson-Vidal, C. Relationship between sulfate groups and biological activities of fucans. Thromb. Res. 2000, 100, 453–459. [Google Scholar] [CrossRef]
- Li, B.; Zhao, R.X.; Wei, X.J. Anticoagulant activity of fucoidan from Hizikia fusiforme. Agro Food Ind. Hi-Tech 2008, 19, 22–24. [Google Scholar]
- Cho, M.L.; Lee, B.Y.; You, S. Relationship between Oversulfation and Conformation of Low and High Molecular Weight Fucoidans and Evaluation of Their in Vitro Anticancer Activity. Molecules 2011, 16, 291–297. [Google Scholar] [CrossRef]
- Koyanagi, S.; Tanigawa, N.; Nakagawa, H.; Soeda, S.; Shimeno, H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol. 2003, 65, 173–179. [Google Scholar] [CrossRef]
- Oliveira, C.; Ferreira, A.S.; Novoa-Carballal, R.; Nunes, C.; Pashkuleva, I.; Neves, N.M.; Coimbra, M.A.; Reis, R.L.; Martins, A.; Silva, T.H. The Key Role of Sulfation and Branching on Fucoidan Antitumor Activity. Macromol. Biosci. 2017, 17, 1600340. [Google Scholar] [CrossRef]
- Soeda, S.; Ishida, S.; Shimeno, H.; Nagamatsu, A. Inhibitory effect of oversulfated fucoidan on invasion through reconstituted basement-membrane by murine lewis lung-carcinoma. Jpn. J. Cancer Res. 1994, 85, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Kasai, A.; Arafuka, S.; Koshiba, N.; Takahashi, D.; Toshima, K. Systematic synthesis of low-molecular weight fucoidan derivatives and their effect on cancer cells. Organ. Biomol. Chem. 2015, 13, 10556–10568. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Popivnich, I.B.; Isakov, V.V.; Scobun, A.S.; Sundukova, E.V.; Elyakova, L.A. A new procedure for the separation of water-soluble polysaccharides from brown seaweeds. Carbohydr. Res. 1999, 322, 32–39. [Google Scholar] [CrossRef]
- Grachev, A.A.; Gerbst, A.G.; Ustyuzhanina, N.E.; Shashkov, A.S.; Usov, A.I.; Nifantiev, N.E. NMR investigation of the influence of sulfate groups at C-2 and C-4 on the conformational behavior of fucoidan fragments with homo-(1 -> 3)-linked backbone. J. Carbohydr. Chem. 2006, 25, 315–330. [Google Scholar] [CrossRef]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and Tissue Distribution of Fucoidan from Fucus vesiculosus after Oral Administration to Rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, H.D.; Li, N.; Zhang, Q.B.; Zhang, H. The Protective Effect of Fucoidan in Rats with Streptozotocin-Induced Diabetic Nephropathy. Mar. Drugs 2014, 12, 3292–3306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.B.; Li, Z.; Xu, Z.H.; Niu, X.Z.; Zhang, H. Effects of fucoidan on chronic renal failure in rats. Planta Med. 2003, 69, 537–541. [Google Scholar] [PubMed]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal Absorption of Fucoidan Extracted from the Brown Seaweed, Cladosiphon okamuranus. Mar. Drugs 2015, 13, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a Fucoidan-Specific Antibody and Measurement of Fucoidan in Serum and Urine by Sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350–357. [Google Scholar] [CrossRef]
- Michel, C.; Lahaye, M.; Bonnet, C.; Mabeau, S.; Barry, J.L. In vitro fermentation by human faecal bacteria of total and purified dietary fibres from brown seaweeds. Br. J. Nutr. 1996, 75, 263–280. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, O.H.; Lee, H.H.; Lee, B.Y. A 4-week repeated oral dose toxicity study of fucoidan from the Sporophyll of Undaria pinnatifida in Sprague-Dawley rats. Toxicology 2010, 267, 154–158. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, O.H.; Lee, B.Y. Genotoxicity studies on fucoidan from Sporophyll of Undaria pinnatifida. Food Chem. Toxicol. 2010, 48, 1101–1104. [Google Scholar] [CrossRef]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Karpiniec, S.S.; Dickinson, J.L. Fucoidan Suppresses the Growth of Human Acute Promyelocytic Leukemia Cells In Vitro and In Vivo. J. Cell. Physiol. 2016, 231, 688–697. [Google Scholar] [CrossRef]
- Huang, T.H.; Chiu, Y.H.; Chan, Y.L.; Wang, H.; Huang, K.C.; Li, T.L.; Hsu, K.H.; Wu, C.J. Prophylactic Administration of Fucoidan Represses Cancer Metastasis by Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs) in Lewis Tumor-Bearing Mice. Mar. Drugs 2015, 13, 1882–1900. [Google Scholar] [CrossRef]
- Marcin Bobiński, K.O.; Bednarek, W.; Wawruszak, A.; Dmoszyńska-Graniczka, M.; Garcia-Sanz, P.; Wertel, I.; Kotarski, J. The Effect of Fucoidan, a Potential New, Natural, Anti-Neoplastic Agent on Uterine Sarcomas and Carcinosarcoma Cell Lines: ENITEC Collaborative Study. Arch. Immunol. Ther. Exp. 2018. submitted for publication. [Google Scholar]
- Zhang, W.J.; Sun, D.L.; Zhao, X.; Jin, W.H.; Wang, J.; Zhang, Q.B. Microanalysis and preliminary pharmacokinetic studies of a sulfated polysaccharide from Laminaria japonica. Chin. J. Oceanol. Limnol. 2016, 34, 177–185. [Google Scholar] [CrossRef]
- Staal, S.P. Molecular cloning of the akt oncogene and its human homologues AKT1 and AKT2: Amplification of AKT1 in a primary human gastric adenocarcinoma. Proc. Natl. Acad. Sci. USA 1987, 84, 5034–5037. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Dan, H.C.; Sun, M.; Liu, Q.Y.; Sun, X.M.; Feldman, R.I.; Hamilton, A.D.; Polokoff, M.; Nicosia, S.V.; Herlyn, M.; et al. Akt/protein kinase B signaling inhibitor-2, a selective small molecule inhibitor of Akt signaling with antitumor activity in cancer cells overexpressing Akt. Cancer Res. 2004, 64, 4394–4399. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Modur, V.; Nagarajan, R.; Evers, B.M.; Milbrandt, J. FOXO proteins regulate tumor necrosis factor-related apoptosis inducing ligand expression—Implications for PTEN mutation in prostate cancer. J. Biol. Chem. 2002, 277, 47928–47937. [Google Scholar] [CrossRef]
