Seaweed Bioactive Compounds against Pathogens and Microalgae: Potential Uses on Pharmacology and Harmful Algae Bloom Control



Abstract
1. Introduction
2. Cyanobacteria
2.1. General Characteristics
2.2. Blooms of Cyanobacteria
2.3. Undesirable Effects of Cyanobacteria Blooms
2.4. Methods Applied in Cyanobacterial Bloom Control
3. Macroalgae
3.1. General Characteristics
3.2. Potential Use of Macroalgal Compounds
3.2.1. Production of Antimicrobial Substances
3.2.2. Antioxidant Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Catherine, Q.; Susanna, W.; Isidora, E.S.; Mark, H.; Aurelie, V.; Jean-Franeois, H. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef] [PubMed]
- Seder-Colomina, M.; Burgos, A.; Maldonado, J.; Solé, A.; Esteve, I. The effect of copper on different phototrophic microorganisms determined in vivo and at cellular level by confocal laser microscopy. Ecotoxicology 2013, 22, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, G.; Huisman, J.; Matthijs, H.C.P. Potassium sensitivity differs among strains of the harmful cyanobacterium Microcystis and correlates with the presence of salt tolerance genes. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Holdren, C.; Jones, W.; Taggart, J.; Holdren, C.W.; Jones, J. Taggart Managing Lakes and Reservoirs; Terrene Institute: Alexandria, VA, USA, 2001. [Google Scholar]
- Visser, P.M.; Ibelings, B.W.; Mur, L.R.; Walsby, A.E. The Ecophysiology of the Harmful Cyanobacterium Microcystis. Harmful Cyanobact. 2005, 3, 109–142. [Google Scholar] [CrossRef]
- Demeke, A. Cyanobacteria blooms and biological control Methods. Int. J. Fauna Boil. Stud. 2016, 3, 32–38. [Google Scholar]
- Pajdak-Stós, A.; Fiałkowska, E.; Fyda, J. Phormidium autumnale (Cyanobacteria) defense against three ciliate grazer species. Aquat. Microb. Ecol. 2001, 23, 237–244. [Google Scholar] [CrossRef]
- Jeong, J.H.; Jin, H.J.; Sohn, C.H.; Suh, K.H.; Hong, Y.K. Algicidal activity of the seaweed Corallina pilulifera against red tide microalgae. J. Appl. Phycol. 2000, 12, 37–43. [Google Scholar] [CrossRef]
- Gao, L.; Xie, L. Analysis of the influence of meteorological condition on cyanobacterial bloom and treatment methods in Taihu Lake. China Resour. 2011, 29, 35–38. [Google Scholar]
- Schrader, K.K. Natural Algicides for the Control of Cyanobacterial-Related Off-Flavor in Catfish Aquaculture; American Chemical Society: Washington, DC, USA, 2003. [Google Scholar]
- Li, F.M.; Hu, H.Y. Allelopathic effects of different macrophytes on the growth of Microcystis aeruginosa. Allelopath. J. 2005, 15, 145–151. [Google Scholar]
- Chiang, I.-Z.; Huang, W.-Y.; Wu, J.-T. Allelochemicals of botryococcus braunii (chlorophyceae). J. Phycol. 2004, 40, 474–480. [Google Scholar] [CrossRef]
- Della Greca, M.; Fiorentino, A.; Monaco, P.; Pinto, G.; Pollio, A.; Previtera, L. Action of antialgal compounds from Juncus effusus L. on Selenastrum capricornutum. J. Chem. Ecol. 1996, 22, 587–603. [Google Scholar] [CrossRef] [PubMed]
- Meepagala, K.M.; Schrader, K.K.; Wedge, D.E.; Duke, S.O. Algicidal and antifungal compounds from the roots of Ruta graveolens and synthesis of their analogs. Phytochemistry 2005, 66, 2689–2695. [Google Scholar] [CrossRef] [PubMed]
- Belattmania, Z.; Engelen, A.H.; Pereira, H.; Serrão, E.A.; Barakate, M.; Elatouani, S.; Zrid, R.; Bentiss, F.; Chahboun, N.; Reani, A.; et al. Potential uses of the brown seaweed Cystoseira humilis biomass: 2-Fatty acid composition, antioxidant and antibacterial activities. J. Mater. Environ. Sci. 2016, 7, 2074–2081. [Google Scholar]
- Kim, M.S.; Kim, J.Y.; Choi, W.H.; Lee, S.S. Effects of seaweed supplementation on blood glucose concentration, lipid profile, and antioxidant enzyme activities in patients with type 2 diabetes mellitus. Nutr. Res. Pract. 2008, 2, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Kumar, M.; Gupta, V.; Reddy, C.R.K.; Jha, B. Tropical marine macroalgae as potential sources of nutritionally important PUFAs. Food Chem. 2010, 120, 749–757. [Google Scholar] [CrossRef]
- Schwartz, N.; Dobretsov, S.; Rohde, S.; Schupp, P.J. Comparison of antifouling properties of native and invasive Sargassum (Fucales, Phaeophyceae) species. Eur. J. Phycol. 2017, 52, 116–131. [Google Scholar] [CrossRef]
- An, Z.; Wang, Z.; Li, F.; Tian, Z.; Hu, H. Allelopathic inhibition on red tide microalgae Skeletonema costatum by five macroalgal extracts. Front. Environ. Sci. Eng. China 2008, 2, 297–305. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.; Tang, X. Identification of the toxic compounds produced by Sargassum thunbergii to red tide microalgae. Chin. J. Oceanol. Limnol. 2012, 30, 778–785. [Google Scholar] [CrossRef]
- Reviers, B. Biologie et Phylohgénie des Algues; Belin Education: Paris, France, 2003; ISBN 2-7011-3512-5. [Google Scholar]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful Freshwater Algal Blooms, with an Emphasis on Cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Seckbach, J. Algae and Cyanobacteria in Extreme Environments; Springer Science & Business Media: Berlin, Germany, 2007. [Google Scholar]
- Rowell, P.; Kerby, N.W. Cyanobacteria and Their Toxins; Study Plant Science: London, UK, 1991. [Google Scholar]
- Stanier, R.Y.; Deruelles, J.; Rippka, R.; Herdman, M.; Waterbury, J.B. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Schopf, J.W. The Fossil Record: Tracing the Roots of the Cyanobacterial Lineage. In The Ecology of Cyanobacteria; Potts, M., Whitton, B.A., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 13–35. [Google Scholar]
- Zhang, C.C.; Laurent, S.; Sakr, S.; Peng, L.; Bédu, S. Heterocyst differentiation and pattern formation in cyanobacteria: A chorus of signals. Mol. Microbiol. 2006, 59, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, Y.; Khudyakov, I.; Fan, Q.; Gao, H.; Ning, D.; Wolk, C.P.; Xu, X. A gene cluster that regulates both heterocyst differentiation and pattern formation in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2007, 66, 1429–1443. [Google Scholar] [CrossRef] [PubMed]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Grant Burgess, J.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Oufdou, K.; Oudra, B. Substances bioactives élaborées par des cyanobactéries isolées de certains écosystèmes aquatiques marocains. Afr. Sci. 2009, 5, 260–279. [Google Scholar] [CrossRef]
