Two Novel Multi-Functional Peptides from Meat and Visceral Mass of Marine Snail Neptunea arthritica cumingii and Their Activities In Vitro and In Vivo

Abstract
:1. Introduction
2. Results
2.1. Proximate Analysis
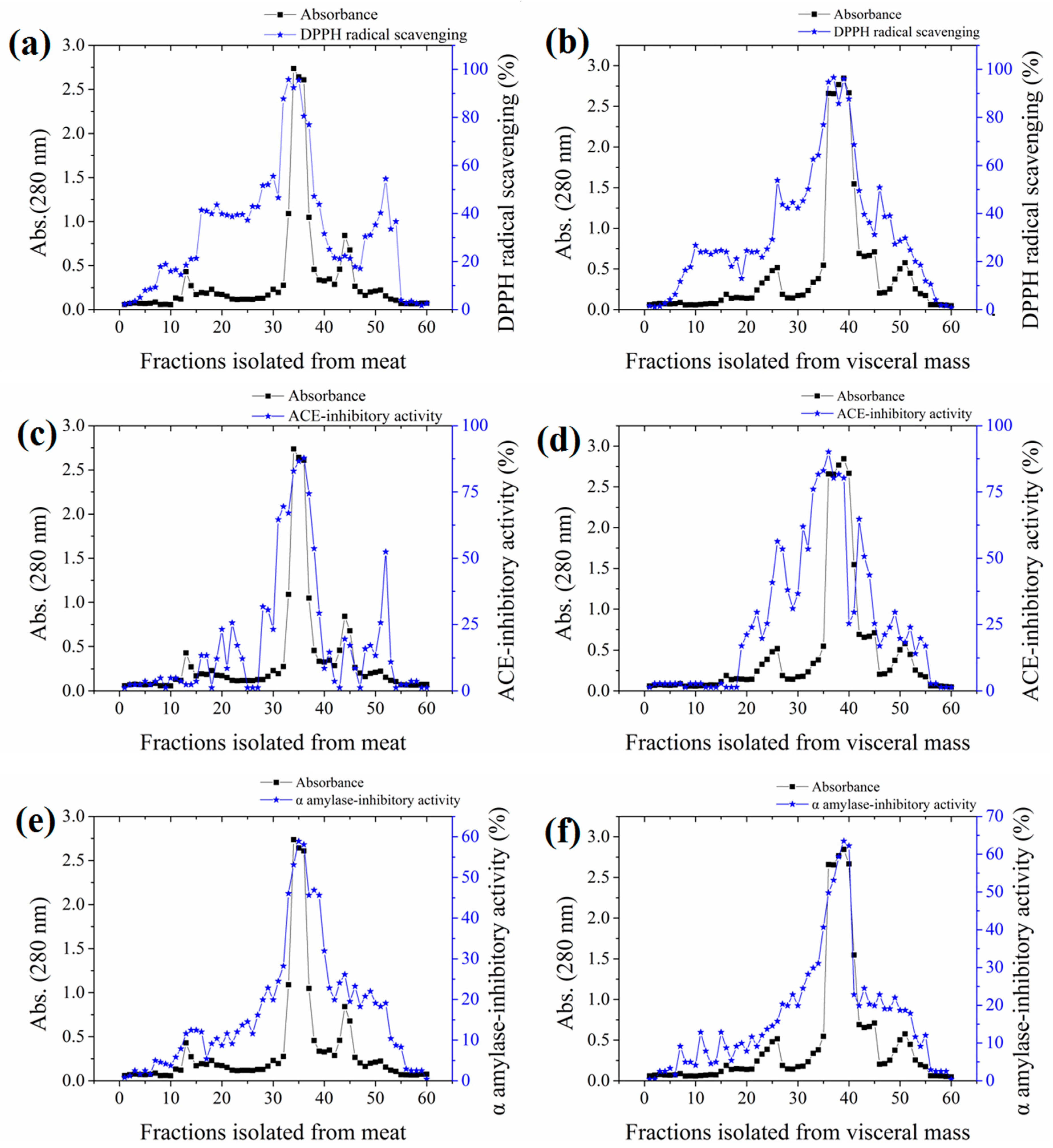
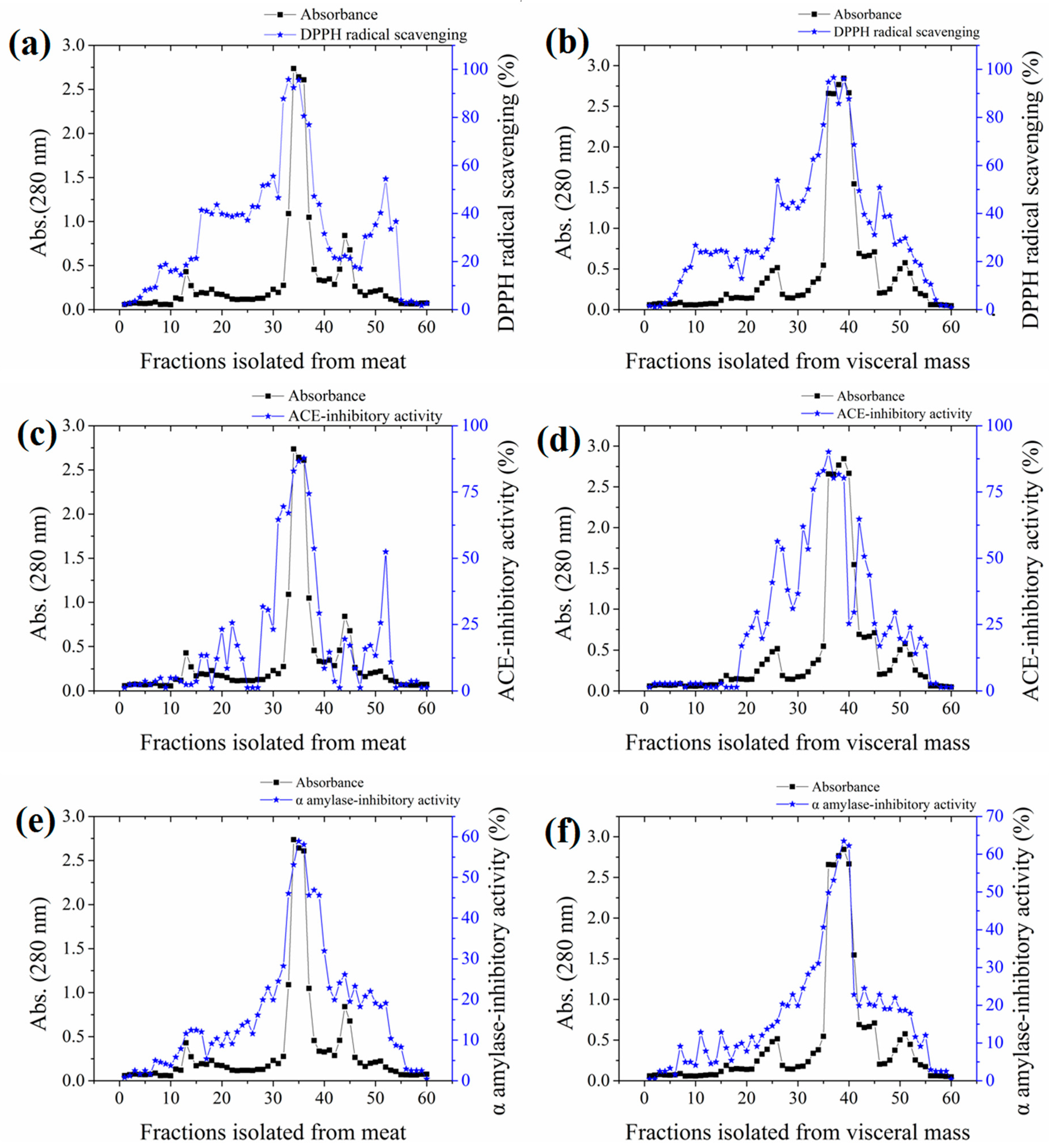
2.2. Bioassay-Guided Isolation of the Active Fraction
2.3. Molecular Weight Distribution
2.4. Amino Acid Profile of Active Fractions
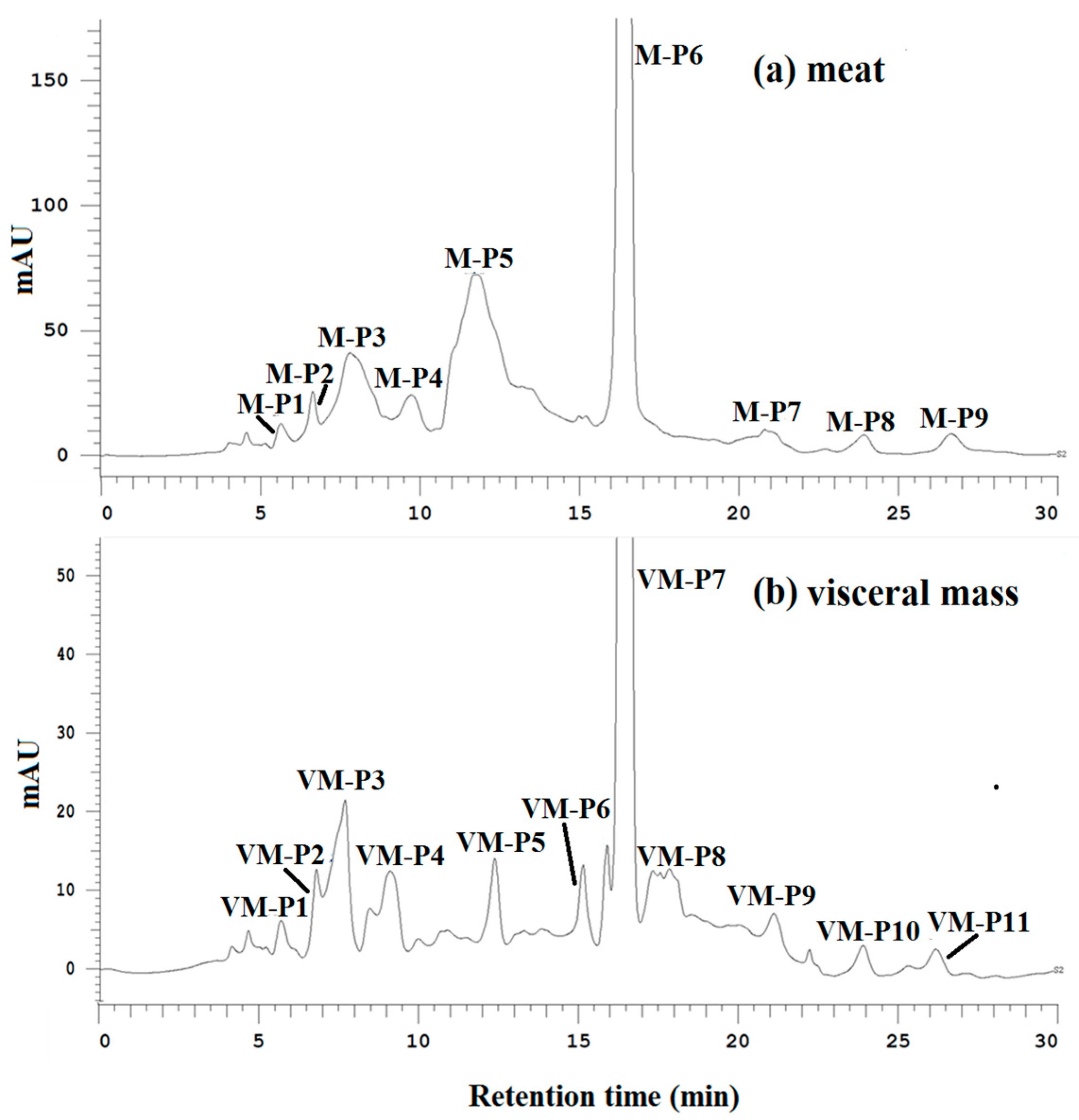
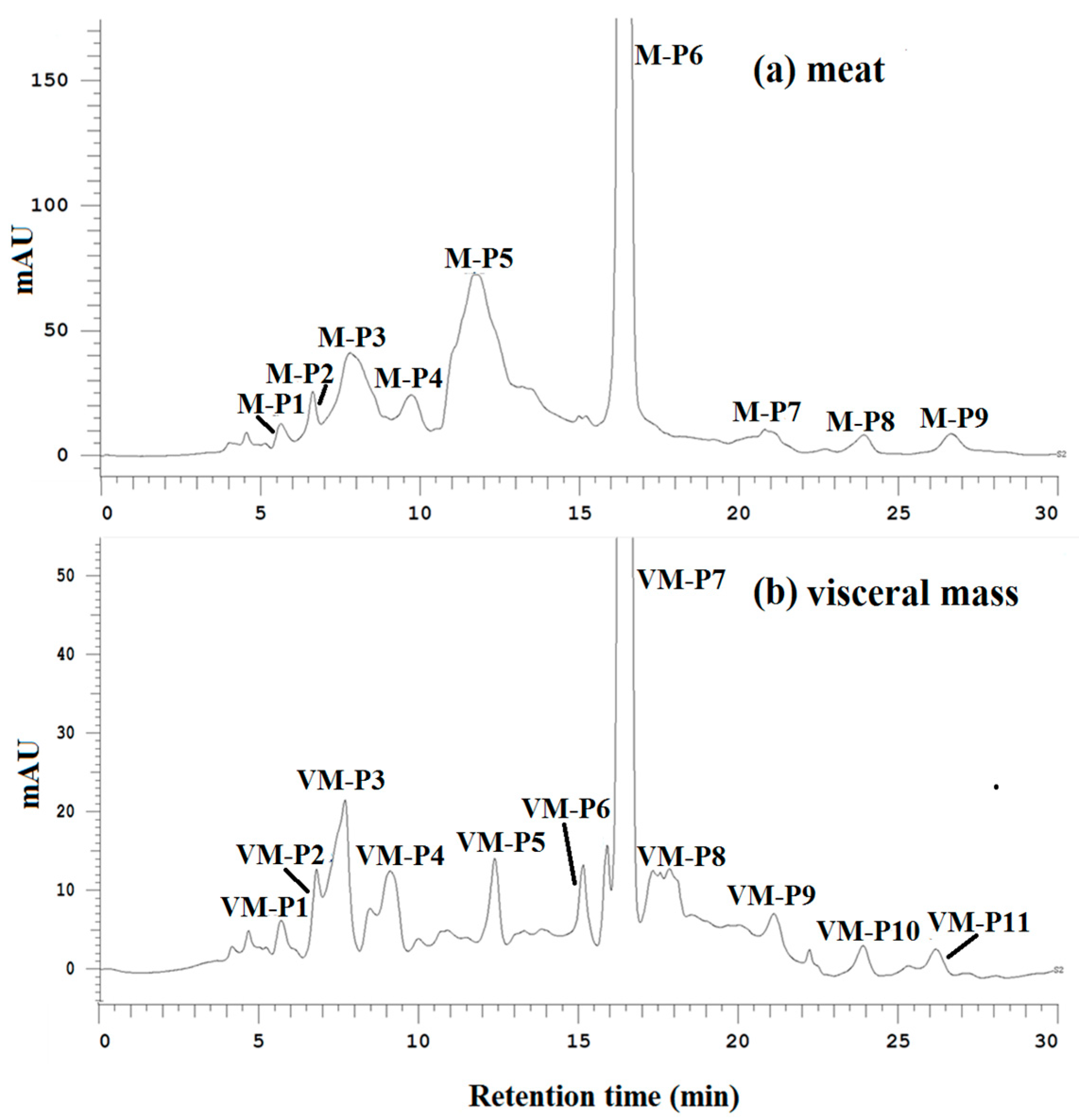
2.5. Bioassay-Guided Purification of Active Peptides
2.6. Amino Acid Sequence of Active Peptides
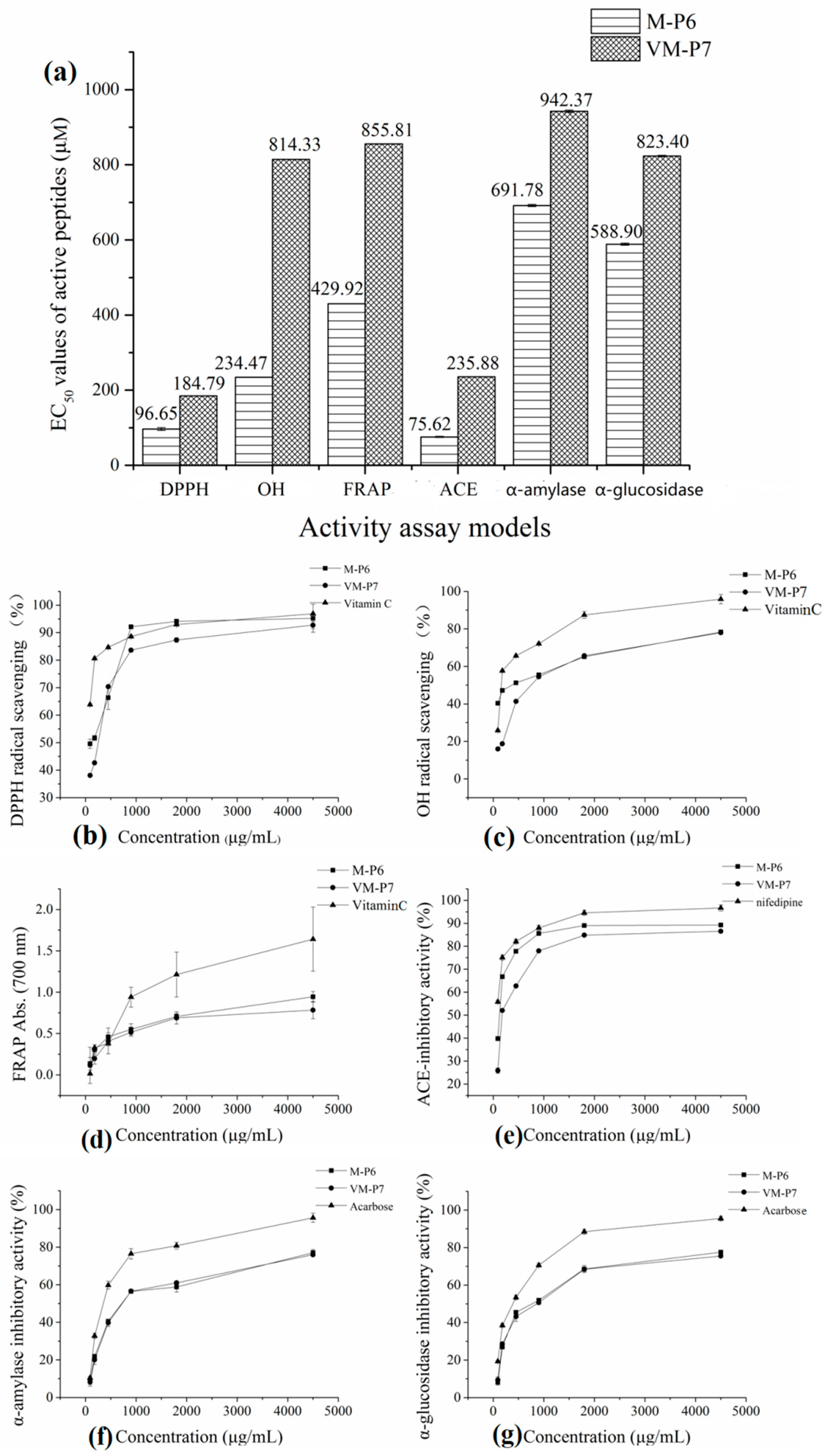
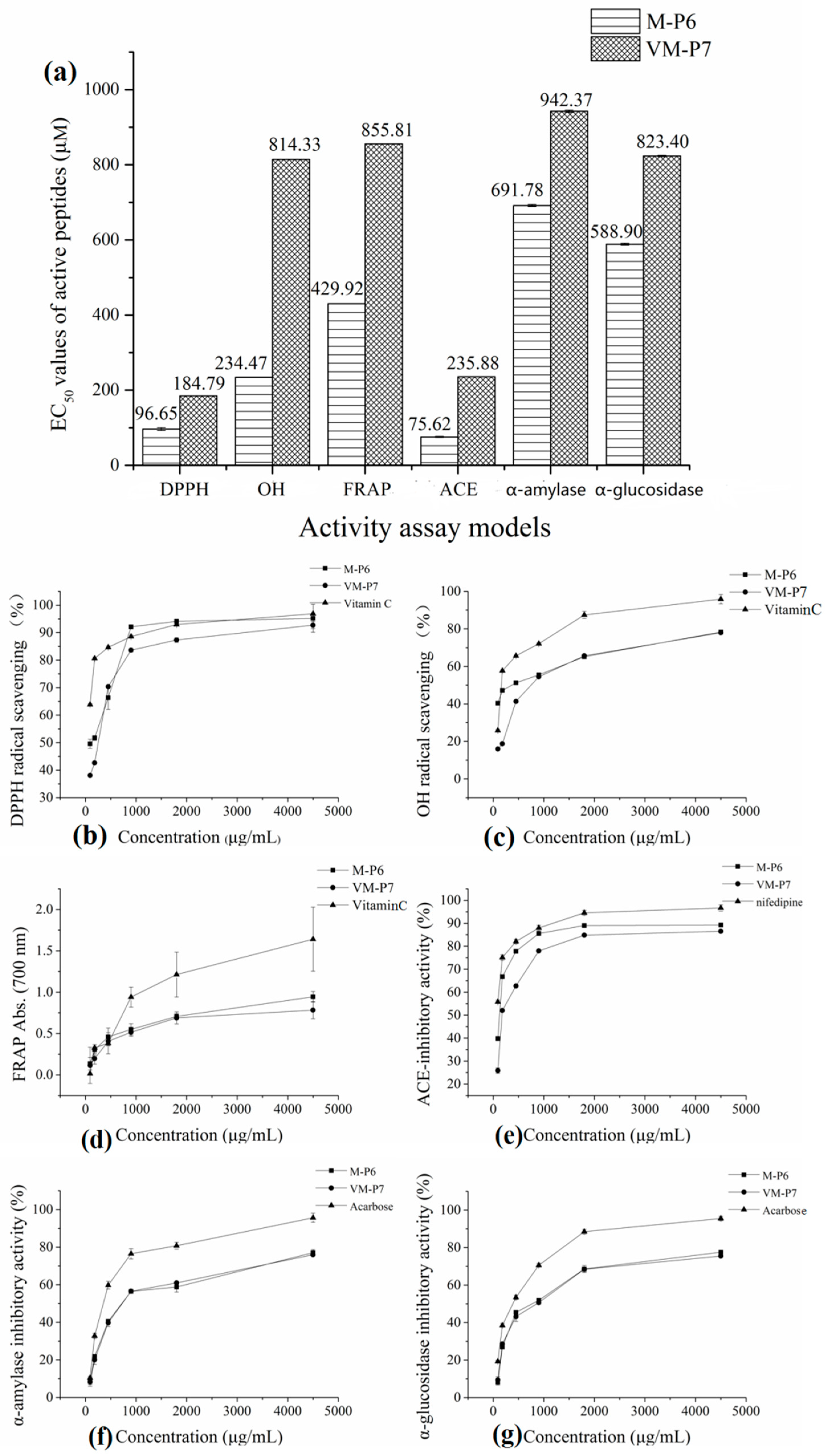
2.7. Analyses of Active Peptide Activity
2.7.1. In Vitro Antioxidant Activity
2.7.2. ACE-Inhibitory Activity
2.7.3. Anti-Diabetic Activity
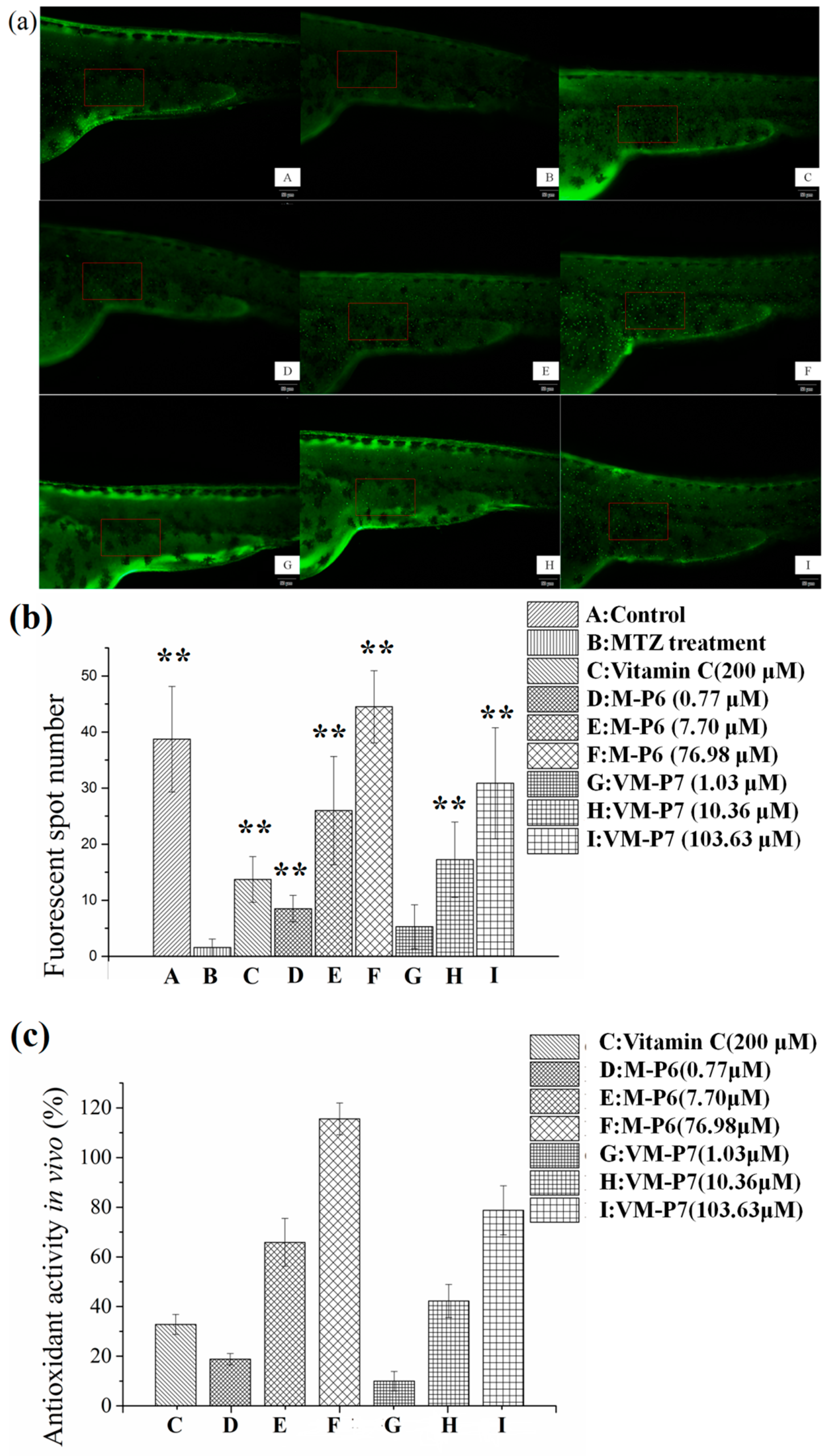
2.7.4. In Vivo Antioxidant Activity in Zebrafish Embryos
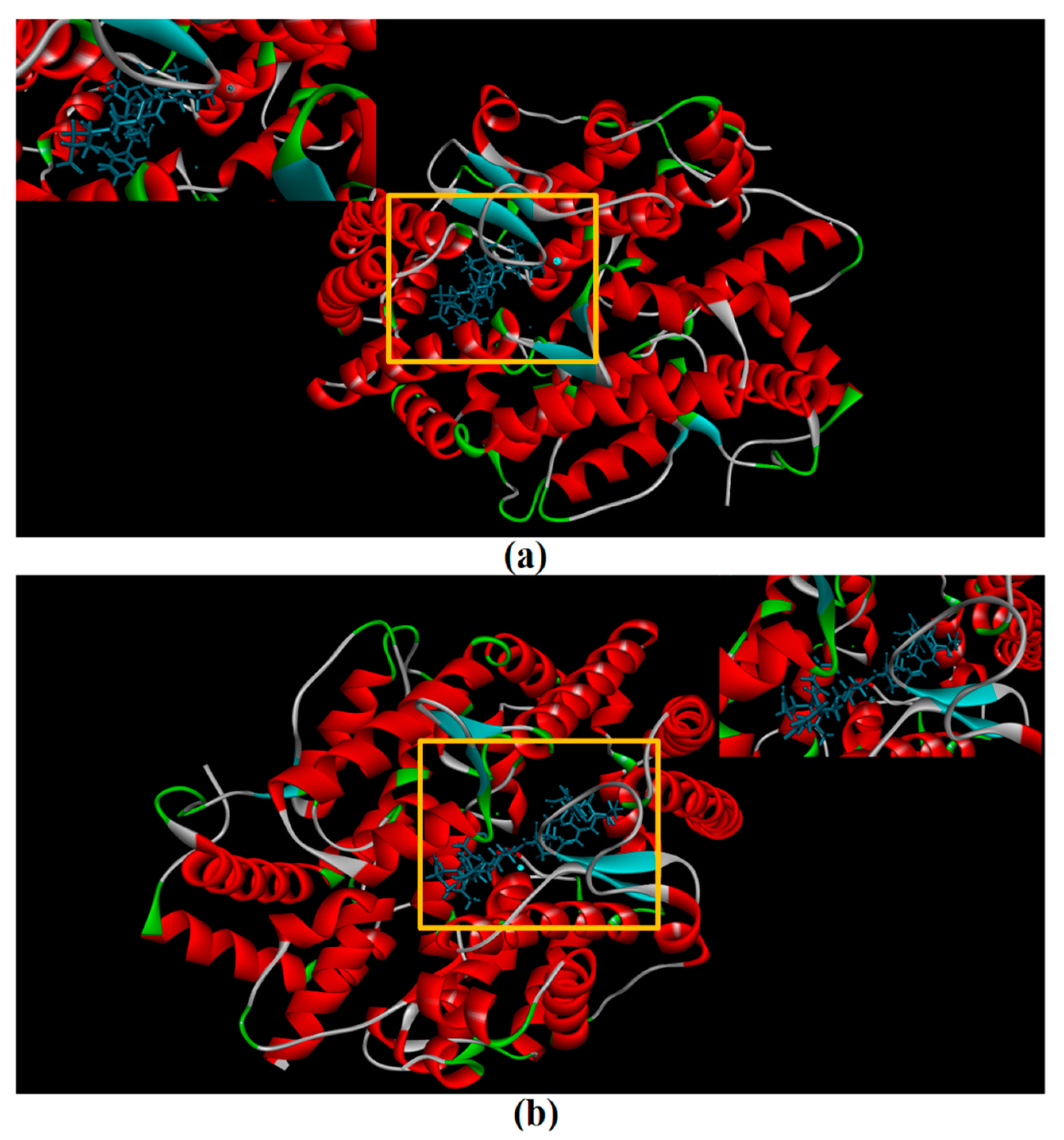
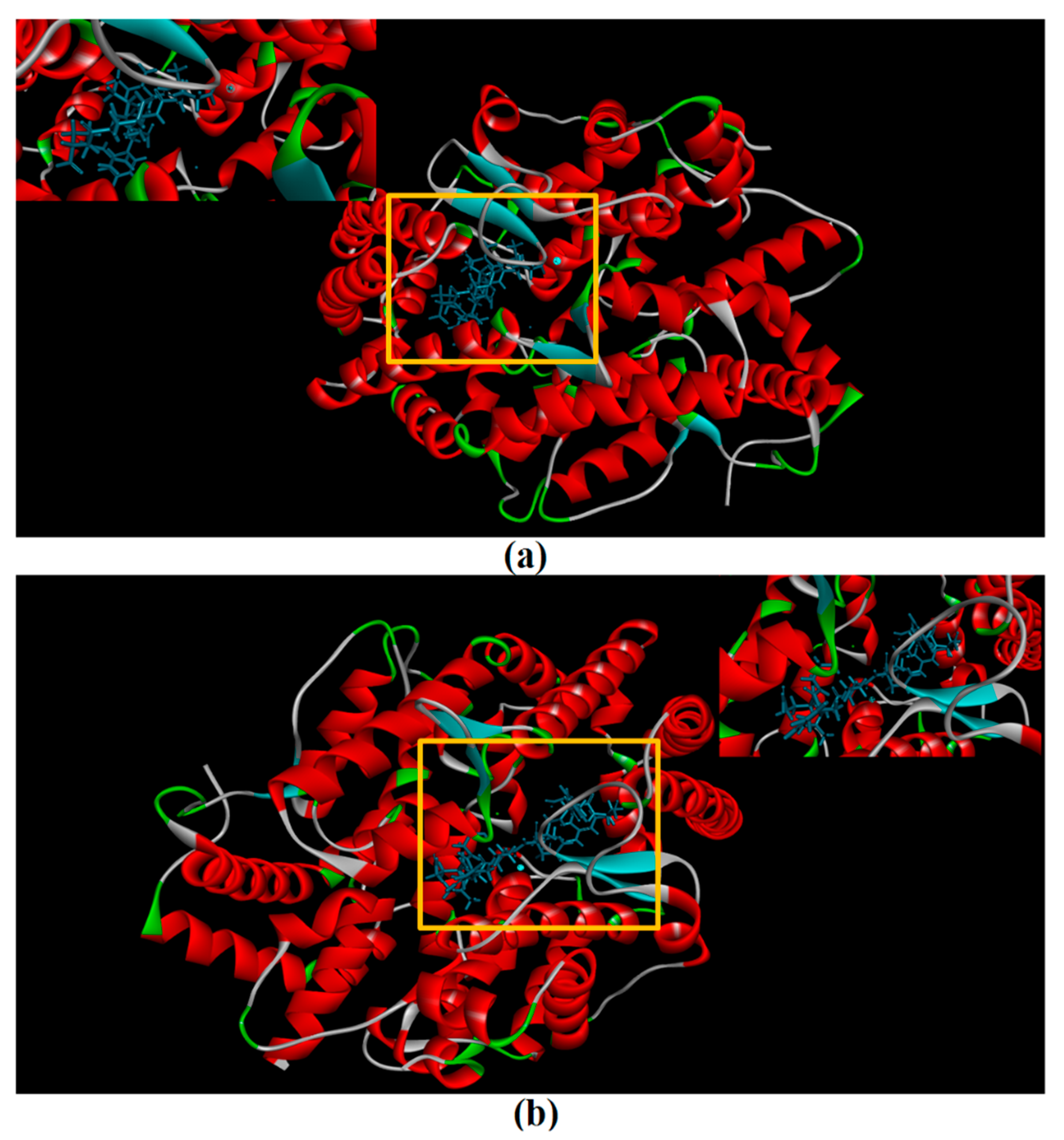
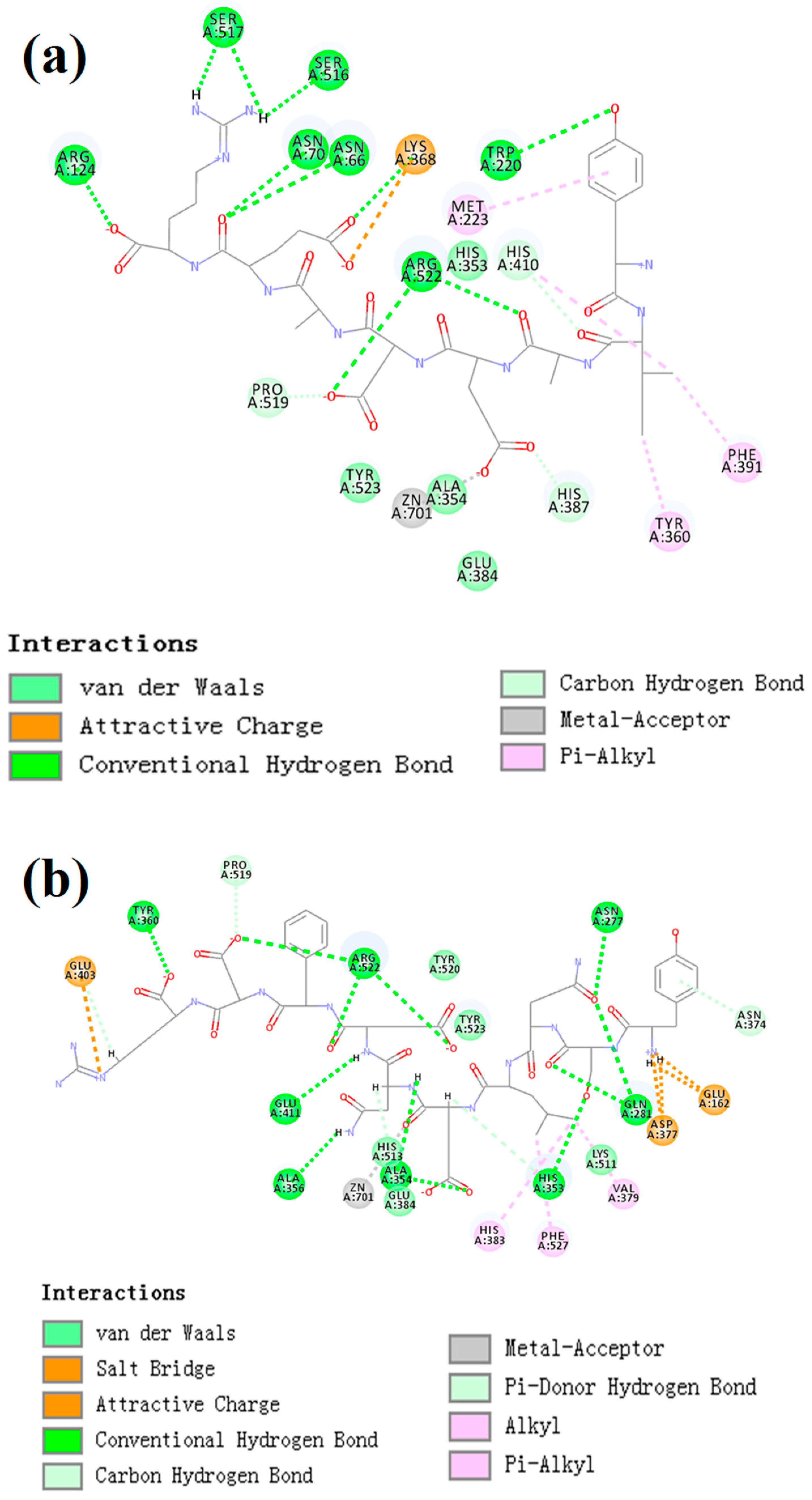
2.8. Molecular Docking Simulation
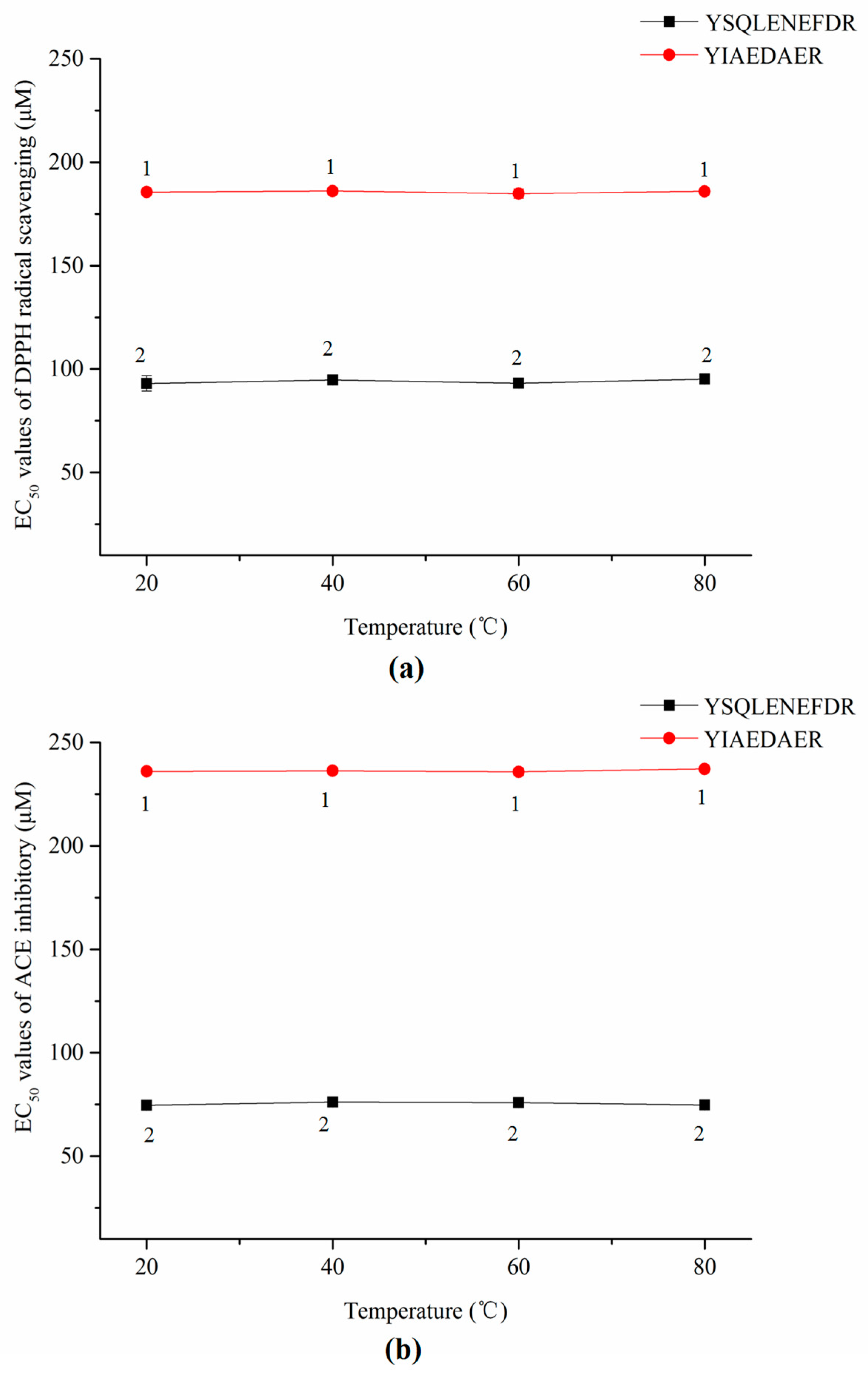
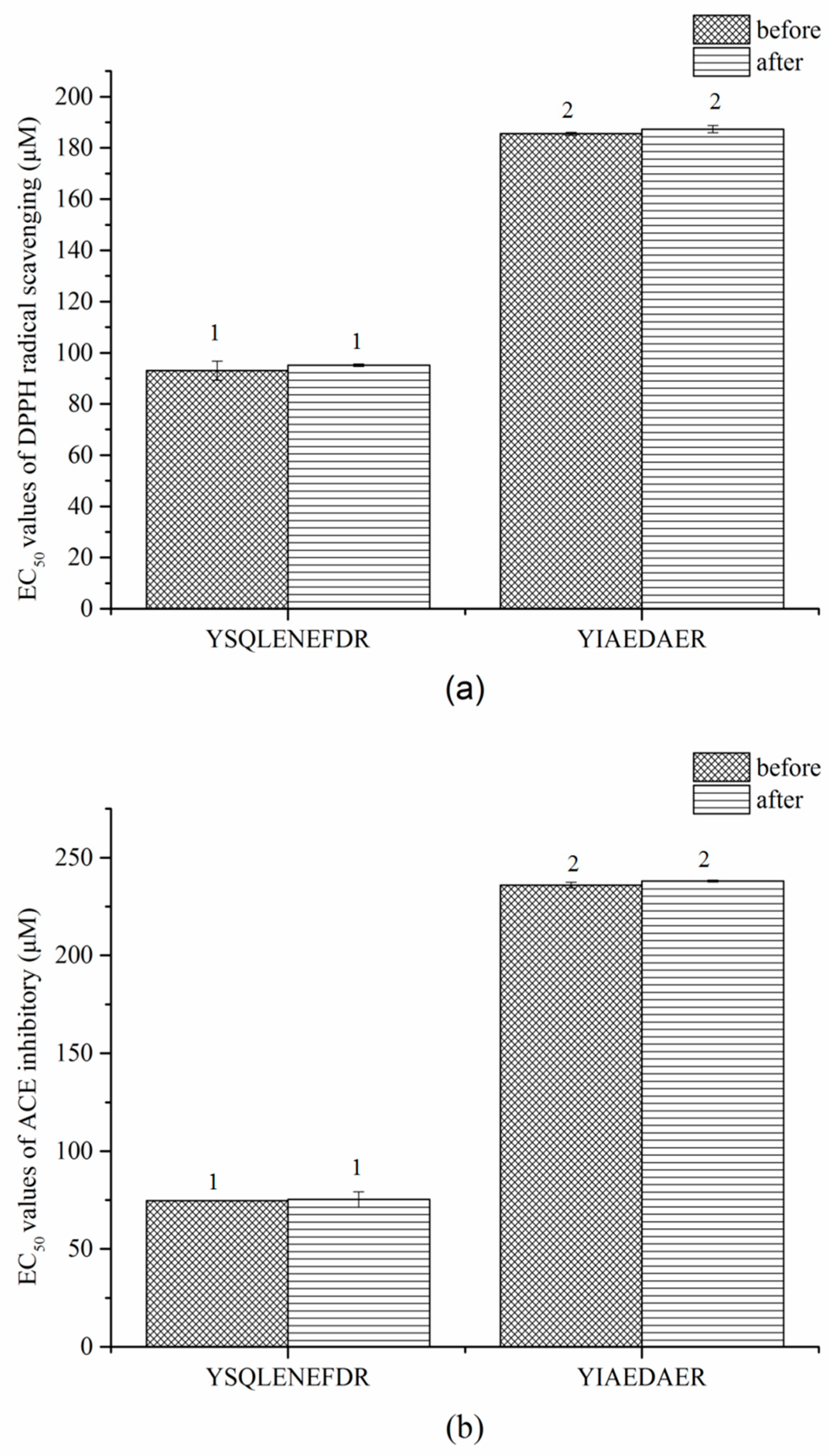
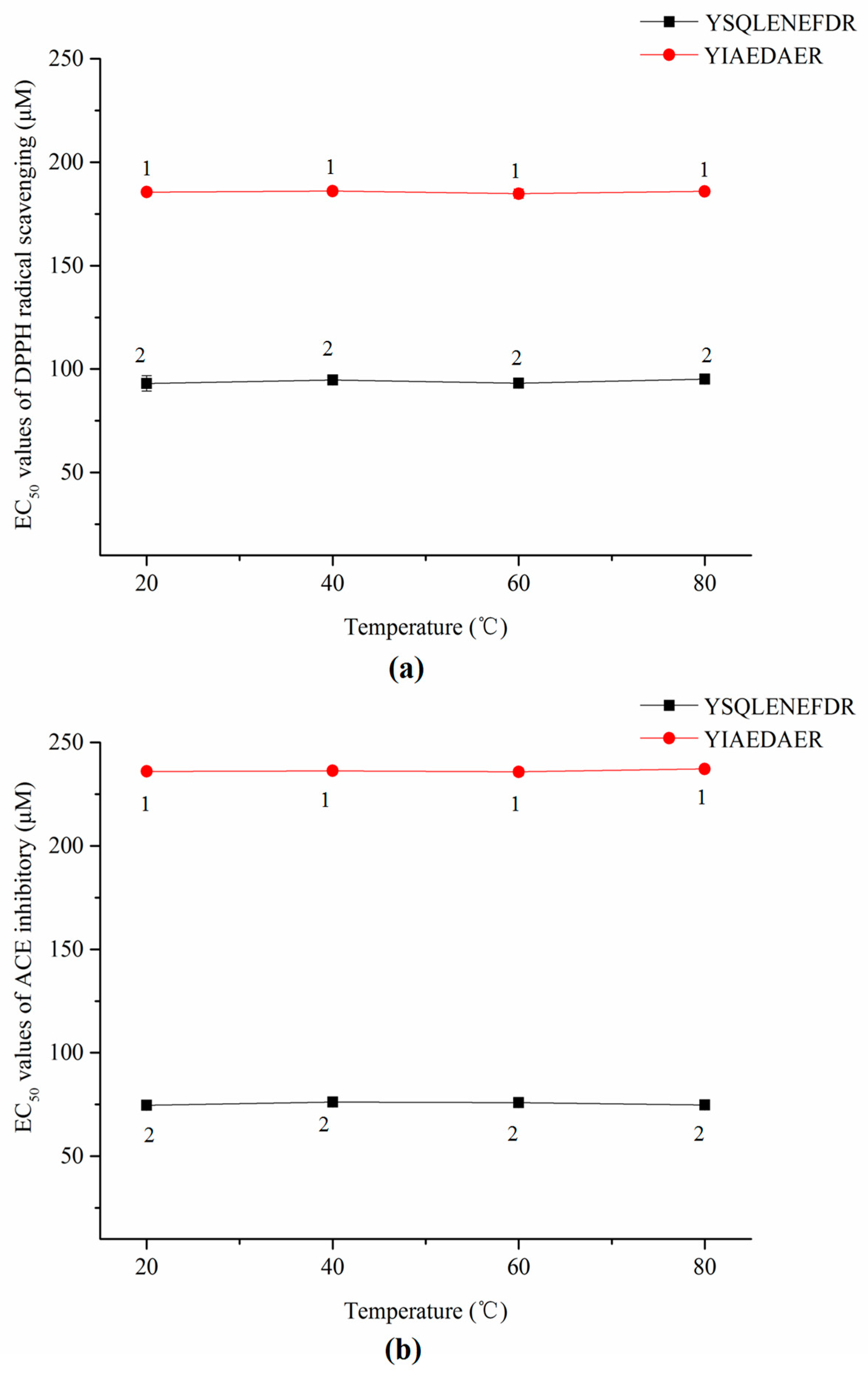
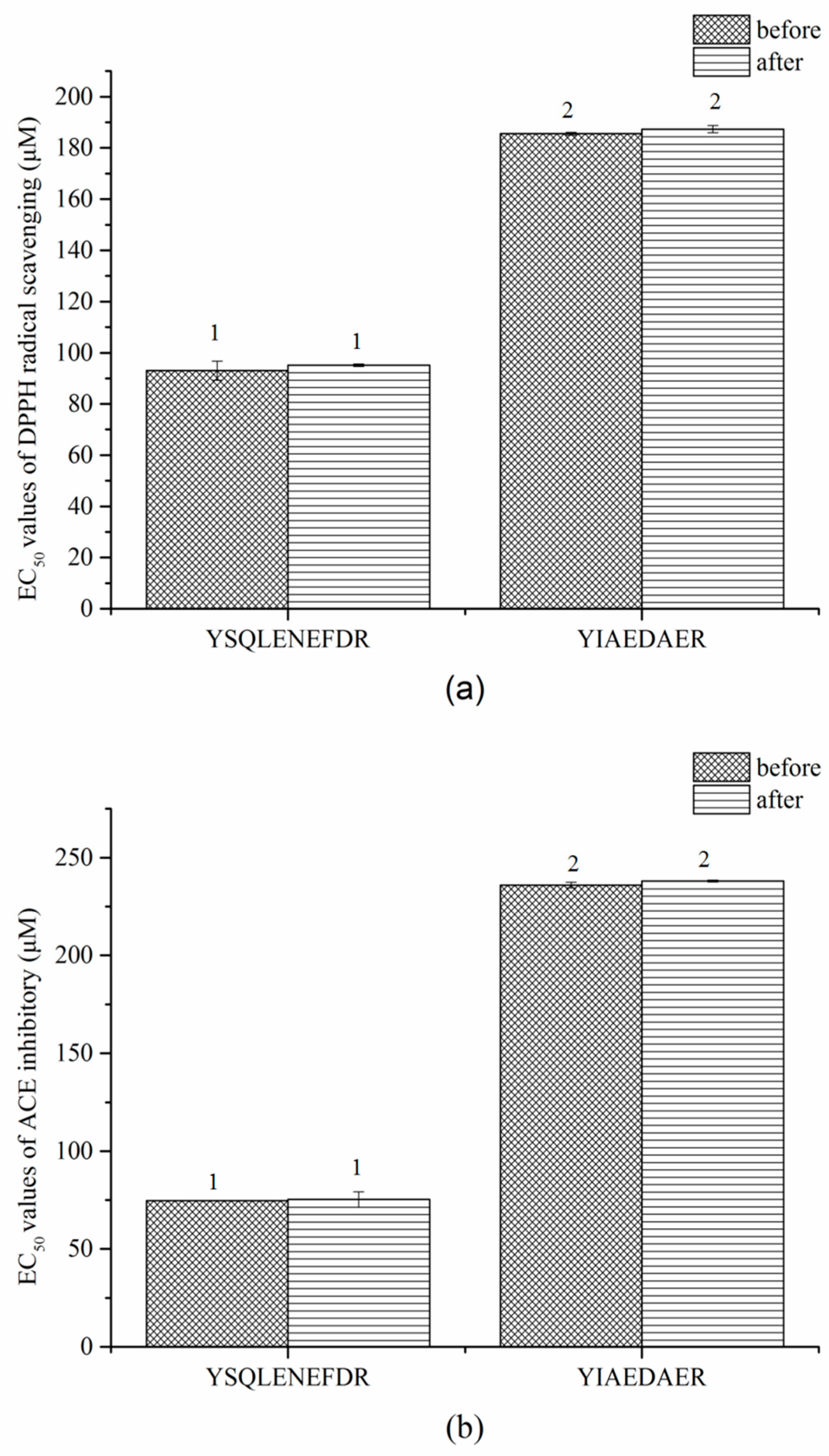
2.9. Stablity of Synthetic Peptides against Thermal and Gastrointestinal Digestion Treatments
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Reagents and Animals
4.3. Zebrafish Maintenance and Embryo Handling
4.4. Proximate Composition
4.5. Peptide Extraction
4.6. Bioassay-Guided Isolation of Active Fractions
4.7. Molecular Weight Distribution of Active Fractions
4.8. Amino Acid Compositions of Active Fractions
4.9. Bioassay-Guided Purification of Active Peptides
4.10. Identification of Active Peptide Sequences by Nano-LC-LTQ-Orbitrap-MS/MS
4.11. Determination of Activities
4.11.1. DPPH Radical Scavenging Activity
4.11.2. Ferric Reducing Capacity
4.11.3. Hydroxyl Radical Scavenging Activity
4.11.4. Determination of Antioxidative Activity in Zebrafish Embryos
4.11.5. Determination of ACE-Inhibitory Activity
4.11.6. Determination of Anti-Diabetic Activity
4.12. Peptide Synthesis
4.13. Molecular Docking
