Abstract
Marine sponges harbour complex microbial communities of ecological and biotechnological importance. Here, we propose the application of the widespread sponge family Irciniidae as an appropriate model in microbiology and biochemistry research. Half a gram of one Irciniidae specimen hosts hundreds of bacterial species—the vast majority of which are difficult to cultivate—and dozens of fungal and archaeal species. The structure of these symbiont assemblages is shaped by the sponge host and is highly stable over space and time. Two types of quorum-sensing molecules have been detected in these animals, hinting at microbe-microbe and host-microbe signalling being important processes governing the dynamics of the Irciniidae holobiont. Irciniids are vulnerable to disease outbreaks, and concerns have emerged about their conservation in a changing climate. They are nevertheless amenable to mariculture and laboratory maintenance, being attractive targets for metabolite harvesting and experimental biology endeavours. Several bioactive terpenoids and polyketides have been retrieved from Irciniidae sponges, but the actual producer (host or symbiont) of these compounds has rarely been clarified. To tackle this, and further pertinent questions concerning the functioning, resilience and physiology of these organisms, truly multi-layered approaches integrating cutting-edge microbiology, biochemistry, genetics and zoology research are needed.
1. Introduction
The phylum Porifera (sponges) represents the oldest extant metazoan lineage on Earth, with fossil records dating to around 580 million years ago [1,2]. The ancestry of marine sponges might even be older than previously thought, as chemical records found for demosponges place the origin of these animals back to more than 635 million years ago at the transition between the Ediacaran (Neoproterozoic) and the Cambrian (Paleozoic) periods [3]. Sponges are sessile filter-feeders, with capability to filter thousands of litres of water per day [4,5,6]. They can be found in almost all of the 232 marine ecoregions of the World [7]. Eleven such ecoregions encompass 201–461 species, and are thus considered hot spots of marine sponge diversity [2,7] (Table 1). These animals are restricted to aquatic habitats and divided into four classes: Demospongiae, Hexactinellida, Calcarea and Homoscleromorpha, which are further distributed into 25 orders, 128 families, and 680 genera. The class Demospongiae is by far the richest and most widespread, containing 83% of the 8553 formally recognized species described so far [2].

Table 1.
Hot spots of marine sponge biodiversity.
| Realms 1 | Provinces | Ecoregions | Number of Species 2 |
|---|---|---|---|
| Tropical Atlantic | Tropical North-western Atlantic | Eastern Caribbean | 208 |
| Tropical Atlantic | Tropical North-western Atlantic | Greater Antilles | 275 |
| Temperate Northern Atlantic | Northern European Seas | Celtic Seas | 295 |
| Temperate Northern Atlantic | Lusitanian | South European Atlantic Shelf | 231 |
| Temperate Northern Atlantic | Lusitanian | Azores, Canaries, Madeira | 458 |
| Temperate Northern Atlantic | Mediterranean Sea | Western Mediterranean | 461 |
| Western Indo-Pacific | West and South Indian Shelf | South India and Sri Lanka | 211 |
| Temperate Northern Pacific | Warn Temperate Northwest Atlantic | Central Kuroshio Current | 377 |
| Central Indo-Pacific | Tropical South-western Pacific | New Caledonia | 256 |
| Temperate Australasia | East Central Australian Shelf | Manning-Hawkesbury | 278 |
| Temperate Australasia | Southeast Australian Shelf | Bassian | 375 |
1 “Realms”, “Provinces” and “Ecoregions” are defined according to Spalding and colleagues [7]; 2 The number of sponge species per Ecoregion [7] was obtained from van Soest and colleagues [2].
Although prokaryotic microorganisms are the most important component of their diet, sponges are known to form symbiotic associations with abundant and diverse bacteria and, in some cases, up to 38% of sponge wet weight consists of bacterial cells [4,5,6]. So far, 10 recognized bacterial phyla and 18 candidate bacterial phyla have been detected in marine sponges [5,6,8]. Several lineages within these phyla are involved in the production of secondary metabolites [9], which may enhance host defence mechanisms against predators and invading pathogens. Besides bacteria, archaea and fungi are consistently found as constituent members of the marine sponge microbiome, and specific host-related functions have been proposed or reported for these groups [6,9]. In the light of the enormous richness of marine sponges across the globe, their contribution as the most prolific sources of marine bioactive compounds and the actual participation of their microbial symbionts in secondary metabolite biosynthesis, it is reasonable to posit that marine sponge microbiomes constitute as-yet uncharted, extremely fertile reservoirs of genetic and metabolic novelties. In this context, the present review covers microbial community structure, diversity and bioactivities reported for marine sponges of the family Irciniidae (Demospongiae, Dictyoceratida), known for its wide geographical distribution (Figure 1), chemical complexity and distinct microbiota. In addition, we make use of the Irciniidae family as a model taxon to revisit our knowledge of marine sponge (host and symbiont) cell culturing, metagenomics-based gene discovery, experimental biology, cell-cell signalling and disease. We follow the concept of symbiosis as originally proposed by Anton de Bary (see Taylor et al. [6]) to describe two or more organisms living in close physical association over time. This definition does not imply any sort of mutual benefit between hosts and symbionts.

Figure 1.
Geographical distribution of exemplarily Irciniidae species. The map was drawn using data available at the World Porifera Database; of note is the wide longitudinal occurrence of the genus Psammocinia.
2. The Irciniidae Family as a Model Taxon in Sponge Microbiology Research
With 8553 catalogued and 17,000 estimated species, comprehensively describing microbial diversity, metabolism and functioning in marine sponges is a complex task. Determining how these features relate to host species, habitat, depth and environmental gradients therefore constitutes a true challenge. Given this, the need to identify and develop model hosts in sponge microbiology research has been discussed by specialists at recent meetings such as the 1st International Symposium on Sponge Microbiology [10] and the Lower Invertebrates Symbiosis with Microorganisms Workshop held in Eilat, Israel in 2012. In the past 15 years, intense molecular microbiology research performed on, for example, Aplysina aerophoba from the Mediterranean Sea [11,12,13,14,15], Rhopaloeides odorabile from the Great Barrier Reef (Australia) [16,17,18,19,20] and Ircinia spp. from the Caribbean [21,22,23,24,25] have naturally turned these organisms into model sponge hosts. Strategic and long-term research on key species will most likely improve our understanding of sponge-microbe interactions in a more mechanistic fashion, allowing us to extract pertinent information from a core of relevant hosts that could represent the whole. In this regard, a likely rewarding approach to the choice of model organisms is looking for taxonomic ranks above the species level. This would widen the biodiversity spectrum of the target animals, expanding the geographical, habitat and niche breadths as well as the phenotypic and genotypic plasticity ranges under study. Such a strategy could thus facilitate the assessment of microbiome diversity and function as a response to environmental and host-related factors within a well-contextualized phylogenetic framework. The family Irciniidae is composed only of marine species divided into three genera, namely Ircinia, Sarcotragus and Psammocinia, embracing 75, 11 and 25 accepted species, respectively [26] (see Figure 2 for pictures of some Irciniidae species). Most species have been registered mainly from tropical to temperate regions [27] (see Figure 1 for the geographical distribution of exemplarily species) and inhabit the epipelagic layer (i.e., photic zone where there is enough light for photosynthesis) from zero to 60 m depths. However, a few specimens have been collected from the mesopelagic layer at about 365 m [28]. Irciniidae species exhibit a wide variety of shapes (for a technical description, see [27]) and lack authentic spicules, the archetypical mineral skeleton-forming structures in the majority of demosponges [29]. Instead, fine collagenous and terminally enlarged spongin filaments are typical features of this family. These constitute the organic fibre skeleton, making the sponges very difficult to tear. Several reasons suggest the Irciniidae as a suitable model taxon for microbiology and biotechnology research, as follows:
- The existence of several sympatric species within the family allows testing the host species-specific hypothesis of microbiome composition in sponges using a solid phylogenetic background;
- The wide geographical distribution of some species permits testing hypotheses of symbiont/metabolite maintenance across biogeographical gradients;
- Irciniidae species are chemically rich, a feature that might relate with microbial diversity and suggests biotechnological potential;
- Irciniidae species have been reported as high microbial abundance sponges. This suggests high metabolic activity and/or selective enrichment of their symbionts, which most likely play essential roles for host fitness and survival;
- Bacterial species from at least 15 phyla have been found to be enriched in Irciniidae hosts when compared with surrounding seawater. This distinct microbiome composition is indicative of close host-symbiont relationships;
- The order Dictyoceratida consists of four families—including Irciniidae—collectively known as “keratose” sponges because their skeleton is made of a complex matrix of protein fibres instead of mineral elements (i.e., true spicules), making these families interesting representatives of a peculiar life strategy within marine sponges.