- Abdullah, E.; Idris, A.; Saparon, A. Papr reduction using scs-slm technique in stfbc mimo-ofdm. ARPN J. Eng. Appl. Sci. 2017, 12, 3218–3221. [Google Scholar] [CrossRef]
- Jayatissa, L.P.; Silva, E.I.L.; McElhiney, J.; Lawton, L.A. Occurrence of toxigenic cyanobacterial blooms in freshwaters of Sri Lanka. Syst. Appl. Microbiol. 2006, 29, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Neilan, B.A. The molecular genetics of cyanobacterial toxicity as a basis for monitoring water quality and public health risk. Curr. Opin. Biotechnol. 2008, 19, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Leflaive, J.; Ten-Hage, L. Algal and cyanobacterial secondary metabolites in freshwaters: A comparison of allelopathic compounds and toxins. Freshw. Biol. 2007, 52, 199–214. [Google Scholar] [CrossRef]
- Leão, P.N.; Vasconcelos, M.T.S.; Vasconcelos, V.M. Allelopathic activity of cyanobacteria on green microalgae at low cell densities. Eur. J. Phycol. 2009, 44, 347–355. [Google Scholar] [CrossRef]
- Havens, K.E. Cyanobacterial blooms: Effects on aquatic ecosystems. Adv. Exp. Med. Biol. 2008, 619, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Millie, D.F.; Weckman, G.R.; Fahnenstiel, G.L.; Carrick, H.J.; Ardjmand, E.; Young, W.A.; Sayers, M.J.; Shuchman, R.A. Using artificial intelligence for CyanoHAB niche modeling: Discovery and visualization of Microcystis—Environmental associations within western Lake Erie. Can. J. Fish. Aquat. Sci. 2014, 71, 1642–1654. [Google Scholar] [CrossRef]
- Bertani, I.; Obenour, D.R.; Steger, C.E.; Stow, C.A.; Gronewold, A.D.; Scavia, D. Probabilistically assessing the role of nutrient loading in harmful algal bloom formation in western Lake Erie. J. Gt. Lakes Res. 2016, 42, 1184–1192. [Google Scholar] [CrossRef]
- Gao, Y.; Cornwell, J.C.; Stoecker, D.K.; Owens, M.S. Influence of cyanobacteria blooms on sediment biogeochemistry and nutrient fluxes. Limnol. Oceanogr. 2014, 59, 959–971. [Google Scholar] [CrossRef]
- Verspagen, J.M.H.; Van De Waal, D.B.; Finke, J.F.; Visser, P.M.; Van Donk, E.; Huisman, J. Rising CO2 levels will intensify phytoplankton blooms in eutrophic and hypertrophic lakes. PLoS ONE 2014, 9, e104325. [Google Scholar] [CrossRef] [PubMed]
- Sigee, D.C. Freshwater Microbiology: Biodiversity and Dynamic Interactions of Microorganisms in the Aquatic Environment; John Wiley Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Castenholz, R.W. General Characteristics of the Cyanobacteria. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley Sons: Hoboken, NJ, USA, 2001; pp. 1–23. [Google Scholar]
- Paerl, H.W.; Huisman, J. CLIMATE: Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Brucet, S.; Naselli-Flores, L.; Papastergiadou, E.; Stefanidis, K.; Nõges, T.; Nõges, P.; Attayde, J.L.; Zohary, T.; Coppens, J.; et al. Ecological impacts of global warming and water abstraction on lakes and reservoirs due to changes in water level and related changes in salinity. Hydrobiologia 2015, 750, 201–227. [Google Scholar] [CrossRef]
- Brasil, J.; Attayde, J.L.; Vasconcelos, F.R.; Dantas, D.D.F.; Huszar, V.L.M. Drought-induced water-level reduction favors cyanobacteria blooms in tropical shallow lakes. Hydrobiologia 2016, 770, 145–164. [Google Scholar] [CrossRef]
- Masó, M.; Garcés, E. Harmful microalgae blooms (HAB); problematic and conditions that induce them. Mar. Pollut. Bull. 2006, 53, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, N.; Antunes, S.C.; Pereira, M.J.; Gonçalves, F. Seasonal succession of cladocerans and phytoplankton and their interactions in a shallow eutrophic lake (Lake Vela, Portugal). Acta Oecol. 2006, 29, 54–64. [Google Scholar] [CrossRef]
- Zingone, A.; Oksfeldt Enevoldsen, H. The diversity of harmful algal blooms: A challenge for science and management. Ocean Coast. Manag. 2000, 43, 725–748. [Google Scholar] [CrossRef]
- Smith, J.L.; Boyer, G.L.; Zimba, P.V. A review of cyanobacterial odorous and bioactive metabolites: Impacts and management alternatives in aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar] [CrossRef]
- Robin, J.; Cravedi, J.P.; Hillenweck, A.; Deshayes, C.; Vallod, D. Off flavor characterization and origin in French trout farming. Aquaculture 2006, 260, 128–138. [Google Scholar] [CrossRef]
- Jüttner, F.; Watson, S.B. Biochemical and ecological control of geosmin and 2-methylisoborneol in source waters. Appl. Environ. Microbiol. 2007, 73, 4395–4406. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin mixtures and taste-and-odor compounds in cyanobacterial blooms from the midwestern united states. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Oudra, B.; Loudiki, M.; Vasconcelos, V.; Sabour, B.; Sbiyyaa, B.; Oufdou, K.; Mezrioui, N. Detection and quantification of microcystins from cyanobacteria strains isolated from reservoirs and ponds in Morocco. Environ. Toxicol. 2002, 17, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Loudiki, M.; Oudra, B.; Sabour, B.; Sbiyyaa, B.; Vasconcelos, V. Taxonomy and geographic distribution of potential toxic cyanobacterial strains in Morocco. Ann. Limnol. Int. J. Limnol. 2002, 38, 101–108. [Google Scholar] [CrossRef]
- Douma, M.; Ouahid, Y.; Campo, F.F.; del Loudiki, M.; Mouhri, K.; Oudra, B. Identification and quantification of cyanobacterial toxins (microcystins) in two Moroccan drinking-water reservoirs (Mansour Eddahbi, Almassira). Environ. Monit. Assess. 2010, 160, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health 2006, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.J. Policy Analysis Eutrophication of U.S. Freshwaters: Damages. Environ. Sci. Technol. 2009, 43, 8. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Chorus, I. Accumulation of cyanobacterial toxins in freshwater “seafood” and its consequences for public health: A review. Environ. Pollut. 2007, 150, 177–192. [Google Scholar] [CrossRef] [PubMed]