4.14. Stability against Thermal and Gastrointestinal Digestion Treatments
4.15. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef] [PubMed]
- Negi, B.; Kumar, D.; Rawat, D.S. Marine peptides as anticancer agents: A remedy to mankind by nature. Curr. Protein Pept. Sci. 2017, 18, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Kim, S.K. Bioactive peptide of marine origin for the prevention and treatment of non-communicable diseases. Mar. Drugs 2017, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Guadalupe, M.; Suarez, J.; Armando, B.H.; Josafat, M.; Ezquerra, B. Bioactive peptides and depsipeptides with anticancer potential: Sources from marine animals. Mar. Drugs 2012, 10, 963–986. [Google Scholar] [CrossRef]
- Luo, F.L.; Xing, R.; Wang, X.Q.; Peng, Q.; Li, P. Proximate composition, amino acid and fatty acid profiles of marine snail Rapana venosa meat, visceral mass and operculum. J. Sci. Food Agric. 2017, 97, 5361–5368. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.H.; Craik, D.J.; Kaas, Q.; Schroeder, C.I. Bioactive compounds isolated from neglected predatory marine gastropods. Mar. Drugs 2018, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- Ponder, W.F. A review of the New Zealand recent and fossil species of Buccinulum deshayes (Mollusca: Neogastropoda: Buccinidae). J. R. Soc. N. Z. 1971, 1, 231–283. [Google Scholar] [CrossRef]
- Hao, Z.L.; Yang, L.M.; Zhan, Y.Y.; Tian, Y.; Mao, J.X.; Wang, L.; Chang, Y.Q. The complete mitochondrial genome of Neptunea arthritica cumingii Crosse, (Gastropoda: Buccinidae). Mitochondrial DNA Part B Resour. 2016, 1, 220–221. [Google Scholar] [CrossRef]
- Lombardo, R.C.; Goshima, S. Sexual conflict in Neptunea arthritica: The power asymmetry and female resistance. J. Mar. Biol. Assoc. 2011, 91, 251–256. [Google Scholar] [CrossRef]
- An, J.E.; Choi, J.D.; Ryu, D.K. Age and Growth of the Neptunea (Barbitonia) Arthritica cumingii in the west sea of Korea. Korean J. Malacol. 2014, 30, 25–32. [Google Scholar] [CrossRef]
- Miranda, R.M.; Fujinaga, K.; Ilano, A.S.; Nakao, S. Effects of imposex and parasite infection on the reproductive features of the Neptune whelk Neptunea arthritica. Mar. Biol. Res. 2009, 5, 268–277. [Google Scholar] [CrossRef]