- Gilley, J.; Coffer, P.J.; Ham, J. FOXO transcription factors directly activate bim gene expression and promote apoptosis in sympathetic neurons. J. Cell Biol. 2003, 162, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Deveraux, Q.L.; Roy, N.; Stennicke, H.R.; Van Arsdale, T.; Zhou, Q.; Srinivasula, S.M.; Alnemri, E.S.; Salvesen, G.S.; Reed, J.C. IAPs block apoptotic events induced by caspase-8 and cytochrome c by direct inhibition of distinct caspases. EMBO J. 1998, 17, 2215–2223. [Google Scholar] [CrossRef]
- Hu, P.; Han, Z.; Couvillon, A.D.; Exton, J.H. Critical role of endogenous Akt/IAPs and MEK1/ERK pathways in counteracting endoplasmic reticulum stress-induced cell death. J. Biol. Chem. 2004, 279, 49420–49429. [Google Scholar] [CrossRef]
- Deveraux, Q.L.; Reed, T.C. IAP family proteins—Suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef]
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, S.; Koh, H.; Yoon, S.O.; Chung, A.S.; Cho, K.S.; Chung, J. Akt/PKB promotes cancer cell invasion via increased motility and metalloproteinase production. FASEB J. 2001, 15, 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Hsieh, H.L.; Hsiao, L.D.; Yang, C.M. PI3-K/Akt/JNIUNF-kappa B is essential for MMP-9 expression and outgrowth in human limbal epithelial cells on intact amniotic membrane. Stem Cell Res. 2012, 9, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Tanno, S.; Mitsuuchi, Y.; Altomare, D.A.; Xiao, G.H.; Testa, J.R. AKT activation up-regulates insulin-like growth factor I receptor expression and promotes invasiveness of human pancreatic cancer cells. Cancer Res. 2001, 61, 589–593. [Google Scholar] [PubMed]
- Jin, E.J.; Park, K.S.; Bang, O.S.; Kang, S.S. Akt signaling regulates actin organization via modulation of MMP-2 activity during chondrogenesis of chick wing limb bud mesenchymal cells. J. Cell. Biochem. 2007, 102, 252–261. [Google Scholar] [CrossRef]
- Stankovic, S.; Konjevic, G.; Gopcevic, K.; Jovic, V.; Inic, M.; Jurisic, V. Activity of MMP-2 and MMP-9 in sera of breast cancer patients. Pathol. Res. Pract. 2010, 206, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.H.; Kim, S.C.; Kang, J.I.; Kim, M.K.; Boo, H.J.; Kwon, J.M.; Koh, Y.S.; Hyun, J.W.; Park, D.B.; Yoo, E.S.; et al. Apoptosis Inducing Activity of Fucoidan in HCT-15 Colon Carcinoma Cells. Biol. Pharm. Bull. 2009, 32, 1760–1764. [Google Scholar] [CrossRef]
- Han, M.H.; Lee, D.S.; Jeong, J.W.; Hong, S.H.; Choi, I.W.; Cha, H.J.; Kim, S.; Kim, H.S.; Park, C.; Kim, G.Y.; et al. Fucoidan Induces ROS-Dependent Apoptosis in 5637 Human Bladder Cancer Cells by Downregulating Telomerase Activity via Inactivation of the PI3K/Akt Signaling Pathway. Drug Dev. Res. 2017, 78, 37–48. [Google Scholar] [CrossRef]
- Boo, H.J.; Hong, J.Y.; Kim, S.C.; Kang, J.I.; Kim, M.K.; Kim, E.J.; Hyun, J.W.; Koh, Y.S.; Yoo, E.S.; Kwon, J.M.; et al. The Anticancer Effect of Fucoidan in PC-3 Prostate Cancer Cells. Mar. Drugs 2013, 11, 2982–2999. [Google Scholar] [CrossRef]
- Wei, C.M.; Xiao, Q.; Kuang, X.Y.; Zhang, T.; Yang, Z.S.; Wang, L. Fucoidan inhibits proliferation of the SKM-1 acute myeloid leukaemia cell line via the activation of apoptotic pathways and production of reactive oxygen species. Mol. Med. Rep. 2015, 12, 6649–6655. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Riby, J.E.; Conde, L.; Grizzle, W.E.; Cui, X.Q.; Skibola, C.F. A Fucus vesiculosus extract inhibits estrogen receptor activation and induces cell death in female cancer cell lines. BMC Complement. Altern. Med. 2016, 16, 151. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, J.S.; Kim, E. Fucoidan from Seaweed Fucus vesiculosus Inhibits Migration and Invasion of Human Lung Cancer Cell via PI3K-Akt-mTOR Pathways. PLoS ONE 2012, 7, e50624. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.S.; Liu, Z.C.; Liu, X.L.; Teng, H.M.; Zhang, C.L.; Hou, L.; Zou, X.Y. Anti-Metastasis Effect of Fucoidan from Undaria pinnatifida Sporophylls in Mouse Hepatocarcinoma Hca-F Cells. PLoS ONE 2014, 9, e106071. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.M.; Kim, W.J.; Moon, S.K. AKT signaling is involved in fucoidan-induced inhibition of growth and migration of human bladder cancer cells. Food Chem. Toxicol. 2014, 64, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Leite, R.; Carreira, I.; Melo, J.; Ferreira, S.I.; Ribeiro, I.; Ferreira, J.; Filipe, M.; Bernardo, C.; Arantes-Rodrigues, R.; Oliveira, P.; et al. Genomic characterization of three urinary bladder cancer cell lines: Understanding genomic types of urinary bladder cancer. Tumor Biol. 2014, 35, 4599–4617. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Lee, J.H.; Lee, S.H. Antitumor Effects of Fucoidan on Human Colon Cancer Cells via Activation of Akt Signaling. Biomol. Ther. 2015, 23, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, V.; Park, Y.; Chen, C.C.; Xu, P.Z.; Chen, M.L.; Tonic, I.; Unterman, T.; Hay, N. Akt Determines Replicative Senescence and Oxidative or Oncogenic Premature Senescence and Sensitizes Cells to Oxidative Apoptosis. Cancer Cell 2008, 14, 458–470. [Google Scholar] [CrossRef]