Figure 2.
In situ pictures of Ircinia variabilis (a); Ircinia felix (b); Ircinia campana (c); Sarcotragus spinosulus (d) and of a fixed specimen of Psammocinia compacta (e); Photos courtesy: Francisco R. Pires (a–d); Panel (e) is reproduced with permission from Guilherme Muricy (Department of Invertebrates, Federal University of Rio de Janeiro, Brazil).
In the following sections, we provide a historical perspective on the marine sponge microbiome, with focus on irciniid hosts (see Figure 3 for a schematic representation of a specimen from the family). It is not the scope of this review to describe all bioactive compounds retrieved from Irciniidae species. Instead, an extensive list of metabolites documented in these sponges is given as Supplementary Information, with notes of a possible microbial origin of the described compounds.

Figure 3.
Schematic overview of a demosponge from the family Irciniidae. Mesohyl (M), ostium (Ot), pinacoderm (Pi), porocyte (Po), choanocyte (C); archeocyte (A); and osculum (Oc); arrows indicate the water flow within the sponge body.
3. Microbial Diversity and Bioactivities in the Family Irciniidae
Microbial diversity surveys have focused primarily on Caribbean species belonging to the genus Ircinia [21,22,23,24,30,31,32,33] and on Ircinia and Sarcotragus species from the eastern Atlantic Ocean and Mediterranean Sea [34,35,36,37,38,39,40]. However, no direct comparison between prokaryotic communities from irciniids inhabiting these regions has been performed. Previous studies classified members of the Irciniidae family as high microbial abundance (HMA) sponges [31,38], highlighting their capability of hosting highly dense microbiomes, ranging from 108–1010 prokaryotic cells·g−1 of sponge (fresh weight). Although several attempts have been made to isolate secondary metabolites from these sponges (see Supplementary Information), little attention was paid to determine the actual producer (i.e., host or symbiont) of such compounds.
3.1. Early Microbiology Studies
First insights into the microbial abundance and morphological diversity in sponges were obtained by electron microscopy—especially transmission electron microscopy (TEM) [4,41], still an important tool in several studies [12,42,43,44,45]. In the early 1970s, Sara [46] showed that I. variabilis was populated by the cyanobacteria Aphanocapsa feldmanni and A. raspaigellae. The former was detected in the cortical mesohyl—the intercellular matrix of sponges—and inside sponge cells, while the latter was found only in the mesohyl, surrounded by a lacunar space. Both species reproduced outside the host’s cells. A true cooperative relationship was suggested between host and symbionts, whereby Aphanocapsa spp. supplied the sponge with organic material originated from photosynthesis, protected the host from excessive illumination and could use the nitrogenous compounds eliminated by the sponge, whereas I. variabilis offered shelter and protection for the Cyanobacteria species [46]. Using scanning and transmission electron microscopy, Wilkinson [47,48] revealed higher abundance of heterotrophic bacteria in the dense mesohyl of I. wistarii from the Great Barrier Reef (Australia) when compared with surrounding seawater (SSW). These symbionts concentrated around the inhalant canals of the sponge. Although pioneering TEM analyses enabled first assessments of the abundance and morphology of bacteria in sponges, the taxonomic diversity and composition of these symbionts remained enigmatic until the first culture-dependent and -independent inventories of sponge-associated bacteria were created. These were ultimately responsible for the enormous increase in knowledge regarding sponge microbiome diversity in the last 35 years.
3.2. Bacteria
3.2.1. Diversity
3.2.1.1. Culture-Dependent Approaches
Wilkinson [48] isolated 87 bacterial colonies from I. wistarii and performed 76 tests measuring morphology, physiology and metabolic capabilities. Fifty-nine isolates were grouped into six well-defined clusters, whereas 28 isolates were too ambiguous to be assigned to any clade. Surprisingly, these isolates were able to metabolize a wide range of compounds and, for the first time, comprehensive information on the physiology of sponge-associated bacteria was provided. During the subsequent 25 years, no attempt was made to cultivate symbionts from Irciniidae species, whereas several other sponge hosts have been approached in this manner. These studies usually reported on Alpha- and Gamma classes of Proteobacteria as the prevailing members of the culturable sponge-associated microbiota [42,43,49]. Likewise, the culturable assemblage of Ircinia sp. (St. Giovanni, Croatia) was later found to be dominated by Alpha and Gammaproteobacteria [50]. In another survey, Esteves and colleagues [37] isolated more than 270 bacterial strains from S. spinosulus and I. variabilis on marine agar. These were classified into 17 genera, and 10 putative new species were detected. In accordance with several cultivation-dependent studies performed with other hosts [42,51,52,53], the bacterial genera Pseudovibrio, Ruegeria and Vibrio (Proteobacteria) prevailed among the retrieved symbionts [37]. Using six Actinobacteria-specific media, Tabares and colleagues [54] isolated only one single strain, Arthrobacter sp. BA51, from I. felix. The main advantage of bacterial cultivation is that this allows the determination of the putative functions of these symbionts. For instance, some studies [37,50] have tested bacterial cultures from Irciniidae specimens for in vitro antimicrobial activity (see Section 3.2.2), an approach widely used in sponge microbiology. However, the biases inherent in culture-dependent methods, such as <1% of the total bacterial cells being recovered [42,55], with most of the isolates belonging to the Proteobacteria [6,43,56], have favoured the use of cultivation-independent techniques in the characterization of sponge-associated bacterial communities.
