Diversity and Antimicrobial Potential of Predatory Bacteria from the Peruvian Coastline

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
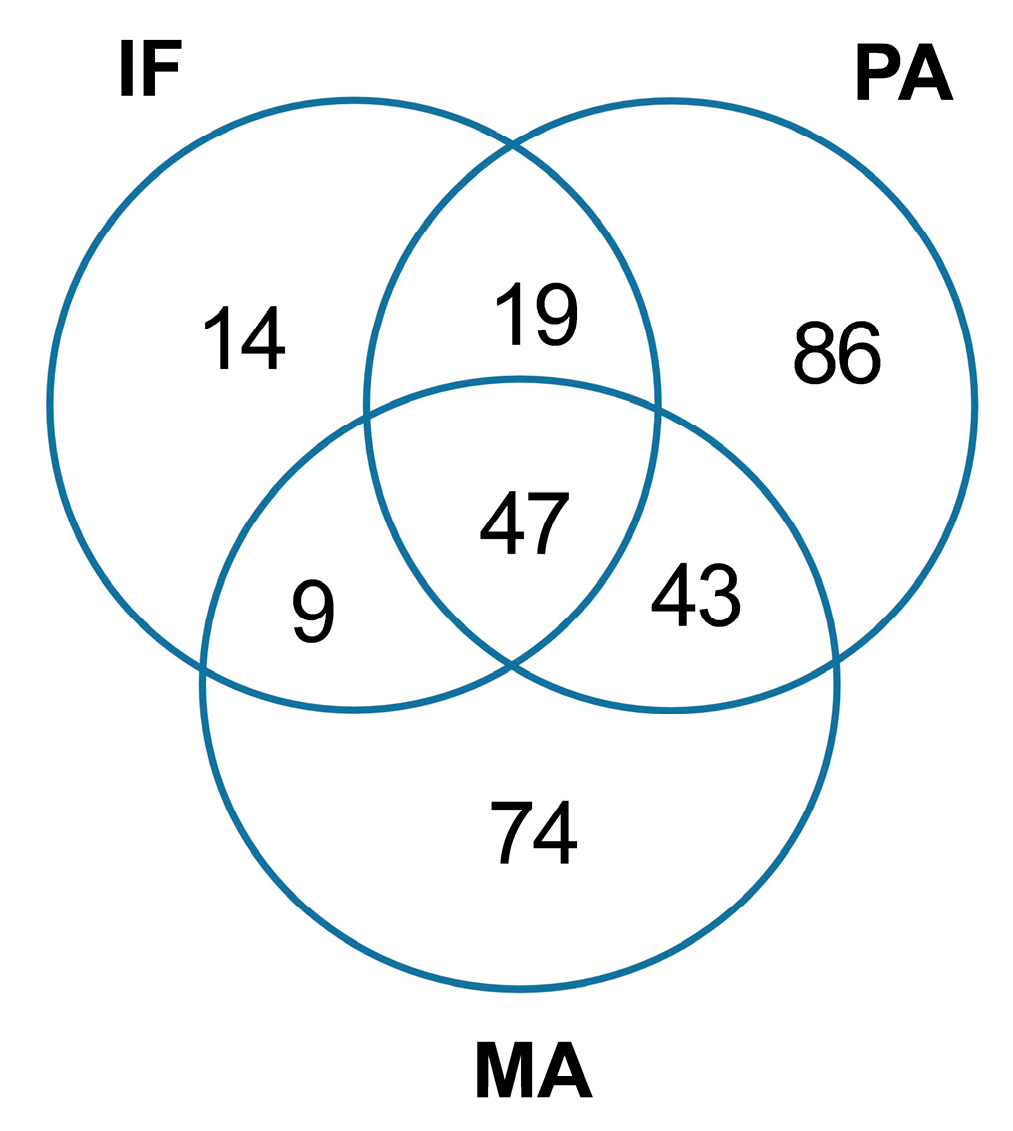
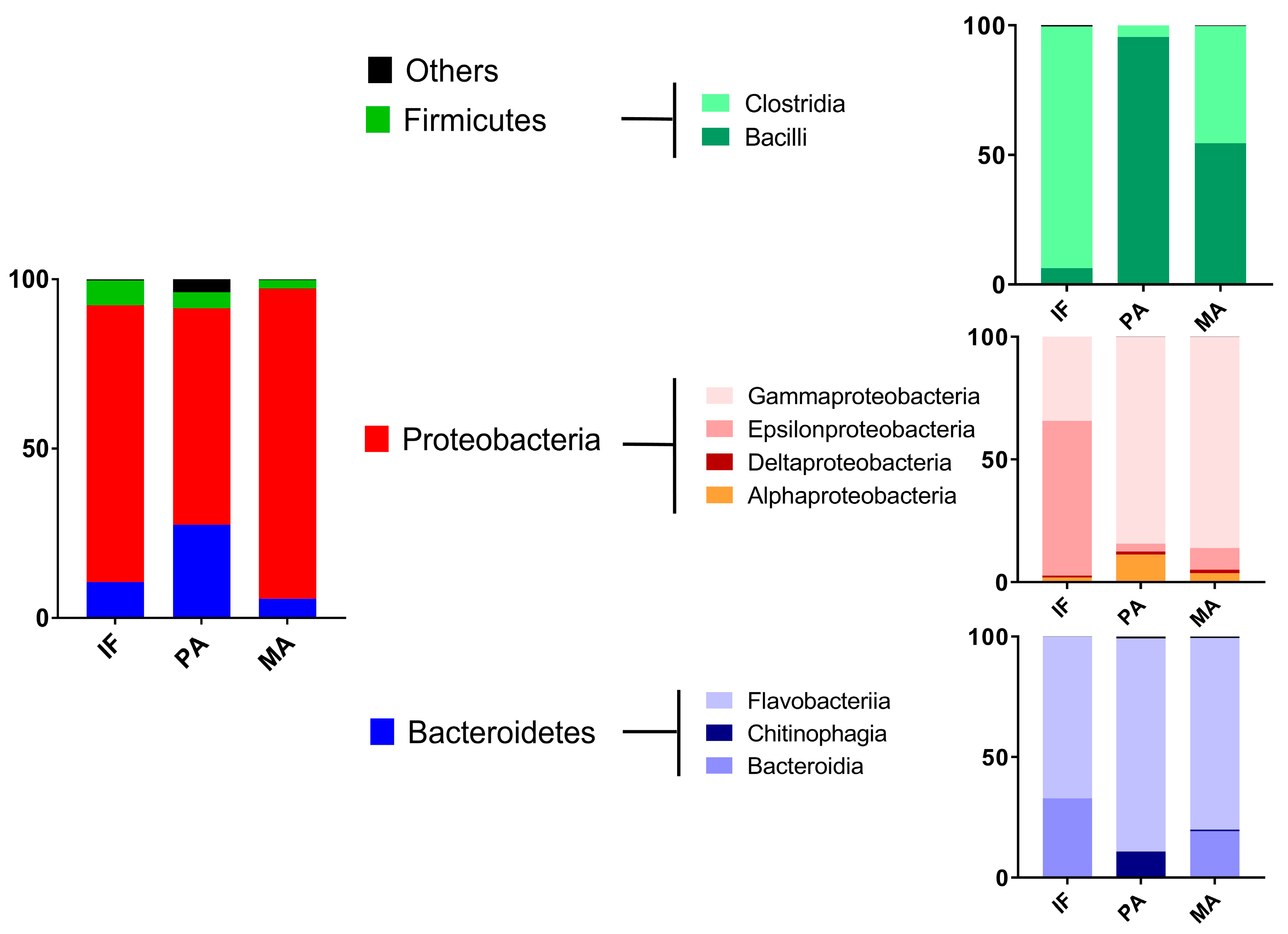
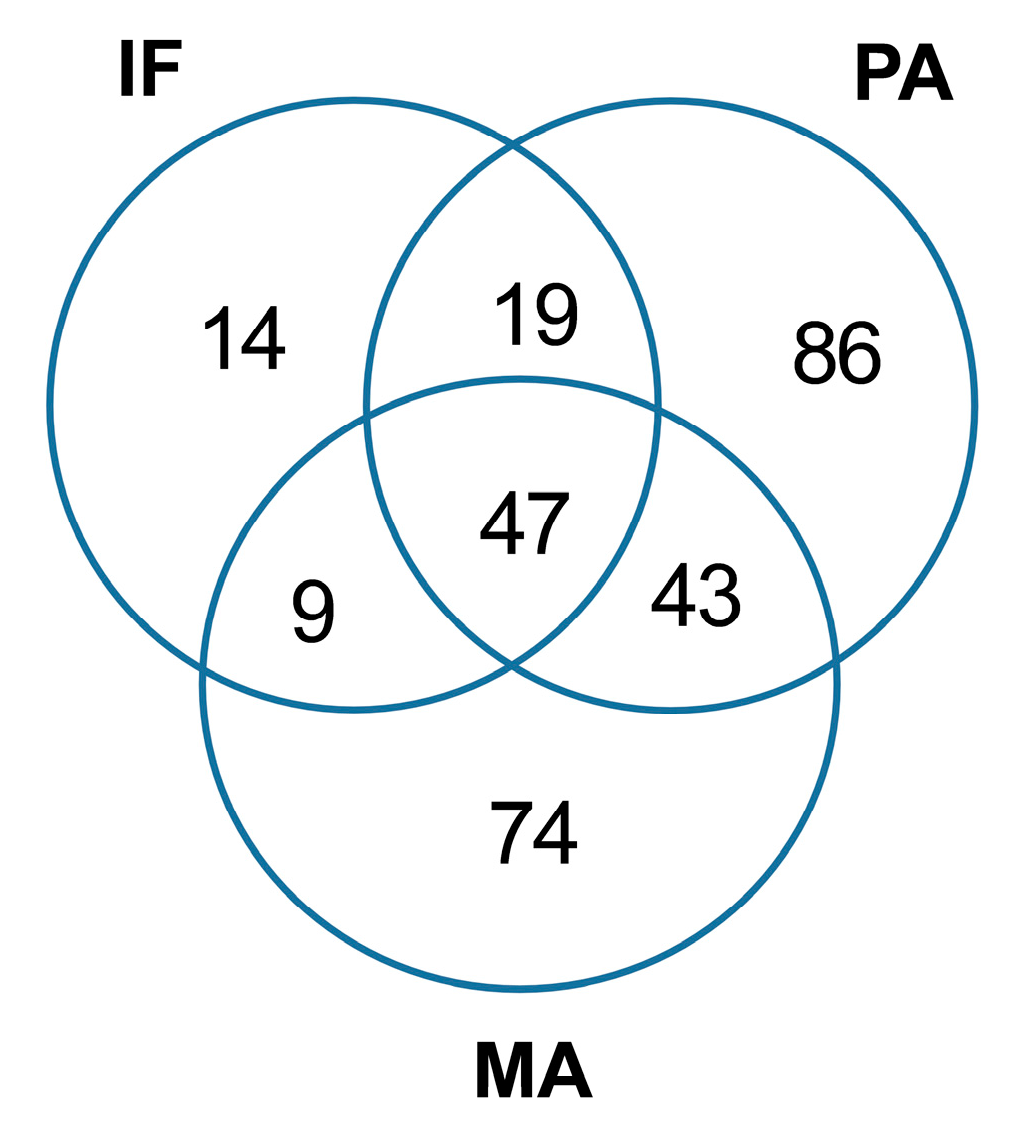
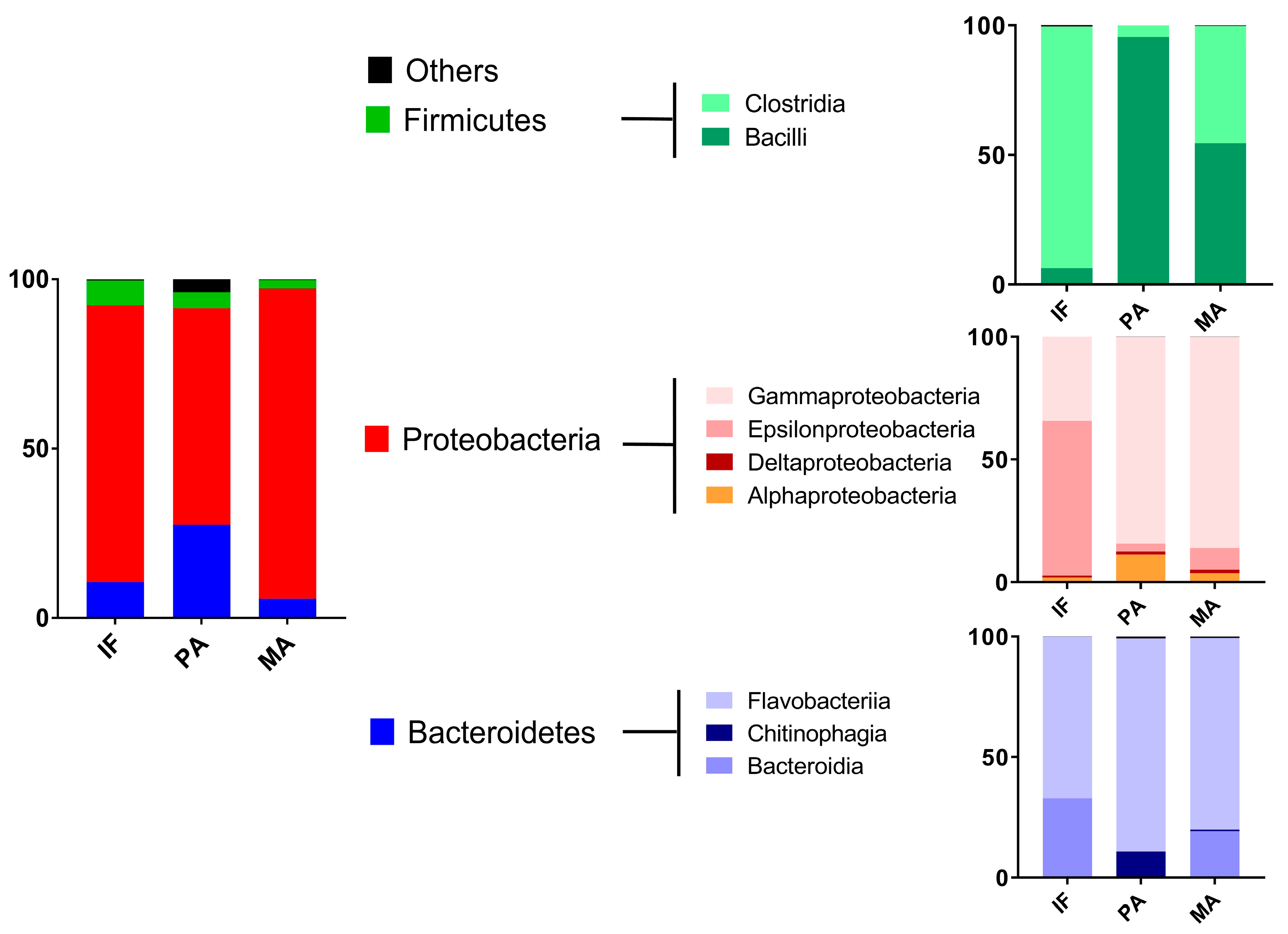
2.1. Coastal Microbiome
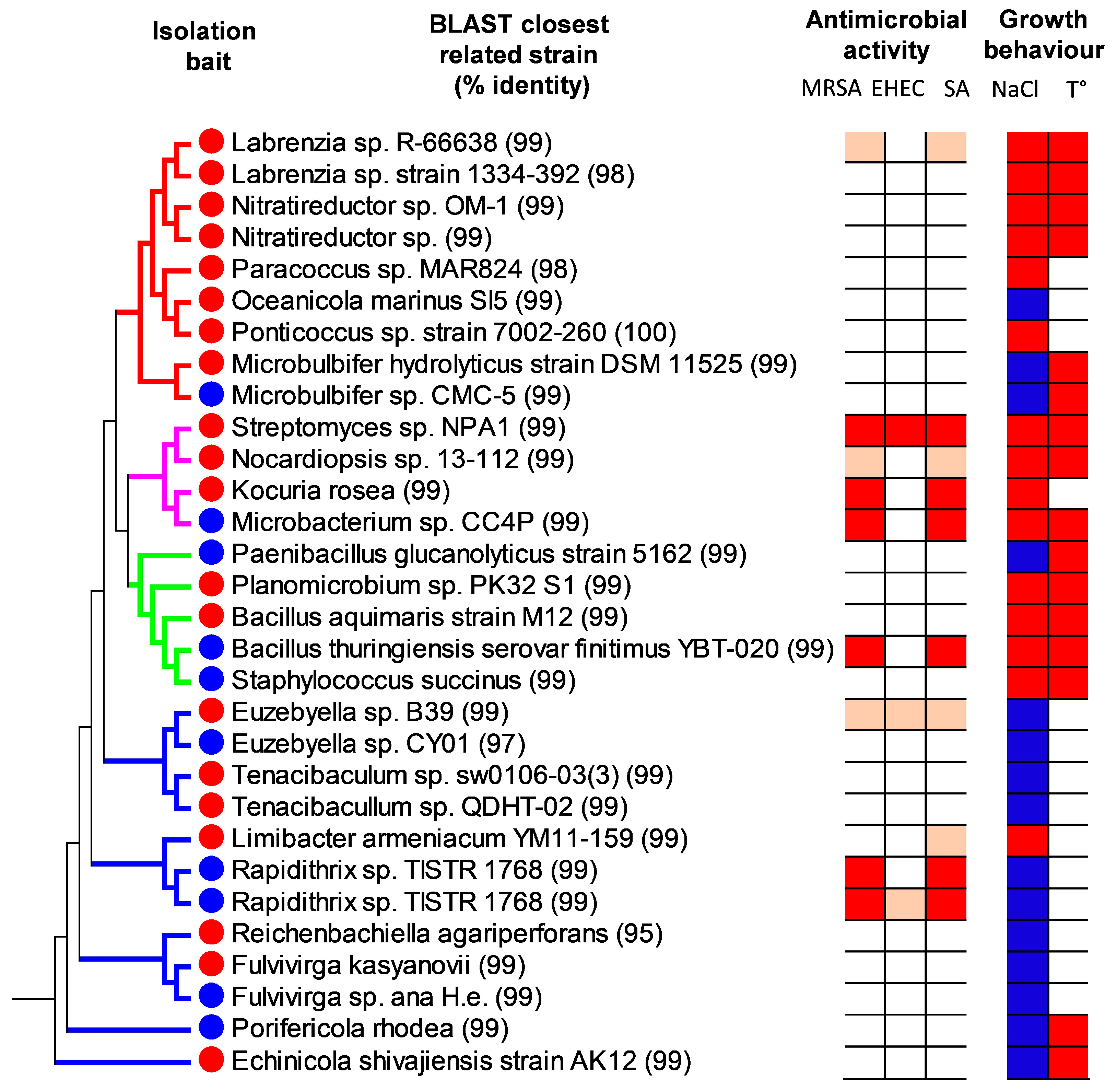
2.2. Isolation of Antibiotic Producing Predatory Bacteria
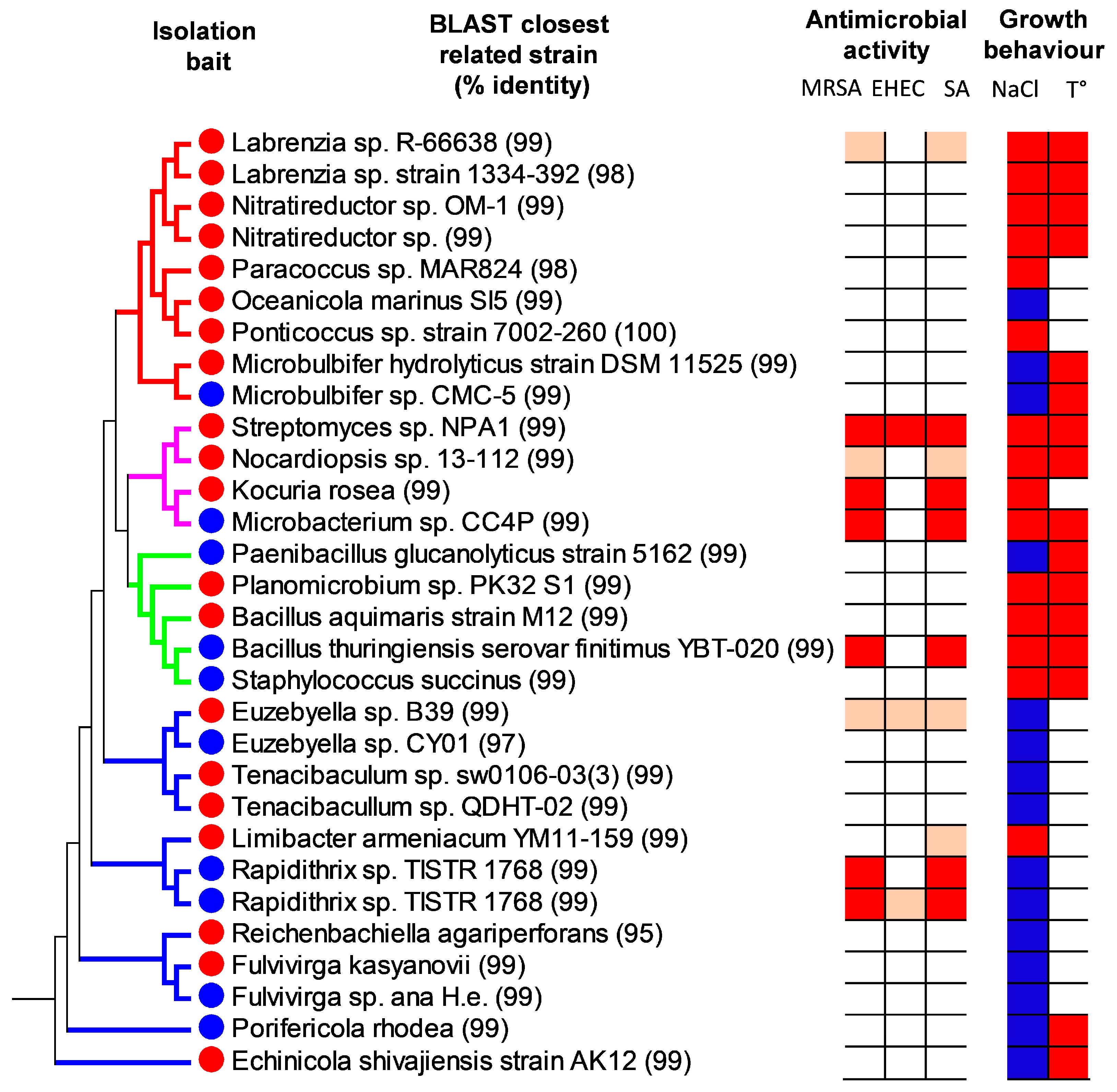
2.3. Phylogenetic Analysis
2.4. Physiological Properties of the Isolated Bacteria
2.5. Identification of Known Compounds—Dereplication
3. Discussion
4. Materials and Methods
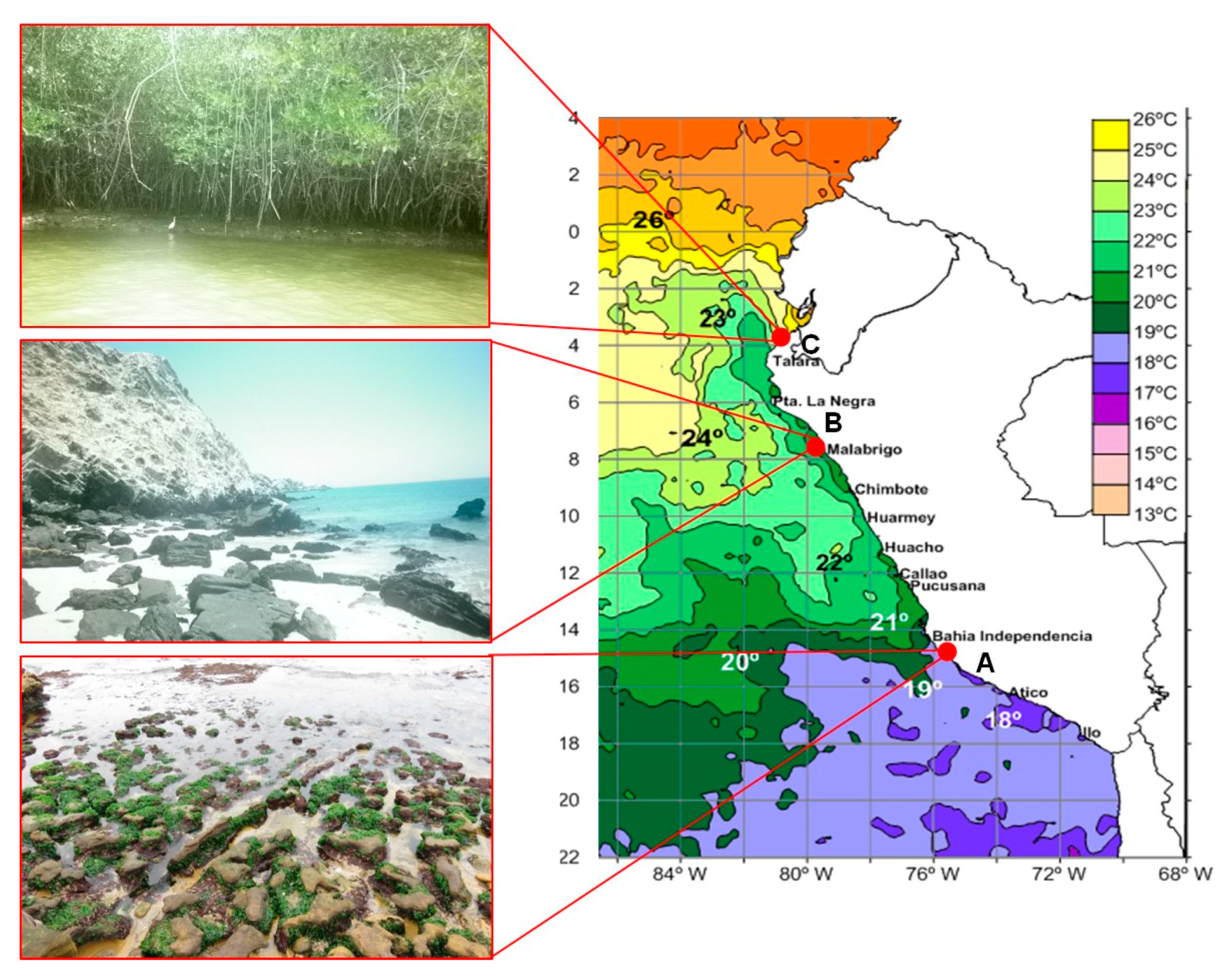
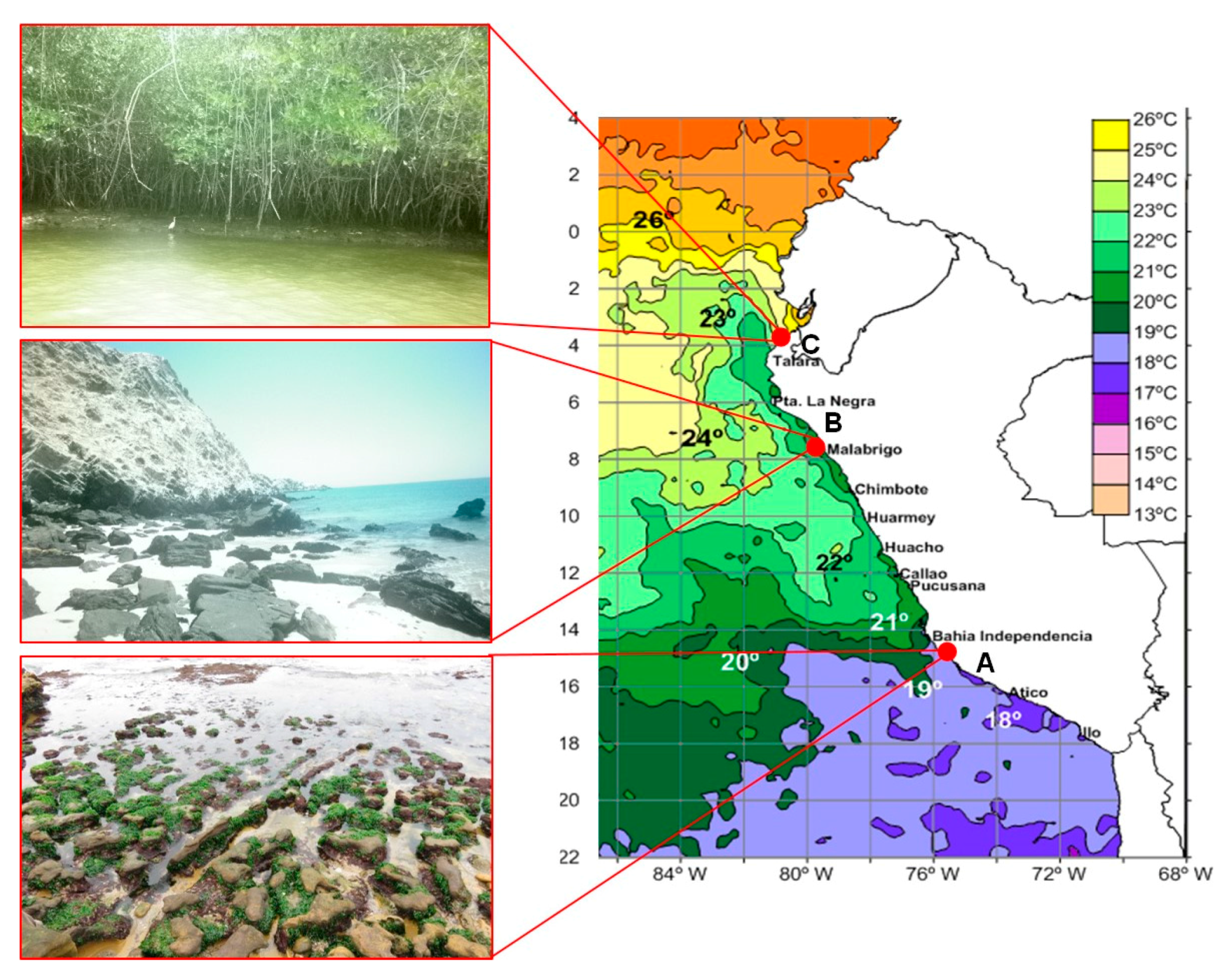
4.1. Collection of Samples and Isolation of Bacterial Strains
4.2. Metagenomic Analysis of 16S rRNA Gene Sequence
4.3. Antimicrobial Activity
4.4. Growth Characteristics
4.5. Phylogenetic Analysis of Isolated Strains
4.6. HPLC-MS/MS Measurements
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eom, S.-H.; Kim, Y.-M.; Kim, S.-K. Marine bacteria: Potential sources for compounds to overcome antibiotic resistance. Appl. Microbiol. Biotechnol. 2013, 97, 4763–4773. [Google Scholar] [CrossRef] [PubMed]
- Boehringer, N.; Fisch, K.M.; Schillo, D.; Bara, R.; Hertzer, C.; Grein, F.; Eisenbarth, J.-H.; Kaligis, F.; Schneider, T.; Waegele, H.; et al. Antimicrobial Potential of Bacteria Associated with Marine Sea Slugs from North Sulawesi, Indonesia. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Schaeberle, T.F.; Goralski, E.; Neu, E.; Erol, O.; Hoelzl, G.; Doermann, P.; Bierbaum, G.; Koenig, G.M. Marine Myxobacteria as a Source of Antibiotics-Comparison of Physiology, Polyketide-Type Genes and Antibiotic Production of Three New Isolates of Enhygromyxa salina. Mar. Drugs 2010, 8, 2466–2479. [Google Scholar] [CrossRef] [PubMed]
- Korp, J.; Vela Gurovic, M.S.; Nett, M. Antibiotics from predatory bacteria. Beilstein J. Org. Chem. 2016, 12, 594–607. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.J.; Zeph, L.R.; Casida, L.E., Jr. Bacterial control of Agromyces-ramosus in soil. Can. J. Microbiol. 1985, 31, 1157–1163. [Google Scholar] [CrossRef]
- Lewin, R.A. Saprospira grandis: A flexibacterium that can catch bacterial prey by “ixotrophy”. Microb. Ecol. 1997, 34, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, M.M.; Hoever, T.; Barra, L.; Kaiser, M.; Dorrestein, P.C.; Dickschat, J.S.; Schaeberle, T.F. Discovery of a Mosaic-Like Biosynthetic Assembly Line with a Decarboxylative Off-Loading Mechanism through a Combination of Genome Mining and Imaging. Angew. Chem. Int. Ed. 2016, 55, 13611–13614. [Google Scholar] [CrossRef] [PubMed]
- Sockett, R.E. Predatory Lifestyle of Bdellovibrio bacteriovorus. Annu. Rev. Microbiol. 2009, 63, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Imai, I.; Ishida, Y.; Hata, Y. Killing of arine-Phytoplankton by a gliding bacterium Cytophaga sp., isolated from the coastal sea of Japan. Mar. Biol. 1993, 116, 527–532. [Google Scholar] [CrossRef]