- He, J.Z. Analysis of trace elements and edibleness of Neptunea cumingi tissue. Food Sci. 2010, 31, 181–184. [Google Scholar] [CrossRef]
- Esipov, A.V.; Busarova, N.G.; Isai, S.V. Composition and fatty-acid contents of the commercial mollusk Neptunea arthritica. Chem. Nat. Comp. 2014, 50, 1099–1100. [Google Scholar] [CrossRef]
- Asano, M.; Ito, M. Occurrence of tetramine and choline compounds in the salivary gland of a marine gastropod Neptunea arthritica, Bernardi. J. Clin. Gastroenterol. 1959, 1, 91–92. [Google Scholar] [CrossRef]
- Lloyd, P.E. Cardioactive neuropeptides in gastropods. Fed. Proc. 1982, 41, 2948–3000. [Google Scholar] [PubMed]
- Liu, K.C.; Biology Institute, Qilu University of Technology, Shandong Academy of Sciences, Jinan, China. Personal communication, 2008.
- Hill, A.; Mesens, N.; Steemans, M.; Xu, J.J.; Aleo, M.D. Comparisons between in vitro whole cell imaging and in vivo zebrafish-based approaches for identifying potential human hepatotoxicants earlier in pharmaceutical development. Drug. Metab. Rev. 2012, 44, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Ingham, P.W. The power of the Zebrafish for disease analysis. Hum. Mol. Genet. 2009, 18, R107–R112. [Google Scholar] [CrossRef] [PubMed]
- Curado, S.; Stainier, D.Y.R.; Anderson, R.M. Nitroreductase-mediated cell/tissue ablation in zebrafish: A spatially and temporally controlled ablation method with applications in developmental and regeneration studies. Nat. Protoc. 2018, 3, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.A.; Conteh, A.M.; Sorrell, C.A.; Mirmira, A.; Tersey, S.A.; Mirnira, R.G.; Linnemann, A.K.; Anderson, R.M. An In Vivo Zebrafish Model for Interrogating ROS-Mediated Pancreatic β-Cell Injury, Response, and Prevention. Oxid. Med. Cell. Long. 2018, 2018, 1324739. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Chu, C.Y.; Chen, T.H.; Lee, S.J.; Shen, C.N.; Ning, C.; Hisiao, C.D. Establishment of a Transgenic Zebrafish Line for Superficial Skin Ablation and Functional Validation of Apoptosis Modulators In Vivo. PLoS ONE 2011, 6, e20654. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Moosman, B.; Behl, C. Secretory peptide hormones are biochemical antioxidants: Structure-activity relationship. Mol. Pharmacol. 2002, 61, 260–268. [Google Scholar] [CrossRef]
