Aplysin Sensitizes Cancer Cells to TRAIL by Suppressing P38 MAPK/Survivin Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
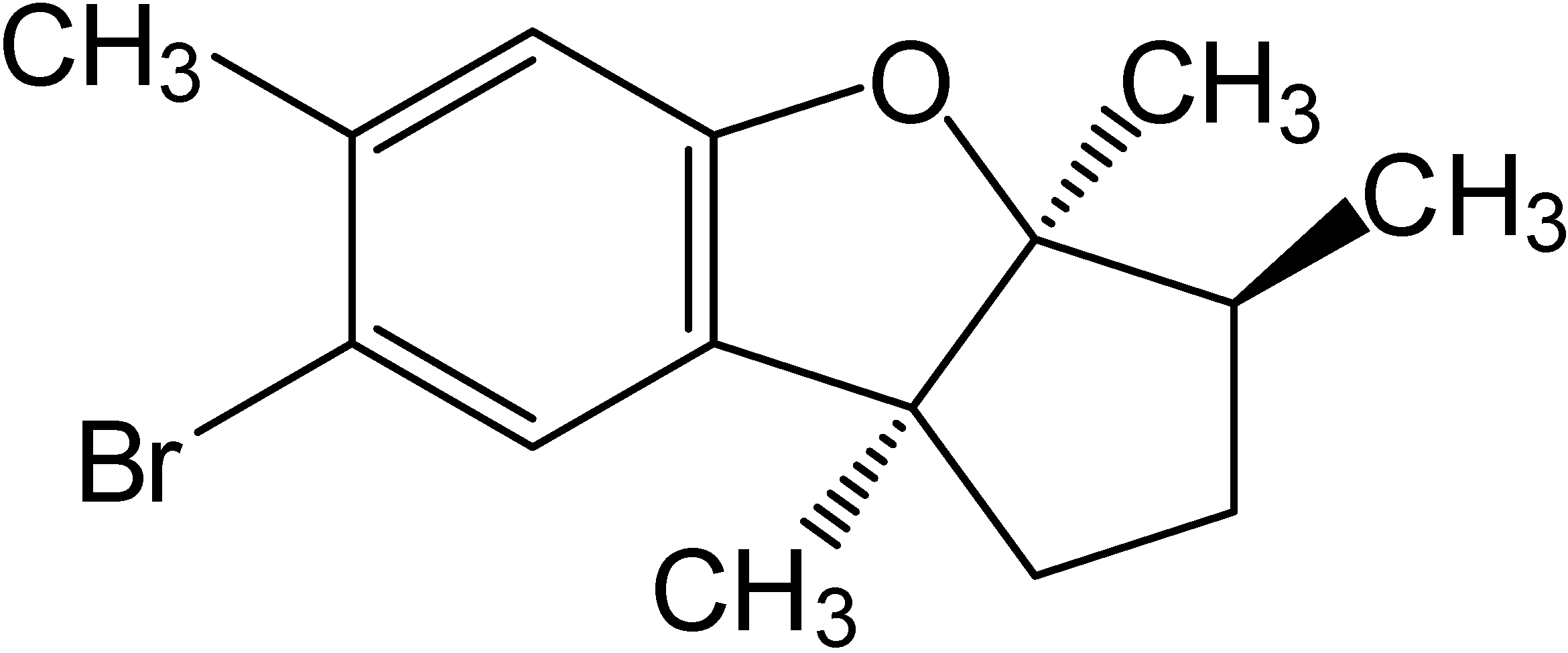
2. Results and Discussion
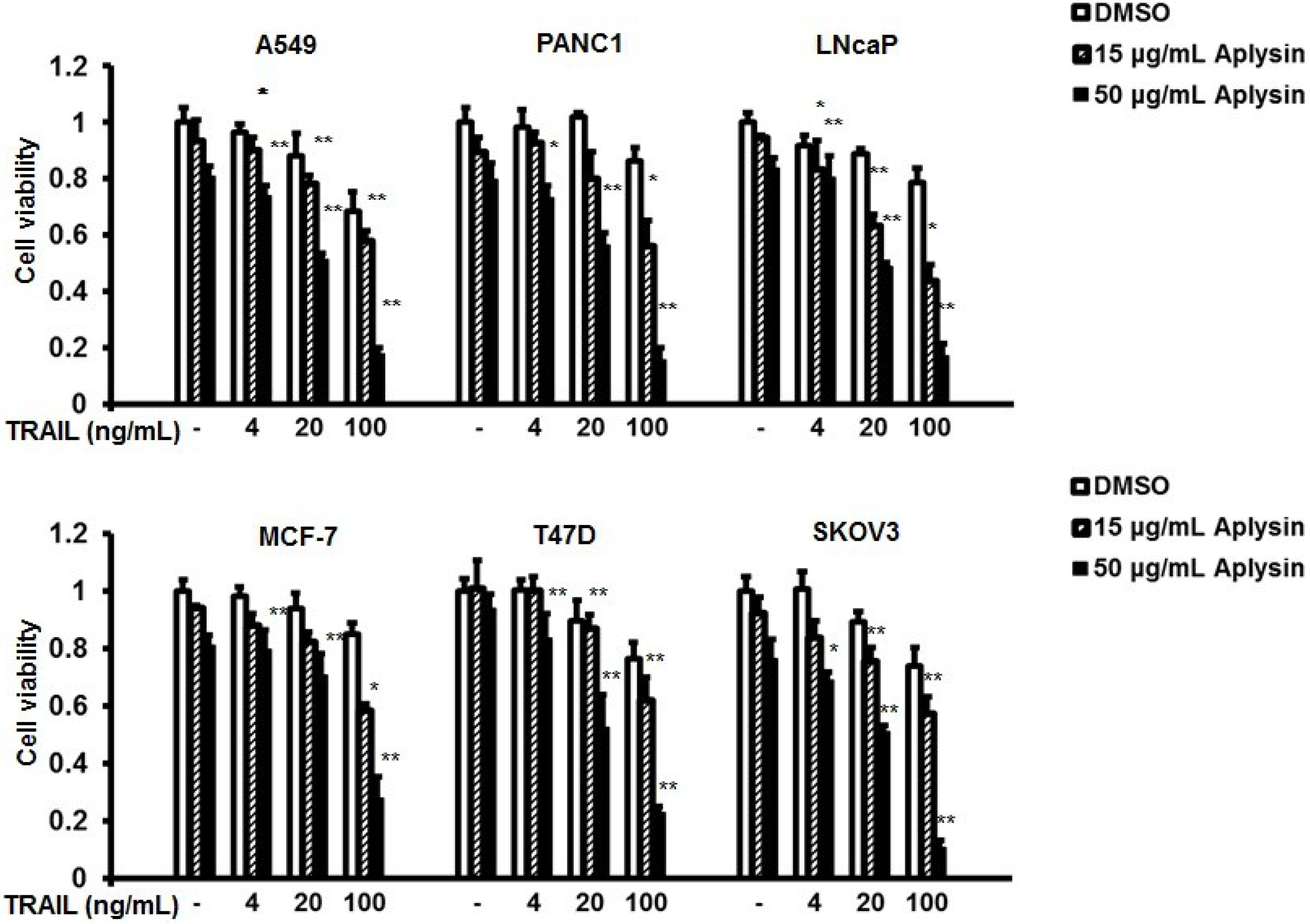
2.1. Aplysin Increases Antitumor Activity of TRAIL on Cancer Cells in Vitro
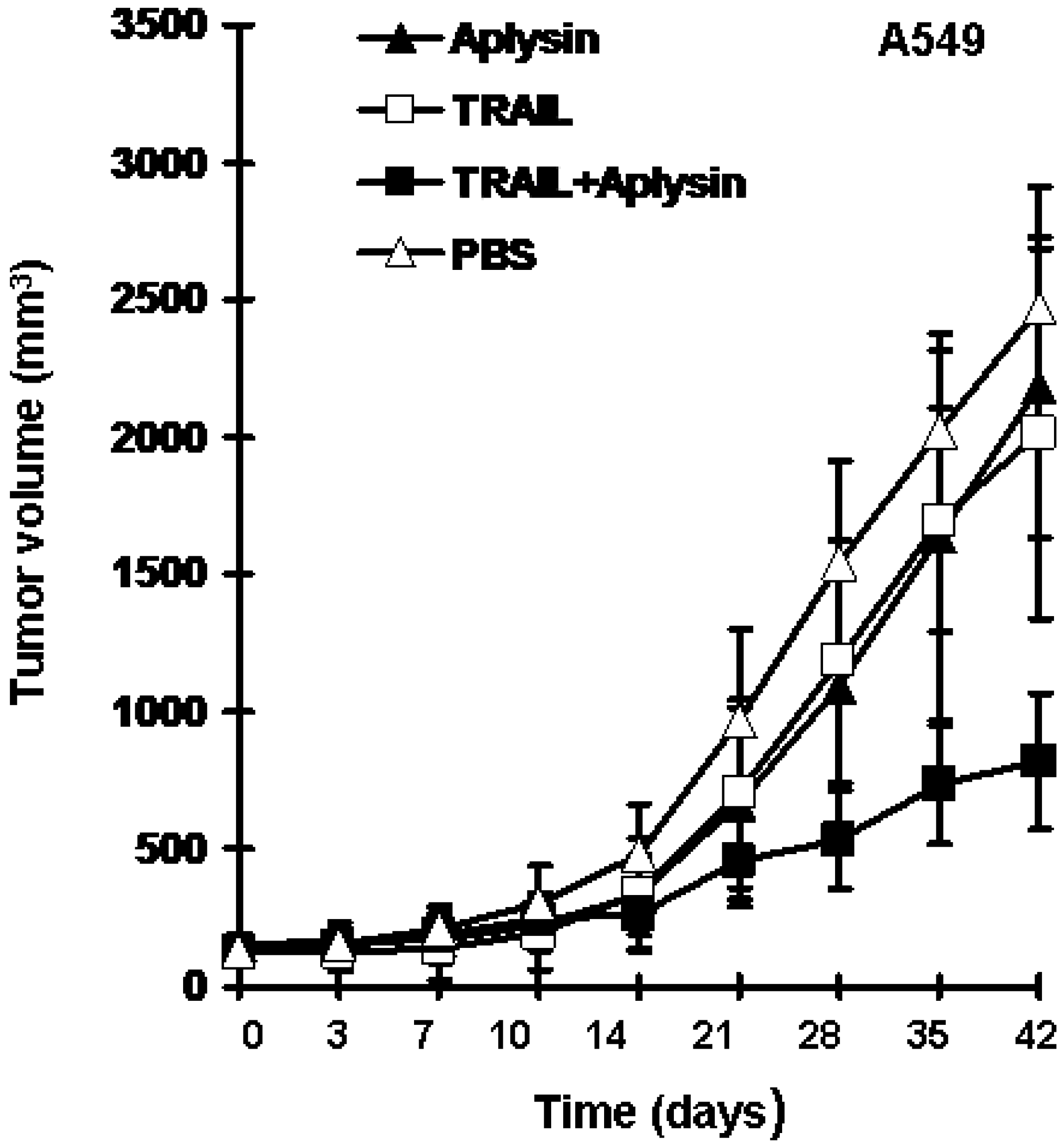
2.2. Aplysin Enhances the Inhibitory Effect of TRAIL on the Growth of Tumor Xenografts in Mice
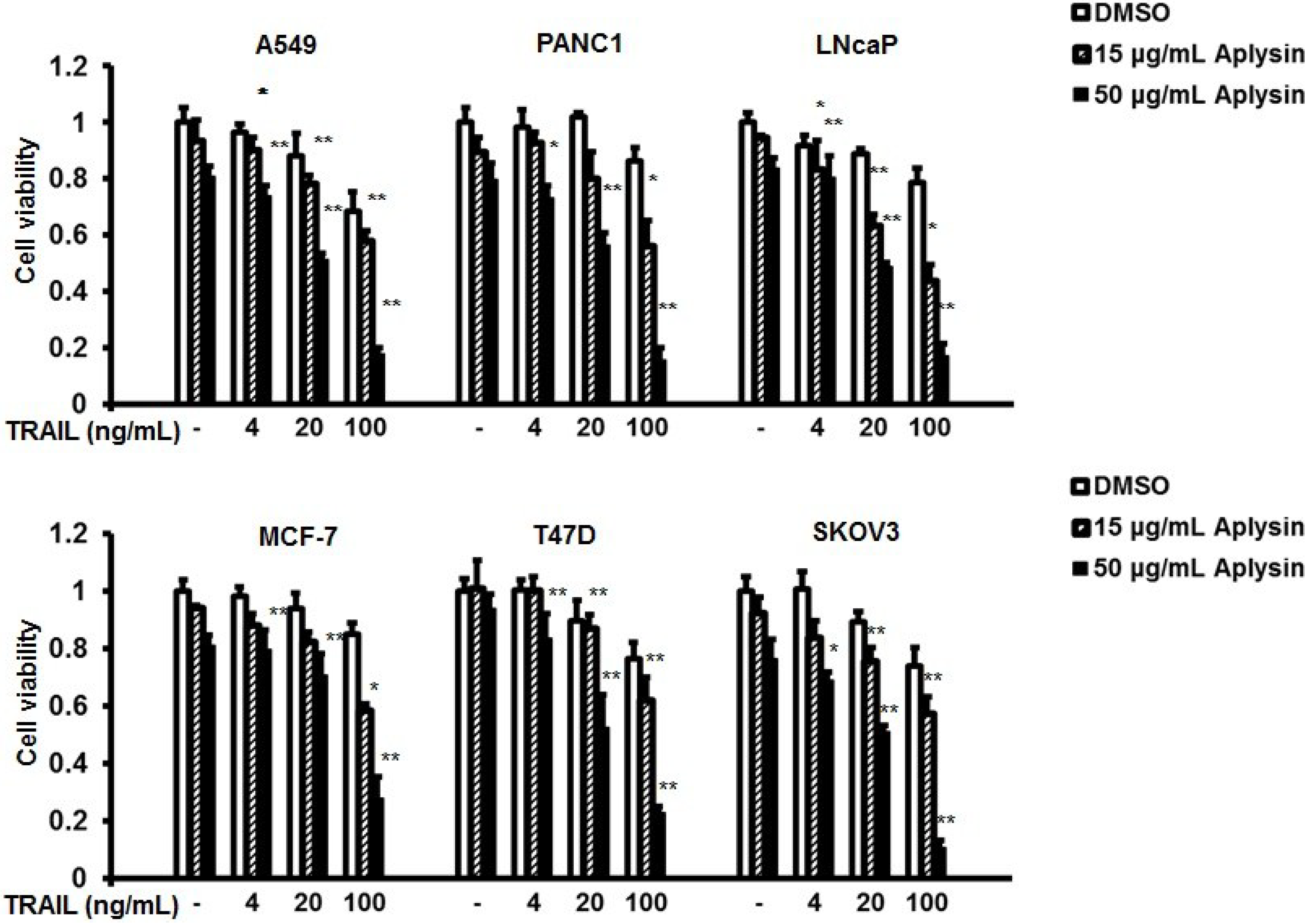
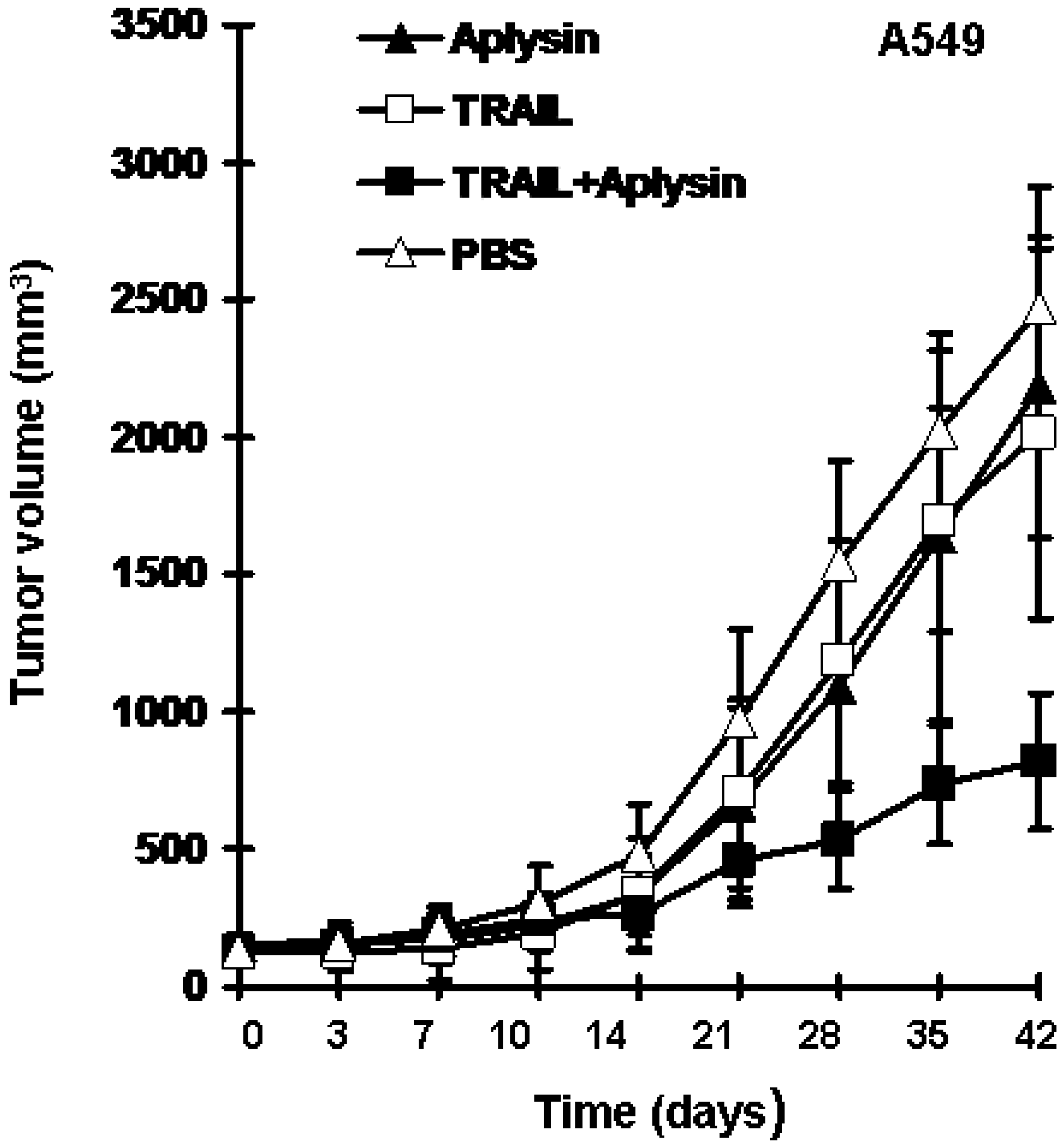
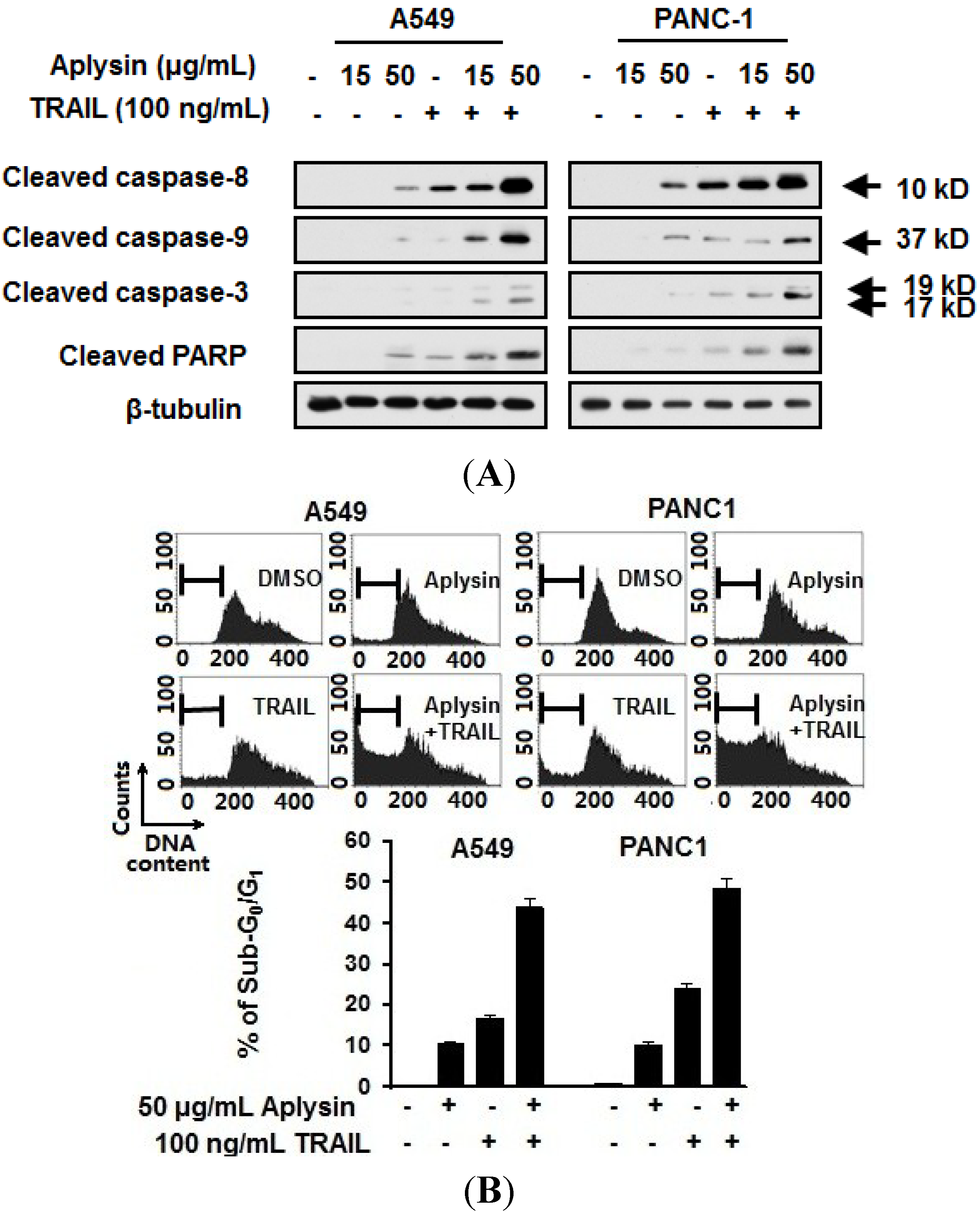
2.3. Aplysin Enhances Apoptosis-Inducing Activity of TRAIL on A549 and PANC1 Cancer Cells
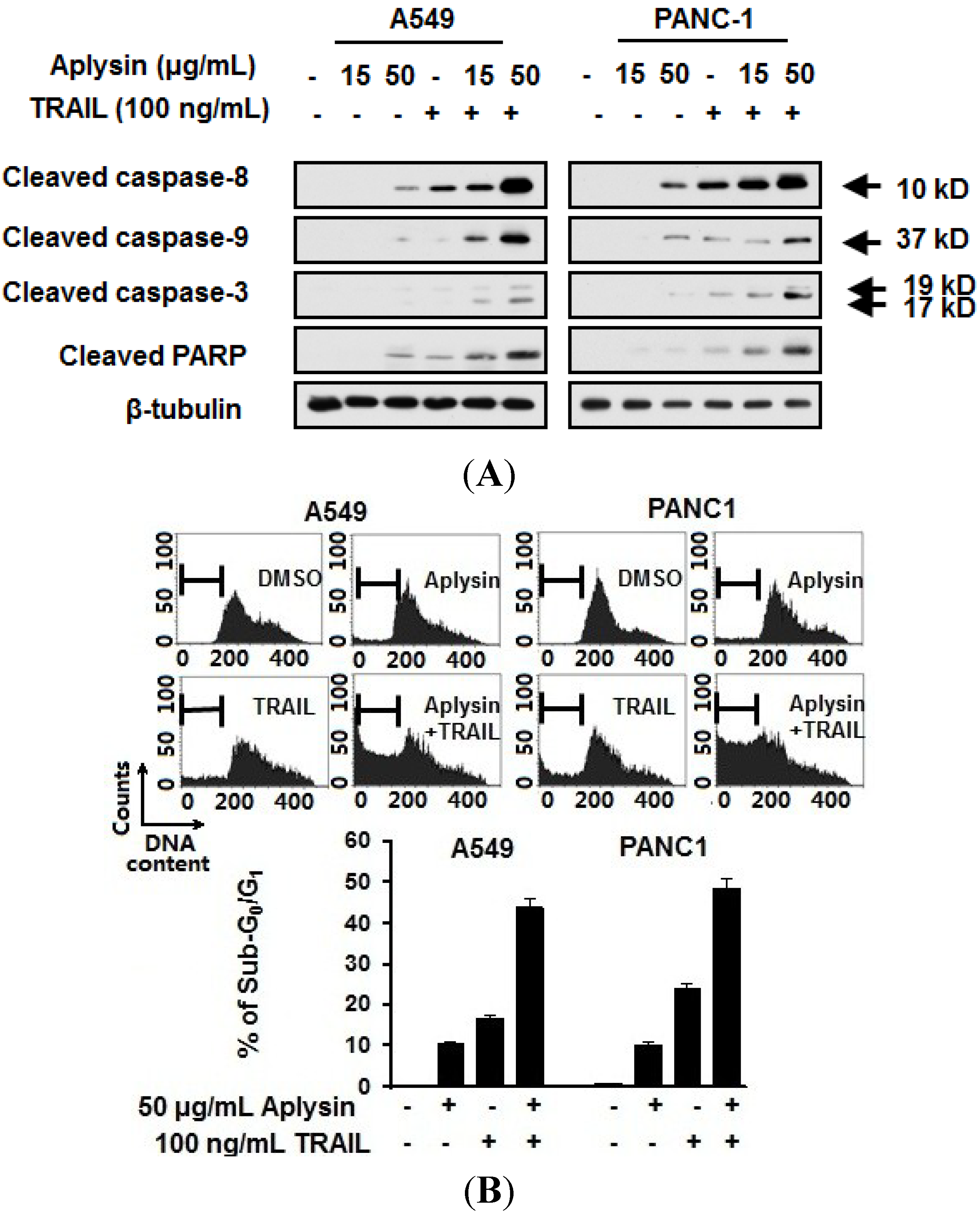
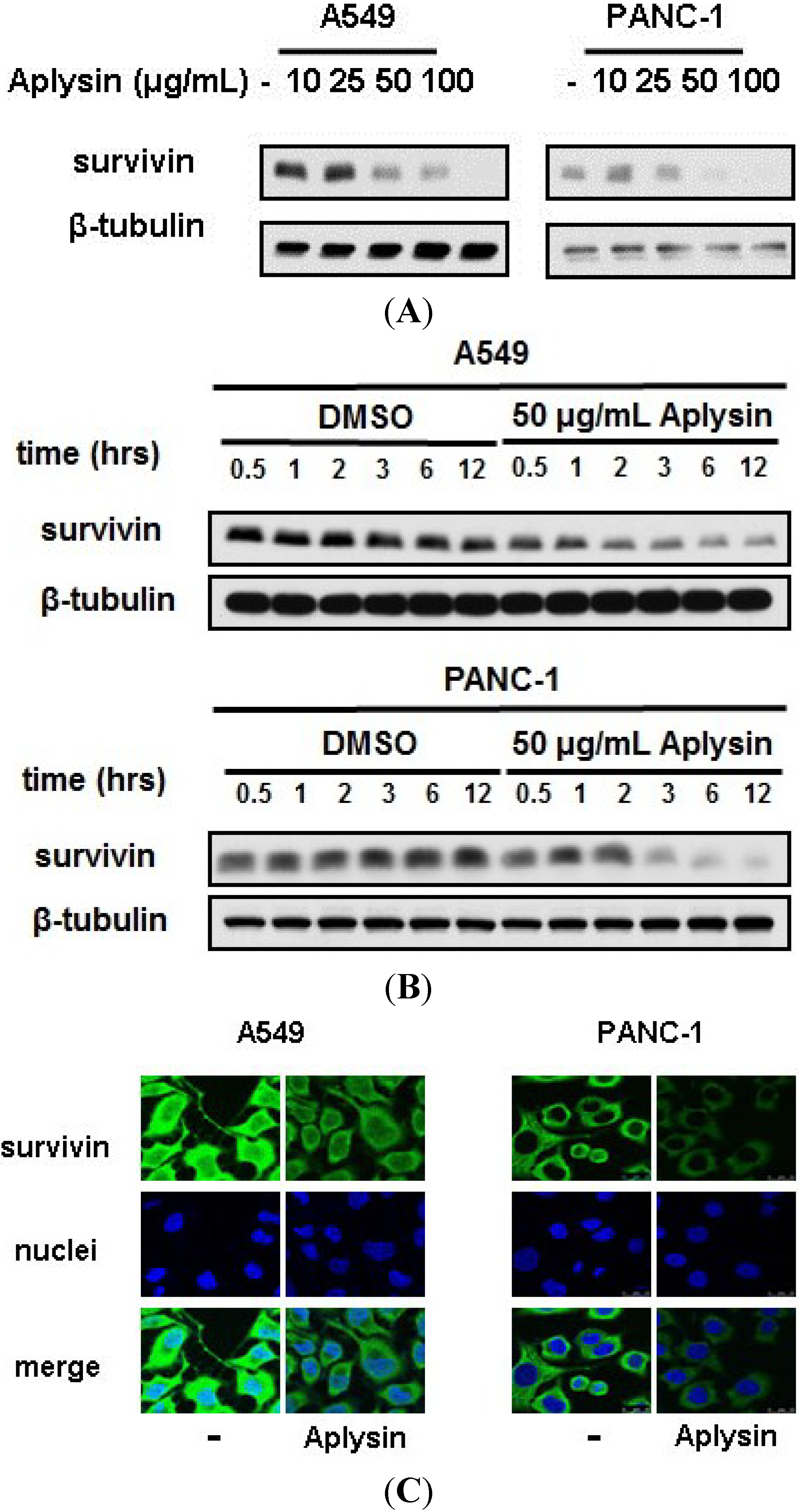
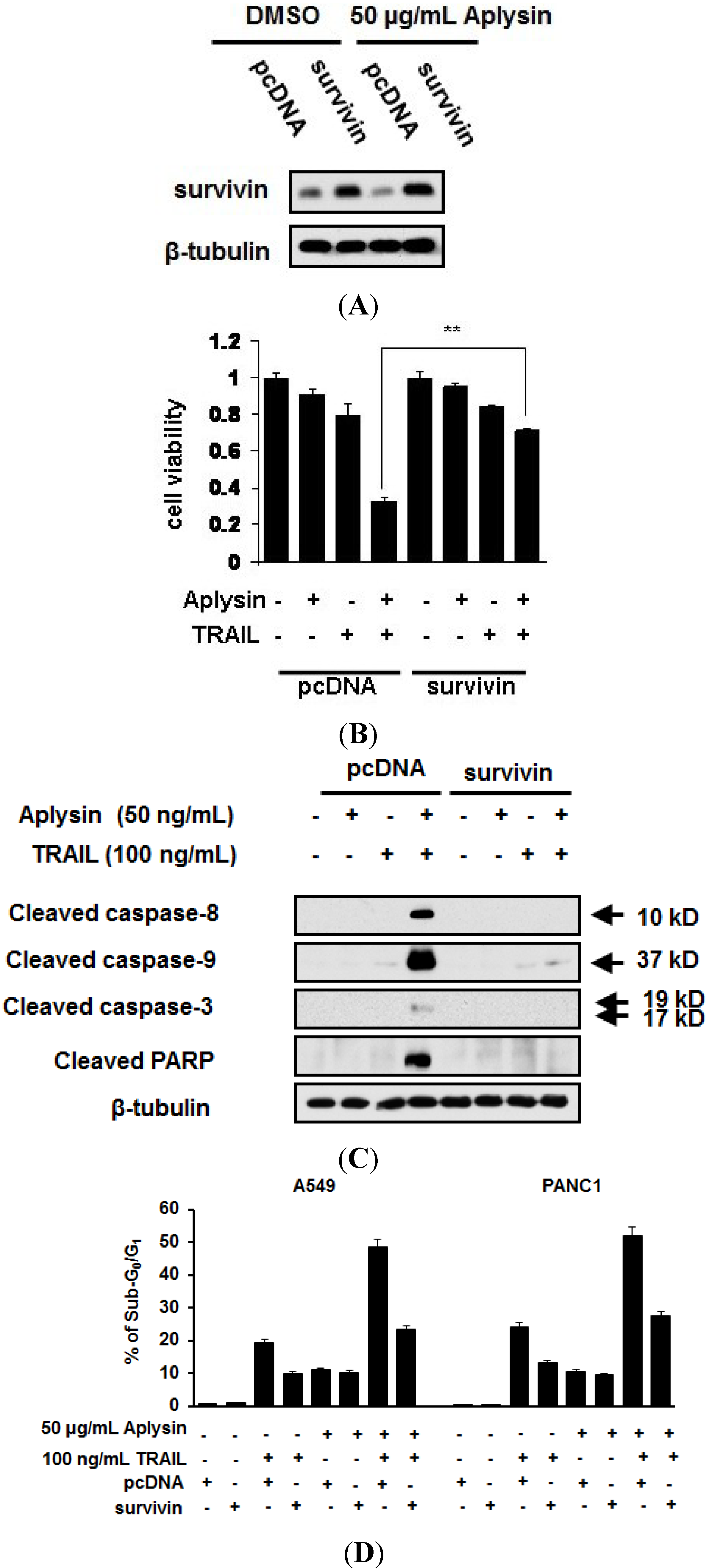
2.4. Survivin Is Downregulated by Aplysin Treatment in TRAIL-Resistant Cancer Cells
2.5. The Decrease in Survivin Expression Level Is Responsible for Aplysin-Caused Cancer Cell Sensitization to TRAIL
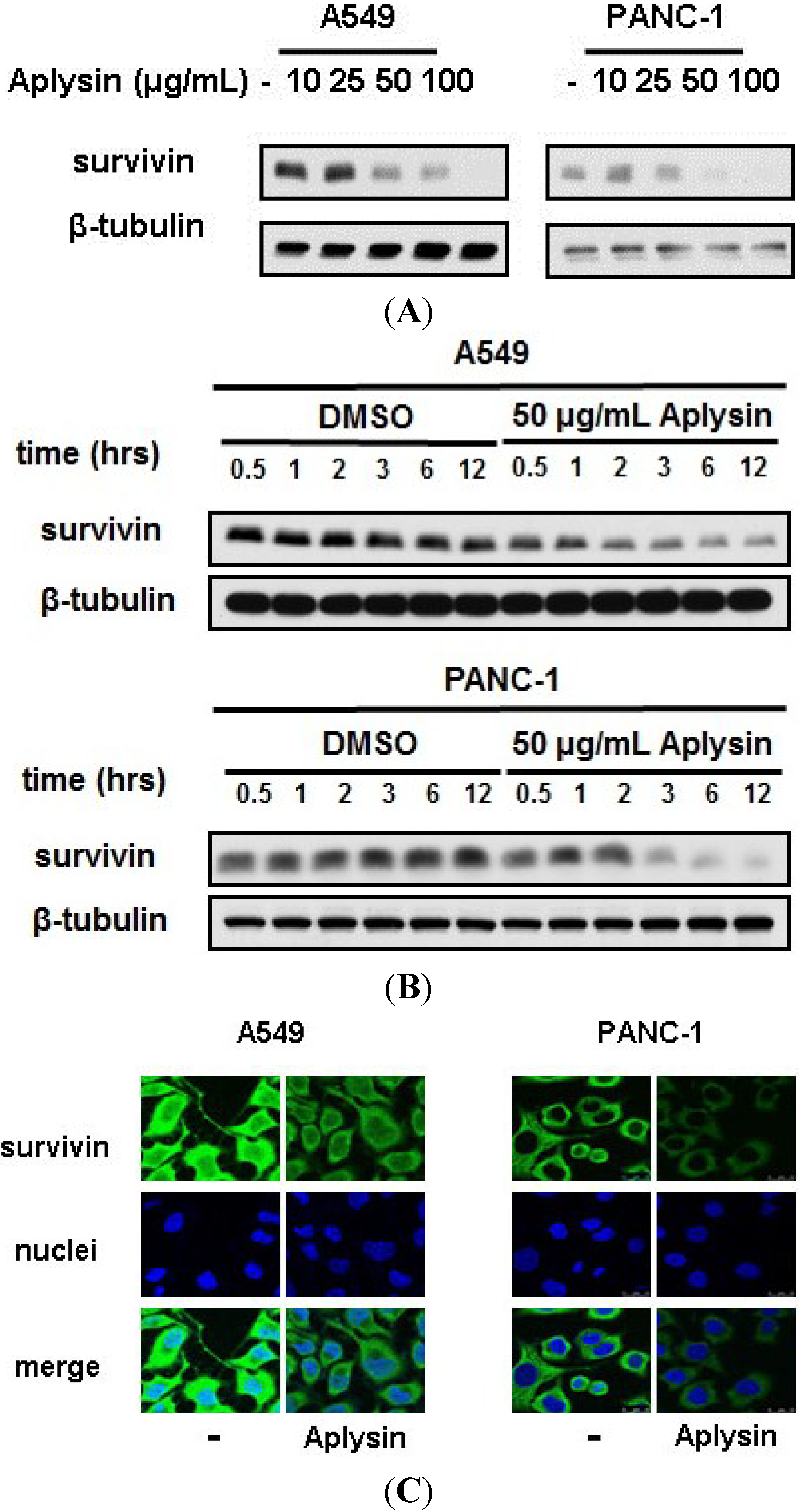
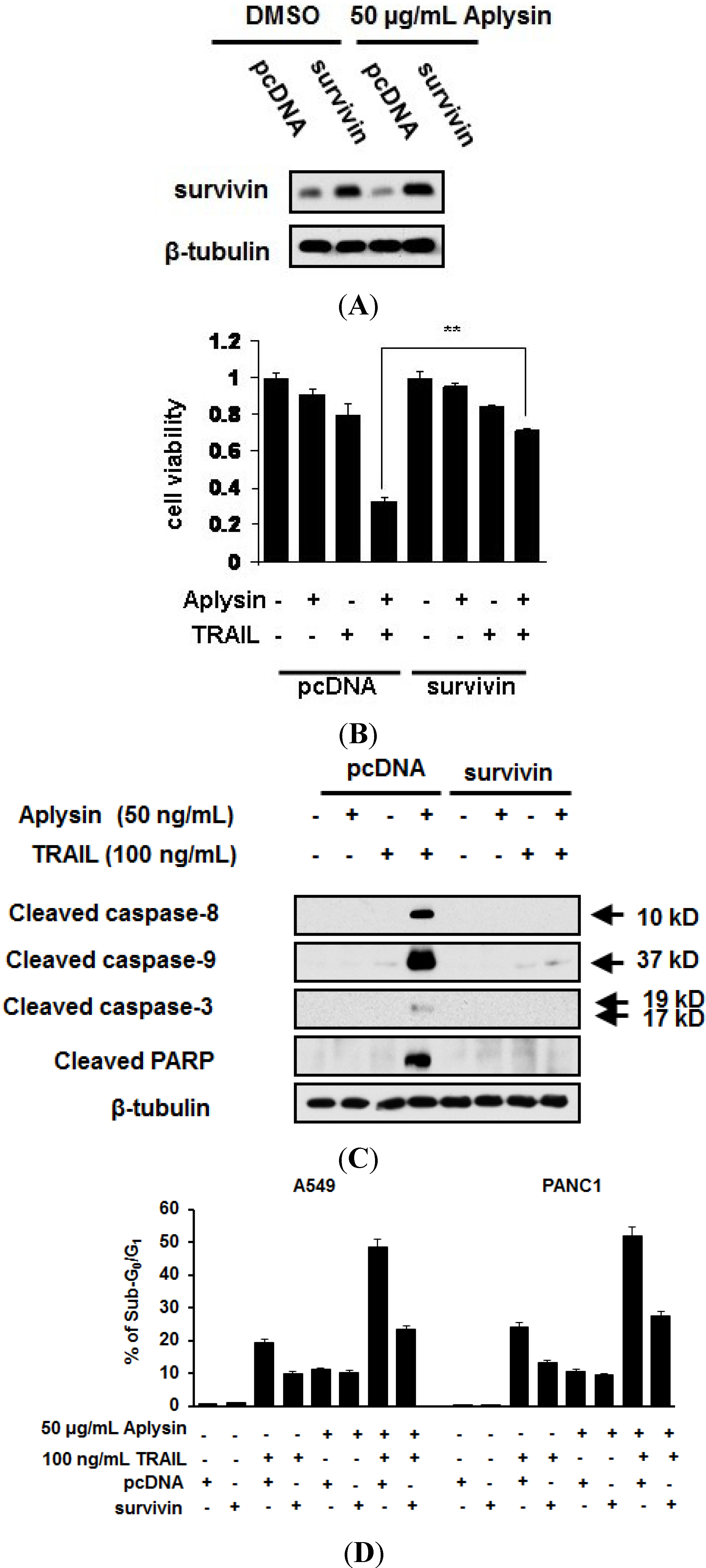
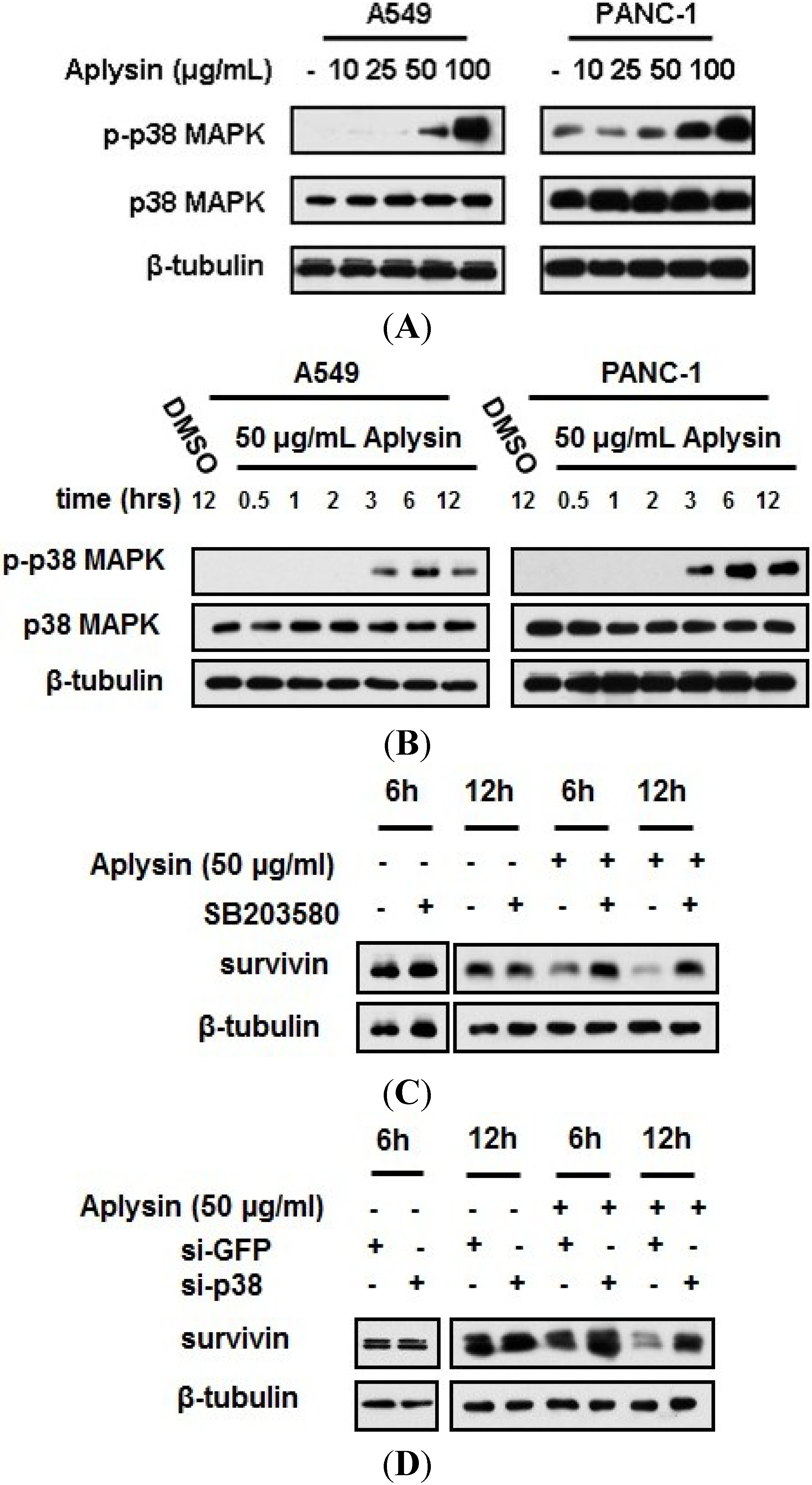