3.2.1.2. Cultivation-Independent Approaches
The application of DNA-based, molecular approaches to the marine sponge microbiome, relying mainly on the sequencing or fingerprinting of 16S rRNA genes, has illuminated our view of the bacterial community structure and diversity in these organisms by circumventing the well-known limitations of culturing techniques. Indeed, a meta-analysis performed with 16S rRNA gene sequences derived from irciniid sponges via cultivation-dependent and -independent methods reveals that the latter strategy uncovers much higher bacterial diversity from these hosts than the former (Figure 4). Below, we first address pioneering fingerprinting/cloning-and-sequencing studies, and then recent surveys that use next generation sequencing to inspect bacterial community structure in marine sponges with cultivation-independent methods.
Using a Cyanobacteria-specific primer pair, Usher and colleagues [57] detected, by polymerase chain reaction-denaturing gradient gel electrophoresis (PCR-DGGE), two closely related species of Synechococcus in I. variabilis. PCR-DGGE fingerprinting and clone-and-sequencing of 16S rRNA gene fragments using universal bacterial primers were later applied to determine the structure of bacterial communities associated with I. felix and I. strobilina from Key Largo, Florida, USA [23,30,31]; I. strobilina from Sweetings Cay, Bahamas [32]; and the Mediterranean I. variabilis, I. fasciculata (currently accepted taxon: Sarcotragus fasciculatus. For the sake of clarity, in this review we will keep the source name Ircinia fasciculata whenever referring to previous studies) and I. oros [34]. These studies revealed Acidobacteria, Actinobacteria, Chloroflexi, Cyanobacteria, Gemmatimonadetes, Nitrospira and Proteobacteria (Alpha, Delta and Gamma classes) as the prevailing bacterial phyla in Ircinia spp. It was also demonstrated that irciniids harboured a distinct microbiome compared to SSW [23,30,31,32,33,34]. Erwin and colleagues [34] showed that 38 out of 56 sequences fell within “sponge-specific” or sponge- and coral-specific clusters. The number of 99% operational taxonomic units (OTUs, here defined at 99% gene sequence similarity) found in I. fasciculata, I. oros and I. variabilis was 29, 33 and 34, respectively. Only four OTUs were registered in all three Ircinia species, revealing that each sponge host harboured its own unique bacterial community. Erwin and colleagues [35] further monitored bacterial community structures in the aforementioned species and SSW once every season for 18 months. Terminal restriction fragment length polymorphism (T-RFLP) analysis revealed that the bacterial communities within each host were highly stable over time. A 16S rRNA gene cloning-and-sequencing procedure using universal bacterial primers was further employed, whereby the same sponge individuals were collected in winter and summer. This showed that >50% of the sponge symbionts were maintained across seasons with no significant differences in community structure. Seven out of eight OTUs were closely related to typical sponge-associated bacteria, of which Candidatus S. spongiarum (Cyanobacteria) was the most dominant followed by OTUs affiliated with Deltaproteobacteria, Acidobacteria, Gammaproteobacteria, Nitrospira and Cyanobacteria. Erwin and colleagues [36] also unravelled Cyanobacteria diversity and abundance in I. fasciculata and I. variabilis using several approaches. Cyanobacterial cells were found in the ectosome of both sponge species in dense populations, but I. fasciculata contained almost twice the level of chlorophyll a and higher abundance of glycogen granules than I. variabilis. Candidatus S. spongiarum was again dominant, being detected intercellularly in the mesohyl, actively reproducing, and seemed to interact with host cells. Synechocystis sp. was sporadically found in I. fasciculata, and it was neither in reproductive process nor in close association with host cells. These results corroborate the findings of Usher and colleagues [57], suggesting that cyanobacterial symbionts play an important role in photosynthesis within these sponges [36]. Hardoim and colleagues [38] determined the bacterial abundance, diversity and community composition in S. spinosulus and I. variabilis. Both sponge species were for the first time classified as HMA sponges based on epifluorescence microscopy analysis. PCR-DGGE profiles of S. spinosulus and I. variabilis were dissimilar, showing that each sponge species harboured its own bacterial community, strengthening the findings of Erwin and colleagues [34] on specific bacterial assemblages inhabiting sympatric Irciniidae species. Pita and colleagues [39] further examined the spatial stability of bacteria associated with I. fasciculata, I. oros and I. variabilis collected at six locations in the western Mediterranean Sea. T-RFLP analyses showed no reliable clustering of sponge symbiont communities according to their respective sampling sites, suggesting high stability of these assemblages across distances from 80–800 km. Pita and colleagues [33] also disclosed the spatial variability and host specificity of bacteria associated with I. strobilina and two colour morphs of I. felix collected at five islands representing different environmental conditions in the Bahamas. T-RFLP fingerprinting uncovered different bacterial communities from I. strobilina and I. felix. However, the same was not true for both colour morphs of I. felix. No significant correlations between bacterial community similarity and sampling sites were detected for both sponge species. The authors suggested that periodic horizontal (i.e., seawater) acquisition together with continuous vertical transmission of symbionts (see below) might contribute to the specificity of bacterial communities within each host sponge [33].
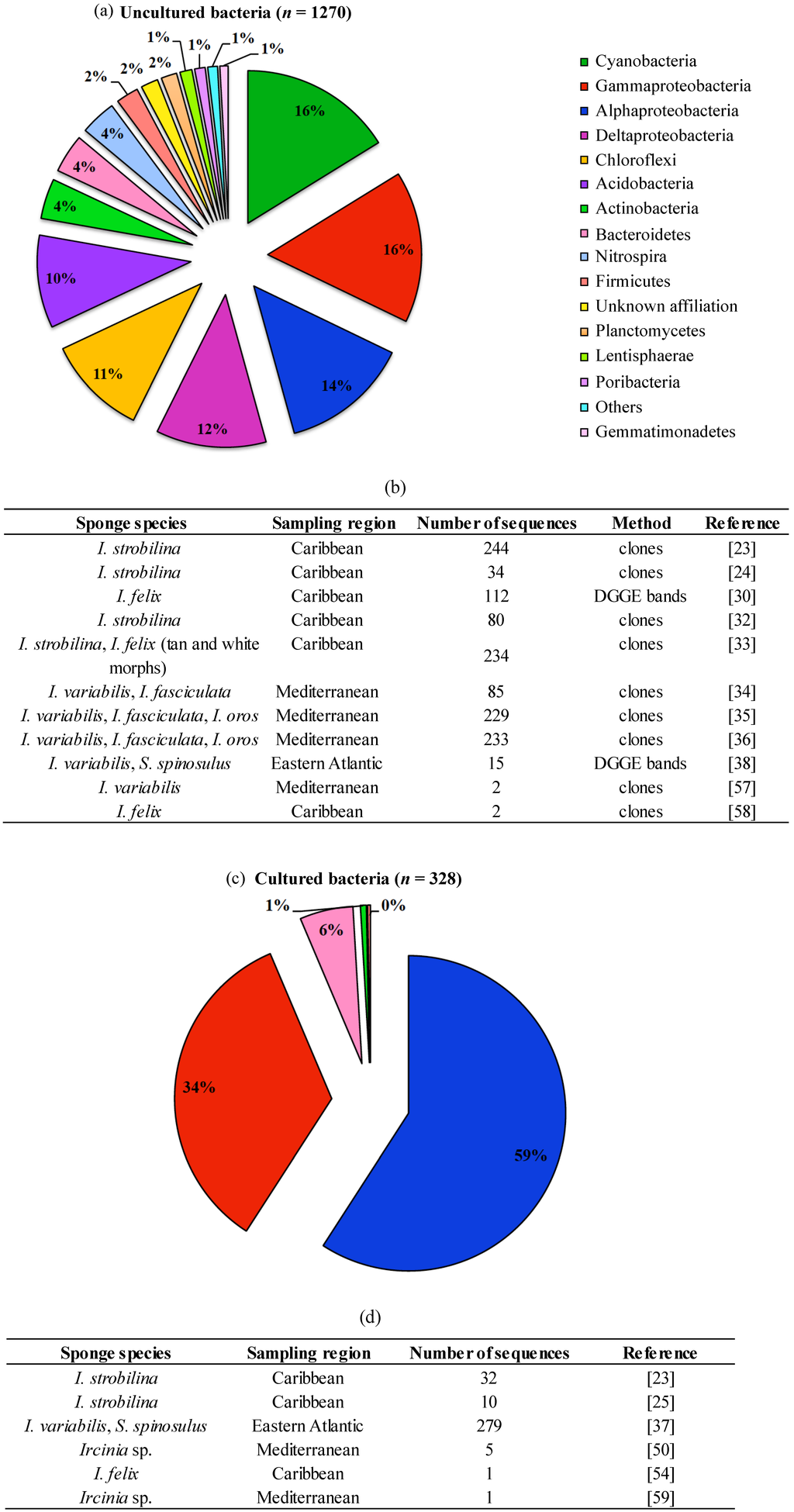
Figure 4.