- El Khalloufi, F.; El Ghazali, I.; Saqrane, S.; Oufdou, K.; Vasconcelos, V.; Oudra, B. Phytotoxic effects of a natural bloom extract containing microcystins on Lycopersicon esculentum. Ecotoxicol. Environ. Saf. 2012, 79, 199–205. [Google Scholar] [CrossRef] [PubMed]
- El Khalloufi, F.; Oufdou, K.; Bertrand, M.; Lahrouni, M.; Oudra, B.; Ortet, P.; Barakat, M.; Heulin, T.; Achouak, W. Microbiote shift in the Medicago sativa rhizosphere in response to cyanotoxins extract exposure. Sci. Total Environ. 2016, 539, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Shavyrina, O.B.; Gapochka, L.D.; Azovskii, A.I. Development of tolerance for copper in cyanobacteria repeatedly exposed to its toxic effect. Biol. Bull. 2001, 28, 183–187. [Google Scholar] [CrossRef]
- García-Villada, L.; Rico, M.; Altamirano, M.; Sánchez-Martín, L.; López-Rodas, V.; Costas, E. Occurrence of copper resistant mutants in the toxic cyanobacteria Microcystis aeruginosa: Characterisation and future implications in the use of copper sulphate as algaecide. Water Res. 2004, 38, 2207–2213. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.; Heimann, K.; Quayle, P.; Negri, A.P. Additive toxicity of herbicide mixtures and comparative sensitivity of tropical benthic microalgae. Mar. Pollut. Bull. 2010, 60, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Prosecka, J.; Orlov, A.V.; Fantin, Y.S.; Zinchenko, V.V.; Babykin, M.M.; Tichy, M. A novel ATP-binding cassette transporter is responsible for resistance to viologen herbicides in the cyanobacterium Synechocystis sp. PCC 6803. FEBS J. 2009, 276, 4001–4011. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.J.; Nakano, K.; Matsumura, M. A novel strategy for cyanobacterial bloom control by ultrasonic irradiation. Water Sci. Technol. 2002, 46, 207–215. [Google Scholar] [PubMed]
- Cooke, G.D.; Welch, E.B.; Peterson, S.; Nichols, S.A. Restoration and Management of Lakes and Reservoirs; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Klapper, H. Technologies for lake restoration. J. Limnol. 2003, 62, 73–90. [Google Scholar] [CrossRef]
- Shen, Y.; Liu, Y.; Wu, G.; Ao, H.; Qiu, C. Mechanical removal of heavy cyanobacterial bloom in the hyper-eutrophic lake Dianchi. Acta Hydrobiol. Sin. 2004, 28, 131–136. [Google Scholar]
- Shapiro, J.; Lamarra, V.; Lynch, M. Biomanipulation: An Ecosystem Approach to Lake Restoration; Limnological Research Center, University of Minnesota: Minneapolis, MN, USA, 1975; pp. 85–96. [Google Scholar]
- Van Donk, E.; Van de Bund, W.J. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Erhard, D.; Gross, E.M. Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquat. Bot. 2006, 85, 203–211. [Google Scholar] [CrossRef]
- Park, M.H.; Chung, I.M.; Ahmad, A.; Kim, B.H.; Hwang, S.J. Growth inhibition of unicellular and colonial Microcystis strains (Cyanophyceae) by compounds isolated from rice (Oryza sativa) hulls. Aquat. Bot. 2009, 90, 309–314. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, B.; Wang, J.; Gao, Y.; Wu, Z. Study on the mechanism of allelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte (Myriophyllum spicatum) and its secretion. Aquat. Toxicol. 2010, 98, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Hu, H. Isolation and Characterization of a Novel Antialgal Allelochemical from Phragmites communis. Society 2005, 71, 6545–6553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, S.; Jiao, L.; Zhao, H.; Zhang, Y.; Li, Y. Physiological response of a submerged plant (Myriophyllum spicatum) to different NH4Cl concentrations in sediments. Ecol. Eng. 2013, 58, 91–98. [Google Scholar] [CrossRef]
- Wang, H.; Liang, F.; Qiao, N.; Dong, J.; Zhang, L.; Guo, Y. Chemical Composition of Volatile Oil from Two Emergent Plants and Their Algae Inhibition Activity. Pol. J. Environ. Stud. 2014, 23, 2371–2374. [Google Scholar]
- Dong, J.; Yang, K.; Li, S.; Li, G.; Song, L. Submerged vegetation removal promotes shift of dominant phytoplankton functional groups in a eutrophic lake. J. Environ. Sci. 2014, 26, 1699–1707. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, F.; Zhang, L. Composition and anti-cyanobacterial activity of essential oils from six different submerged macrophytes. Pol. J. Environ. Stud. 2015, 24, 333–338. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Zhang, P.; Zeng, G.; Cai, X.; Liu, S.; Yin, Y.; Hu, X.; Hu, X.; Tan, X. Growth inhibition and oxidative damage of Microcystis aeruginosa induced by crude extract of Sagittaria trifolia tubers. J. Environ. Sci. 2016, 43, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Meng, P.; Pei, H.; Hu, W.; Liu, Z.; Li, X.; Xu, H. Allelopathic effects of Ailanthus altissima extracts on Microcystis aeruginosa growth, physiological changes and microcystins release. Chemosphere 2015, 141, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Najem, A.M.; Abed, I.J.; Al-haidari, A.M.D. Evaluation the Activity of Rosemary (Rosmarinus officinalis L.) Essential Oil Against Some Cyanobacteria. Iraqi J. Biotechnol. 2016, 15, 97–102. [Google Scholar]
- Tellez, M.R.; Dayan, F.E.; Schrader, K.K.; Wedge, D.E.; Duke, S.O. Composition and some biological activities of the essential oil of Callicarpa americana (L.). J. Agric. Food Chem. 2000, 48, 3008–3012. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jin, H.; Zhang, L.; Hu, W.; Li, Y.; Xu, N. Screening and isolation of the algicidal compounds from marine green alga Ulva intestinalis. Chin. J. Oceanol. Limnol. 2016, 34, 781–788. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Meng, K.; Su, Z.; Guo, G.L.; Pu, Y.F.; Wang, C. Isolation and purification of antialgal compounds from the red alga Gracilaria lemaneiformis for activity against common harmful red tide microalgae. Environ. Sci. Pollut. Res. 2017, 24, 4964–4972. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Mathaiyan, M. Emerging novel anti HIV biomolecules from marine Algae: An overview. J. Appl. Pharm. Sci. 2015, 5, 153–158. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar] [CrossRef] [PubMed]