- Dong, Z.Y.; Tian, G.; Xu, Z.G.; Li, M.Y.; Zhou, Y.J.; Ren, H. Antioxidant Activities of Peptide Fractions Derived from Freshwater Mussel Protein Using Ultrasound-Assisted Enzymatic Hydrolysis. Czech J. Food Sci. 2017, 35, 328–338. [Google Scholar] [CrossRef]
- Liu, R.; Zheng, W.W.; Li, J.; Wang, L.C.; Wu, H.; Wang, X.Z.; Shi, L. Rapid identification of bioactive peptides with antioxidant activity from the enzymatic hydrolysate of Mactra veneriformis by UHPLC-Q-TOF mass spectrometry. Food Chem. 2015, 167, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Miguel, M.; Bartolomé, B.; Lopez-Fandino, R. Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Protect. 2004, 67, 1939–1944. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Bandara, N.; Wu, J.P. Purification and characterization of antioxidant peptides from enzymatically hydrolyzed chicken egg white. Food Chem. 2015, 188, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Soffer, R.L. Angiotensin-converting enzyme and the regulation of vasoactive peptides. Annu. Rev. Biochem. 1976, 45, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.M.; Liu, H.Z.; Liu, L.; Hu, H.; Wang, Q.; Adhikari, B. Isolation, Purification and Molecular Mechanism of a Peanut Protein-Derived ACE-Inhibitory Peptide. PLoS ONE 2014, 9, e111188. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.; Hayes, M.; Carney, B. Angiotensin-I-converting enzyme and prolyl endopeptidase inhibitory peptides from natural sources with a focus on marine processing by-products. Food Chem. 2011, 129, 235–244. [Google Scholar] [CrossRef]
- Segura-Campos, M.R.; Peralta-González, F.; Castellanos-Ruelas, A.; Chel-Guerrero, L.A.; Betancur-Ancona, D.A. Effect of Jatropha curcas peptide fractions on the angiotensin I-converting enzyme inhibitory activity. Biomed. Res. Int. 2013, 2013, 541974. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.A.; FitzGerald, R.J. Angiotensin converting enzyme inhibitory peptides derived from food proteins: Biochemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 773–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sun, L.C.; Yan, L.J.; Lin, Y.C.; Liu, G.M.; Cao, M.J. Production, optimisation and characterisation of angiotensin converting enzyme inhibitory peptides from sea cucumber (Stichopus japonicus) gonad. Food Funct. 2018, 9, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Uraipong, C.; Zhao, J. Rice bran protein hydrolysates exhibit strong in vitro α-amylase, β-glucosidase and ACE-inhibition activities. J. Sci. Food Agric. 2016, 96, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H.; Lucius, A.; Meyer, T.; de Mejia, E.G. Cultivar evaluation and effect of fermentation on antioxidant capacity and in vitro inhibition of a-amylase and a-glucosidase by highbush blueberry (Vaccinium corombosum). J. Agric. Food Chem. 2011, 59, 8923–8930. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Martíne-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, a-amylase and a-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef]
- Siow, H.L.; Gan, C.Y. Extraction, identification, and structure-activity relationships of antioxidative and a-amylase inhibitory peptides from cumin seeds (Cuminum cyminum). J. Funct. Foods 2016, 22, 1–12. [Google Scholar] [CrossRef]
- Ren, Y.; Liang, K.; Jin, Y.Q.; Zhang, M.M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel a-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Pina, A.S.; Roque, A.C. Studies on the molecular recognition between bioactive peptides and angiotensin-converting enzyme. J. Mol. Recognit. 2009, 22, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.D.; Cao, J.X.; Guo, H.Q.; Zhao, B. Studies on purification and the molecular mechanism of a novel ACE inhibitory peptide from whey protein hydrolysate. Food Chem. 2012, 130, 121–126. [Google Scholar] [CrossRef]
- Zhao, Y.H.; Li, B.F.; Liu, Z.Y.; Dong, S.Y.; Zhao, X.; Zeng, M.Y. Antihypertensive effect and purification of an ACE inhibitory peptide from sea cucumber gelatin hydrolysate. Process Biochem. 2007, 42, 1586–1591. [Google Scholar] [CrossRef]
