
Marine Organisms with Anti-Diabetes Properties



Abstract
:1. Introduction
2. Common Targets for Anti-Diabetes Assays
3. Marine Microorganisms with Anti-Diabetes Properties
4. Marine Macroorganisms with Anti-Diabetes Properties
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014, 943162. [Google Scholar] [CrossRef] [PubMed]
- Barde, S.R.; Sakhare, R.S.; Kanthale, S.B.; Chandak, P.G.; Jamkhande, P.G. Marine bioactive agents: A short review on new marine antidiabetic compounds. Asian Pac. J. Trop. Dis. 2015, 5, S209–S213. [Google Scholar] [CrossRef]
- Mathers, C.D.; Loncar, D. Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med. 2006, 3, e442. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, Z.; Guo, Q.; Wang, T.; Lu, C.; Chen, Y.; Sheng, Q.; Chen, J.; Nie, Z.; Zhang, Y.; et al. Anti-Diabetic Effects of CTB-APSL Fusion Protein in Type 2 Diabetic Mice. Mar. Drugs 2014, 12, 1512–1529. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.R.; Parikh, R.M.; Das, A.K. Insulin—History, biochemistry, physiology and pharmacology. J. Assoc. Physicians India 2007, 55, S19–S25. [Google Scholar]
- Newsholme, P.; Cruzat, V.; Arfuso, F.; Keane, K. Nutrient regulation of insulin secretion and action. J. Endocrinol. 2014, 221, R105–R120. [Google Scholar] [CrossRef] [PubMed]
- Sonksen, P.; Sonksen, J. Insulin: Understanding its action in health and disease. Br. J. Anaesth. 2000, 85, 69–79. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; De Girolamo, M.; Cusenza, S.; Riccioni, G. Marine bioactives: Pharmacological properties and potential applications against inflammatory diseases. Mar. Drugs 2012, 10, 812–833. [Google Scholar] [CrossRef] [PubMed]
- Rewers, M.; Ludvigsson, J. Environmental risk factors for type 1 diabetes. Lancet 2016, 387, 2340–2348. [Google Scholar] [CrossRef]
- Reimann, M.; Bonifaci, E.; Solimena, M.; Schwarz, P.E.; Ludwig, B.; Hanefeld, M.; Bornstein, S.R. An update on preventive and regenerative therapies in diabetes mellitus. Pharmacol. Ther. 2009, 121, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Popov, A.M.; Krivoshapko, O.N. Protective effects of polar lipids and redox-active compounds from marine organisms at modeling of hyperlipidemia and diabetes. J. Biomed. Sci. Eng. 2013, 6, 543–550. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, M.; Wang, J.; Ding, Y.; Dai, X.; Li, Y. Oral administration of skin gelatin isolated from chum salmon (Oncorhynchus keta) enhances wound healing in diabetic rats. Mar. Drugs 2011, 9, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.F.; Peng, H.B.; Liu, G.Q.; Zhang, F.; Li, Y. Beneficial effects of oligopeptides from marine salmon skin in a rat model of type 2 diabetes. Nutrition 2010, 26, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. Ectyoceramide, the First Natural Hexofuranosylceramide from the Marine Sponge Ectyoplasia ferox. Eur. J. Org. Chem. 2003, 1433–1437. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. Glycolipids from sponges. 20. J-coupling analysis for stereochemical assignments in furanosides: Structure elucidation of vesparioside B, a glycosphingolipid from the marine sponge Spheciospongia vesparia. J. Org. Chem. 2008, 73, 6158–6165. [Google Scholar] [PubMed]