- Siriwardhana, N.; Jeon, Y.-J.; Kim, S.-H.; Ha, J.-H.; Heo, S.-J.; Lee, K.-W. Enzymatic Hydrolysis for Effective Extraction of Antioxidative Compounds from Hizikia fusiformis. Algae 2004, 19, 59–68. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Rasoul-Amini, S.; Morowvat, M.H. Algae for the Production of SCP; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 163–184. [Google Scholar]
- Laouina, A. Prospective “Maroc 2030”: Gestion Durable des Ressources Naturelles de la Biodiversité au Maroc; Haut-Commissariat au Plan (HCP): Casablanca, Marocco, 2006. [Google Scholar]
- Chalabi, A.; Semroud, R.; Grimes, S. Plan d’Action Stratégique pour la Conservation de la Diversité Biologique en Région Méditerranéenne; National PAS-BIO Algérie, Ministère de l’Aménagement du Territoire et de l’Environnement: Algiers, Algérie, 2015. [Google Scholar]
- Sasidharan, S.; Aravindran, S.; Latha, L.Y.; Vijenthi, R.; Saravanan, D.; Amutha, S. In vitro antioxidant activity and hepatoprotective effects of lentinula edodes against paracetamol-induced hepatotoxicity. Molecules 2010, 15, 4478–4489. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.L.; Mular, L.; Drenkard, E.J.; Shearer, T.L.; Fredericq, S.; Fairchild, C.R.; Prudhomme, J.; Roch, K.; Hay, M.E.; Aalbersberg, W.; et al. Ecological leads for natural product discovery: Novel sesquiterpene hydroquinones from the red macroalga Peyssonnelia sp. Tetrahedron 2011, 66, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Suzuki, M.; Ishii, T.; Okino, T.; Abe, T.; Masuda, M. Antibacterial activity of halogenated sesquiterpenes from Malaysian Laurencia spp. Phytochemistry 2008, 69, 2490–2494. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, H.; Craige, S.J.; Mcinnes, G. The occurrence of 2,3-dibromobenzyl alcohol 4,5-disulfate, dipotassium salt, in Polysiphonia lanosa. Can. J. Chem. 1966, 44, 74–78. [Google Scholar] [CrossRef]
- Fenical, W.; Sims, J.J.; Wing, R.M.; Radlick, P.C. Zonarene, a sesquiterpene from the brown seaweed Dictyopteris zonarioides. Phytochemistry 1972, 11, 1161–1163. [Google Scholar] [CrossRef]
- Kamei, Y.; Sueyoshi, M.; Hayashi, K.; Terada, R.; Nozaki, H. The novel anti-Propionibacterium acnes compound, Sargafuran, found in the marine brown alga Sargassum macrocarpum. J. Antibiot. 2009, 62, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Ji, N.Y.; Li, X.M.; Li, K.; Ding, L.P.; Gloer, J.B.; Wang, B.G. Diterpenes, sesquiterpenes, and a C15-acetogenin from the marine red alga Laurencia mariannensis. J. Nat. Prod. 2007, 70, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- Rosaline, X.D.; Sakthivelkumar, S.; Rajendran, K.; Janarthanan, S. Screening of selected marine algae from the coastal Tamil Nadu, South India for antibacterial activity. Asian Pac. J. Trop. Biomed. 2012, 2. [Google Scholar] [CrossRef]
- Mishra, J.K.; Srinivas, T.; Sawhney, S. Antibacterial Activity of Seaweed Halimeda Opuntia From the coasts of south andaman. Glob. J. Biosci. Biotechnol. 2016, 5, 345–348. [Google Scholar]
- Taskin, E.; Ozturk, M.; Kurt, O. Antibacterial activities of some marine algae from the Aegean Sea (Turkey). Afr. J. Biotechnol. 2007, 6, 2746–2751. [Google Scholar] [CrossRef]
- Cortés, Y.; Hormazábal, E.; Leal, H.; Urzúa, A.; Mutis, A.; Parra, L.; Quiroz, A. Novel antimicrobial activity of a dichloromethane extract obtained from red seaweed Ceramium rubrum (Hudson) (Rhodophyta: Florideophyceae) against Yersinia ruckeri and Saprolegnia parasitica, agents that cause diseases in salmonids. Electron. J. Biotechnol. 2014, 17, 126–131. [Google Scholar] [CrossRef]
- Salvador, N.; Garreta, A.G.; Lavelli, L.; Ribera, M.A. Antimicrobial activity of Iberian macroalgae. Sci. Mar. 2007, 71, 101–113. [Google Scholar] [CrossRef]
- Rangaiah, G.S.; Lakshmi, P.; Sruthikeerthi, K. Antimicrobial activity of the crude extracts of Chlorophycean seaweeds Ulva, Caulerpa and Spongomorpha sps. against clinical and phytopathogens. Drug Invent. Today 2010, 2, 311–314. [Google Scholar] [CrossRef]
- Seenivasan, R.; Indu, H.; Archana, G.; Geetha, S. The Antibacterial Activity of Some Marine Algae from South East Coast of India. Environ. Sci. 2010, 9, 480–489. [Google Scholar]
- Lavanya, R.; Veerappan, N.; Nadu, P.-T. Antibacterial Potential of Six Seaweeds Collected from Gulf of Mannar of Southeast Coast of India. Adv. Biol. Res. 2011, 5, 38–44. [Google Scholar]
- Pandian, P.; Selvamuthukumar, S.; Manavalan, R. Screening of antibacterial and antifungal activities of red marine algae Acanthaphora spicifera (Rhodophyceae). J. Biomed. Sci. Res. 2011, 3, 444–448. [Google Scholar]
- Kausalya, M.; Rao, G.M.N. Antimicrobial activity of marine Algae. J. Algal Biomass Util. 2015, 6, 78–87. [Google Scholar]
- Shanmughapriya, S.; Manilal, A.; Sujith, S.; Selvin, J.; Kiran, G.S.; Natarajaseenivasan, K. Antimicrobial activity of seaweeds extracts against multiresistant pathogens. Ann. Microbiol. 2008, 58, 535–541. [Google Scholar] [CrossRef]
- Kolsi, R.B.A.; Frikha, D.; Jribi, I.; Hamza, A.; Feki, L.; Belghith, K. Screening of antibacterial and antifongical activity in marine macroalgae and magnoliophytea from the coast of Tunisia. Int. J. Pharm. Pharm. Sci. 2015, 7, 47–51. [Google Scholar]
- Baleta, F.N.; Bolaños, J.M.; Ruma, O.C.; Baleta, A.N.; Cairel, J.D. Phytochemicals screening and antimicrobial properties of Sargassum oligocystum and Sargassum crassifolium Extracts. J. Med. Plants 2017, 5, 382–387. [Google Scholar]
- Radhika, D.; Priya, R. Antifungal Activity of Acanthophora Spicifera, Padina Tetrastomatica and Caulerpa Scalpelliformis Against Some Fungal Pathogens in Crude and Fractionated. World J. Pharm. Pharm. Sci. 2016, 5, 1155–1162. [Google Scholar]
- Tüney, I.; Çadirci, B.H.; Ünal, D.; Sukatar, A. Antimicrobial Activities of the Extracts of Marine Algae from the Coast of Urla (Izmir, Turkey). Turk. J. Biol. 2006, 30, 171–175. [Google Scholar]