- Deng, Z.Z.; Liu, Y.J.; Wang, J.; Wu, S.H.; Geng, L.H.; Sui, Z.H.; Zhang, Q.B. Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs). Mar. Drugs 2018, 16, 299. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.P.; Ding, X.L. Characterization of inhibition and stability of soy-protein derived angiotensin I-converting enzyme inhibitory peptides. Food Res. Int. 2002, 35, 367–375. [Google Scholar] [CrossRef]
- Erak, M.; Bellmann-Sickert, K.; Els-Heindl, S.; Beck-Sickinger, A.G. Peptide chemistry toolbox—Transforming natural peptides into peptides therapeutics. Bioorg. Med. Chem. 2018, 26, 2759–2765. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 19th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Li, Z.R.; Luo, H.Y. Influence of Amino Acid Compositions and Peptide Profiles on Antioxidant Capacities of Two Protein Hydrolysates from Skipjack Tuna (Katsuwonus pelamis) Dark Muscle. Mar. Drugs 2015, 13, 2580–2601. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Han, X.D.; Zhang, M.T.; Xu, W.W.; Zhou, W.; Jiao, H. Application AQC as a pre-column derivatization reagent for HPLC determination of free amino acids in Borojo sorbilis cuter. Acta Sci. Nat. Univ. Sunyatseni 2013, 52, 100–104. [Google Scholar] [CrossRef]
- Lassoued, I.; Mora, L.; Barkia, A.; Aristory, M.C.; Nasri, M.; Toldráb, F. Bioactive peptides identified in thornback ray skin’s gelatin hydrolysates by proteases from Bacillus subtilis and Bacillus amyloliquefaciens. J. Proteomic 2015, 128, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Moon, S.H.; Kim, H.S.; Park, E.; Ahn, D.; Paik, H.D. Antioxidant and anticancer effects of functional peptides from Ovotransferrin hydrolysates. J. Sci. Food Agric. 2017, 97, 4857–4864. [Google Scholar] [CrossRef] [PubMed]
- Moayedi, A.; Mora, L.; Aristoy, M.-C.; Hashemi, M.; Safari, M.; Toldrá, F. ACE inhibitory and antioxidant activities of peptide fragments obtained from tomato processing by-products fermented using Bacillus subtilis: Effect of amino acid composition and peptides molecular mass distribution. Appl. Biochem. Biotechnol. 2017, 181, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.G.; Chu, J.; Chen, X.Q.; Wang, J.G.; Wu, X.M.; Liu, K.C. Study on the antioxidant activity evaluation of Jujube(Ziziphus) leaf flavonoids in vitro and zebrafish (Danio rerio) with fluorescent skin. Sci. Technol. Food Ind. 2014, 35, 58–61. [Google Scholar] [CrossRef]