- Jaspars, M.; DePascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The marine biodiscovery pipeline and ocean medicines of tomorrow. J. Mar. Biol. Assoc. U. K. 2016, 96, 151–158. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Löwenberg, B. Sense and nonsense of high-dose cytarabine for acute myeloid leukemia. Blood 2013, 121, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Schoffski, P.; Dumez, H.; Wolter, P.; Stefan, C.; Wozniak, A.; Jimeno, J.; Van Oosterom, A.T. Clinical impact of trabectedin (ecteinascidin-743) in advanced/metastatic soft tissue sarcoma. Expert Opin. Pharmacother. 2008, 9, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Menis, J.; Twelves, C. Eribulin (Halaven): A new, effective treatment for women with heavily pretreated metastatic breast cancer. Breast Cancer Targets Ther. 2011, 3, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Klotz, U. Ziconotide—A novel neuron-specific calcium channel blocker for the intrathecal treatment of severe chronic pain—A short review. Int. J. Clin. Pharmacol. Ther. 2006, 44, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.; Kaur, M.; Minneman, K.P. Antiviral lead compounds from marine sponges. Mar. Drugs 2010, 8, 2619–2638. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the ocean for new developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Bidon-Chanal, A.; Fuertes, A.; Alonso, D.; Perez, D.I.; Martinez, A.; Luque, F.J.; Medina, M. Evidence for a new binding mode to GSK-3: Allosteric regulation by the marine compound palinurin. Eur. J. Med. Chem. 2013, 60, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; FitzGerald, R.J. In vitro assessment of the cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Imada, G. Enzyme inhibitors and other bioactive compounds from marine actinomycetes. Antonie Van Leeuwenhoek 2005, 87, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Nam, K.A.; Kurihara, H.; Kim, S.M. Potent α-glucosidase inhibitors purified from the red algae Grateloupia elliptica. Phytochemistry 2008, 69, 2820–2825. [Google Scholar] [CrossRef] [PubMed]
- Krish, S.; Das, A. In-vitro bioactivity of marine seaweed, Cladophora rupestris. Int. J. Pharm. Biol. Sci. 2014, 5, 898–908. [Google Scholar]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Pandey, S.; Sree, A.; Dash, S.S.; Sethi, D.P.; Chowdhury, L. Diversity of marine bacteria producing beta-glucosidase inhibitors. Microb. Cell Fact. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Chen, F. Evaluation of the Green Alga Chlorella pyrenoidosa for management of diabetes. J. Food Drug Anal. 2012, 20, 246–249. [Google Scholar]
- Lebovitz, H.E. Oral antidiabetic agents: The emergence of α-glucosidase inhibitors. Drugs 1992, 44, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Suzen, S.; Buyukbingol, E. Recent studies of aldose reductase enzyme inhibition for diabetic complications. Curr. Med. Chem. 2003, 10, 1329–1352. [Google Scholar] [CrossRef] [PubMed]