- Jurkevitch, E.; Jacquet, S. Bdellovibrio and related organisms Unusual bacterial predators! MS Med. Sci. 2017, 33, 519–527. [Google Scholar]
- Martin, M.O. Predatory prokaryotes: An emerging research opportunity. J. Mol. Microbiol. Biotechnol. 2002, 4, 467–477. [Google Scholar] [PubMed]
- Baumann, S.; Herrmann, J.; Raju, R.; Steinmetz, H.; Mohr, K.I.; Huettel, S.; Harmrolfs, K.; Stadler, M.; Mueller, R. Cystobactamids: Myxobacterial Topoisomerase Inhibitors Exhibiting Potent Antibacterial Activity. Angew. Chem. Int. Ed. 2014, 53, 14605–14609. [Google Scholar] [CrossRef] [PubMed]
- Schaeberle, T.F.; Lohr, F.; Schmitz, A.; Koenig, G.M. Antibiotics from myxobacteria. Nat. Prod. Rep. 2014, 31, 953–972. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.; Abou Fayad, A.; Muller, R. Natural products from myxobacteria: Novel metabolites and bioactivities. Nat. Prod. Rep. 2017, 34, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Seccareccia, I.; Kost, C.; Nett, M. Quantitative Analysis of Lysobacter Predation. Appl. Environ. Microbiol. 2015, 81, 7098–7105. [Google Scholar] [CrossRef] [PubMed]
- Srisukchayakul, P.; Suwanachart, C.; Sangnoi, Y.; Kanjana-Opas, A.; Hosoya, S.; Yokota, A.; Arunpairojana, V. Rapidithrix thailandica gen. nov., sp nov., a marine gliding bacterium isolated from samples collected from the Andaman sea, along the southern coastline of Thailand. Int. J. Syst. Evol. Microbiol. 2007, 57, 2275–2279. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Oku, N.; Park, S.; Kasai, H.; Yokota, A. Porifericola rhodea gen. nov., sp nov., a new member of the phylum Bacteroidetes isolated by the bait-streaked agar technique. Antonie Van Leeuwenhoek 2011, 100, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Miloslavich, P.; Klein, E.; Diaz, J.M.; Hernandez, C.E.; Bigatti, G.; Campos, L.; Artigas, F.; Castillo, J.; Penchaszadeh, P.E.; Neill, P.E.; et al. Marine Biodiversity in the Atlantic and Pacific Coasts of South America: Knowledge and Gaps. PLoS ONE 2011, 6, e14631. [Google Scholar] [CrossRef] [PubMed]
- Davila-Cespedes, A.; Hufendiek, P.; Cruesemann, M.; Schaeberle, T.F.; Koenig, G.M. Marine-derived myxobacteria of the suborder Nannocystineae: An underexplored source of structurally intriguing and biologically active metabolites. Beilstein J. Org. Chem. 2016, 12, 969–984. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Linares-Otoya, L.; Linares-Otoya, V.; Armas-Mantilla, L.; Blanco-Olano, C.; Crüsemann, M.; Ganoza-Yupanqui, M.L.; Campos-Florian, J.; König, G.M.; Schäberle, T.F. Identification and heterologous expression of the kocurin biosynthetic gene cluster. Microbiology 2017, 163, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; Los Santos, E.L.C.; de Kim, H.U.; Nave, M.; et al. AntiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.P.; Fay, J.P.; Dickens, K.A.; Parent, M.A.; Soroka, D.S.; Boyd, E.F. Predatory bacteria as natural modulators of Vibrio parahaemolyticus and Vibrio vulnificus in seawater and oysters. Appl. Environ. Microbiol. 