- Chen, J.W.; Wang, Y.M.; Zhong, Q.X.; Wu, Y.N.; Xia, W.S. Purification and characterization of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide derived from enzymatic hydrolysate of grass carp protein. Peptides 2012, 33, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.D.; Liu, Y.; Wang, G.Y.; Sun, S.S.; Liu, R.; Hong, B.H.; Gao, R.; Bai, K.K. Processing Optimization and Characterization of Angiotensin-I-Converting Enzyme Inhibitory Peptides from Lizardfish (Synodus macrops) Scale Gelatin. Mar. Drugs 2018, 16, 228. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Meat (g/kg) | Visceral Mass (g/kg) |
|---|---|---|
| asp | 42.24 ± 7.69 | 73.39 ± 9.67 ** |
| glu | 99.87 ± 5.60 | 65.21 ± 6.73 ** |
| ser | 21.82 ± 1.09 | 24.55 ± 5.99 * |
| gly | 30.65 ± 3.15 | 36.29 ± 7.46 * |
| his | 31.12 ± 1.03 | 10.90 ± 3.84 ** |
| arg | 388.77 ± 5.93 | 366.20 ± 8.02 |
| thr | 21.78 ± 0.88 | 18.03 ± 8.46 |
| ala | 98.54 ± 5.92 | 132.58 ± 7.21 ** |
| pro | 94.58 ± 5.26 | 47.18 ± 0.57 ** |
| cys | 1.04 ± 0.11 | 0.72 ± 0.11 * |
| tyr | 5.05 ± 0.67 | 3.15 ± 5.60 ** |
| val | 24.60 ± 2.55 | 25.50 ± 5.15 |
| met | 3.09 ± 0.95 | 3.24 ± 0.15 |
| lys | 14.63 ± 1.25 | 7.23 ± 1.50 ** |
| ile | 18.10 ± 1.16 | 19.92 ± 4.15 |
| leu | 34.87 ± 1.18 | 32.34 ± 4.81 |
| phe | 51.34 ± 1.61 | 3.98 ± 0.96 ** |
| Sum | 982.11 ± 3.20 | 870.40 ± 2.23 ** |
| EAA | 168.42 ± 1.03 | 110.23 ± 4.43 ** |
| HAA | 332. 56 ± 8.61 | 268.6 ± 3.56 ** |
| AAA | 56.40 ± 8.61 | 7.13 ± 3.83 ** |
| PCAA | 434.52 ± 6.95 | 384.33 ± 5.73 |
| NCAA | 140.78 ± 1.81 | 138.60 ± 6.29 |
| Origin | Fractions | DPPH Radical Scavenging Activity (%) | ACE-Inhibitory Activity (%) | α-Amylase Inhibitory Activity (%) |
|---|---|---|---|---|
| M-F | M-P1 | 14.73 ± 3.36 | 12.25± 1.67 | 10.25 ± 2.00 |
| M-P2 | 25.03 ± 3.11 | 15.26 ± 3.34 | 12.55 ± 0.62 | |
| M-P3 | 20.75 ± 2.39 | 16.76 ± 3.12 | 15.26 ± 1.30 | |
| M-P4 | 27.25 ±1.82 | 20.77 ± 0.85 | 15.76 ± 1.26 | |
| M-P5 | 33.80 ± 2.93 | 23.07 ± 2.61 | 24.55 ± 0.29 | |
| M-P6 | 91.87 ± 0.62 | 84.81 ± 0.35 | 56.15 ± 1.64 | |
| M-P7 | 34.80 ± 1.22 | 28.20 ± 3.54 | 17.85 ± 2.77 | |
| M-P8 | 32.60 ± 2.17 | 26.45 ± 1.90 | 16.45 ± 3.01 | |