Phylum-level classification of bacteria detected in Irciniidae sponges by cultivation-independent (a) and cultivation-dependent (c) 16S rRNA gene surveys, and their respective metadata (b,d); the analysis covers all sequences available at NCBI until December 2013, with the exception of 454-pyrosequencing sequence reads. The phylum Proteobacteria is represented in (a) and (c) by the classes Alpha, Gamma and Deltaproteobacteria. The total number of sequences analysed in (a) and (c) is given in brackets.
To overcome limitations common to PCR-DGGE and T-RFLP fingerprinting (i.e., impossibility to identify the entire prokaryotic community) and cloning-and-sequencing procedures (i.e., restrictions regarding library sizes), next-generation sequencing (NGS, e.g., 454-pyrosequencing) has been more often applied in recent years for the characterization of sponge symbiont diversity [20,60]. Using 454-pyrosequencing, 16 bacterial phyla and 1199 OTUs (95% gene similarity cut-off) were recovered from I. ramosa sampled from the Great Barrier Reef (Australia), whereas 30 bacterial phyla and 6800 OTUs were recovered from SSW [20]. With this technology, Schmitt and colleagues [14] uncovered 159, 179, 86 and 111 bacterial OTUs (97% cut-off) from Ircinia sp., I. felix, I. variabilis and I. gigantea (note by authors: non-valid taxon) specimens sampled from the Indian Pacific, Caribbean and Mediterranean Seas, and the Great Barrier Reef, respectively. Overall, the dominant bacterial phyla associated with Ircinia spp. were Acidobacteria, Chloroflexi, Poribacteria and Proteobacteria (Alpha and Gamma classes). Both surveys suggested that indeed the bacterial community associated with marine sponges was species-specific [14,20]. Likewise, 454-pyrosequencing was employed in recent studies covering cultivation bias, host specificity and temporal dynamics of microbial communities associated with Irciniidae species in the north Atlantic. Hardoim and colleagues [61] addressed the effects of three methods of sample handling on bacterial community composition in S. spinosulus and I. variabilis. Using cultivation-independent methods, the bacteriome of S. spinosulus and I. variabilis was indeed found to be species-specific, deepening the observations made by means of PCR-DGGE profiling [38], whereas similar bacterial assemblages were detected in both sponge species via culturing, highlighting the bias associated with describing sponge microbiome diversity based solely on cultivation attempts. These authors also localized bacterial symbionts in I. variabilis and S. spinosulus by fluorescence in situ hybridization (FISH). They observed that bacterial cells were almost exclusively detected in-between sponge cells and ususally absent on the spongin fibers that form the skeleton of Irciniidae sponges (Figure 5). In addition, Hardoim and Costa [62] unravelled the temporal dynamics of the bacterial community associated with S. spinosulus over three sucessive years. PCR-DGGE and tag-pyrosequencing showed that the bacterial assemblage was stable over time, with the majority of dominant bands and OTUs detected in all years. S. spinosulus was dominated by eight bacterial phyla, of which only two presented different relative abundances over the years. Only 27 OTUs, distributed in nine bacterial phyla, were detected in all 12 sponge specimens. Interestingly, in spite of its low diversity at the phylotype level, this minimal core of symbionts shared by all animals analysed displayed high bacterial richness at the phylum level, comparable to that observed in a single sponge hosting about 200 OTUs. This suggests that phylum diversification is fundamental to sponge functioning, and that high redundancy of phylotypes within each phylum likely aids the sponge host in maintaining its repertoire of bacterial phyla in face of local environmental changes or across different developmental stages.
In summary, the recent application of NGS has enabled a better understanding of the “species-level” bacterial diversity in irciniid sponges in comparison with previous molecular technologies. For instance, between 29 and 33 OTUs (99% cut-off) have been reported for Ircinia spp. using the cloning-and-sequencing method [34], whereas hundreds to thousands of OTUs have been recovered from irciniids via NGS [14,20,61,62], unmasking the complex microbiomes that inhabit these animals. Nevertheless, previous findings made based on less powerful tools, such as the trends for spatial and temporal stability of these symbiont assemblages, have been confirmed by deep sequencing surveys.
Besides the use of the 16S rRNA gene as a phylogenetic marker to describe sponge-associated bacterial communities, other genes have been selected to uncover putative functions from sponge symbionts, especially those involved in biogeochemical cycling [6]. Among the elements essential to life, nitrogen is one of the most limiting factors affecting ecosystem functioning [63]. The nifH gene (which encodes an iron-containing dinitrogenase reductase) has been widely used in diversity assessments of N-fixing bacteria in aquatic and terrestrial environments [64,65,66]. Mohamed and colleagues [21] determined the expression of nifH genes in I. strobilina. From the cDNA clone library made for this species, 44 nifH gene clones were retrieved. They clustered within a group formed exclusively by cyanobacterial phylotypes, indicating that Cyanobacteria symbionts were the prevailing N-fixers in I. strobilina. Mohamed and colleagues [24] also disclosed the diversity of aerobic ammonia-oxidizing bacteria (AAOB) associated with I. strobilina using cloning-and-sequencing of the amoA gene (which encodes the catalytic α-subunit of the ammonia-monooxygenase enzyme). All amoA sequences fell into two clusters affiliated with Nitrosospira (Betaproteobacteria), suggesting that they were the main nitrifiers in I. strobilina. However, no expression of the amoA gene could be detected [24]. Hardoim and Costa [62] used the amoA gene to unravel the temporal dynamics of ammonia-oxidizing bacteria (AOB) in S. spinosulus by PCR-DGGE fingerprinting. Few AOB phylotypes could be detected in the PCR-DGGE profiles of S. spinosulus over the three successive years, suggesting that the AOB community in this species was stable over time. As sponges comprise a major portion of the biomass in many coral reefs, microbial mediated nitrogen metabolism within the animal host is expected to have a major impact on the nitrogen budget of these environments [21,24]. More studies are still necessary to ascertain the activity and identity of N-cycling players in marine sponges.