- Moorthi, P.V.; Balasubramanian, C. Antimicrobial properties of marine seaweed, Sargassum muticum against human pathogens. J. Coast. Life Med. 2015, 3, 1–5. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Reguant, C.; Bordons, A.; Arola, L.; Rozès, N. Influence of phenolic compounds on the physiology of Oenococcus oeni from wine. J. Appl. Microbiol. 2000, 88, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Alberto, M.R.; Farías, M.E.; Manca De Nadra, M.G. Effect of gallic acid and catechin on Lactobacillus hilgardii 5w growth and metabolism of organic compounds. J. Agric. Food Chem. 2001, 49, 4359–4363. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liu, Q.; Xu, C.; Yu, J.; Zhao, L.; Guo, Q. Damage to the Membrane Permeability and Cell Death of Vibrio parahaemolyticus Caused by Phlorotannins with Low Molecular Weight from Sargassum thunbergii. J. Aquat. Food Prod. Technol. 2016, 25, 323–333. [Google Scholar] [CrossRef]
- Nagayama, K.; Shibata, T.; Fujimoto, K.; Honjo, T.; Nakamura, T. Algicidal effect of phlorotannins from the brown alga Ecklonia kurome on red tide microalgae. Aquaculture 2003, 218, 601–611. [Google Scholar] [CrossRef]
- Ameer, J.; Begum, M.; Selvaraju, P.; Vijayakumar, A. Evaluation of antifungal activity of seaweed extract (Turbinaria conoides) against Fusarium oxysporum. Chem. Sci. Rev. Lett. 2016, 9411, 2014–2016. [Google Scholar]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological potential of marine macroalgae of the genus Cystoseira. Acta Biol. Hung. 2015, 66, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Xavier, D.R. Antibacterial activity of the seaweeds Chaetomorpha linum and Padina gymnospora on human bacterial pathogens. J. Environ. Biotechnol. Res. 2017, 6, 43–52. [Google Scholar]
- Moubayed, N.M.S.; Al Houri, H.J.; Al Khulaifi, M.M.; Al Farraj, D.A. Antimicrobial, antioxidant properties and chemical composition of seaweeds collected from Saudi Arabia (Red Sea and Arabian Gulf). Saudi J. Biol. Sci. 2017, 24, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Bhagavathy, S.; Sumathi, P.; Jancy Sherene Bell, I. Green algae Chlorococcum humicola—A new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop. Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Al-Saif, S.S.A.; Abdel-Raouf, N.; El-Wazanani, H.A.; Aref, I.A. Antibacterial substances from marine algae isolated from Jeddah coast of Red sea, Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Renata, C.C.; Paula, F.F.; Isabel, R.A.; Ana, P.S.; Vera, L.; Maria, T.; Alexandre, G. Antimicrobial activity of seaweeds of Pernambuco, northeastern coast of Brazil. Afr. J. Microbiol. Res. 2016, 10, 312–318. [Google Scholar] [CrossRef]
- El Shafay, S.M.; Ali, S.S.; El-Sheekh, M.M. Antimicrobial activity of some seaweeds species from Red sea, against multidrug resistant bacteria. Egypt. J. Aquat. Res. 2016, 42, 65–74. [Google Scholar] [CrossRef]
- Khelil-Radji, F.; Belhouari, M.Y.; Chemlal-Kherraz, D.; Matallah-Boutiba, A.; Boutiba, Z. Antimicrobial activity of aqueous and ethanol extracts of two marine algae collected from Algerian west coast. Electron. J. Environ. Agric. Food Chem. 2017, 6655, 100–104. [Google Scholar]
- Genovese, G.; Leitner, S.; Minicante, S.A.; Lass-Flörl, C. The Mediterranean red alga Asparagopsis taxiformis has antifungal activity against Aspergillus species. Mycoses 2013, 56, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Eom, S.; Lee, E.; Jung, Y.; Kim, H.; Jo, M.; Son, K.; Lee, H.; Kim, J.H.; Lee, M.; et al. In vitro antibacterial and synergistic effect of phlorotannins isolated from edible brown seaweed Eisenia bicyclis against acne-related bacteria. Algae 2014, 29, 47–55. [Google Scholar] [CrossRef]
- Nithya, P.; Dhanalakshmi, B. Antibacterial activity of methanol extracts from selected seaweed of south east coast of India. Int. J. Adv. Res. 2016, 2, 714–718. [Google Scholar]
- Sahnouni, F.; Benattouche, Z.; Matallah-Boutiba, A.; Benchohra, M.; Moumen Chentouf, W.; Bouhadi, D.; Boutiba, Z. Antimicrobial activity of two marine algae Ulva rigida and Ulva intestinalis collected from Arzew gulf (Western Algeria). J. Appl. Environ. Biol. Sci. 2016, 6, 242–248. [Google Scholar]
- Shobier, A.H.; Abdel Ghani, S.A.; Barakat, K.M. GC/MS spectroscopic approach and antifungal potential of bioactive extracts produced by marine macroalgae. Egypt. J. Aquat. Res. 2016, 42, 289–299. [Google Scholar] [CrossRef]
- Al-zahrani, A.; Omer, H.; Al-judaibi, A. Impact of Antibacterial Activity of Physical Storage Extracts on Pathogenic Bacteria. J. Biosci. Med. 2016, 4, 54–62. [Google Scholar] [CrossRef]
- Daisy, A.; Indra, V.; Geetha, S.; Seetharaman, S.; Selvamuthu, B.; Nadu, T. Phytochemical Profiling and Antibacterial Activity of Seaweeds Collected From Pulicate Lake, Coramandal Coast of South India. World J. Pharm. Pharm. Sci. 2016, 5, 740–755. [Google Scholar] [CrossRef]
- Alagan, V.; Valsala, R.; Rajesh, K. Bioactive Chemical Constituent Analysis, in vitro Antioxidant and Antimicrobial Activity of Whole Plant Methanol Extracts of Ulva lactuca Linn. Br. J. Pharm. Res. 2017, 15, 1–14. [Google Scholar] [CrossRef]
- Rani, V.; Jawahar, P.; Shakila, R.J.; Srinivasan, A. Antibacterial Activity of Some Brown Seaweeds of Gulf Of Mannar, South East Coast of India. J. Pharm. Biosci. 2017, 4, 14–21. [Google Scholar]
- Alghazeer, R.; Whida, F.; Abduelrhman, E.; Gammoudi, F.; Azwai, S. Screening of antibacterial activity in marine green, red and brown macroalgae from the western coast of Libya. Nat. Sci. 2013, 5, 7–14. [Google Scholar] [CrossRef]
- Sebaaly, C.; Kassem, S.; Grishina, E.; Kanaan, H.; Sweidan, A.; Chmit, M.S.; Kanaan, H.M. Anticoagulant and antibacterial activities of polysaccharides of red algae Corallina collected from lebanese coast. J. Appl. Pharm. Sci. 2014, 4, 30–37. [Google Scholar] [CrossRef]
- Rameshkumar, S.; Kolidoss, R.; Arasan, S. Antibacterial activity of four Gracilaria species of red seaweeds collected from Mandapam Coast, Gulf of Mannar Marine Biosphere Reserve, India. J. Coast. Life Med. 2016, 4, 703–707. [Google Scholar] [CrossRef]