2.6. Aplysin-Induced Survivin Downregulation Was Partially Mediated by p38 MAPK Signaling
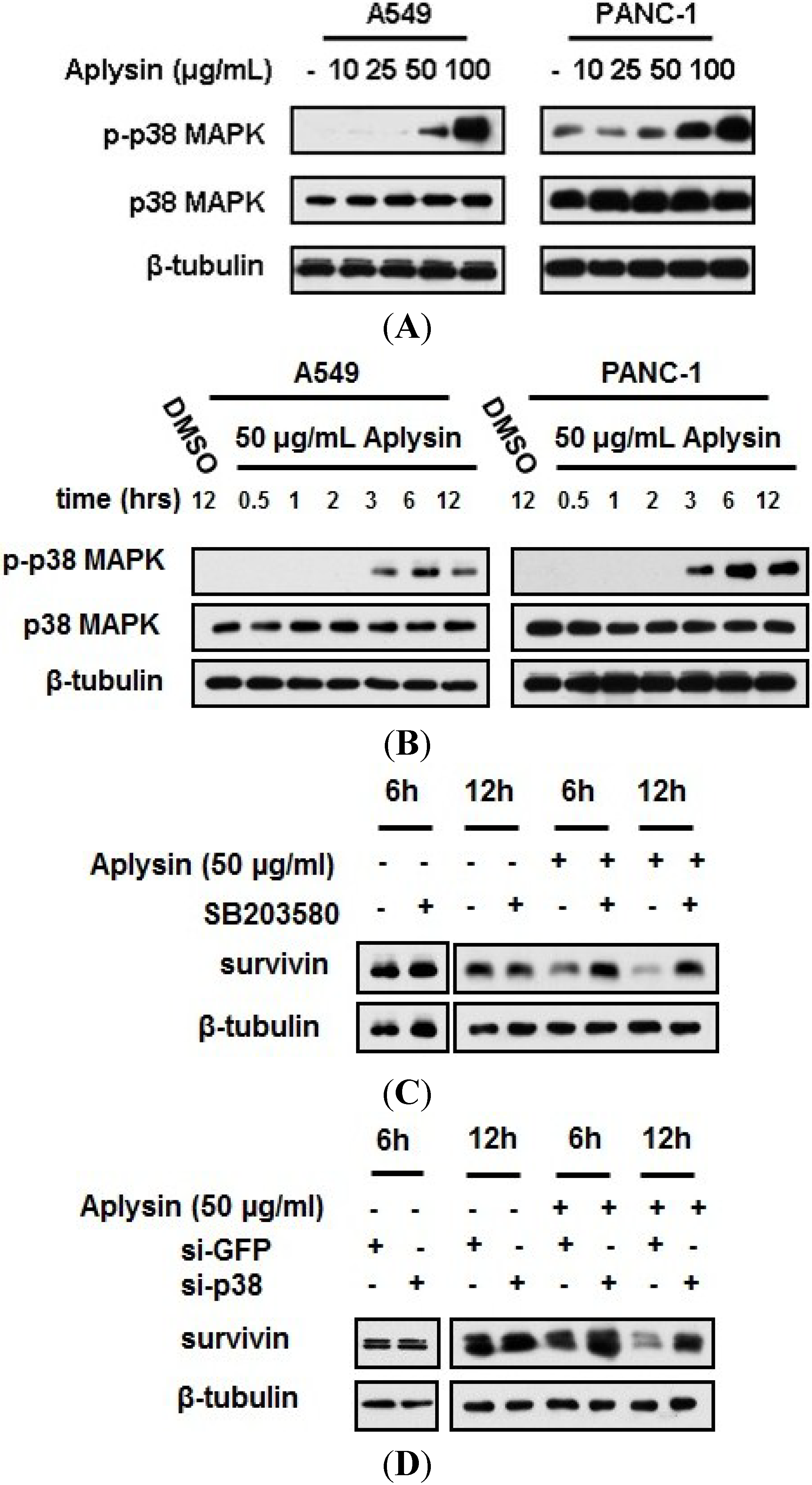
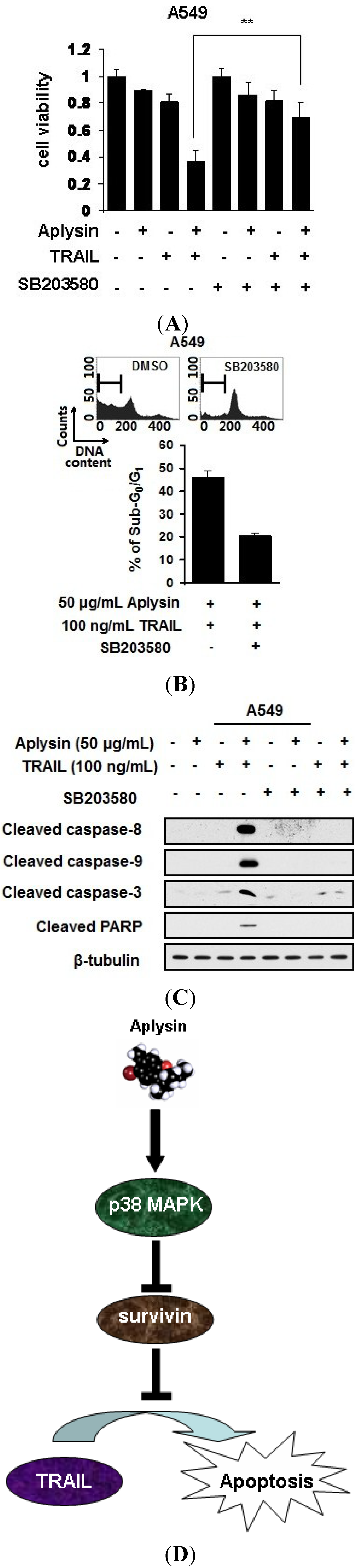
2.7. Aplysin-Induced p38 MAPK Activation Was Partially Required for the Sensitization of Cancer Cells to TRAIL
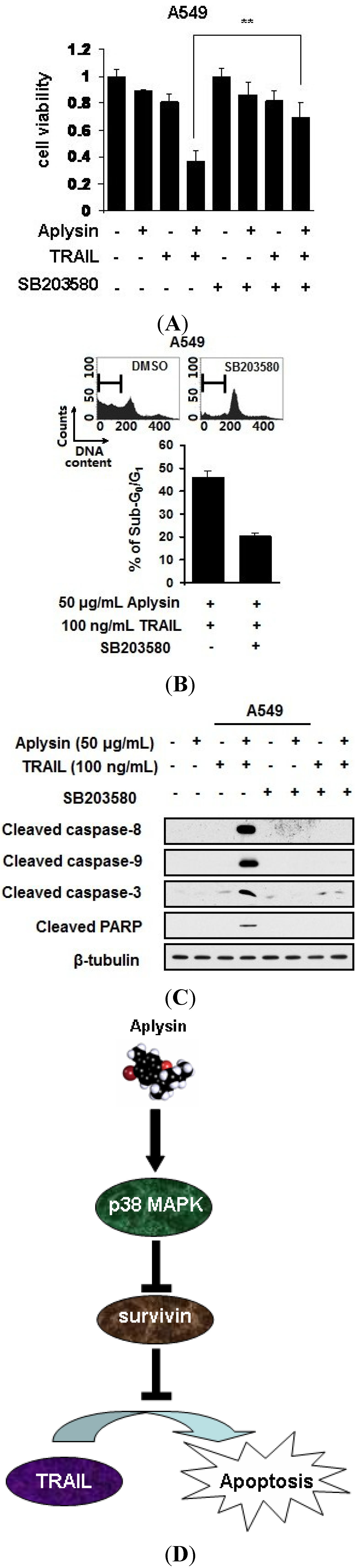
3. Experimental Section
3.1. Cell Culture
3.2. Chemical Reagents
3.3. Cell Viability Assay
3.4. Animal Experiments
3.5. Apoptosis Assays
3.6. Immunoblotting Assay
3.7. Vector Construction and Transfection
3.8. Statistical Analysis
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgements
Author Contributions
Conflicts of Interest
References
- Pan, G.; O’Rourke, K.; Chinnaiyan, A.M.; Gentz, R.; Ebner, R.; Ni, J.; Dixit, V.M. The receptor for the cytotoxic ligand TRAIL. Science 1997, 276, 111–113. [Google Scholar] [CrossRef]
- Yerbes, R.; Palacios, C.; Lopez-Rivas, A. The therapeutic potential of TRAIL receptor signaling in cancer cells. Clin. Transl. Oncol. 2011, 13, 839–847. [Google Scholar] [CrossRef]