2012, 78, 7455–7466. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.I.; Baumann, P.; Reichelt, J.L.; Allen, R.D. Isolation, enumeration, and host range of marine Bdellovibrios. Arch. Microbiol. 1974, 98, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Grote, J.; Jost, G.; Labrenz, M.; Herndl, G.J.; Juergens, K. Epsilonproteobacteria Represent the Major Portion of Chemoautotrophic Bacteria in Sulfidic Waters of Pelagic Redoxclines of the Baltic and Black Seas. Appl. Environ. Microbiol. 2008, 74, 7546–7551. [Google Scholar] [CrossRef] [PubMed]
- Kandel, P.P.; Pasternak, Z.; van Rijn, J.; Nahum, O.; Jurkevitch, E. Abundance, diversity and seasonal dynamics of predatory bacteria in aquaculture zero discharge systems. FEMS Microbiol. Ecol. 2014, 89, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Coates, R.C.; Clark, B.R.; Engene, N.; Gonzalez, D.; Esquenazi, E.; Dorrestein, P.C.; Gerwick, W.H. Biosynthetic origin of natural products isolated from marine microorganism-invertebrate assemblages. Proc. Natl. Acad. Sci. USA 2008, 105, 4587–4594. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; Fernández-López, M.; Muñoz-Adelantado, E.; Goris, J.; Vos, P.; de Martínez-Romero, E.; Toro, N.; Gillis, M. Description of new Ensifer strains from nodules and proposal to transfer Ensifer adhaerens Casida 1982 to Sinorhizobium as Sinorhizobium adhaerens comb. nov. Request for an opinion. Int. J. Syst. Evol. Microbiol. 