- Los, M.; Maddika, S.; Erb, B.; Schulze-Osthoff, K. Switching Akt: From survival signaling to deadly response. Bioessays 2009, 31, 492–495. [Google Scholar] [CrossRef]
- Liu, J.H.; Babaian, D.C.; Liebert, M.; Steck, P.A.; Kagan, J. Inactivation of MMAC1 in bladder transitional-cell carcinoma cell lines and specimens. Mol. Carcinogen. 2000, 29, 143–150. [Google Scholar] [CrossRef]
- Hsu, C.P.; Kao, T.Y.; Chang, W.L.; Nieh, S.; Wang, H.L.; Chung, Y.C. Clinical significance of tumor suppressor PTEN in colorectal carcinoma. Eur. J. Surg. Oncol. EJSO 2011, 37, 140–147. [Google Scholar] [CrossRef]
- Vlietstra, R.J.; van Alewijk, D.; Hermans, K.G.L.; van Steenbrugge, G.J.; Trapman, J. Frequent inactivation of PTEN in prostate cancer cell lines and xenografts. Cancer Res. 1998, 58, 2720–2723. [Google Scholar] [PubMed]
- Yap, T.A.; Walton, M.I.; Grimshaw, K.M.; te Poele, R.H.; Eve, P.D.; Valenti, M.R.; Brandon, A.K.D.; Martins, V.; Zetterlund, A.; Heaton, S.P.; et al. AT13148 Is a Novel, Oral Multi-AGC Kinase Inhibitor with Potent Pharmacodynamic and Antitumor Activity. Clin. Cancer Res. 2012, 18, 3912–3923. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Gossett, D.R.; Wang, S.; Yang, D.; Cao, Y.; Chen, J.; Guo, R.; Reynolds, R.K.; Lin, J. Inhibition of AKT survival pathway by a small molecule inhibitor in human endometrial cancer cells. Br. J. Cancer 2004, 91, 1808–1812. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, L.; Kim, M.K.; Lyle, L.T.; Bunch, K.P.; House, C.D.; Ning, F.; Noonan, A.M.; Annunziata, C.M. Characterization of ovarian cancer cell lines as in vivo models for preclinical studies. Gynecol. Oncology 2016, 142, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Cellai, C.; Laurenzana, A.; Bianchi, E.; Sdelci, S.; Manfredini, R.; Vannucchi, A.M.; Caporale, R.; Balliu, M.; Mannelli, F.; Ferrari, S.; et al. Mechanistic insight into WEB-2170-induced apoptosis in human acute myelogenous leukemia cells: The crucial role of PTEN. Exp. Hematol. 2009, 37, 1176–1185. [Google Scholar] [CrossRef]
- Ioffe, Y.J.; Chiappinelli, K.B.; Mutch, D.G.; Zighelboim, I.; Goodfellow, P.J. Phosphatase and tensin homolog (PTEN) pseudogene expression in endometrial cancer: A conserved regulatory mechanism important in tumorigenesis? Gynecol. Oncol. 2012, 124, 340–346. [Google Scholar] [CrossRef]
- Milella, M.; Falcone, I.; Conciatori, F.; Matteoni, S.; Sacconi, A.; De Luca, T.; Bazzichetto, C.; Corbo, V.; Simbolo, M.; Sperduti, I.; et al. PTEN status is a crucial determinant of the functional outcome of combined MEK and mTOR inhibition in cancer. Sci. Rep. 2017, 7, 43013. [Google Scholar] [CrossRef]
- Yin, J.X.; Li, X.X.; Zhang, Z.L.; Luo, X.H.; Wang, L.; Liu, L. SPAG6 silencing induces apoptosis in the myelodysplastic syndrome cell line SKM-1 via the PTEN/PI3K/AKT signaling pathway in vitro and in vivo. Int. J. Oncol. 2018, 53, 297–306. [Google Scholar] [CrossRef]
- Teimourian, S.; Moghanloo, E. Role of PTEN in neutrophil extracellular trap formation. Mol. Immunol. 2015, 66, 319–324. [Google Scholar] [CrossRef]
- Philip, C.A.; Laskov, I.; Beauchamp, M.C.; Marques, M.; Amin, O.; Bitharas, J.; Kessous, R.; Kogan, L.; Baloch, T.; Gotlieb, W.H.; et al. Inhibition of PI3K-AKT-mTOR pathway sensitizes endometrial cancer cell lines to PARP inhibitors. BMC Cancer 2017, 17, 638. [Google Scholar] [CrossRef]
- Williamson, R.M.; Jackson, J.H. Common evolutionary origin of the ilvgmeda attenuation locus and transfer rna1-leu in Escherichia-coli. J. Bacteriol. 1987, 169, 2893–2895. [Google Scholar] [CrossRef] [PubMed]
- Saal, L.H.; Gruvberger-Saal, S.K.; Persson, C.; Loevgren, K.; Jumppanen, M.; Staaf, J.; Jonsson, G.; Pires, M.M.; Maurer, M.; Holm, K.; et al. Recurrent gross mutations of the PTEN tumor suppressor gene in breast cancers with deficient DSB repair. Nat. Genet. 2008, 40, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Dithmer, M.; Kirsch, A.-M.; Richert, E.; Fuchs, S.; Wang, F.; Schmidt, H.; Coupland, S.; Roider, J.; Klettner, A. Fucoidan Does Not Exert Anti-Tumorigenic Effects on Uveal Melanoma Cell Lines. Mar. Drugs 2017, 15, 193. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.X.; Luo, J.; Mo, J.S.; Liu, G.B.; Kim, Y.C.; Meng, Z.P.; Zhao, L.; Peyman, G.; Ouyang, H.; Jiang, W.; et al. Mutant Gq/11 Promote Uveal Melanoma Tumorigenesis by Activating YAP. Cancer Cell 2014, 25, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Shoushtari, A.N.; Carvajal, R.D. GNAQ and GNA11 mutations in uveal melanoma. Melanoma Res. 2014, 24, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Burotto, M.; Chiou, V.L.; Lee, J.M.; Kohn, E.C. The MAPK Pathway Across Different Malignancies: A New Perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef]
- Millington, G.W.M. Mutations of the BRAF gene in human cancer, by Davies et al. (Nature 2002; 417: 94954). Clin. Exp. Dermatol. 2013, 38, 222–223. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.B.; Kong, N.; Ye, L.; Han, W.D.; Zhou, J.C.; Zhang, Q.; He, C.; Pan, H.M. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014, 344, 174–179. [Google Scholar] [CrossRef]
- Cuadrado, A.; Nebreda, A.R. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar] [CrossRef]
- Brancho, D.; Tanaka, N.; Jaeschke, A.; Ventura, J.J.; Kelkar, N.; Tanaka, Y.; Kyuuma, M.; Takeshita, T.; Flavell, R.A.; Davis, R.J. Mechanism of p38 MAP kinase activation in vivo. Genes Dev. 2003, 17, 1969–1978. [Google Scholar] [CrossRef]