- Dimberg, L.Y.; Anderson, C.K.; Camidge, R.; Behbakht, K.; Thorburn, A.; Ford, H.L. On the TRAIL to successful cancer therapy? Predicting and counteracting resistance against TRAIL-based therapeutics. Oncogene 2012, 32, 1341–1350. [Google Scholar] [CrossRef]
- Maksimovic-Ivanic, D.; Stosic-Grujicic, S.; Nicoletti, F.; Mijatovic, S. Resistance to TRAIL and how to surmount it. Immunol. Res. 2012, 52, 157–168. [Google Scholar] [CrossRef]
- Kawasaki, H.; Altieri, D.C.; Lu, C.D.; Toyoda, M.; Tenjo, T.; Tanigawa, N. Inhibition of apoptosis by survivin predicts shorter survival rates in colorectal cancer. Cancer Res. 1998, 58, 5071–5074. [Google Scholar]
- Griffith, T.S.; Fialkov, J.M.; Scott, D.L.; Azuhata, T.; Williams, R.D.; Wall, N.R.; Altieri, D.C.; Sandler, A.D. Induction and regulation of tumor necrosis factor-related apoptosis-inducing ligand/Apo-2 ligand-mediated apoptosis in renal cell carcinoma. Cancer Res. 2002, 62, 3093–3099. [Google Scholar]
- Shigeno, M.; Nakao, K.; Ichikawa, T.; Suzuki, K.; Kawakami, A.; Abiru, S.; Miyazoe, S.; Nakagawa, Y.; Ishikawa, H.; Hamasaki, K.; et al. Interferon-alpha sensitizes human hepatoma cells to TRAIL-induced apoptosis through DR5 upregulation and NF-kappa B inactivation. Oncogene 2003, 22, 1653–1662. [Google Scholar] [CrossRef]