2003, 53, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Banning, E.C.; Casciotti, K.L.; Kujawinski, E.B. Novel strains isolated from a coastal aquifer suggest a predatory role for flavobacteria. FEMS Microbiol. Ecol. 2010, 73, 254–270. [Google Scholar] [CrossRef] [PubMed]
- Mohr, K.I.; Stechling, M.; Wink, J.; Wilharm, E.; Stadler, M. Comparison of myxobacterial diversity and evaluation of isolation success in two niches: Kiritimati Island and German compost. Microbiol. Open 2016, 5, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaeberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef] [PubMed]
- De Lima Procopio, R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araujo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, G.; Thomas, B.; Parab, R.; Patel, Z.E.; Kuldharan, S.; Yemparala, V.; Mishra, P.D.; Ranadive, P.; D’Souza, L.; Pari, K.; et al. In vitro and in vivo activities of antibiotic PM181104. Antimicrob. Agents Chemother. 2013, 57, 5315–5319. [Google Scholar] [CrossRef] [PubMed]
- Lucena, T.; Pascual, J.; Giordano, A.; Gambacorta, A.; Garay, E.; Arahal, D.R.; Carmen Macian, M.; Pujalte, M.J. Euzebyella saccharophila gen. nov., sp nov., a marine bacterium of the family Flavobacteriaceae. Int. J. Syst. Evol. Microbiol. 2010, 60, 2871–2876. [Google Scholar] [CrossRef] [PubMed]
- Oku, N.; Adachi, K.; Matsuda, S.; Kasai, H.; Takatsuki, A.; Shizuri, Y. Ariakemicins a and b, novel polyketide-peptide antibiotics from a marine gliding bacterium of the genus Rapidithrix. Org. Lett. 2008, 10, 2481–2484. [Google Scholar] [CrossRef] [PubMed]