- Alamsjah, M.A.; Hirao, S.; Ishibashi, F.; Fujita, Y. Isolation and structure determination of algicidal compounds from Ulva fasciata. Biosci. Biotechnol. Biochem. 2005, 69, 2186–2192. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Feng, L.; Tang, X.; Wang, J.; Dong, S. Allelopathic growth inhibition of Heterosigma akashiwo by the three Ulva spcieces (Ulva Pertusa, Ulva Linza, Enteromorpha intestinalis) under laboratory conditions. Acta Oceanol. Sin. 2012, 31, 138–144. [Google Scholar] [CrossRef]
- Manilal, A.; Sujith, S.; Seghal Kiran, G.; Selvin, J.; Panikkar, M.V.N. Evaluation of seaweed bioactives on common aquatic floral and faunal weeds of shrimp ponds. Thalassas 2011, 27, 47–56. [Google Scholar]
- Chowdhury, M.T.H.; Bangoura, I.; Kang, J.-Y.; Cho, J.Y.; Joo, J.; Choi, Y.S.; Hwang, D.S.; Hong, Y.-K. Comparison of Ecklonia cava, Ecklonia stolonifera and Eisenia bicyclis for phlorotannin extraction. J. Environ. Biol. 2014, 35, 713. [Google Scholar] [PubMed]
- Nan, C.; Zhang, H.; Zhao, G. Allelopathic interactions between the macroalga Ulva pertusa and eight microalgal species. J. Sea Res. 2004, 52, 259–268. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B.; Wang, C. Isolation, purification, and identification of antialgal substances in green alga Ulva prolifera for antialgal activity against the common harmful red tide microalgae. Environ. Sci. Pollut. Res. 2016, 23, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kwon, E.H.; Choi, J.S.; Hong, S.Y.; Shin, H.W.; Hong, Y.K. Antifouling activity of seaweed extracts on the green alga Enteromorpha prolifera and the mussel Mytilus edulis. J. Appl. Phycol. 2001, 13, 117–125. [Google Scholar] [CrossRef]
- Sun, X.; Xia, Y.; Xu, N.; Cai, X. Inhibitory effects of marine alga Enteromorpha intestinalis on the growth of the red tide microalga Prorocentrum micans. In Proceedings of the World Automation Congress, Puerto Vallarta, Mexico, 24–28 June 2012. [Google Scholar]
- Wang, Y.; Zhou, B.; Tang, X. Effects of two species of macroalgae—Ulva pertusa and Gracilaria lemaneiformis—On growth of Heterosigma akashiwo (Raphidophyceae). J. Appl. Phycol. 2009, 21, 375–385. [Google Scholar] [CrossRef]
- Ye, C.; Zhang, M. Allelopathic Effect of Macroalga Gracilaria Tenuistipitata (Rhodophyta) on the Photosynthetic Apparatus of Red-tide Causing Microalga Prorocentrum micans. IERI Procedia 2013, 5, 209–215. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Gobler, C.J. The green macroalga, Ulva lactuca, inhibits the growth of seven common harmful algal bloom species via allelopathy. Harmful Algae 2011, 10, 480–488. [Google Scholar] [CrossRef]
- Wang, R.; Xiao, H.; Wang, Y.; Zhou, W.; Tang, X. Effects of three macroalgae, Ulva linza (Chlorophyta), Corallina pilulifera (Rhodophyta) and Sargassum thunbergii (Phaeophyta) on the growth of the red tide microalga Prorocentrum donghaiense under laboratory conditions. J. Sea Res. 2007, 58, 189–197. [Google Scholar] [CrossRef]
- Manilal, A.; Sujith, S.; Sabarathnam, B.; Kiran, G.S.; Selvin, J.; Shakir, C.; Lipton, A.P. Bioactivity of the red algae Asparagopsis taxiformis collected from the southwestern coast of India. Braz. J. Oceanogr. 2010, 58, 93–100. [Google Scholar] [CrossRef]
- Pramitha, V.S.; Lipton, A.P. Growth responses of microalgae, chlorella salina and isochrysis galbana exposed to extracts of the macroalga, Hypnea musciformis. Indian J. Fish. 2011, 58, 95–99. [Google Scholar]
- Ying, S.; Hui, W.; Gan, G.; Yin, P.; Bin, Y.; Chang, W. Green Alga Ulva pertusa—A new source of bioactive compounds with antialgal activity. Environ. Sci. Pollut. Res. 2015, 22, 10351–10359. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.Y.; Lee, S.B.; Jin, D.H.; Hong, Y.K.; Jin, H.J. Isolation of algicidal compounds from the red Alga Corallina pilulifera against red tide microalgae. J. Appl. Phycol. 2010, 22, 453–458. [Google Scholar] [CrossRef]
- Hamouda, M.H.H.; Karim-Eldeen, N.E. Potentiality and Biological Activities of the Polysaccharide. J. Agric. Food Chem. 2015, 6, 373–392. [Google Scholar]
- Wang, R.; Wang, Y.; Zhou, J.; Sun, J.; Tang, X. Algicidal activity of Ulva pertusa and Ulva prolifera on Prorocentrum donghaiense under laboratory conditions. Afr. J. Microbiol. Res. 2013, 7, 4389–4396. [Google Scholar] [CrossRef]
- Kakisawa, H.; Asari, F.; Kusumi, T.; Toma, T.; Sakurai, T.; Oohusa, T.; Hara, Y.; Chiharai, M. An allelopathic fatty acid from the brown alga Cladosiphon okamuranus. Phytochemistry 1988, 27, 731–735. [Google Scholar] [CrossRef]
- Chang, V.S.; Teo, S.S. Evaluation of heavy metal, antioxidant and anti-tyrosinase activities of red seaweed (Eucheuma cottonii). Int. Food Res. J. 2016, 23, 2370–2373. [Google Scholar]
- Shalaby, E.A.; Shanab, S.M.M.; El-Fayoumy, E.A. Enteromorpha compressa exhibits potent antioxidant activity. J. Biomed. Biotechnol. 2011, 2011, 726405. [Google Scholar] [CrossRef]
- Mole Megha, N.; Sabale Anjali, B. Antioxidant Potential of Seaweeds from Kunakeshwar along the West Coast Maharashtra. Asian J. Biomed. Pharm. Sci. 2013, 3, 45–50. [Google Scholar]
- Matsukawa, R.; Dubinsky, Z.; Kishimoto, E.; Masaki, K.; Masuda, Y.; Takeuchi, T.; Chihara, M.; Yamamoto, Y.; Niki, E.; Karube, I. A comparison of screening methods for antioxidant activity in seaweeds. J. Appl. Phycol. 1997, 9, 29. [Google Scholar] [CrossRef]
- Lim, S.N.; Cheung, P.C.K.; Ooi, V.E.C.; Ang, P.O. Evaluation of antioxidative activity of extracts from a brown seaweed, Sargassum siliquastrum. J. Agric. Food Chem. 2002, 50, 3862–3866. [Google Scholar] [CrossRef] [PubMed]
- Zubia, M.; Robledo, D.; Freile-Pelegrin, Y. Antioxidant activities in tropical marine macroalgae from the Yucatan Peninsula, Mexico. J. Appl. Phycol. 2007, 19, 449–458. [Google Scholar] [CrossRef]