- Lankas, G.R.; Leiting, B.; Roy, R.S.; Eiermann, G.J.; Beconi, M.G.; Biftu, T.; Chan, C.C.; Edmondson, S.; Feeney, W.P.; He, H.; et al. Dipeptidyl peptidase IV inhibition for the treatment of type 2 diabetes: potential importance of selectivity over dipeptidyl peptidases 8 and 9. Diabetes 2005, 54, 2988–2994. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, E.J.; Dokken, B.B. Role of glycogen synthase kinase-3 in insulin resistance and type 2 diabetes. Curr. Drug Targets 2006, 7, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Sharifuddin, Y.; Chin, Y.X.; Lim, P.E.; Phang, S.M. Potential bioactive compounds from seaweed for diabetes management. Mar. Drugs 2015, 13, 5447–5491. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Guo, S.; Jiang, B.; Guo, C.; Wang, T.; Zhang, L.; Li, J. HPN, a synthetic analogue of bromophenol from red alga Rhodomela confervoides: Synthesis and anti-diabetic effects in C57BL/KsJ-db/db mice. Mar. Drugs 2013, 11, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Nakazawa, T.; Sumilat, D.A.; Takahashi, O.; Ukai, K.; Takahashi, S.; Namikoshi, M. Euryspongins A–C, three new unique sesquiterpenes from a marine sponge Euryspongia sp. Bioorg. Med. Chem. Lett. 2013, 23, 2151–2154. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Jin, Y.B.; Lee, H.; Cha, M.; Sohn, E.-T.; Moon, J.; Park, C.; Chun, S.; Jung, E.-S.; Hong, J.-S.; et al. Brown alga Ecklonia cava attenuates type 1 diabetes by activating ampk and akt signaling pathways. Food Chem. Toxicol. 2010, 48, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Nuño, K.; Villarruel-López, A.; Puebla-Pérez, A.M.; Romero-Velarde, E.; Puebla-Mora, A.G.; Ascencio, F. Effects of the marine microalgae Isochrysis galbana and Nannochloropsis oculata in diabetic rats. J. Funct. Foods 2013, 106–115. [Google Scholar] [CrossRef]
- Tamrakar, A.K.; Tiwari, P.; Ahmad, R.; Kumar, R.; Lakshmi, V.; Srivastava, M.N.; Srivastava, A.K. Antihyperglycaemic activity of Sinularia firma and Sinularia erecta in streptozotocin-induced diabetic rats. Med. Chem. Res. 2008, 17, 62–73. [Google Scholar] [CrossRef]
- Tiwari, P.; Rahuja, N.; Kumar, R.; Lakshmi, V.; Srivastava, M.N.; Agarwal, S.C.; Raghubir, R.; Srivastava, A.K. Search for antihyperglycemic activity in few marine flora and fauna. Indian J. Sci. Technol. 2008, 1, 1–5. [Google Scholar]
- Sun, Z.; Peng, X.; Liu, J.; Fan, K.-W.; Wang, M.; Chen, F. Inhibitory effects of microalgal extracts on the formation of advanced glycation endproducts (AGEs). Food Chem. 2010, 120, 261–267. [Google Scholar] [CrossRef]
- Zorena, K.; Raczyńska, D.; Raczyńska, K. Biomarkers in diabetic retinopathy and the therapeutic implications. Mediat. Inflamm. 2013, 2013, 193604. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, J.; Zeng, X.; Huangfu, J.; Jiang, Y.; Wang, M.; Chen, F. Protective actions of microalgae against endogenous and exogenous advanced glycation endproducts (AGEs) in human retinal pigment epithelial cells. Food Funct. 2011, 2, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Cannell, R.J.; Kellam, S.J.; Owsianka, A.M.; Walker, J.M. Microalgae and cyanobacteria as a source of glycosidase inhibitors. J. Gen. Microbiol. 1987, 133, 1701–1705. [Google Scholar] [CrossRef] [PubMed]