- Sangnoi, Y.; Plubrukarn, A.; Arunpairojana, V.; Kanjana-Opas, A. A new antibacterial amino phenyl pyrrolidone derivative from a novel marine gliding bacterium Rapidithrix thailandica. World J. Microbiol. Biotechnol. 2014, 30, 1135–1139. [Google Scholar] [CrossRef] [PubMed]
- Sangnoi, Y.; Sakulkeo, O.; Yuenyongsawad, S.; Kanjana-Opas, A.; Ingkaninan, K.; Plubrukarn, A.; Suwanborirux, K. Acetylcholinesterase-Inhibiting Activity of Pyrrole Derivatives from a Novel Marine Gliding Bacterium, Rapidithrix thailandica. Mar. Drugs 2008, 6, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Kunze, B.; Wray, V.; Reichenbach, H.; Jurkiewicz, E.; Hunsmann, G.; Hofle, G. Antibiotics from gliding bacteria 43. Phenoxan—A novel inhibitor of HIV-1 infection in cell-cultures from Polyangium-sp., strain PI-VO19 (Myxobacteria). Eur. J. Org. Chem. 1991, 1991, 707–708. [Google Scholar]
- Schaeberle, T.F.; Hack, I.M. Overcoming the current deadlock in antibiotic research. Trends Microbiol. 2014, 22, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Site of Collection (N° of Reads) a | ||
|---|---|---|---|
| Isla Foca (213,991) | Paracas (161,958) | Manglares (163,056) | |
| Bacteroidetes | |||
| Cytophagales | 6 | 203 | 11 |
| Cellulophaga | 2 | 0 | 0 |
| Tenacibaculum | 4 | 131 | 179 |
| Saprospiraceae | 2 | 334 | 0 |
| Delta-proteobacteria | |||
| Bdellovibrionales-like bacteria | 2 | 81 | 1184 |
| Bacteriovoracaceae | 1 | 324 | 2 |