- Katz, M.; Amit, I.; Yarden, Y. Regulation of MAPKs by growth factors and receptor tyrosine kinases. Biochim. Biophys. Acta-Mol. Cell Res. 2007, 1773, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-Sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol. 2005, 78, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.L.; Ge, Y.L.; Zhang, J.Y.; Wang, Q.; Hou, L.; Liu, Y.C.; Sun, L.L.; Li, Q. Anticancer Properties and Mechanisms of Fucoidan on Mouse Breast Cancer In Vitro and In Vivo. PLoS ONE 2012, 7, e43483. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.Y.; Lin, T.Y.; Lu, M.K.; Leng, P.J.; Tsao, S.M.; Wu, Y.C. Fucoidan induces Toll-like receptor 4-regulated reactive oxygen species and promotes endoplasmic reticulum stress-mediated apoptosis in lung cancer. Sci. Rep. 2017, 7, 44990. [Google Scholar] [CrossRef]
- Choo, G.S.; Lee, H.N.; Shin, S.A.; Kim, H.J.; Jung, J.Y. Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression. Mar. Drugs 2016, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.E.; Choi, E.S.; Shin, J.A.; Lee, S.O.; Park, K.S.; Cho, N.P.; Cho, S.D. Fucoidan induces caspase-dependent apoptosis in MC3 human mucoepidermoid carcinoma cells. Exp. Ther. Med. 2014, 7, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.J.; Park, K.S.; Ku, M.J.; Lee, M.S.; Jeong, S.H.; Imbs, T.I.; Zvyagintseva, T.N.; Ermakova, S.P.; Lee, Y.H. Effect of Costatia costata Fucoidan on Expression of Matrix Metalloproteinase-1 Promoter, mRNA, and Protein. J. Nat. Prod. 2009, 72, 1731–1734. [Google Scholar] [CrossRef] [PubMed]
- Tower, G.B.; Coon, C.C.; Benbow, U.; Vincenti, M.P.; Brinckerhoff, C.E. Erk 1/2 differentially regulates the expression from the 1G/2G single nucleotide polymorphism in the MMP-1 promoter in melanoma cells. Biochim. Biophys. Acta-Mol. Basis Dis. 2002, 1586, 265–274. [Google Scholar] [CrossRef]
- Xiu, B.; Zhang, W.J.; Huang, B.B.; Chen, J.D.; Lu, H.N.; Fu, J.F.; Xiong, H.; Liang, A.B. Genetic inhibition of vascular endothelial growth factor receptor-1 significantly inhibits the migration and proliferation of leukemia cells and increases their sensitivity to chemotherapy. Oncol. Rep. 2013, 29, 2030–2038. [Google Scholar] [CrossRef]
- Hotz, B.; Keilholz, U.; Fusi, A.; Buhr, H.J.; Hotz, H.G. In vitro and in vivo antitumor activity of cetuximab in human gastric cancer cell lines in relation to epidermal growth factor receptor (EGFR) expression and mutational phenotype. Gastric Cancer 2012, 15, 252–264. [Google Scholar] [CrossRef]
- Milella, M.; Kornblau, S.M.; Estrov, Z.; Carter, B.Z.; Lapillonne, H.; Harris, D.; Konopleva, M.; Zhao, S.R.; Estey, E.; Andreeff, M. Therapeutic targeting of the MEK/MAPK signal transduction module in acute myeloid leukemia. J. Clin. Investig. 2001, 108, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Hwang, H.J.; Kim, G.Y.; Cha, H.J.; Kim, W.J.; Kim, N.D.; Yoo, Y.H.; Choi, Y.H. Induction of Apoptosis by Fucoidan in Human Leukemia U937 Cells through Activation of p38 MAPK and Modulation of Bcl-2 Family. Mar. Drugs 2013, 11, 2347–2364. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Hirata, Y.; Nakagawa, H.; Sakamoto, K.; Hikiba, Y.; Kinoshita, H.; Nakata, W.; Takahashi, R.; Tateishi, K.; Tada, M.; et al. Apoptosis signal-regulating kinase 1 and cyclin D1 compose a positive feedback loop contributing to tumor growth in gastric cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, M.; Higaki, K.; Nanba, E.; Ikeguchi, M. Anti-Proliferation Activity of Fucoidan in MKN45 Gastric Cancer Cells and Downregulation of Phosphorylated ASK1, a Cell Cycle-Regulated Kinase. Yonago Acta Med. 2015, 58, 1–7. [Google Scholar] [PubMed]
- Min, E.Y.; Kim, I.H.; Lee, J.; Kim, E.Y.; Choi, Y.H.; Nam, T.J. The effects of fucodian on senescence are controlled by the p16(INK4a)-pRb and p14(Arf)-p53 pathways in hepatocellular carcinoma and hepatic cell lines. Int. J. Oncol. 2014, 45, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, B.A.; Restuccia, D.F. PI3K-PKB/Akt Pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011189. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X.D. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The BCL2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X.D. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 1998, 94, 481–490. [Google Scholar] [CrossRef]
- Xue, M.L.; Ji, X.Q.; Xue, C.X.; Liang, H.; Ge, Y.L.; He, X.J.; Zhang, L.; Bian, K.; Zhang, L.C. Caspase-dependent and caspase-independent induction of apoptosis in breast cancer by fucoidan via the PI3K/AKT/GSK3 beta pathway in vivo and in vitro. Biomed. Pharmacother. 2017, 94, 898–908. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds Inhibit Proliferation of Melanoma Cells and Induce Apoptosis by Activation of Caspase-3 in Vitro. Mar. Drugs 2011, 9, 2605–2621. [Google Scholar] [CrossRef] [PubMed]
- Boucher, M.J.; Morisset, J.; Vachon, P.H.; Reed, J.C.; Laine, J.; Rivard, N. MEK/ERK signaling pathway regulates the expression of Bcl-2, Bcl-X-L, and Mcl-1 and promotes survival of human pancreatic cancer cells. J. Cell. Biochem. 2000, 79, 355–369. [Google Scholar] [CrossRef]
- Dijkers, P.F.; Medema, R.H.; Lammers, J.W.J.; Koenderman, L.; Coffer, P.J. Expression of the pro-apoptotic Bcl-2 family member Bim is regulated by the forkhead transcription factor FKHR-L1. Curr. Biol. 2000, 10, 1201–1204. [Google Scholar] [CrossRef]