- Lee, S.; Lee, Y.S.; Jung, S.H.; Kang, S.S.; Shin, K.H. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res. 2003, 26, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Farasat, M.; Khavari-Nejad, F.A.; Nabavi, S.M.B.; Namjooyan, F. Antioxidant Properties of two Edible Green Seaweeds from northern coasts of the Persian Gulf. Jundishapur J. Nat. Pharm. Prod. 2013, 8, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.T.; Kwek, M.T.; Ismail, N.I.M.; Ooi, J.L.S.; Amri, A.Y.; Manan, F.A.; Law, Y.C.; Wong, F.C. Antioxidant Activities of Methanol Extract and Solvent Fractions of Marine Macroalga, Avrainvillea Erecta (Berkeley) a. Gepp and E.S. Gepp (Dichotomosiphonaceae). Trop. J. Pharm. Res. 2015, 14, 503–509. [Google Scholar] [CrossRef]
- Alghazeer, R.; Elmansori, A.; Sidati, M.; Gammoudi, F.; Azwai, S. In Vitro Antibacterial Activity of Flavonoid Extracts of Two Selected Libyan Algae against Multi-Drug Resistant Bacteria Isolated from Food Products. J. Biosci. Med. 2017, 5, 27–48. [Google Scholar] [CrossRef]
- Pinteus, S.; Silva, J.; Alves, C.; Horta, A.; Fino, N.; Rodrigues, A.I.; Mendes, S.; Pedrosa, R. Cytoprotective effect of seaweeds with high antioxidant activity from the Peniche coast (Portugal). Food Chem. 2017, 218, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Mamelona, J.; Pelletier, É.; Girard-Lalancette, K.; Legault, J.; Karboune, S.; Kermasha, S. Quantification of phenolic contents and antioxidant capacity of Atlantic sea cucumber, Cucumaria frondosa. Food Chem. 2007, 104, 1040–1047. [Google Scholar] [CrossRef]
- Cho, M.; Lee, H.S.; Kang, I.J.; Won, M.H.; You, S. Antioxidant properties of extract and fractions from Enteromorpha prolifera, a type of green seaweed. Food Chem. 2011, 127, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Raja, R.; Hemaiswarya, S.; Arunkumar, K.; Carvalho, I.S. Antioxidant activity and lipid profile of three seaweeds of Faro, Portugal. Braz. J. Bot. 2016, 39, 9–17. [Google Scholar] [CrossRef]
- Nahas, R.; Abatis, D.; Anagnostopoulou, M.A.; Kefalas, P.; Vagias, C.; Roussis, V. Radical-scavenging activity of Aegean Sea marine algae. Food Chem. 2007, 102, 577–581. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Source | Reference |
|---|---|---|
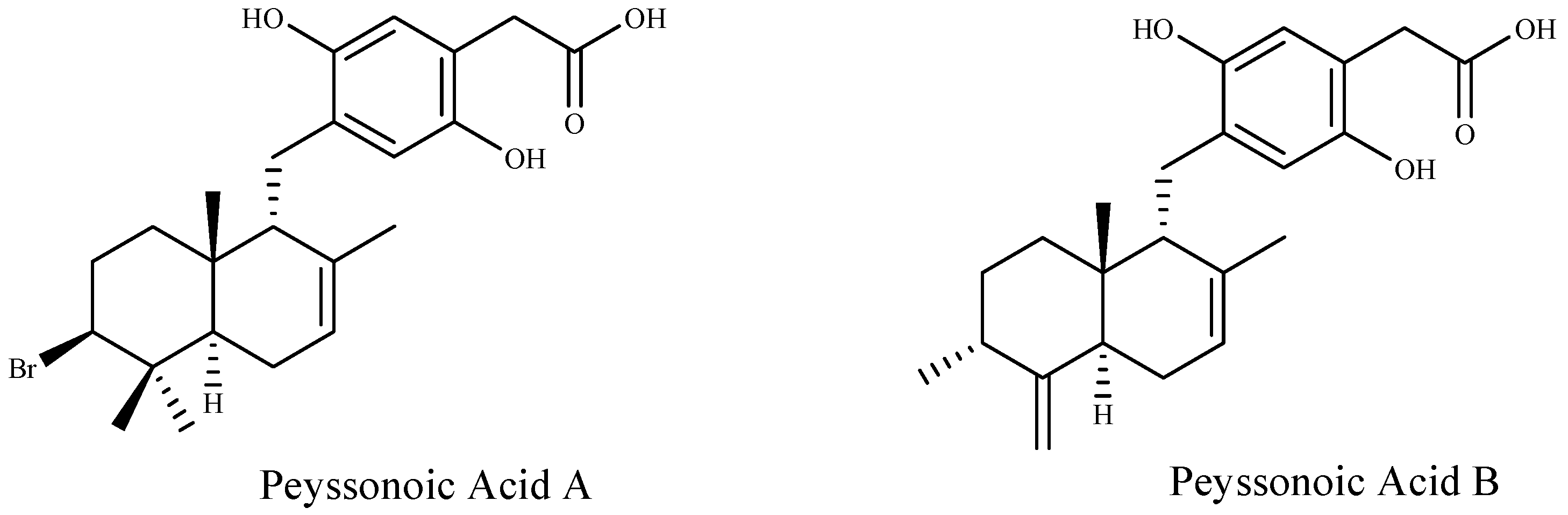
| Peyssonoic acid A and B | Peyssonnelia sp. | [92] |
| Tiomanene Acetylmajapolene (A and B) | Laurencia sp. | [93] |
| 3-Dibromobenzaldehyde-4,5-disulfate potassium salt 5-Bromo-3,4-dihydroxybenzaldehyde | Polysiphonia lanora | [94] |
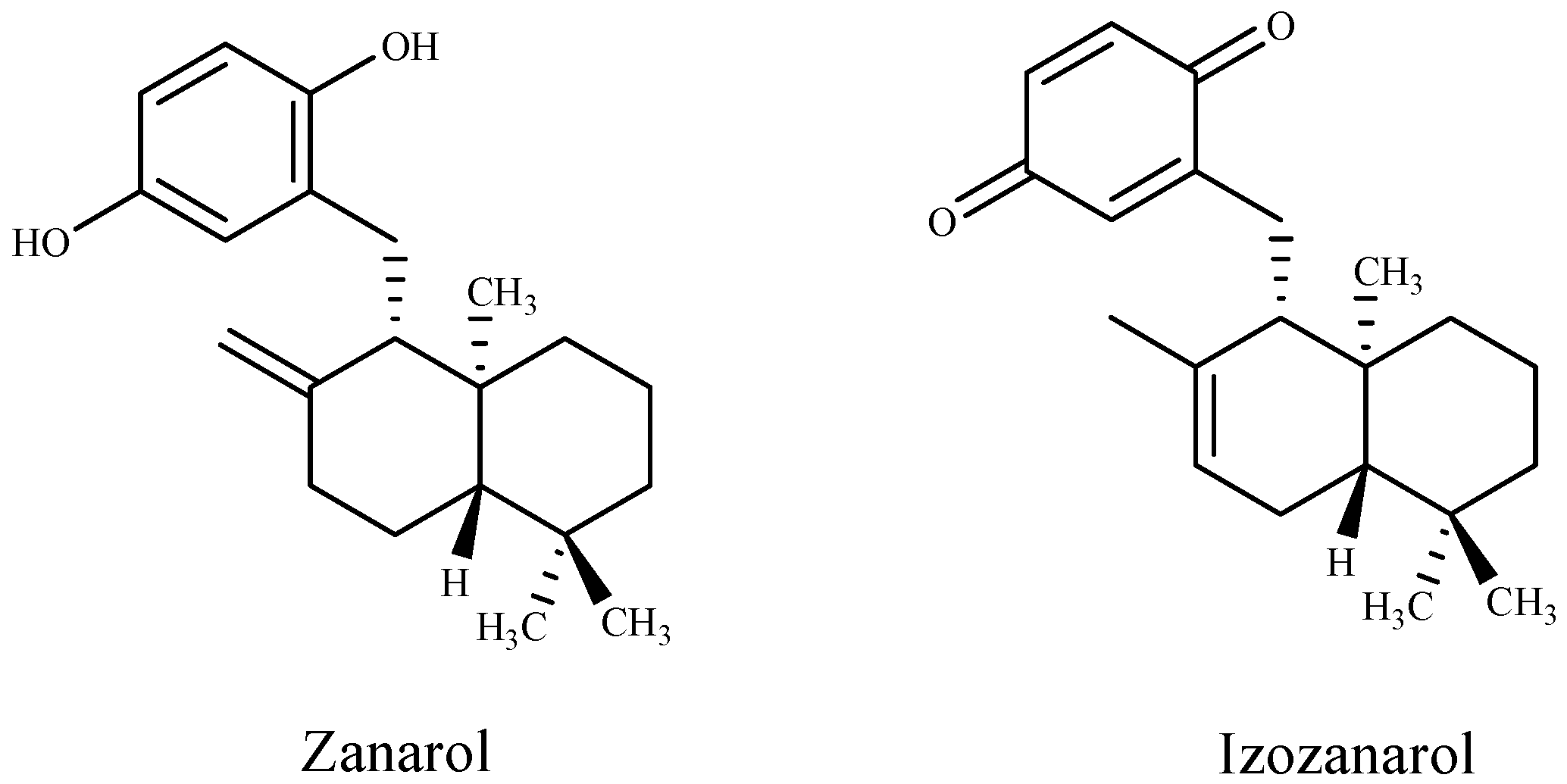
| Zonarol and isozonarol sesquiterpenes | Dictyopteris zonarioides | [95] |
| Diterpene sargafuran | Sargassum macrocarpum | [96] |
| 10-Hydroxy kahukuene B | Laurencia mariannensis | [97] |
| Solvents | Seaweed | Target Organisms | Reference |
|---|---|---|---|
| Acetone | Turbinaria conoides | Fusarium oxysporum | [121] |
| Ulva lactuca | Escherichia coli, Staphylococcus aureus, Bacillus mycoides, Bacillus subtilis, Klebsiella pneumoniae, Aspergillus flavus, Aspergillus fumigatus, penicillium purpurescens, Candida albicans and Penicillium verrucosum | [122] | |
| Acetone, Chloroform | Sargassum muticum | Micrococcus sp., Staphylococcus aureus (methicillinresistance), Salmonella paratyphi B, Staphylococcus epidermis, Enterobacter aerogenus, Shigella fleschneri, Proteus vulgaris, Staphylococcus aureus Salmonella typhymurium | [113] |
| Acetone, Ethyl, acetate, Hexane | Sargassum wightii, Chaetomorpha linum and Padina gymnospora | Erwinia amylovora, Enterobacter aerogenes, Proteus vulgaris, Escherichia coli, Staphylococcus aureus, Bacillus subtilis Enterococcus faecalis | [98] |
| Acetone, Ethyl acetate, Hexane, Methanol | Chaetomorpha linum | Pseudomonas aeruginosa Bacillus subtilis | [123] |
| Acetone, Methanol | Sargassum platycarpum, Sargassum latifolium | Escherichia coli, Salmonella sp., Staphylococcus xylosus, Staphylococcus aureus, Bacillus subtilis, Enterococcus faecalis Candida albicans | [124] |
| Benzene, Diethyl ether, Ethyl acetate, Hexane | Chlorococcum humicola | Escherichia coli, Pseudomonas aeruginosa, Salmonella typhimurium, Klebsiella pneumoniae, Vibreo cholerae, Staphylococcus aureus, Bacillus subtilis, Candida albicans, Aspergillus niger and Aspergillus flavus | [125] |
| Chloroform, Ethanol | Ulva reticulata, Caulerpa occidentalis, Cladophora socialis, Dictyota ciliolata, and Gracilaria dendroides | Escherichia coli, Pseudomonas aeruginosa, Stapylococcus aureus Enterococcus faecalis | [126] |
| Chloroform, Hexane, Ethyl acetate, Methanol | Ulva lactuca, Sargassum polyceratium, Caulerpa racemosa | Bacillus subtilis, Micrococcus luteus, Staphylococcus aureus, bacteria Escherichia coli and Klebsiella pneumoniae | [127] |
| Chloroform, Hexane, Ethyl acetate, Methanol | Jania adhaerens, Padina gymnospora | Bacillus subtilis and Micrococcus luteus | [127] |
| Diethyl ether, Methanol, Ethanol | Ceramium rubrum, Sargassum vulgare, Sargassum fusiforme and Padina pavonia | Pseudomonas aeruginosa, Shigellaflexneri, and Klebsiella pneumoniae | [128] |
| Ethanol | Stypocaulon scoparium and Halopitys Incurvus | Staphylococcus aureus, Bacillus subtilis, Escherichia coli, pseudomonas aeruginosa, Fusarium oxysporum f. sp. Albedinis and Penicillium sp. | [129] |
| Asparagopsis taxiformis | Aspergillus fumigatus, Aspergillus terreus and Aspergillus flavus | [130] | |
| Ethyl acetate | Eisenia bicyclis | Propionibacter iumacnes, Staphylococcus aureus, Staphylococcus epidermidis and Pseudomonas aeruginosa | [131] |
| Methanol | Ulva lactuca, Sargassum wightii and Gracilaria edulis | Bacillus cereus, Streptococcus faecali, Staphylococcus aureus, E-coli, Pseudomonas aeruginosa, Salmonella typhi and Vibreo cholerae | [132] |
| Ulva rigida and Ulva intestinalis | Esherichia coli, Streptococcus pyogenes, Staphylococcus epidermidis, Candida albicans and Aspergillus niger | [133] | |
| Ulva lactuca, Ulva fasciata Enteromorpha compressa, Pterocladia capillacea, Hypnea musciformis and Padinapavonica | Fusarium solani, Fusarium oxysporum, Tricodermahamatum, Aspergillus flavipes and Candida albicans | [134] | |
| Ulva lactuca | Staphylococcus aureus and Pseudomonas aeruginosa | [135] | |
| Sargassum wightii | Staphylococcus aureus., Klebsiella pneumonia, Proteus mirabilis, Escherichia coli and Proteus valgaris | [136] | |