- Debbab, A.; Aly, A.H.; Lin, W.H.; Proksch, P. Bioactive compounds from marine bacteria and fungi. Microb. Biotechnol. 2010, 3, 544–563. [Google Scholar] [CrossRef] [PubMed]
- Ianora, A.; Miralto, A.; Poulet, S.A.; Carotenuto, Y.; Buttino, I.; Romano, G.; Casotti, R.; Pohnert, G.; Wichard, T.; Colucci-D’Amato, L.; et al. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, Y.; Esposito, F.; Pisano, F.; Lauritano, C.; Perna, M.; Miralto, A.; Ianora, A. Multigeneration cultivation of the copepod Calanus helgolandicus in a re-circulating system. J. Exp. Mar. Biol. Ecol. 2012, 418, 46–58. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, J.; Zeng, X.; Huangfu, J.; Jiang, Y.; Wang, M.; Chen, F. Astaxanthin is responsible for antiglycoxidative properties of microalga Chlorella zofingiensis. Food Chem. 2011, 126, 1629–1635. [Google Scholar] [CrossRef] [PubMed]
- Miki, W. Biological functions and activities of animal carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes and antibacterial activities. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and temperature effects on bioactivity in diatoms. J. Appl. Phycol. 2015, 28, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, X.; Gao, L.; Cui, C.; Li, C.; Li, J.; Wang, B. Extraction and PTP1B inhibitory activity of bromophenols from the marine red alga Symphyocladia latiuscula. Chin. J. Oceanol. Limnol. 2011, 29, 686–690. [Google Scholar] [CrossRef]
- Shi, D.; Li, J.; Jiang, B.; Guo, S.; Su, H.; Wang, T. Bromophenols as inhibitors of protein tyrosine phosphatase 1B with antidiabetic properties. Bioorg. Med. Chem. Lett. 2012, 22, 2827–2832. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Okada, Y.; Shi, H.; Wang, Y.; Okuyama, T. Structures and aldose reductase inhibitory effects of bromophenols from the red alga Symphyocladia latiuscula. J. Nat. Prod. 2005, 68, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Shin, K.H.; Kim, B.K.; Lee, S. Anti-diabetic activities of fucosterol from Pelvetia siliquosa. Arch. Pharm. Res. 2004, 27, 1120–1122. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K. Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK-A(y) mice. Plant Foods Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Yoon, N.Y.; Woo, M.H.; Choi, J.S. Inhibitory activities of extracts from several kinds of seaweeds and phlorotannins from the brown alga Ecklonia stolonifera on glucose-mediated protein damage and rat lens aldose reductase. Fish. Sci. 2008, 74, 1363–1365. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Ishimaru, A.; Suzuki, R.; Okuyama, T. A new phloroglucinol derivative from the brown alga Eisenia bicyclis: Potential for the effective treatment of diabetic complications. J. Nat. Prod. 2004, 67, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Han, J.S.; Kim, H.J.; Jeon, Y.J. Diphlorethohydroxycarmalol isolated from Ishige okamurae, a brown algae, a potent alpha-glucosidase and alpha-amylase inhibitor, alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2009, 615, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Celikler, S.; Tas, S.; Vatan, O.; Ziyanok-Ayvalik, S.; Yildiz, G.; Bilaloglu, R. Anti-hyperglycemic and antigenotoxic potential of Ulva rigida ethanolic extract in the experimental diabetes mellitus. Food Chem. Toxicol. 2009, 47, 1837–1840. [Google Scholar] [CrossRef] [PubMed]