- Gimenez-Bonafe, P.; Tortosa, A.; Perez-Tomas, R. Overcoming drug resistance by enhancing apoptosis of tumor cells. Curr. Cancer Drug Targets 2009, 9, 320–340. [Google Scholar] [CrossRef]
- Sakai, T.; Eskander, R.N.; Guo, Y.; Kim, K.J.; Mefford, J.; Hopkins, J.; Bhatia, N.N.; Zi, X.; Hoang, B.H. Flavokawain B, a kava chalcone, induces apoptosis in synovial sarcoma cell lines. J. Orthop. Res. 2012, 30, 1045–1050. [Google Scholar] [CrossRef]
- Jacquemin, G.; Granci, V.; Gallouet, A.S.; Lalaoui, N.; Morle, A.; Iessi, E.; Morizot, A.; Garrido, C.; Guillaudeux, T.; Micheau, O. Quercetin-mediated Mcl-1 and survivin downregulation restores TRAIL-induced apoptosis in non-Hodgkin’s lymphoma B cells. Haematologica 2012, 97, 38–46. [Google Scholar] [CrossRef]
- Chen, W.; Wang, X.; Zhuang, J.; Zhang, L.; Lin, Y. Induction of death receptor 5 and suppression of survivin contribute to sensitization of TRAIL-induced cytotoxicity by quercetin in non-small cell lung cancer cells. Carcinogenesis 2007, 28, 2114–2121. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Resveratrol-mediated sensitisation to TRAIL-induced apoptosis depends on death receptor and mitochondrial signaling. Eur. J. Cancer 2005, 41, 786–798. [Google Scholar] [CrossRef]
- Pellerito, O.; Calvaruso, G.; Portanova, P.; de Blasio, A.; Santulli, A.; Vento, R.; Tesoriere, G.; Giuliano, M. The synthetic cannabinoid WIN 55,212-2 sensitizes hepatocellular carcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by activating p8/CCAAT/enhancer binding protein homologous protein (CHOP)/death receptor 5 (DR5) axis. Mol. Pharmacol. 2010, 77, 854–863. [Google Scholar] [CrossRef]
- Ge, N.; Liang, H.; Liu, Y.; Ma, A.G.; Han, L. Protective effect of Aplysin on hepatic injury in ethanol-treated rats. Food Chem. Toxicol. 2013, 62, 361–372. [Google Scholar] [CrossRef]
- Hasegawa, H.; Yamada, Y.; Komiyama, K.; Hayashi, M.; Ishibashi, M.; Sunazuka, T.; Izuhara, T.; Sugahara, K.; Tsuruda, K.; Masuda, M.; et al. A novel natural compound, a cycloanthranilylproline derivative (Fuligocandin B), sensitizes leukemia