| Ulva lactuca | Bacillus subtilis, Corynebacterium diphtheria, Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa, Salmonella paratyphi, Aspergillus niger Aspergillus fumigatus | [137] | |
| Turbinaria ornata, Padina tetrastromatica | Micrococcus luteus and Bacillus subtilis | [138] | |
| Methanol, Water | Cytoseira crinite | Taphylococcus aureus, Bacillus subtilis, Bacillus spp., Staphylococcus epidermidis Escherichia coli and Salmonella typhi, Klebsiella spp., and Pseudomonas aeruginosa | [139] |
| Polysaccharides | Corallina | Staphylococcus epidermidis, Staphylococcus aureus, Enterococcus feacalis Escherichia coli and Pseudomonas aeruginosa | [140] |
| Toluene | Gracilaria crassa, Gracilaria debilis, and Gracilaria corticata | Escherichia coli, Shigella sp., Staphylococcus aureus, Vibriocholerae, Proteus sp., Bacillus subtilis and Pseudomonas fluroscens | [141] |
| Macroalgae | Fraction Used or Solvent | Target Species | Effects | Reference |
|---|---|---|---|---|
| Enteromorpha intestinalis | Ethanol extract Fresh tissue | Prorocentrum micans | Fresh tissue and ethanol extracts significantly inhibited the growth of P. micans. | [149] |
| Sargassum thunbergii | Methanol extract | Heterosigma akashiwo Skeletonema costatum Prorocentrum micans | Stronger inhibitory effects on the growths of red tide microalgae tests. | [20] |
| Enteromorpha clathrata, Undaria pinnatifida Laminaria japonica Porphyra tenera Ulva pertusa | Seawater extracts | Skeletonema costatum | The macroalgal extracts of P. tenera, E. clathrata, and U. pertusa showed strong growth inhibition on S. costatum. | [19] |
| Gracilaria lemaneiformis Ulva pertusa | Fresh thalli Water-soluble extract Dry powder | Heterosigma akashiwo | Algicidal effects of both macroalgae on H. akashiwo, cells were entirely killed. | [150] |
| Gracilaria tenuistipitata | Dry powder | Prorocentrum micans | Inhibitory effect on the photosynthesis of P. micans. | [151] |
| Ulva lactuca | Dry powder Fresh thalli Extracts | Aureococcus anophagefferens Cochlodinium polykrikoides Pseudo-Nitzschiamultiseries Prorocentrum minimum Karlodinium veneficum Chattonella marina Karenia brevis | The fresh thalli and dry powder strongly inhibited the growth of all seven HAB species with advantage of dry powder. The extracts of U. lactuca exhibited dramatic allelopathic effect on the HAB species. | [152] |
| Sargassum thunbergii Corallina pilulifera Ulva pertusa | Aqueous extracts Fresh tissue Dry powder | Heterosigma akashiwo Alexandrium tamarense | The growth of the two microalgae was strongly inhibited | [153] |
| Asparagopsis taxiformis | Methanol extract | Trichodesmium sp. | Total inhibition of Trichodesmium sp. growth | [154] |
| Hypnea musciformis | Methanol extract | Isochrysis galbana Chlorella salina | Enhancement growing of both microalgae even at low concentration | [155] |
| Gracilaria lemaneiformis | Ethanol extract | Prorocentrum donghaiense Skeletonem acostatum Heterosigma akashiwo Amphidinium carterae Phaeocystis globa Karenia mikimitoi | Inhibitory effect on the growth of all microalgae | [84] |
| Ulva pertusa | Methanol extract | Red tide microalgae Feed microalgae | The methanolic extract showed antialgal activity against red tide, with no growth inhibition for feed microalgae. | [156] |
| Corallina pilulifera | Methanol extract | Red tide microalgae Feed microalgae | Stronger inhibitory effects on the growths of red tide microalgae, with no growth inhibition for feed microalgae. | [157] |
| Ulva intestinalis | Ethanol extract Fresh tissue Dry powder | Heterosigma akashiwo Prorocentrum micans | The fresh tissue, dry powder and extract, all exhibited obvious algicidal effects on red tide microalgae. | [83] |
| 37 species | Methanol extract Water extract | Heterosigma akashiwo | The green alga Ulva fasciata showed the strongest algicidal activity among the 37 seaweeds tested | [142] |
| Ulva lactuca Ulvafasciata | Ethanol extract | Chlorella vulgaris | Stimulation of growth and progressive increase of Chlorella vulgaris biomass | [158] |
| Ulva pertusa Ulva prolifera | Extracts (acetone, ether chloroform, methanol) Fresh tissue Dry powder | Prorocentrum donghaiense | Stronger inhibitory growing effects by fresh tissue and dry powder of both seaweeds. Methanol extracts of the macroalgae were found to strongly inhibit the growth of P. donghaiense. | [159] |
| Ecklonia kurome | Phlorotannins extract | Cochlodinium polykrikoides Chattonella antiqua Kareniam ikimotoi | Destruction of 99% cells of ride tide microalgae, with no mortality observed among other organisms such us: Pagrus major, tiger puffer Fugu rubripes or larval blue crab Portunustrituberculatus. | [120] |
| Ulva pertusa | Fresh tissue | Heterosigma akashiwo, Skeletonema costatum, Tetraselmis subcordiformis, Nitzschia closterium, Chaetoceros gracile, Chroomonas placoidea, Isochrysis galbana, Alexandrium tamarense | Algicidal interaction between green alga Ulva pertusa and all phytoplankton species | [146] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zerrifi, S.E.A.; El Khalloufi, F.; Oudra, B.; Vasconcelos, V. Seaweed Bioactive Compounds against Pathogens and Microalgae: Potential Uses on Pharmacology and Harmful Algae Bloom Control. Mar. Drugs 2018, 16, 55. https://doi.org/10.3390/md16020055
Zerrifi SEA, El Khalloufi F, Oudra B, Vasconcelos V. Seaweed Bioactive Compounds against Pathogens and Microalgae: Potential Uses on Pharmacology and Harmful Algae Bloom Control. Marine Drugs. 2018; 16(2):55. https://doi.org/10.3390/md16020055
Chicago/Turabian StyleZerrifi, Soukaina El Amrani, Fatima El Khalloufi, Brahim Oudra, and Vitor Vasconcelos. 2018. "Seaweed Bioactive Compounds against Pathogens and Microalgae: Potential Uses on Pharmacology and Harmful Algae Bloom Control" Marine Drugs 16, no. 2: 55. https://doi.org/10.3390/md16020055
APA StyleZerrifi, S. E. A., El Khalloufi, F., Oudra, B., & Vasconcelos, V. (2018). Seaweed Bioactive Compounds against Pathogens and Microalgae: Potential Uses on Pharmacology and Harmful Algae Bloom Control. Marine Drugs, 16(2), 55. https://doi.org/10.3390/md16020055