Figure 5.
Confocal laser scanning microscopy images of fluorescent in situ hybridization (FISH-CLSM) stained bacteria in S. spinosulus; volume rendering images (left in each panel) and their corresponding 3D reconstructions (right in each panel) are shown for hybridizations with the Cy3-labeled universal bacterial probe (red cells) coupled to ALEXA488- or Cy5-labeled group-specific probes targeting the Alpha- and Gammaproteobacteria classes; Gammaproteobacteria cells appear as yellowish cells in the volume rendering images (a) and as green objects in the 3D reconstruction (b). For co-hybridizations including bacterial, alpha and gammaproteobacterial probes, the latter two groups are represented by yellowish and pink cells, respectively (c,d). Sponge background structure, vastly dominated by profuse spongin filaments, is shown overall in blue or cyan.
Altogether, Irciniidae species harbour complex and diverse bacteriomes. These are to a large extent maintained by the host species over seasons, years, environmental gradients and geographical regions. Through an in-depth phylogenetic assessment comprising all bacterial 16S rRNA gene sequences (excluding NGS and PCR-DGGE short sequence reads) from Irciniidae sponges publicly available until December 2013, we here substantiate the view of high spatial bacteriome stability in these sponges (Supplementary Figure S1). In fact, bacterial phylogenetic signatures common to the major oceanic regions studied so far could be found within several typical sponge-associated phyla such as the Acidobacteria, Actinobacteria, Bacteroidetes, Chloroflexi, Cyanobacteria and Alpha, Delta and Gamma classes of the Proteobacteria (Supplementary Figure S1a–i). Irciniidae species appear, moreover, to hold their own distinct bacterial communities. These differ from host to host within the same habitat and from the surrounding bacterioplankton communities found in seawater. Thus, intrinsic aspects of the host species—for instance habitat preference, specific geographic distribution and evolutionary history—may cooperatively shape the structure of bacterial symbiont communities in these sponges [34]. Several bacterial taxa associated with Irciniidae species are known for their physiological and metabolic features—of which inhibitory, primary production and nitrogen metabolism capacities can be highlighted—indicating that they play essential roles in the establishment and maintenance of the sponge host.
3.2.2. Bioactivities
Few surveys have described biological activities obtained from bacterial strains—or from their respective metabolite extracts—isolated from Irciniidae species [37,50,67,68]. Although these studies usually do not unveil the identity of the compounds underlying the observed activities, they collectively highlight a broad spectrum of functions of ecological or applied interest within the culturable sponge-associated microbiota. For instance, Thakur and colleagues [68] addressed the hypothetical importance of I. fusca-associated bacteria in host defence against bacterial epibionts. In total, 25 isolates were recovered from the sponge surface in five sampling events. Crude sponge extracts were obtained from all events, but only two were active against isolates retrieved from the same sampling. Strains identified as Micrococcus sp. and Bacillus sp. exhibited antibacterial and antifouling activities, highlighting the potential of both genera to inhibit the growth of epibiotic bacteria on the sponge surface. These results suggest that sponge-associated bacteria can modulate the growth of their own cells as well as their neighbours by producing bioactive metabolites. Esteves and colleagues [37] described the in vitro antagonistic activity of 155 distinct genotypes isolated from S. spinosulus and I. variabilis towards two relevant clinical strains: Escherichia coli NCTC 9001 (Gram-negative) and Staphylococcus aureus NCTC 6571 (Gram-positive). A few isolates (n = 18) were active against both strains, whereas 44 and 27 isolates showed antimicrobial activity towards E. coli and S. aureus, respectively. The most active genus isolated from Irciniidae species was Vibrio with 27 and 12 of the isolates inhibiting E. coli and S. aureus, respectively. From S. fasciculatus collected from the Bay of Bengal (India), five Firmicutes isolates and one Proteobacteria isolate inhibited β-glucosidase activity [69]. Glucosidase inhibitors may be of use in the treatment of viral infections, cancer and genetic disorders [70]. These studies emphasized the biotechnological potential of the culturable bacterial fraction of Irciniidae sponges, encouraging more research into the mechanisms and compounds underpinning such activities.
3.2.3. Bioactive Compounds
Some studies identified the bioactive compounds extracted from Irciniidae-associated bacteria. Several cyclic peptides have been retrieved from Bacillus and Staphylococcus strains isolated from I. variabilis from the Bay of Naples, Italy [71]. Of these, the compound cyclo-(l-prolyl-l-tyrosine) could modulate the activity of the LuxR-based quorum sensing system of bacteria [72], whereas all other compounds investigated presented structural similarities to compounds known to interact with LuxR-based biosensors [72,73]. Thus, such cyclic peptides may play a role in the marine sponge microbiome as quorum sensing signal molecules [71]. Furthermore, a Bacillus pumilus strain was isolated from Ircinia sp. and found to produce five surfactin-like substances (cyclic acyldepsipeptides) called bacircines [74,75]. These compounds showed cytotoxic effects on the development of sea urchin eggs. One exhibited antitumour activity against Ehrlich ascites carcinoma and anti-HIV activity [74,75]. These studies demonstrate that the functional screening of compounds extracted from sponge-associated bacteria is a promising, direct route to manifold interesting bioactivities, which nevertheless remains underexplored.
3.3. Archaea
In comparison with the multi-layered molecular research undertaken in the past decade on sponge-associated bacteria, fewer surveys have addressed archaeal community composition and diversity in marine sponges. Overall, these studies uncovered less diverse archaeal assemblages from sponges than the corresponding bacterial communities [6,8,76].
3.3.1. Diversity
Phylogenetic inference performed with three distinct archaeal 16S rRNA gene sequences obtained from Sarcotragus sp. (Jeju Island, Korea) revealed that they were affiliated to marine group I Crenarchaeota (now known as Thaumarchaeota, see [77,78,79]). Two of them grouped within clusters containing only sponge-derived sequences, but without bootstrap support [80]. Further analysis showed that one of these sequences indeed belonged to a Thaumarchaeota “sponge-specific” cluster (SC175) [76].
Using PCR-DGGE and 454-pyrosequencing, we demonstrated that the archaeal community associated with S. spinosulus was stable over three successive years. In 11 out of 12 S. spinosulus specimens, one single OTU affiliated to Nitrosopumilus dominated the archaeal assemblage, whereas the remaining specimen was dominated by Cenarchaeum sp. Candidatus Nitrosopumilus maritimus and Cenarchaeum sp. are known ammonia oxidizers. This indicates that the replacement of the dominant archaeal symbiont observed for one S. spinosulus individual likely does not interfere with the ammonia-oxidation potential in this host [62]. Overall, the diversity of archaea associated with other sponge species was as well encompassed by only few phylotypes [6,76,81,82]. Despite their low diversity, it appears that Archaea, and not Bacteria, are the prevalent players in the process of ammonia-oxidation in marine sponges [83,84,85]. It still needs to be determined which other possible roles, if any, archaeal symbionts play in association with Irciniidae and marine sponges as a whole.