cells to apoptosis induced by tumor necrosis factor related apoptosis-inducing ligand (TRAIL) through 15-deoxy-Delta 12, 14 prostaglandin J2 production. Blood 2007, 110, 1664–1674. [Google Scholar] [CrossRef]
- Lee, J.; Hwangbo, C.; Lee, J.J.; Seo, J.; Lee, J.H. The sesquiterpene lactone eupatolide sensitizes breast cancer cells to TRAIL through down-regulation of c-FLIP expression. Oncol. Re. 2010, 23, 229–237. [Google Scholar]
- Minakawa, T.; Toume, K.; Ahmed, F.; Sadhu, S.K.; Ohtsuki, T.; Arai, M.A.; Ishibashi, M. Constituents of Pongamia pinnata isolated in a screening for activity to overcome tumor necrosis factor-related apoptosis-inducing ligand-resistance. Chem. Pharm. Bull. 2010, 58, 1549–1551. [Google Scholar] [CrossRef]
- Depraetere, V.; Golstein, P. Fas and other cell death signaling pathways. Semin. Immunol. 1997, 9, 93–107. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, X.; Yue, P.; Benbrook, D.M.; Berlin, K.D.; Khuri, F.R.; Sun, S.Y. Involvement of c-FLIP and survivin down-regulation in flexible heteroarotinoid-induced apoptosis and enhancement of TRAIL-initiated apoptosis in lung cancer cells. Mol. Cancer Ther. 2008, 7, 3556–3565. [Google Scholar] [CrossRef]