| M-P9 | 35.25 ± 1.40 | 25.85 ± 1.37 | 15.85 ± 2.27 | |
| VM-F | VM-P1 | 14.63 ± 3.38 | 12.50 ± 2.26 | 11.50 ± 2.46 |
| VM-P2 | 17.03 ± 2.07 | 14.56 ± 2.33 | 13.55 ± 2.61 | |
| VM-P3 | 28.70 ± 0.99 | 20.56 ± 1.10 | 15.55 ± 1.36 | |
| VM-P4 | 24.77 ± 3.71 | 23.85 ± 3.88 | 16.35 ± 1.34 | |
| VM-P5 | 33.73 ± 1.47 | 25.80 ± 3.81 | 16.30 ± 1.74 | |
| VM-P6 | 38.40 ± 1. 84 | 28.05 ± 2.80 | 18.05 ± 4.27 | |
| VM-P7 | 82.60 ± 0.86 | 74.95 ± 1.24 | 53.85 ± 0.92 | |
| VM-P8 | 35.60 ± 1.99 | 27.95 ± 0.76 | 26.95 ± 4.49 | |
| VM-P9 | 29.93 ± 0.71 | 24.95 ± 0.85 | 24.96 ± 0.79 | |
| VM-P10 | 28.03 ± 3.78 | 21.55 ± 1.64 | 11.56 ± 2.87 | |
| VM-P11 | 32. 90 ± 0.86 | 27. 35 ± 3.36 | 17. 91 ± 1.70 |
| Peptides | - CDocker Energy (kcal/mol) | - CDocker Interaction Energy (kcal/mol) |
|---|---|---|
| YIAEDAER | 174.672 | 130.72 |
| YSQLENEFDR | 193.884 | 175.07 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.-S.; Han, L.-W.; Shi, Y.-P.; Li, X.-B.; Zhang, X.-M.; Hou, H.-R.; Lin, H.-W.; Liu, K.-C. Two Novel Multi-Functional Peptides from Meat and Visceral Mass of Marine Snail Neptunea arthritica cumingii and Their Activities In Vitro and In Vivo. Mar. Drugs 2018, 16, 473. https://doi.org/10.3390/md16120473
Zhang S-S, Han L-W, Shi Y-P, Li X-B, Zhang X-M, Hou H-R, Lin H-W, Liu K-C. Two Novel Multi-Functional Peptides from Meat and Visceral Mass of Marine Snail Neptunea arthritica cumingii and Their Activities In Vitro and In Vivo. Marine Drugs. 2018; 16(12):473. https://doi.org/10.3390/md16120473
Chicago/Turabian StyleZhang, Shan-Shan, Li-Wen Han, Yong-Ping Shi, Xiao-Bin Li, Xuan-Ming Zhang, Hai-Rong Hou, Hou-Wen Lin, and Ke-Chun Liu. 2018. "Two Novel Multi-Functional Peptides from Meat and Visceral Mass of Marine Snail Neptunea arthritica cumingii and Their Activities In Vitro and In Vivo" Marine Drugs 16, no. 12: 473. https://doi.org/10.3390/md16120473
APA StyleZhang, S.-S., Han, L.-W., Shi, Y.-P., Li, X.-B., Zhang, X.-M., Hou, H.-R., Lin, H.-W., & Liu, K.-C. (2018). Two Novel Multi-Functional Peptides from Meat and Visceral Mass of Marine Snail Neptunea arthritica cumingii and Their Activities In Vitro and In Vivo. Marine Drugs, 16(12), 473. https://doi.org/10.3390/md16120473