3.3.2. Bioactivities and Bioactive Compounds
Neither in vitro bioactivities of archaeal cultures and their metabolite extracts nor identified bioactive compounds from sponge-associated Archaea were found in our literature survey.
3.4. Fungi
As pointed out by Webster and Taylor [8], the study of the diversity and functioning of sponge-derived fungi still requires more effort. Fungi have been approached primarily in a cultivation-dependent manner, probably because of their higher amenability to laboratory domestication than that displayed by bacteria and archaea. Whereas this facilitates functional screenings for manifold activities and full genome exploration of the obtained pure cultures, it is felt that comprehensive analyses of fungal diversity and maintenance across temporal and spatial scales could benefit more from dedicated cultivation-independent studies.
3.4.1. Diversity
The diversity of fungi associated with I. oros and I. variabilis collected at Malta was assessed using several media and incubation for over 30 days at room temperature (~20 °C) [86]. Both sponge species were dominated by Aspergillus spp. corresponding to 39.4% and 50%, respectively, of the total isolates followed by Penicillium spp. (13.6% and 27%). Whereas 13 and 10 fungal genera were found in I. oros and I. variabilis, respectively, only five of them were shared by both [86]. The diversity of culturable fungi was also assessed in Psammocinia sp. (Sedot-Yam, Israel) [87]. Potato dextrose medium amended with fungicides was used in two approaches: direct plating of inner sponge fragments and direct plating of sponge extracts (“sponge-compressed” method). The Petri dishes were incubated at 25 °C in the dark from three to 30 days. Sequencing of the internal transcribed spacer (ITS) of rRNA genes was used to identify the fungal strains. Overall, 220 pure cultures were isolated and assigned to 85 fungal taxa, of which the majority was obtained using the sponge-compressed method (n = 76). Most of the strains (94%) were identified as Ascomycota, whereas Basidiomycota (4%) and Glomeromycota (2%) were minor components of the cultured fungal community. From Ascomycota, 80 “species-level” taxa were recovered and among them 25 were identified as Eurotiales (Aspergillus and Penicillium spp.), five as Capnodiales (Cladosporium spp.), 15 as Pleosporales (Bionectria, Fusarium, Phoma, and Preusia spp.) and 35 taxa as Hypocreales (Acremonium and Trichoderma spp.). When sponge-compressed samples were plated onto medium amended with different fungicides, further 28 taxa that were not obtained with the previous approach could be isolated, thus enabling an increase in the diversity of fungi cultured from the sponge. Using a coverage-based algorithm, the authors estimated that only 15% of the fungi associated with Psammocinia sp. could be recovered [87]. Notably, even though both studies used distinct approaches, the fungal diversity obtained from I. oros, I. variabilis [86] and Psammocinia sp. [87] was alike.
3.4.2. Bioactivities
Accompanying their assessment of fungal diversity, Paz and colleagues [87] also investigated the inhibitory activities of fungi isolated from Psammocinia sp. using a diffusion bioassay. Thirty-six fungal cultures showed in vitro antagonism against at least one of the test fungi (Alternaria alternata, Rhizoctonia solani and Neurospora crassafour) or against the oomycete Pythium aphanidermatum. Some isolates (Trichoderma, Acremonium, Bionectria, Verticillium, Penicillium and Aspergillus spp.) were able to secret inhibitory compounds into the growth medium, although those were not identified. It was demonstrated that distinct Trichoderma spp. isolated from Psammocinia sp. were capable to mycoparasite Fusarium equiseti isolated from the same sponge species [87].
3.4.3. Bioactive Compounds
Penicillium chrysogenum was isolated from the Mediterranean I. fasciculata collected in the Bight of Fetovaia (Italy) and was shown to produce the sorbicillinoid alkaloids sorbicillactones A and B [88]. Sorbicillactone A exhibited remarkable cytotoxic activity towards murine leukaemia lymphoblast (L5178y) cells and anti-HIV activity. This compound is also a promising neuroprotective metabolite. Three other sorbicillinoid compounds (oxosorbicillinol, sorbicillin and bisvertinolone) were identified from P. chrysogenum, along with the alkaloids meleagrine and roquefortine C [88]. The isolate Microascus sp. K14 from I. variabilis also produced the anti-fungal compound fungerin [86].
Irciniids harbour a complex, readily culturable fungal diversity. The potential for uncovering novel fungal-derived bioactive compounds from sponges will become more apparent with the improvement of cultivation and screening techniques and/or the accomplishment of in-depth molecular surveys such as whole genome sequencing of fungal symbionts. This will also improve our understanding of the roles played by sponge-associated fungi within the host.
3.5. Other Microeukaryotes
Although diatoms and dinoflagellates have been detected in marine sponges [6], no study involved irciniids. There is thus a need to unravel microeukaryote taxonomic, functional and metabolic diversities within sponges as a first step in the pursuit of their biotechnological potential.
In summary, much of the microbial diversity research undertaken so far, considering both marine sponges as a whole and the family Irciniidae, has focused on bacterial communities. Such an interest may reflect the higher relevance of this particular group of symbionts in host fitness. Nevertheless, more effort to unveil the spatial-temporal diversity, composition and abundance of archaeal and microeukaryote assemblages—and their corresponding biological activities—in marine sponges is needed for a balanced picture of the marine sponge holobiont. Within the family Irciniidae, more attention to the genus Ircinia was found. Attempts to approach the microbial ecology of S. spinosulus have been published only recently [37,38,62], and the highly selective and conserved character of its microbiome makes it a valuable species for comparative studies with other irciniids in the Atlanto-Mediterranean zone and beyond. Besides the recent study by Paz and colleagues on fungal diversity and inhibitory activities [87], very limited information exists on the microbial ecology of Psammocinia spp., in spite of their distinct geographical range (Figure 1) and manifold bioactivities (see Supplementary Information). Collectively, approaching the microbiology of these three genera might constitute a rewarding strategy to understanding the microbiology of marine sponges on a global scale with a robust, underlying comparative framework.
4. Vertical Transmission of Sponge Symbionts
Vertical transmission is an important mechanism by which sponge-specific symbionts are passed from the parental to the next generation via larvae or gametes. It has been investigated in several sponges and, when present, has usually been interpreted as evidence for an intimate pattern of relationship between host and the transmitted symbionts [30,52,58,89,90,91].