- Waiwut, P.; Inujima, A.; Inoue, H.; Saiki, I.; Sakurai, H. Bufotalin sensitizes death receptor-induced apoptosis via Bid- and STAT1-dependent pathways. Int. J. Oncol. 2012, 40, 203–208. [Google Scholar]
- Ryu, B.J.; Hwang, M.K.; Park, M.; Lee, K.; Kim, S.H. Thiourea compound AW00178 sensitizes human H1299 lung carcinoma cells to TRAIL-mediated apoptosis. Bioorg. Med. Chem. Lett. 2013, 22, 3862–3865. [Google Scholar] [CrossRef]
- Hsu, Y.F.; Sheu, J.R.; Lin, C.H.; Yang, D.S.; Hsiao, G.; Ou, G.; Chiu, P.T.; Huang, Y.H.; Kuo, W.H.; Hsu, M.J. Trichostatin A and sirtinol suppressed survivin expression through AMPK and p38MAPK in HT29 colon cancer cells. Biochim. Biophys. Acta 2012, 1820, 104–115. [Google Scholar] [CrossRef]
- Sun, J.; Shi, D.; Ma, M.; Li, S.; Wang, S.; Han, L.; Yang, Y.; Fan, X.; Shi, J.; He, L. Sesquiterpenes from the red alga Laurencia tristicha. J. Nat. Prod. 2005, 68, 915–919. [Google Scholar] [CrossRef]