- Dijkers, P.F.; Birkenkamp, K.U.; Lam, E.W.F.; Thomas, N.S.B.; Lammers, J.W.J.; Koenderman, L.; Coffer, P.J. FKHR-L1 can act as a critical effector of cell death induced by cytokine withdrawal: Protein kinase B-enhanced cell survival through maintenance of mitochondrial integrity. J. Cell Biol. 2002, 156, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Park, S.Y.; Lee, J.Y.; Park, J.H.Y. Fucoidan present in brown algae induces apoptosis of human colon cancer cells. BMC Gastroenterol. 2010, 10, 96. [Google Scholar] [CrossRef]
- Colak, S.; ten Dijke, P. Targeting TGF-beta Signaling in Cancer. Trends Cancer 2017, 3, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Buzdar, A.U. Fulvestrant—A new type of estrogen receptor antagonist for the treatment of advanced breast cancer. Drugs Today 2004, 40, 751–764. [Google Scholar] [CrossRef]
- Hasan, J.; Jayson, G.C. VEGF antagonists. Expert Opin. Biol. Ther. 2001, 1, 703–718. [Google Scholar]
- Massague, J. TGF beta in cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef]
- Kim, T.H.; Lee, E.K.; Lee, M.J.; Kim, J.H.; Yang, W.S. Fucoidan inhibits activation and receptor binding of transforming growth factor-beta 1. Biochem. Biophys. Res. Commun. 2013, 432, 163–168. [Google Scholar] [CrossRef]
- Hong, S.W.; Jung, K.H.; Lee, H.S.; Zheng, H.M.; Choi, M.J.; Lee, C.; Hong, S.S. Suppression by Fucoidan of Liver Fibrogenesis via the TGF-beta/Smad Pathway in Protecting against Oxidative Stress. Biosci. Biotechnol. Biochem. 2011, 75, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.D.; Yao, C.J.; Chow, J.M.; Chang, C.L.; Hwang, P.A.; Chuang, S.E.; Whang-Peng, J.; Lai, G.M. Fucoidan Elevates MicroRNA-29b to Regulate DNMT3B-MTSS1 Axis and Inhibit EMT in Human Hepatocellular Carcinoma Cells. Mar. Drugs 2015, 13, 6099–6116. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.Y.; Lin, T.Y.; Hwang, P.A.; Tseng, L.M.; Chen, R.H.; Tsao, S.M.; Hsu, J. Fucoidan induces changes in the epithelial to mesenchymal transition and decreases metastasis by enhancing ubiquitin-dependent TGF beta receptor degradation in breast cancer. Carcinogenesis 2013, 34, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.Y.; Lin, T.Y.; Wu, Y.C.; Tsao, S.M.; Hwang, P.A.; Shih, Y.W.; Hsu, J. Fucoidan inhibition of lung cancer in vivo and in vitro: Role of the Smurf2-dependent ubiquitin proteasome pathway in TGF beta receptor degradation. Oncotarget 2014, 5, 7870–7885. [Google Scholar] [CrossRef] [PubMed]
- Urist, M.R. Bone morphogenetic protein: The molecularization of skeletal system development. J. Bone Miner. Res. 1997, 12, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Chen, H.J.; Zhang, L.; Qin, Y.; Cong, Q.F.; Wang, P.P.; Ding, K. A fucoidan from Nemacystus decipiens disrupts angiogenesis through targeting bone morphogenetic protein 4. Carbohydr. Polym. 2016, 144, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Kang, H.J.; Park, J.Y.; Lee, J. Fucoidan promotes osteoblast differentiation via JNK- and ERK-dependent BMP2-Smad 1/5/8 signaling in human mesenchymal stem cells. Exp. Mol. Med. 2015, 47, e128. [Google Scholar] [CrossRef]
- Cho, Y.S.; Jung, W.K.; Kim, J.A.; Choi, I.W.; Kim, S.K. Beneficial effects of fucoidan on osteoblastic MG-63 cell differentiation. Food Chem. 2009, 116, 990–994. [Google Scholar] [CrossRef]
- Hwang, P.A.; Hung, Y.L.; Phan, N.N.; Hieu, B.T.N.; Chang, P.M.; Li, K.L.; Lin, Y.C. The in vitro and in vivo effects of the low molecular weight fucoidan on the bone osteogenic differentiation properties. Cytotechnology 2016, 68, 1349–1359. [Google Scholar] [CrossRef]
- Trinh, X.B.; Tjalma, W.A.A.; Vermeulen, P.B.; Van den Eynden, G.; Van der Auwera, I.; Van Laere, S.J.; Helleman, J.; Berns, E.; Dirix, L.Y.; van Dam, P.A. The VEGF pathway and the AKT/mTOR/p70S6K1 signalling pathway in human epithelial ovarian cancer. Br. J. Cancer 2009, 100, 971–978. [Google Scholar] [CrossRef]
- Doanes, A.M.; Hegland, D.D.; Sethi, R.; Kovesdi, I.; Bruder, J.T.; Finkel, T. VEGF stimulates MAPK through a pathway that is unique for receptor tyrosine kinases. Biochem. Biophys. Res. Commun. 1999, 255, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Nam, J.O.; Jean, C.; Lawson, C.; Walsh, C.T.; Goka, E.; Lim, S.T.; Tomar, A.; Tancioni, I.; Uryu, S.; et al. VEGF-Induced Vascular Permeability Is Mediated by FAK. Dev. Cell 2012, 22, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, J.; Chang, A.K.; Liu, B.; Yang, L.L.; Li, Q.M.; Wang, P.S.; Zou, X.Y. Fucoidan extract derived from Undaria pinnatifida inhibits angiogenesis by human umbilical vein endothelial cells. Phytomedicine 2012, 19, 797–803. [Google Scholar] [CrossRef]
- Dithmer, M.; Fuchs, S.; Shi, Y.; Schmidt, H.; Richert, E.; Roider, J.; Klettner, A. Fucoidan Reduces Secretion and Expression of Vascular Endothelial Growth Factor in the Retinal Pigment Epithelium and Reduces Angiogenesis In Vitro. PLoS ONE 2014, 9, e89150. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Z.; Gao, Z.X.; Ma, Y.H.; Teng, H.M.; Liu, Z.D.; Wei, H.Y.; Lu, Y.B.; Cheng, X.F.; Hou, L.; Zou, X.Y. Fucoidan inhibits lymphangiogenesis by downregulating the expression of VEGFR3 and PROX1 in human lymphatic endothelial cells. Oncotarget 2016, 7, 38025–38035. [Google Scholar] [CrossRef]
- Liu, F.; Luo, G.P.; Xiao, Q.; Chen, L.P.; Luo, X.H.; Lv, J.L.; Chen, L.X. Fucoidan inhibits angiogenesis induced by multiple myeloma cells. Oncol. Rep. 2016, 36, 1963–1972. [Google Scholar] [CrossRef]