- Gokce, G.; Haznedaroglu, M.Z. Evaluation of antidiabetic, antioxidant and vasoprotective effects of Posidonia oceanica extract. J. Ethnopharmacol. 2008, 115, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, Y.A.; Lee, J.I.; Lee, B.J.; Seo, Y.W. Screening of Korean marine plants extracts for inhibitory activity on protein tyrosine phosphatase 1B. J. Appl. Biol. Chem. 2007, 50, 74–77. [Google Scholar]
- Kimura, Y.; Watanabe, K.; Okuda, H. Effects of soluble sodium alginate on cholesterol excretion and glucose tolerance in rats. J. Ethnopharmacol. 1996, 54, 47–54. [Google Scholar] [CrossRef]
- Esposito, G.; Teta, R.; Miceli, R.; Ceccarelli, L.S.; Della Sala, G.; Camerlingo, R.; Irollo, E.; Mangoni, A.; Pirozzi, G.; Costantino, V. Isolation and assessment of the in vitro anti-tumor activity of smenothiazole A and B, chlorinated thiazole-containing peptide/polyketides from the Caribbean sponge, Smenospongia aurea. Mar. Drugs 2015, 13, 444–459. [Google Scholar] [CrossRef] [PubMed]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine Sponge Derived Natural Products between 2001 and 2010: Trends and Opportunities for Discovery of Bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [PubMed]
- Lamoral-Theys, D.; Fattorusso, E.; Mangoni, A.; Perinu, C.; Kiss, R.; Costantino, V. Evaluation of the antiproliferative activity of diterpene isonitriles from the sponge Pseudoaxinella flava in apoptosis-sensitive and apoptosis-resistant cancer cell lines. J. Nat. Prod. 2011, 74, 2299–2303. [Google Scholar] [CrossRef] [PubMed]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A.; Freigang, S.; Teyton, L. Corrugoside, a new immunostimulatory alpha-galactoglycosphingolipid from the marine sponge Axinella corrugata. Bioorg. Med. Chem. 2008, 16, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Khanfar, M.A.; Asal, B.A.; Mudit, M.; Kaddoumi, A.; El Sayed, K.A. The marine natural-derived inhibitors of glycogen synthase kinase-3β phenylmethylene hydantoins: In vitro and in vivo activities and pharmacophore modeling. Bioorg. Med. Chem. 2009, 17, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Uehara, T.; Matunaga, S.; Fusetani, N.; van Soest, R.W.M. Callyspongynic acid, a polyacetylenic acid which inhibits α-glucosidase, from the marine sponge Callyspongia truncata. J. Nat. Prod. 2002, 65, 922–924. [Google Scholar] [CrossRef] [PubMed]
- Van Kaer, L. Drugs from the Sea: A marine sponge-derived compound prevents Type 1 diabetes. Sci. World J. 2001, 1, 630–632. [Google Scholar] [CrossRef] [PubMed]