- Weng, D.; Song, X.; Xing, H.; Ma, X.; Xia, X.; Weng, Y.; Zhou, J.; Xu, G.; Meng, L.; Zhu, T.; et al. Implication of the Akt2/survivin pathway as a critical target in paclitaxel treatment in human ovarian cancer cells. Cancer Lett. 2009, 273, 257–265. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, J.; Ma, L.; Wu, N.; Liu, G.; Zheng, L.; Lin, X. Aplysin Sensitizes Cancer Cells to TRAIL by Suppressing P38 MAPK/Survivin Pathway. Mar. Drugs 2014, 12, 5072-5088. https://doi.org/10.3390/md12095072
Liu J, Ma L, Wu N, Liu G, Zheng L, Lin X. Aplysin Sensitizes Cancer Cells to TRAIL by Suppressing P38 MAPK/Survivin Pathway. Marine Drugs. 2014; 12(9):5072-5088. https://doi.org/10.3390/md12095072
Chicago/Turabian StyleLiu, Jia, Leina Ma, Ning Wu, Ge Liu, Lanhong Zheng, and Xiukun Lin. 2014. "Aplysin Sensitizes Cancer Cells to TRAIL by Suppressing P38 MAPK/Survivin Pathway" Marine Drugs 12, no. 9: 5072-5088. https://doi.org/10.3390/md12095072
APA StyleLiu, J., Ma, L., Wu, N., Liu, G., Zheng, L., & Lin, X. (2014). Aplysin Sensitizes Cancer Cells to TRAIL by Suppressing P38 MAPK/Survivin Pathway. Marine Drugs, 12(9), 5072-5088. https://doi.org/10.3390/md12095072