- Cong, Q.F.; Chen, H.J.; Liao, W.F.; Xiao, F.; Wang, P.P.; Qin, Y.; Dong, Q.; Ding, K. Structural characterization and effect on anti-angiogenic activity of a fucoidan from Sargassum fusiforme. Carbohydr. Polym. 2016, 136, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Cong, Q.F.; Du, Z.Y.; Liao, W.F.; Zhang, L.; Yao, Y.L.; Ding, K. Sulfated fucoidan FP08S2 inhibits lung cancer cell growth in vivo by disrupting angiogenesis via targeting VEGFR2/VEGF and blocking VEGFR2/Erk/VEGF signaling. Cancer Lett. 2016, 382, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.L.; Schmidt, H.; Pavleska, D.; Wermann, T.; Seekamp, A.; Fuchs, S. Crude Fucoidan Extracts Impair Angiogenesis in Models Relevant for Bone Regeneration and Osteosarcoma via Reduction of VEGF and SDF-1. Mar. Drugs 2017, 15, 186. [Google Scholar] [CrossRef]
- Lake, A.C.; Vassy, R.; Di Benedetto, M.; Lavigne, D.; Le Visage, C.; Perret, G.Y.; Letourneur, D. Low molecular weight fucoidan increases VEGF(165)-induced endothelial cell migration by enhancing VEGF(165) binding to VEGFR-2 and NRP1. J. Biol. Chem. 2006, 281, 37844–37852. [Google Scholar] [CrossRef]
- Kim, B.S.; Yang, S.S.; You, H.K.; Shin, H.I.; Lee, J. Fucoidan-induced osteogenic differentiation promotes angiogenesis by inducing vascular endothelial growth factor secretion and accelerates bone repair. J. Tissue Eng. Regener. Med. 2018, 12, E1311–E1324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Teruya, K.; Yoshida, T.; Eto, H.; Shirahata, S. Fucoidan Extract Enhances the Anti-Cancer Activity of Chemotherapeutic Agents in MDA-MB-231 and MCF-7 Breast Cancer Cells. Mar. Drugs 2013, 11, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Banafa, A.M.; Roshan, S.; Liu, Y.Y.; Chen, H.J.; Chen, M.J.; Yang, G.X.; He, G.Y. Fucoidan induces G(1) phase arrest and apoptosis through caspases-dependent pathway and ROS induction in human breast cancer MCF-7 cells. J. Huazhong Univ. Sci. Technol.-Med. Sci. 2013, 33, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Zhao, Y.; Zhang, Y.; Zhang, D.H. Fucoidan Induces Cancer Cell Apoptosis by Modulating the Endoplasmic Reticulum Stress Cascades. PLoS ONE 2014, 9, e108157. [Google Scholar] [CrossRef]
- Wu, S.Y.; Yan, M.D.; Wu, A.T.H.; Yuan, K.S.P.; Liu, S.H. Brown Seaweed Fucoidan Inhibits Cancer Progression by Dual Regulation of mir-29c/ADAM12 and miR-17-5p/PTEN Axes in Human Breast Cancer Cells. J. Cancer 2016, 7, 2408–2419. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki-Miyamoto, Y.; Yamasaki, M.; Tachibana, H.; Yamada, K. Fucoidan Induces Apoptosis through Activation of Caspase-8 on Human Breast Cancer MCF-7 Cells. J. Agric. Food Chem. 2009, 57, 8677–8682. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, Q.Q.; Kong, Y.Y.; Xie, B.Q.; Gao, M.J.; Tao, Y.; Xu, H.W.; Zhan, F.H.; Dai, B.J.; Shi, J.M.; et al. Antitumor activity of fucoidan against diffuse large B cell lymphoma in vitro and in vivo. Acta Biochim. Biophys. Sin. 2015, 47, 925–931. [Google Scholar] [CrossRef]
- Schneider, T.; Ehrig, K.; Liewert, I.; Alban, S. Interference with the CXCL12/CXCR4 axis as potential antitumor strategy: Superiority of a sulfated galactofucan from the brown alga Saccharina latissima and Fucoidan over heparins. Glycobiology 2015, 25, 812–824. [Google Scholar] [CrossRef]
- Ishikawa, C.; Mori, N. In vitro and in vivo anti-primary effusion lymphoma activities of fucoidan extracted from Cladosiphon okamuranus Tokida. Oncol. Rep. 2017, 38, 3197–3204. [Google Scholar] [CrossRef]
- Haneji, K.; Matsuda, T.; Tomita, M.; Kawakami, H.; Ohshiro, K.; Uchihara, J.N.; Masuda, M.; Takasu, N.; Tanaka, Y.; Ohta, T.; et al. Fucoidan extracted from Cladosiphon okamuranus tokida induces apoptosis of human T-cell leukemia virus type 1-infected T-cell lines and primary adult T-cell leukemia cells. Nutr. Cancer 2005, 52, 189–201. [Google Scholar] [CrossRef]
- Ye, J.; Li, Y.P.; Teruya, K.; Katakura, Y.; Ichikawa, A.; Eto, H.; Hosoi, M.; Nishimoto, S.; Shirahata, S. Enzyme-digested fucoidan extracts derived from seaweed Mozuku of Cladosiphon novae-caledoniae kylin inhibit invasion and angiogenesis of tumor cells. Cytotechnology 2005, 47, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Zhurishkina, E.V.; Stepanov, S.I.; Shvetsova, S.V.; Kulminskaya, A.A.; Lapina, I.M. A comparison of the effect of fucoidan from alga Fucus vesiculosus and its fractions obtained by anion-exchange chromatography on HeLa G-63, Hep G2, and Chang liver cells. Cell Tissue Biol. 2017, 11, 242–249. [Google Scholar] [CrossRef]
- Alekseyenko, T.V.; Zhanayeva, S.Y.; Venediktova, A.A.; Zvyagintseva, T.N.; Kuznetsova, T.A.; Besednova, N.N.; Korolenko, T.A. Antitumor and antimetastatic activity of fucoidan, a sulfated polysaccharide isolated from the Okhotsk sea Fucus evanescens brown alga. Bull. Exp. Biol. Med. 2007, 143, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Nagamine, T. Effect of Fucoidan on the Biotinidase Kinetics in Human Hepatocellular Carcinoma. Anticancer Res. 2009, 29, 1211–1217. [Google Scholar]