- Pascual, I.; Lopéz, A.; Gómez, H.; Chappé, M.; Saroyán, A.; González, Y.; Cisneros, M.; Charli, J.L.; Chávez, M. Screening of inhibitors of porcine dipeptidyl peptidase iv activity in aqueous extracts from marine organisms. Enzyme Microb. Technol. 2007, 40, 414–419. [Google Scholar] [CrossRef]
- Yamazaki, H.; Sumilat, D.A.; Kanno, S.; Ukai, K.; Rotinsulu, H.; Wewengkang, D.S.; Ishikawa, M.; Mangindaan, R.E.; Namikoshi, M. A polybromodiphenyl ether from an Indonesian marine sponge Lamellodysidea herbacea and its chemical derivatives inhibit protein tyrosine phosphatase 1B, an important target for diabetes treatment. J. Nat. Med. 2013, 67, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Guo, Y.W.; Jiang, H.L.; Shen, X. A sesquiterpene quinone, dysidine, from the sponge Dysidea villosa, activates the insulin pathway through inhibition of PTPases. Acta Pharm. Sin. 2009, 30, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Giannini, C.; Debitus, C.; Lucas, R.; Ubeda, A.; Payá, M.; Hooper, J.N.; D’Auria, M.V. New sesquiterpene derivatives from the sponge Dysidea species with a selective inhibitor profile against human phospholipase A2 and other leukocyte functions. J. Nat. Prod. 2001, 64, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Shen, X.; Guo, Y.W. A novel sesquiterpene quinone from Hainan sponge Dysidea villosa. Bioorg. Med. Chem. Lett. 2009, 19, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.F.; Li, G.-Z.; Peng, H.B.; Zhang, F.; Chen, Y.; Li, Y. Effect of marine collagen peptides on markers of metabolic nuclear receptors in type 2 diabetic patients with/without hypertension. Biomed. Environ. Sci. 2010, 23, 113–120. [Google Scholar] [CrossRef]
- Taouis, M.; Dagou, C.; Ster, C.; Durand, G.; Pinault, M.; Delarue, J. N-3 polyunsaturated fatty acids prevent the defect of insulin receptor signaling in muscle. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E664–E671. [Google Scholar] [CrossRef] [PubMed]
- Delarue, J.; Couet, C.; Cohen, R.; Bréchot, J.F.; Antoine, J.M.; Lamisse, F. Effects of fish oil on metabolic responses to oral fructose and glucose loads in healthy humans. Am. J. Physiol. 1996, 270, E353–E362. [Google Scholar] [PubMed]
- Lauritano, C.; Procaccini, G.; Ianora, A. Gene Expression Patterns and Stress Response in Marine Copepods. Mar. Environ. Res. 2012, 76, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Mendis, E. Bioactive compounds from marine processing byproducts—A review. Int. Food Res. 2006, 39, 383–393. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive peptides from marine processing waste and shellfish: A review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Berry, E.M. Dietary fatty acids in the management of diabetes mellitus. Am. J. Clin. Nutr. 1997, 66, 991–997. [Google Scholar]
- Sheehan, J.P.; Wei, I.W.; Ulchaker, M.; Tserng, K.Y. Effect of high fiber intake in fish oil-treated patients with non-insulin-dependent diabetes mellitus. Am. J. Clin. Nutr. 1997, 66, 1183–1187. [Google Scholar] [PubMed]
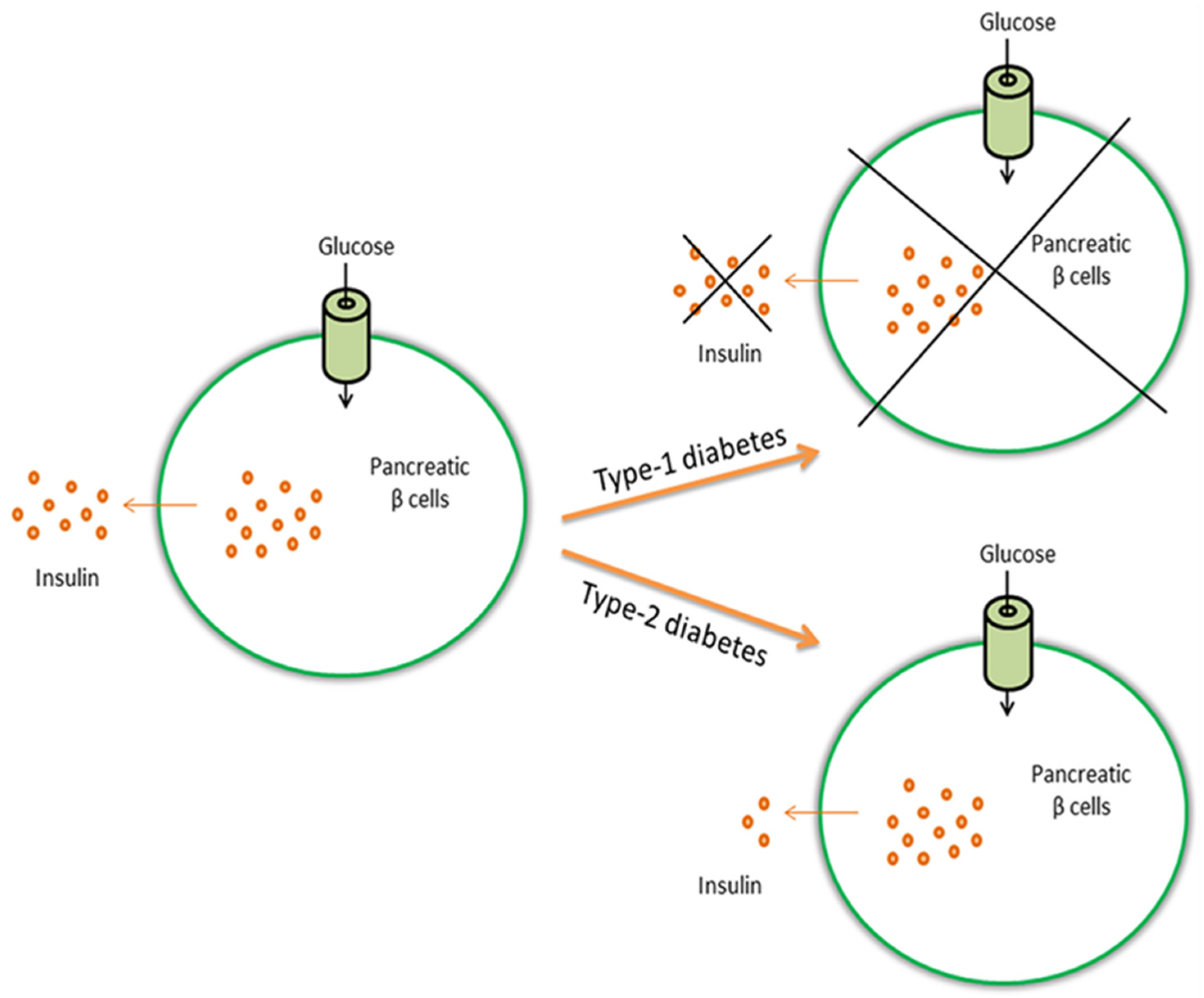

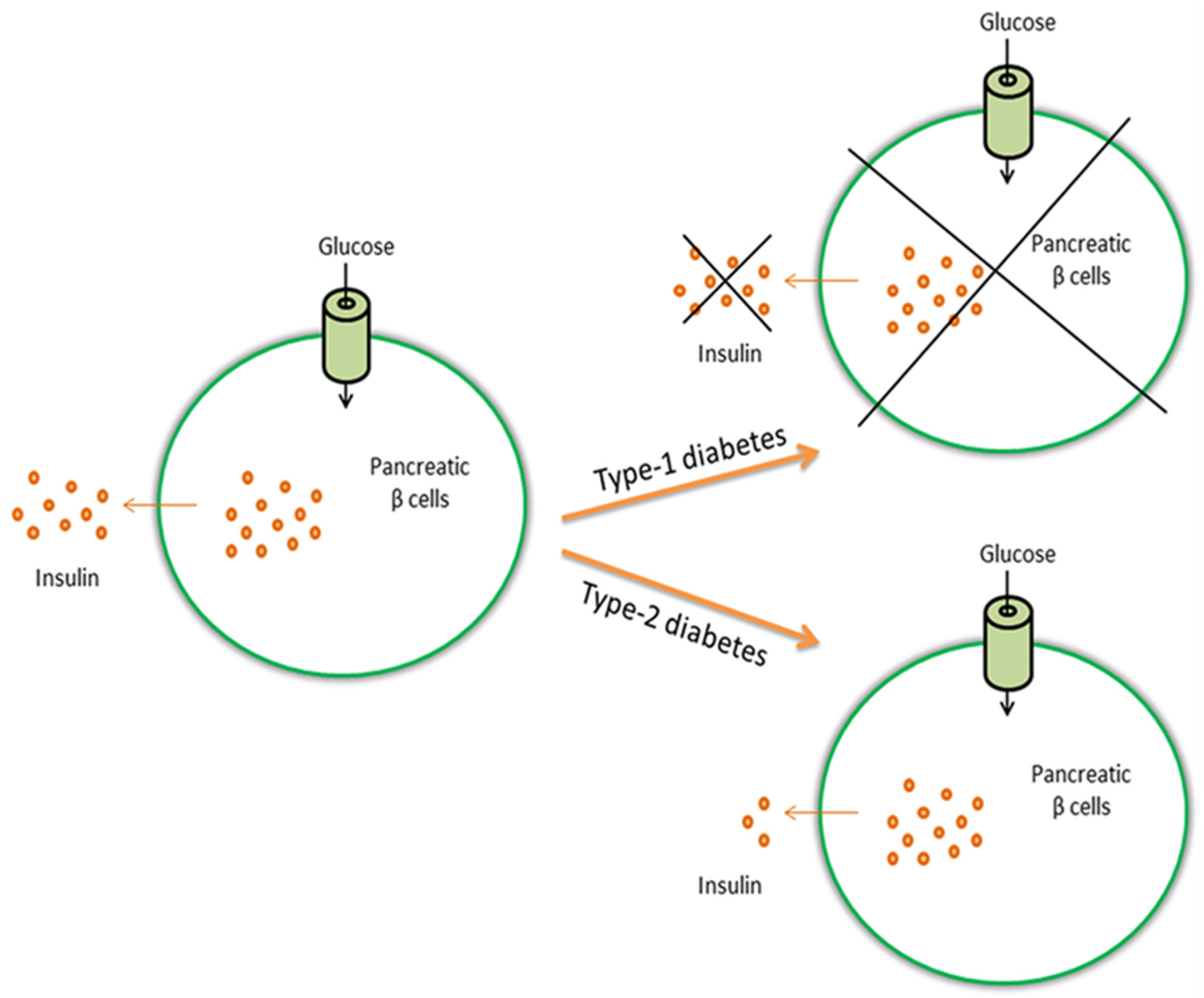{kind=link}
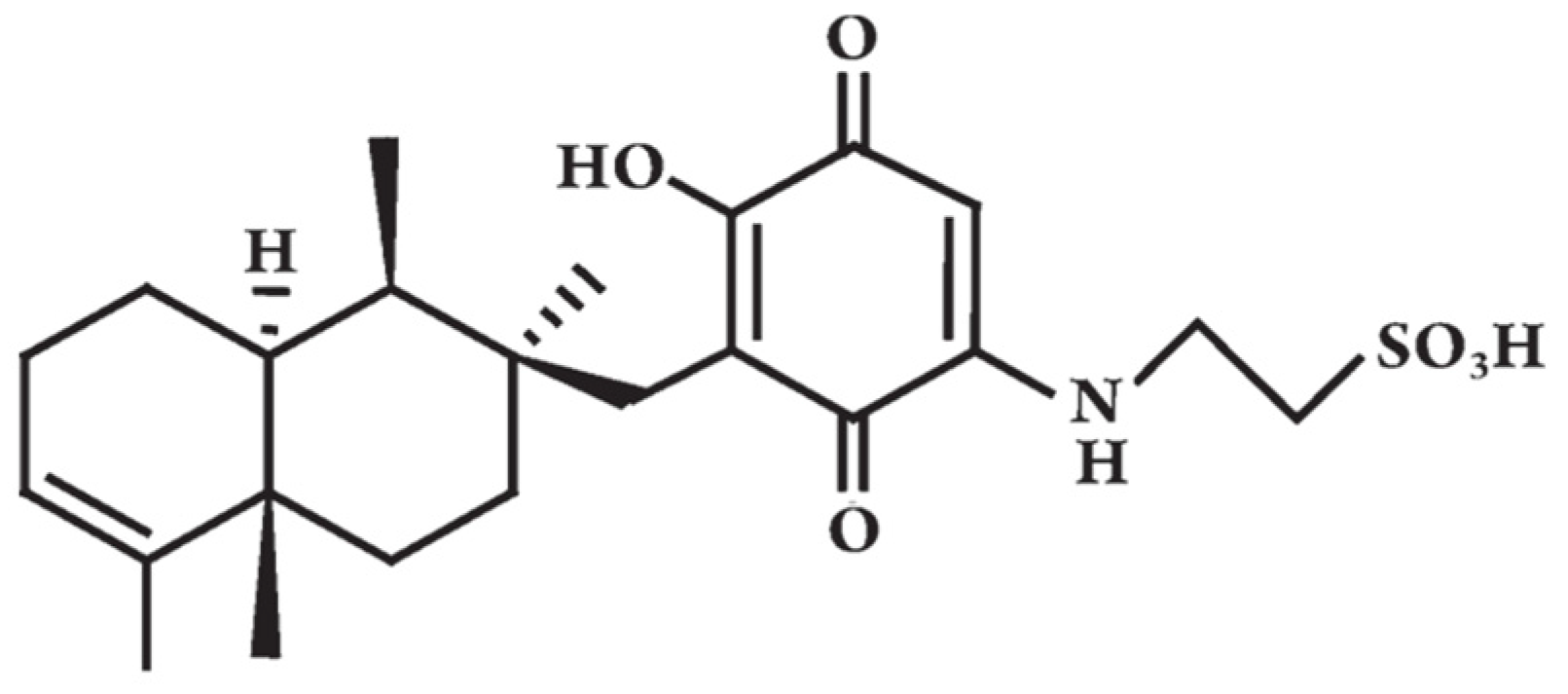
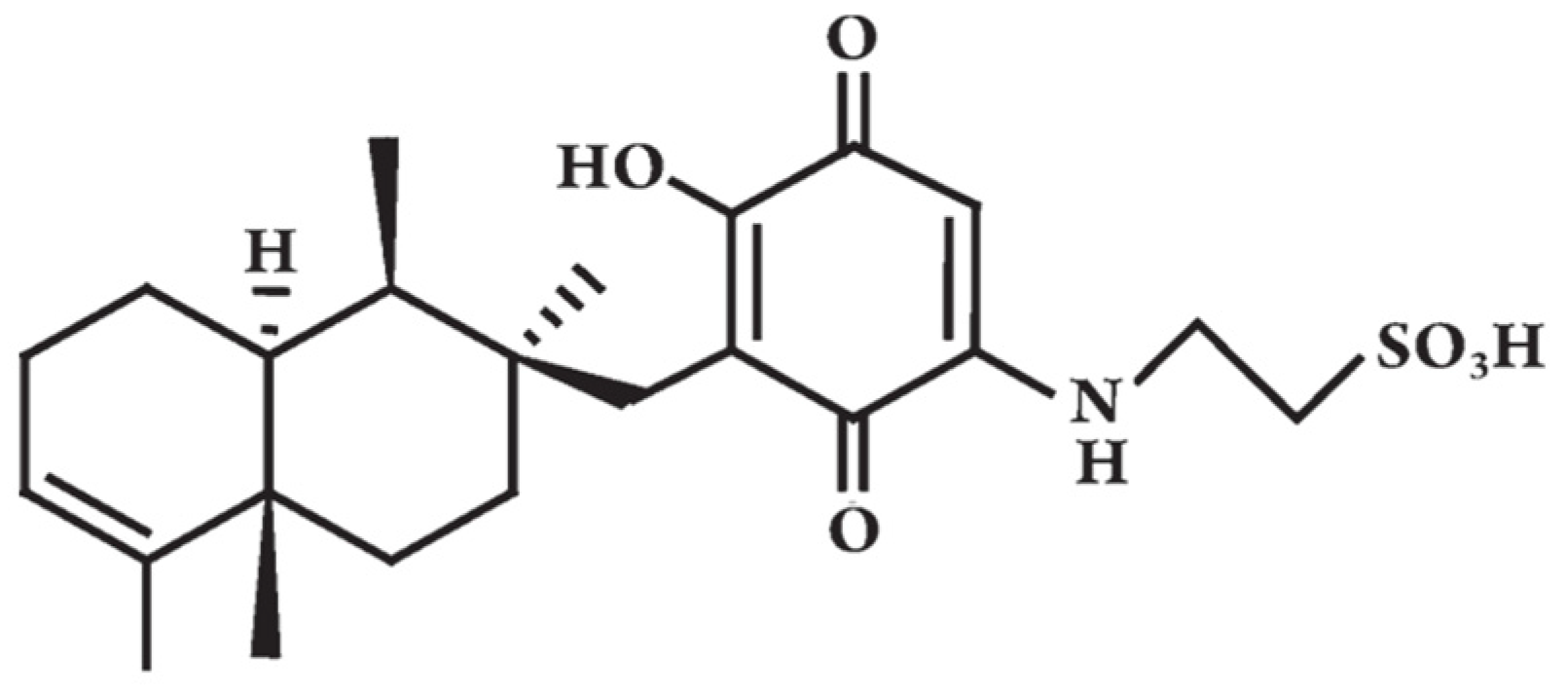{kind=link}