Vertical transmission was proposed as a mode of symbiont inheritance in I. felix through the detection of bacteria in adult, larval and juvenile stages of the sponge [30]. This study showed, via TEM, that adults contained large and complex bacterial communities, and that high bacterial abundance was also observed extracellularly in the inner region of the larvae, whereas the outer region was almost free of microorganisms. When the host entered the juvenile stage, bacterial cells were found primarily in the mesohyl. Bacterial 16S rRNA gene sequences obtained from I. felix adults and offspring (larvae and/or juvenile) belonged to the phyla Acidobacteria, Chloroflexi, Gemmatimonadetes and Proteobacteria (Alpha, Delta, and Gamma classes), and to one lineage of uncertain affiliation. In a subsequent study, Schmitt and colleagues [58] detected several vertically transmitted phylogenetic clusters (VT-clusters), composed by sequences obtained from I. felix adults and offspring. VT-clusters affiliated with the Acidobacteria, Chloroflexi, Gemmatimonadetes, Nitrospira and Proteobacteria phyla, suggesting vertical transmission as a mechanism through which a complex bacterial consortium can be formed and maintained within I. felix. We still do not know what their function may be in the larvae, nor in settlement, which may occur from a few minutes to several hours after release [92]. Once settled, sponge larvae undergo a rapid metamorphosis to an early juvenile stage. Thus, bacterial symbionts might protect the juvenile from predators by producing antibiotics and deterrent compounds. Arguably, the best example of such an ecological function was obtained by Lopanik and colleagues [93,94,95,96] and Sudek and colleagues [97] while studying the association between the bacterial symbiont Candidatus Endobugula sertula and the bryozoan Bugula neritina. Candidatus E. sertula was found to produce the bioactive polyketides bryostatins. These protected the larvae against predators and were observed in all life stages of B. neritina. Bacterial symbionts in sponge larvae may also perform important housekeeping functions or constitute a source of food, since the larvae are unable to take up food particles from seawater [98].
Even though Schmitt and colleagues [58] also described two archaeal VT-clusters from the marine sponges Agelas conifera and Luffariella variabilis, there is to date no specific documentation of archaeal vertical transmission in Irciniidae sponges, although its existence can be presumed based on available data. Our understanding of fungal acquisition by marine sponges remains otherwise very limited, with neither dedicated studies of this group in sponge larvae or juveniles nor an existing conceptual model on how sponges recruit and maintain these symbionts.
5. Bacterial Communication and Signalling Molecules
Quorum sensing (QS) is a mechanism by which microorganisms monitor and regulate their population size through chemical signalling [99]. When quorum-sensing molecules (QSM) released by microorganisms into the extracellular environment reach a threshold concentration, they initiate a signalling transduction cascade that regulates the expression of several target genes in an orchestrated response to the prevailing conditions. QS is a complex mechanism involved in bacterial virulence, swarming motility, conjugal plasmid transfer, biofilm maturation, and antibiotic production and resistance [99]. As such, it affects several activities or areas relevant to human health and the environment (e.g., bacterial multidrug resistance, aquaculture, water purification). These might be impacted by negative feedbacks ensued from QS and its mediation of bacterial metabolism [99,100]. Solutions to QS-derived drawbacks may arise from the activities of the symbiont communities themselves. For instance, from the marine sponge Luffariella variabilis, the compounds manoalide, secomanolide and manoalide monoacetate were obtained and showed to be important QS inhibitors [101]. Furthermore, the alkaloids ageliferin and mauritamide B obtained from Agelas conifera and A. nakamurai, respectively, inhibited the QS of the bacterial reporter Chromobacterium violaceum CV017, whereas seven compounds isolated from distinct marine sponges inhibited the QS of C. violaceum CV017 and showed antibiotic properties [102].
One of the most studied groups of QSM, N-acyl homoserine lactones (AHLs), was investigated in I. strobilina inhabiting shallow waters in Key Largo, Florida [22]. Two Alphaproteobacteria and three Gammaproteobacteria isolates were found to produce AHLs, indicating that QS systems play an important role in sponge microbiome dynamics as observed for other systems. The sponge mesohyl constitutes a nutrient rich environment when compared to oligotrophic seawater, thus colonizing bacteria may reach high densities within this microenviroment, allowing them to perform activities mediated by quorum sensing.
Autoinducer-2 (AI-2) is one well-characterized molecule involved in bacterial interspecies communication [99,100]. Briefly, during AI-2 biosynthesis, S-adenosylmethionine is transformed into 4,5-dihydroxy-2,3-pentanedione (DPD) by a sequence of three enzymatic reactions, where the last is catalyzed by the enzyme LuxS. Since DPD is unstable, it naturally cyclizes, generating a range of furanones in the presence of water. Vibrio harveyi, for example, detects the borate diester form of AI-2 using the LuxP/LuxQ signaling cascade, which may be exclusive to Vibrio organisms. AI-2 is associated with a broad range of functions; for instance, virulence in V. vulnificus and V. cholera, type III secretion, protease production, luminescence, colony morphology and siderophore production in V. harveyi [99]. Several surveys detected Vibrio spp. associated with different marine sponges [37,49,50,51,103,104,105,106], indicating that they may be capable of performing an array of functions within the sponge host. From I. strobilina (Key Largo, USA), 10 out of 40 isolates were identified as Vibrio spp. based on 16S rRNA gene sequencing [25] and found to be closely related to V. harveyi (n = 5) and V. campbelli (n = 4). Notably, all 10 Vibrio isolates were capable of synthesizing AI-2 molecules. Furthermore, 30 distinct Vibrio luxS gene sequences were identified, of which 28 were closely related to the luxS gene of V. harveyi and the other two were affiliated with the luxS gene of V. parahaemolyticus. Interestingly, two luxS gene sequences were distantly related to any known Vibrio luxS sequence. These were proposed to represent a sponge-specific luxS cluster [25].
Two of the five known classes of QSM have already been detected in the bacterial community associated with I. strobilina, and more may be found as research advances. Considering the complexity of the Irciniidae microbiome, future investigations involving, for example, full genome sequencing of symbionts and deep sponge metagenomic mining (Figure 6) for QS regulatory genes may result in the discovery of other classes of QSM. It is too early to determine the precise role(s) of QS in gene regulation and metabolism of both symbionts and host. The application potential of QS regulation can be nevertheless exemplified from other host-microbe associations in the marine realm. For instance, Shewanella sp. strain MIB010, isolated from the intestine of the ayu fish, is an effective agent against the QS-regulated biofilm formed by the fish pathogen V. anguillarum [100].
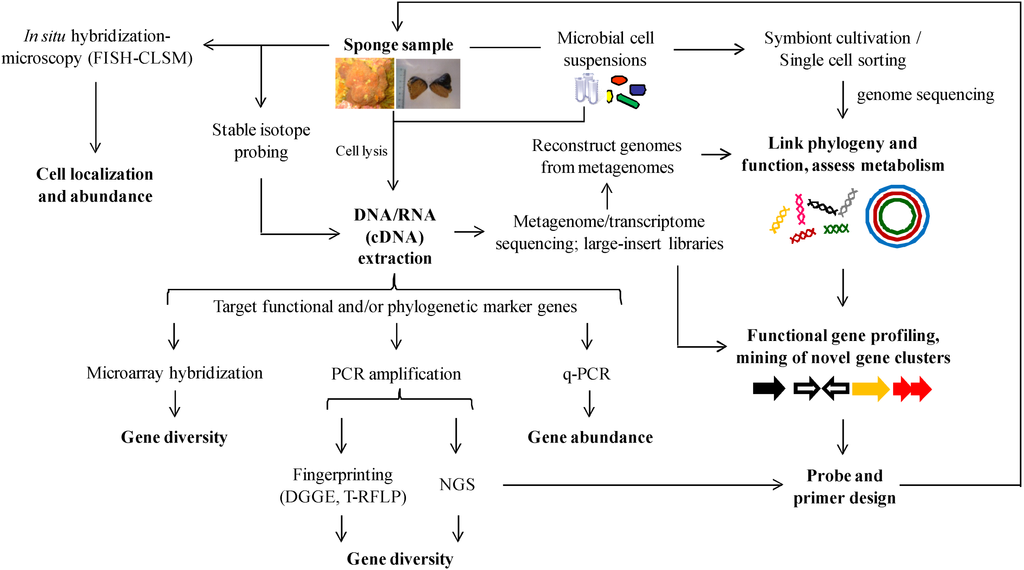
Figure 6.