- Song, Y.F.; Wang, Q.K.; Wang, Q.J.; He, Y.H.; Ren, D.D.; Liu, S.; Wu, L. Structural characterization and antitumor effects of fucoidans from brown algae Kjellmaniella crassifolia farmed in northern China. Int. J. Biol. Macromol. 2018, 119, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Vishchuk, O.S.; Sun, H.M.; Wang, Z.; Ermakova, S.P.; Xiao, J.J.; Lu, T.; Xue, P.P.; Zvyagintseva, T.N.; Xiong, H.; Shao, C.; et al. PDZ-binding kinase/T-LAK cell-originated protein kinase is a target of the fucoidan from brown alga Fucus evanescens in the prevention of EGF-induced neoplastic cell transformation and colon cancer growth. Oncotarget 2016, 7, 18763–18773. [Google Scholar] [CrossRef]
- Chen, L.M.; Liu, P.Y.; Chen, Y.A.; Tseng, H.Y.; Shen, P.C.; Hwang, P.A.; Hsu, H.L. Oligo-Fucoidan prevents IL-6 and CCL2 production and cooperates with p53 to suppress ATM signaling and tumor progression. Sci. Rep. 2017, 7, 11864. [Google Scholar] [CrossRef]
- Park, H.Y.; Park, S.H.; Jeong, J.W.; Yoon, D.; Han, M.H.; Lee, D.S.; Choi, G.; Yim, M.J.; Lee, J.M.; Kim, D.H.; et al. Induction of p53-Independent Apoptosis and G1 Cell Cycle Arrest by Fucoidan in HCT116 Human Colorectal Carcinoma Cells. Mar. Drugs 2017, 15, 154. [Google Scholar] [CrossRef]
- Lv, J.L.; Xiao, Q.; Wang, L.; Liu, X.; Wang, X.; Yang, Z.S.; Zhang, H.B.; Dong, P.J. Fucoidan prevents multiple myeloma cell escape from chemotherapy-induced drug cytotoxicity. Fitoterapia 2013, 84, 257–263. [Google Scholar] [CrossRef]
- Rui, X.; Pan, H.F.; Shao, S.L.; Xu, X.M. Anti-tumor and anti-angiogenic effects of Fucoidan on prostate cancer: Possible JAK-STAT3 pathway. BMC Complement. Altern. Med. 2017, 17, 378. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.B.; Trotman, L.C.; Shaffer, D.; Lin, H.K.; Dotan, Z.A.; Niki, M.; Koutcher, J.A.; Scher, H.I.; Ludwig, T.; Gerald, W.; et al. Crucial role of p53-dependent cellular senescence in suppression of Pten-deficient tumorigenesis. Nature 2005, 436, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Dickinson, J.L. Fucoidan and cancer: A multifunctional molecule with anti-tumor potential. Mar. Drugs 2015, 13, 2327–2346. [Google Scholar] [CrossRef] [PubMed]
- Pugazhenthi, S.; Nesterova, A.; Sable, C.; Heidenreich, K.A.; Boxer, L.M.; Heasley, L.E.; Reusch, J.E.B. Akt/protein kinase B up-regulates Bcl-2 expression through cAMP-response element-binding protein. J. Biol. Chem. 2000, 275, 10761–10766. [Google Scholar] [CrossRef]
- Guicciardi, M.E.; Gores, G.J. Life and death by death receptors. FASEB J. 2009, 23, 1625–1637. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Bach, D.H.; Park, H.J.; Lee, S.K. The Dual Role of Bone Morphogenetic Proteins in Cancer. Mol. Ther.-Oncolytics 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, T.J.; Meier, J.B.; Jansen, V.M. Current Landscape of Targeted Therapies for Hormone-Receptor Positive, HER2 Negative Metastatic Breast Cancer. Front. Oncol. 2018, 8, 308. [Google Scholar] [CrossRef]
- Duygu, A.; Canfeza, S.; Ruchan, U.; Buket, K. Cytotoxic, apoptotic and molecular effects of fucoidan as single agent and in combination with trastuzumab and docetaxel in HER2(+) breast cancer cells. J. Clin. Oncol. 2015, 33, e13520. [Google Scholar] [CrossRef]
- Tocaciu, S.; Oliver, L.J.; Lowenthal, R.M.; Peterson, G.M.; Patel, R.; Shastri, M.; McGuinness, G.; Olesen, I.; Fitton, J.H. The Effect of Undaria pinnatifida Fucoidan on the Pharmacokinetics of Letrozole and Tamoxifen in Patients With Breast Cancer. Integr. Cancer Ther. 2018, 17, 99–105. [Google Scholar] [CrossRef]
- Andrade, E.L.; Bento, A.F.; Cavalli, J.; Oliveira, S.K.; Freitas, C.S.; Marcon, R.; Schwanke, R.C.; Siqueira, J.M.; Calixto, J.B. Non-clinical studies required for new drug development—Part I: Early in silico and in vitro studies, new target discovery and validation, proof of principles and robustness of animal studies. Braz. J. Med. Biol. Res. 2016, 49. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cancer Type | Cell Line | Fucoidan | Mechanism | Research Methods | References |
|---|---|---|---|---|---|
| Breast cancer | MDA-MB-231 and MCF-7 | Fucus vesiculosus | Inhibiting proliferation and metastasis | In vitro | [153] |
| MCF-7 | Fucus vesiculosus | Inhibiting proliferation, inducing cell cycle arrest, and inducing apoptosis | In vitro | [173,174] | |
| MDA-MB-231 | Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [174] | |
| MDA-MB-231 and MCF-7 | Sargassum hemiphyllum | Inhibiting proliferation, inducing cell cycle arrest, and inducing apoptosis | In vitro | [175] | |
| MDA-MB-231 and MCF-7 | Fucus vesiculosus (extract) | Inhibiting proliferation and inducing apoptosis | In vitro | [91] | |
| MCF-7 | Not stated (supposed Fucus vesiculosus) | Inhibiting proliferation and inducing apoptosis | In vitro | [176] | |
| MCF-7 | Fucus vesiculosus (derivatives) | inhibiting proliferation | In vitro | [57] | |
| T47D | Fucus vesiculosus (extract) | Inhibiting proliferation and inducing apoptosis | In vitro | [91] | |
| B-cell lymphoma | HS-sultan and IM-9 | Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [122] |
| DLBCL lines | Fucus vesiculosus | Inhibiting proliferation, inducing cell cycle arrest, and inducing apoptosis | In vitro and In vivo | [177] | |
| Raji cells | Saccharina latissima and Fucus vesiculosus | Inhibiting metastasis | In vitro | [178] | |
| BCBL-1 and TY-1 | Cladosiphon okamuranus | Inhibiting proliferation and inducing apoptosis | In vitro and In vivo | [179] | |
| T-cell lymphoma | MOLT-4 | Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [122] |
| MT-2, MT-4, HUT-102, and MT-1 | Cladosiphon okamuranus | Inhibiting growth and inducing apoptosis | In vitro | [180] | |