| Species | Possible Compounds | Tested Activity | Reference |
|---|---|---|---|
| 500 freshwater and marine cyanobacteria | N.A. | α-glucosidase inhibition | [46] |
| Bacteria | N.A. | α-amylase and α-glucosidase inhibition | [30] |
| Actinomycetes Streptomyces corchorusii | N.A. | α-amylase inhibition | [26] |
| Actinomycetes Streptomyces sp. | Pyrostatins A and B | N-acetyl-glucosaminidase inhibition | [26] |
| Fungus Cosmospora sp. | Aquastatin A | PTP1B Inhibition | [47] |
| Three clones of the microalgae Chlorella pyrenoidosa, Chlorella protothecoides, three clones of Chlorella vulgaris, four clones of Crypthecodinium cohnii, Nitzschia laevis | Carotenoids, linoleic acid, arachidonic acid, eicosapentaenoic acid | AGE formation inhibition | [43] |
| Microalgae Chlorella zofingiensis | Astaxanthin | AGE formation inhibition | [50] |
| Microalgae Chlorella protothecoides, Chlorella zofingiensis, Nitzschia laevis | Astaxanthin, lutein and eicosapentaenoic acid | AGE formation inhibition | [45] |
| Microalgae Chlorella pyrenoidosa | N.A. | Antioxidant potential, α-amylase and α-glucosidase inhibition | [31] |
| Microalgae Isochrysis galbana, Nannochloropsis oculata | Docosahexaenoic and Eicosapentaenoic acids | Clinical values and intestinal inflammation in rats | [40] |
| Microalgae Attheya longicornis, Chaetoceros socialis, Chaetoceros furcellatus, Skeletonema marinoi and Porosira glacialis | N.A. | PTP1B Inhibition | [53] |
| Several microalgae | N.A. | PTP1B Inhibition | [52] |
| Species | Compounds/Extracts | Mechanism of Action | Reference |
|---|---|---|---|
| Red algae Rhodomela confervoides | 3,4-dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)benzyl)benzene-1,2-diol | PTP1B inhibition | [37,55] |
| Red algae Grateloupia elliptica | 2,4,6-tribromophenol and 2,4-dibromophenol | α-glucosidase, sucrase and maltase inhibition | [27] |
| Red algae Symphyocladia latiuscula | Bromophenols | Aldose reductase inhibition | [56] |
| Red algae Palmaria sp. | Phenolic extracts | α-amylase inhibition | [29] |
| Red algae Palmaria palmata | Protein hydrolysates | Dipeptidyl peptidase IV inhibition | [25] |
| Brown algae Ecklonia cava | Methanolic extracts | Reduce plasma glucose levels in rats | [39] |
| Brown algae Pelvetica siliquosa | Raw extracts | Reduce plasma glucose levels in rats, increase insulin concentration | [57] |
| Brown algae Alaria sp. | Phenolic extracts | α-amylase inhibition | [29] |
| Brown algae Ascophyllum sp. | Phenolic extracts | α-amylase and α-glucosidase inhibition | [29] |
| Brown algae Ecklonia stolonifera | Phlorofucofuroeckol-A | AGEs inhibition | [59] |
| Brown algae Ecklonia stolonifera | Methanolic extracts | Reduce plasma glucose levels in rats | [58] |
| Brown algae Ecklonia cava | Polyphenol-rich extracts | Reduce plasma glucose levels in rats | [39] |
| Brown algae Eisenia bicyclis | Dieckol | α-amylase inhibition | [61] |
| Brown algae Eisenia bicyclis | Eckol | α-amylase inhibition | [61] |
| Brown algae Ishige okamurae | Diphlorethohydroxycarmalol | α-amylase and α-glucosidase inhibition | [62] |