| Bdellovibrionaceae | 2 | 3 | 0 |
| Myxococcales | 0 | 1 | 0 |
| Cyanobacteria | |||
| Vampirovibrio | 0 | 0 | 2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linares-Otoya, L.; Linares-Otoya, V.; Armas-Mantilla, L.; Blanco-Olano, C.; Crüsemann, M.; Ganoza-Yupanqui, M.L.; Campos-Florian, J.; König, G.M.; Schäberle, T.F. Diversity and Antimicrobial Potential of Predatory Bacteria from the Peruvian Coastline. Mar. Drugs 2017, 15, 308. https://doi.org/10.3390/md15100308
Linares-Otoya L, Linares-Otoya V, Armas-Mantilla L, Blanco-Olano C, Crüsemann M, Ganoza-Yupanqui ML, Campos-Florian J, König GM, Schäberle TF. Diversity and Antimicrobial Potential of Predatory Bacteria from the Peruvian Coastline. Marine Drugs. 2017; 15(10):308. https://doi.org/10.3390/md15100308
Chicago/Turabian StyleLinares-Otoya, Luis, Virginia Linares-Otoya, Lizbeth Armas-Mantilla, Cyntia Blanco-Olano, Max Crüsemann, Mayar L. Ganoza-Yupanqui, Julio Campos-Florian, Gabriele M. König, and Till F. Schäberle. 2017. "Diversity and Antimicrobial Potential of Predatory Bacteria from the Peruvian Coastline" Marine Drugs 15, no. 10: 308. https://doi.org/10.3390/md15100308
APA StyleLinares-Otoya, L., Linares-Otoya, V., Armas-Mantilla, L., Blanco-Olano, C., Crüsemann, M., Ganoza-Yupanqui, M. L., Campos-Florian, J., König, G. M., & Schäberle, T. F. (2017). Diversity and Antimicrobial Potential of Predatory Bacteria from the Peruvian Coastline. Marine Drugs, 15(10), 308. https://doi.org/10.3390/md15100308