| Fibroblastic sarcoma | HT 1080 | Cladosiphon novae-caledoniae (extract) | Inhibiting metastasis | In vitro | [181] |
| Uterine sarcoma | HeLa | Cladosiphon novae-caledoniae (extract) | Inhibiting metastasis | In vitro | [181] |
| HeLa | Fucus vesiculosus (derivatives) | inhibiting proliferation | In vitro | [57] | |
| HeLa | Fucus vesiculosus (fractions) | Inhibiting proliferation, growth, and inducing apoptosis | In vitro | [182] | |
| MES-SA | Fucus vesiculosus (extract) | Inhibiting proliferation and inducing apoptosis | In vitro | [91] | |
| Lung cancer | LLC1 | Fucus evanescens | Inhibiting proliferation and metastasis | In vitro | [183] |
| LLC1 | Sargassum sp. And Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [184] | |
| A549 | Fucus vesiculosus | Inhibiting metastasis | In vitro | [92] | |
| LLC1, A549, and CL1-5 | Fucus vesiculosus | Inhibiting proliferation, metastasis and inducing apoptosis | In vitro and In vivo | [154] | |
| LLC1 | Fucus vesiculosus | Inhibiting proliferation and metastasis | In vitro and In vivo | [70] | |
| A549 | Sargassum fusiforme (sulfated extract) | Inhibiting proliferation and metastasis | In vitro and In vivo | [168] | |
| A549, LLC1, and CL1-5 | Fucus vesiculosus and Saccharina Japonica | Inhibiting proliferation and inducing apoptosis | In vitro and In vivo | [124] | |
| Hepatocellular carcinoma | HuH-6 | Cladosiphon okamuranus | Inhibiting biotinidase activity | In vitro | [185] |
| Huh-6, HUH-7, SK-Hep1, and HepG2 | Sargassum hemiphyllum | Inhibiting proliferation and metastasis | In vitro | [152] | |
| HepG2 | Fucus vesiculosus (fractions) | Inhibiting proliferation, growth and inducing apoptosis | In vitro | [182] | |
| Hca-F | Kjellmaniella crassifolia | inhibiting proliferation | In vitro | [186] | |
| Colorectal cancer | HCT-15 | Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [87] |
| HT-29 and HCT-116 | Fucus vesiculosus | Inhibiting proliferation and metstasis | In vitro | [145] | |
| HCT-116 | Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [174] | |
| HT-29 | Fucus vesiculosus | Inhibiting proliferation, inducing cell cycle arrest, and inducing apoptosis | In vitro and In vivo | [96] | |
| HCT-116, HT-29, and WiDr | Fucus evanescens | Inhibiting colony formation and growth | In vitro and In vivo | [187] | |
| HCT116 | Sargassum hemiphyllum (LMWF, oligo-fucoidan) | Inhibiting proliferation, inducing cell cycle arrest, and inducing apoptosis | In vitro and In vivo | [188] | |
| HCT116 | Fucus vesiculosus | Inhibiting proliferation, inducing cell cycle arrest, and inducing apoptosis | In vitro | [189] | |
| Keratinocytes | HaCaT | Costaria costata | Inhibiting metastasis | In vitro | [127] |
| Melanoma | B16 | Sargassum sp. And Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [184] |
| Bladder cancer | 5637 and T-24 | Not stated | Inhibiting proliferation, growth and inducing cell cycle arrest | In vitro | [94] |
| T-24 | Sargassum hemiphyllum | Inhibiting angiogenesis | In vitro and In vivo | [45] | |
| 5637 | Fucus vesiculosus | induction apoptosis | In vitro | [88] | |
| Plasma cell myeloma | RPMI8226 and U266 | Not stated | Inhibiting angiogenesis | In vitro | [190] |
| Leukemia | U937, HL60, K562, THP1 | Fucus vesiculosus | Inhibiting proliferation and inducing apoptosis | In vitro | [132] |
| NB4, HL60, and K562 | Fucus vesiculosus | Inhibiting proliferation and inducing cell cycle arrest | In vitro and In vivo | [69] | |
| SKM-1 | Not stated (supposed Fucus vesiculosus) | Inhibiting proliferation and inducing apoptosis | In vitro | [90] | |
| Stomach cancer | MKN45 | Cladosiphon okamuranus | Inhibiting proliferation and inducing cell cycle arrest | In vitro | [133] |
| Pancreatic cancer | MiaPaCa-2 and Panc-1 | Turbinaria conoides | Inhibiting proliferation, metastasis and inducing apoptosis | In vitro and ex vivo | [49] |
| Ovarian cancer | OVCAR-3 | Fucus vesiculosus (extract) | Inhibiting proliferation and inducing apoptosis | In vitro | [91] |
| Endometrium carcinoma | HEC-1B, RL95-2, and AN3CA | Fucus vesiculosus (extract) | Inhibiting proliferation and inducing apoptosis | In vitro | [91] |
| Prostate cancer | DU-145 | Not stated (supposed Fucus vesiculosus) | Inhibiting proliferation and metastasis | In vitro and In vivo | [191] |
| Osteosarcoma | MG63 | Fucus vesiculosus | Inhibiting angiogenesis | In vitro | [169] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Weelden, G.; Bobiński, M.; Okła, K.; Van Weelden, W.J.; Romano, A.; Pijnenborg, J.M.A. Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms. Mar. Drugs 2019, 17, 32. https://doi.org/10.3390/md17010032
Van Weelden G, Bobiński M, Okła K, Van Weelden WJ, Romano A, Pijnenborg JMA. Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms. Marine Drugs. 2019; 17(1):32. https://doi.org/10.3390/md17010032
Chicago/Turabian StyleVan Weelden, Geert, Marcin Bobiński, Karolina Okła, Willem Jan Van Weelden, Andrea Romano, and Johanna M. A. Pijnenborg. 2019. "Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms" Marine Drugs 17, no. 1: 32. https://doi.org/10.3390/md17010032
APA StyleVan Weelden, G., Bobiński, M., Okła, K., Van Weelden, W. J., Romano, A., & Pijnenborg, J. M. A. (2019). Fucoidan Structure and Activity in Relation to Anti-Cancer Mechanisms. Marine Drugs, 17(1), 32. https://doi.org/10.3390/md17010032