| Green algae Ulva rigida | Raw extracts | Reduce plasma glucose levels in rats | [63] |
| Seagrass Posidonia oceanica | Raw extracts | Reduce plasma glucose levels in rats | [64] |
| Macrophytes Sargassum pallidum, Ulva fenestrata and Zostera marina | Mixture of lipids, Echinochrome A and polyphenols | Protective effects in mice models | [11] |
| Seaweed Cladophora rupestris | Raw extracts | α-amylase and α-glucosidase inhibition | [28] |
| Seaweeds Derbesia marina and Symphycladia latiscula | Raw extracts | PTP1B inhibition | [65] |
| Seaweed Laminaria angustata | Raw extracts | Reduce plasma glucose levels in rats | [66] |
| Brown algae | Fucoxanthin | Inflammation reduction | [8] |
| Sponge Ircinia dendroides | Palinurin | GSK-3β inhibition | [24] |
| Sponge Hemimycale arabica | Phenylmethylene hydantoins | GSK-3β inhibition, increase liver glycogen in rat | [71] |
| Sponge Callyspongia truncata | Callyspongynic acid | α-glucosidase inhibition | [72] |
| Sponge Lamellodysidea herbacea | Polybromodiphenyl ether | PTP1B inhibition | [75] |
| Sponge Xetospongia muta | Aqueous extracts | Dipeptidyl peptidase IV inhibition | [74] |
| Sponge Agelas mauritianus | α-GalCer | Protection beta pancreatic cells | [2] |
| Sponge Dysidea villosa | Dysidine | PTP1B inhibition | [76] |
| Corals Sinularia firma and Sinularia erecta | Methanolic extracts | Reduce plasma glucose levels in rats | [41] |
| Corals Lobophytum pauciflorum and Sarcophyton glaucum, and sponge Sigmadocia pumila | Raw extracts | Reduce plasma glucose levels in rats | [42] |
| Wild fishes | Marine collagen peptides | Decrease free fatty acids and regulate metabolic nuclear receptors in type-2 diabetes patients | [79] |
| Fish oil | n-3 PUFAs | Restoration insulin receptor and its substrate phosphorylation in rat | [80] |
| Sea anemones Bunodosoma granulifera and Bartholomea annulata | Aqueous extracts | Dipeptidyl peptidase IV inhibition | [74] |
| Salmon Oncorhynchus keta skin | Oligopeptides | Antioxidant activity | [13] |
| Salmon Oncorhynchus keta | Gelatin skin | Wound repair in rat skin | [12] |
| Shark | Cholera toxin B subunit and peptide shark liver fusion protein | Protective effects in rat model, inflammation reduction, promote insulin secretion, reduce plasma glucose levels | [4] |
| Fish and shellfish wastes | oil | Lower blood pressure and triacylglycerol concentrations, maintain normal glucose metabolism | [83,84] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauritano, C.; Ianora, A. Marine Organisms with Anti-Diabetes Properties. Mar. Drugs 2016, 14, 220. https://doi.org/10.3390/md14120220
Lauritano C, Ianora A. Marine Organisms with Anti-Diabetes Properties. Marine Drugs. 2016; 14(12):220. https://doi.org/10.3390/md14120220
Chicago/Turabian StyleLauritano, Chiara, and Adrianna Ianora. 2016. "Marine Organisms with Anti-Diabetes Properties" Marine Drugs 14, no. 12: 220. https://doi.org/10.3390/md14120220
APA StyleLauritano, C., & Ianora, A. (2016). Marine Organisms with Anti-Diabetes Properties. Marine Drugs, 14(12), 220. https://doi.org/10.3390/md14120220