Integrated molecular approach for the study of symbiont communities in Irciniidae sponges.
6. Diseases Affecting Irciniids
Many studies have shown that microorganisms might also cause disease in marine sponges [107]. In 1884, a disease affected Indian Ocean Ircinia spp., whereby fungal filaments were shown to destroy the sponge body, leaving only the hard spongin fibres [107]. In addition, the abundance of I. variabilis in the benthic community of the Marsala Lagoon (Italy) decreased from 6.9% in 1986 to 3.0% in 1989, due to a disease characterized by white patches on the surface of affected individuals [108]. Several Ircinia specimens from Guigalatupo, Panamá, were lost over five censuses performed from 1984 to 1998, whereby spreading lesions were seen in I. felix, I. campana and I. strobilina [109]. A mass mortality event was reported in 1994 affecting S. spinosulus and Ircinia sp. in the Mediterranean Sea, characterized by the decay of their spongin filaments [107]. Four more recent disease outbreaks affected Irciniidae species in the Mediterranean, Ionian and Adriatic Seas [59,110,111,112], eventually with severe consequences for the infected populations [111]. Whereas loss of photosynthetic cyanobacteria was documented among diseased specimens in one case [111], higher abundance of Vibrio spp. was observed in another [59]. In one study [110], a twisted rod bacterium was proposed as the etiological agent because it penetrated the sponge body, proliferated, and outcompeted a large range of bacterial symbionts. Shifts in symbiont communities towards a more virulent state because of elevated seawater temperature have been hypothesized to trigger the mass mortality events [59,110,111,112]. In this context, Pita and colleagues [113] subjected I. fasciculata and I. oros to aquaria experiments under high temperature (25 °C), food shortage (0.1 μm-filtered seawater) and both factors in combination. T-RFLP analysis showed that the sponge-associated bacterial communities did not change after acclimation time and during the next three weeks of experiment. TEM revealed that the ectosome of I. fasciculata was dominated by Candidatus S. spongiarum. Under high temperature and food shortage, both healthy and damaged cyanobacterial cells were observed. For I. oros under all treatments, no signs of sponge or bacterial cell degradation or cyanobacterial cells were detected. Thus, high temperature and food shortage up to three weeks did not disrupt the bacterial communities associated with both sponge hosts. The authors suggested that these factors could not alone explain the mass mortality events registered for Ircinia spp. in the Mediterranean Sea [113].
To date, few surveys have identified bacterial species as the etiological agents of disease in sponges [59,107]. More often, a clear cause-effect relationship between agent and disease is difficult to achieve [107,114]. More research is needed to improve our capability of identifying etiological agents and unravelling how biotic and abiotic features influence marine sponge disease incidence in a changing climate.
7. Experimental Microbial Ecology
Sponge microbiology is a fertile field for experimentation. Most of the first, experimental sponge microbial ecology studies kept the host animals for varying periods in aquaria, and tested whether the structure and function of their associated bacterial communities shifted during the maintenance periods [23,115,116]. Surveys using other sponge hosts reported changes in bacterial community composition of specimens kept in aquaria over six months and two years [115,116]. In contrast, evidence exists of recovery within Irciniidae symbiont communities back to the in-situ stage after some time in captivity [23]. Specifically, Mohamed and colleagues applied molecular techniques to monitor bacterial communities associated with I. strobilina maintained in aquaculture [23]. Thirty-five OTUs were retrieved from wild specimens, while 48 and 47 OTUs were recovered from specimens kept in aquaria for three or nine months, respectively. Overall, these OTUs were affiliated to five or six bacterial phyla depending on the category (wild, 3- and 9-month sponges). Although the highest diversity was observed for the 3-month sponges, 9-month sponges showed an intermediate diversity between wild and 3-month sponges, which suggested acclimatization of the holobiont to the aquaculture system. From the sponges maintained in aquaculture for three and nine months, chemical fingerprints of small molecules including primary and secondary metabolites extracted with ethanol were analysed by liquid chromatography-mass spectrometry and compared to the wild type. The analyses revealed no major changes in the natural product profile of I. strobilina upon transfer to aquaculture [23]. This suggests that, although the bacterial community from aquarium-maintained I. strobilina differed from those of the wild specimens along the assay, the major functions were retained. This could be explained by the functional redundancy of sponge bacterial symbionts [117,118]. So far, no study approached sponge-associated archaeal or fungal communities during maintenance in aquaria.
It is still to be determined whether sponge microbial communities can be successfully stabilized in aquaculture, and longer surveys may shed more light. Technical problems in maintaining sponges in captivity have hindered experimental progress, which is fundamental to elucidate how the marine sponge microbiome might be affected by e.g., temperature increase [119,120], water acidification [121] or invasive pathogens (see Section 6 above). However, Ircinia spp. have shown amenability to experimental handling [23,113], a feature that reinforces their suitability as a model taxon in microbiology and biochemistry studies.
8. Metagenomics-Based Discovery of Secondary Metabolites Biosynthetic Gene Clusters
Polyketides are usually found in sponges and comprise a structurally varied class of compounds that have attracted considerable attention due to their highly potent cytotoxicity [122]. Polyketide synthases (PKS) are multifunctional enzymes of typical microbial origin that catalyse polyketides biosynthesis. However, the complexity of the bacterial assemblages associated with marine sponges makes identification of the actual PKS producers difficult. Some techniques have been applied to overcome these limitations; for instance, cloning-and-sequencing, genome, single-cell genome and metagenome mining of PKS encoding operons (Figure 6). Targeting the biosynthetic gene clusters, two pederin-type PKS systems putatively involved in the synthesis of antitumour polyketides were located in the metagenomic DNA of Theonella swinhoei [123]. Likewise, metagenomic libraries generated from cell fractions enriched for filamentous bacteria associated with Discodermia dissoluta revealed that they consisted of several non-ribosomal synthase (NRPS) as well as mixed PKS-NRPS gene clusters [124]. Moreover, genomic mining of two bacterial cells sorted from Aplysina aerophoba and belonging to the phyla Poribacteria and Chloroflexi revealed Sup-PKS (“sponge symbionts ubiquitous PKS gene”) and NRPS gene clusters in the screened genomes, respectively [15]. Single-cell genomics further uncovered at least two PKSs, one of which affiliated with the sponge-specific “Sup-type PKS” from a Poribacteria cell sourced from A. aerophoba [125]. Collectively, these studies demonstrate the wide applicability of metagenomics to studying the function and bioactive potential of marine sponge symbionts.
Within Irciniidae sponges, psymberin was isolated from Psammocinia aff. bulbosa collected at Milne Bay (Papua New Guinea) [126]. The complete sequencing of three fosmids revealed common features of bacterial genomic architecture: no introns were found, the genes were preceded by Shine-Dalgarno sequences, the space between genes suggesting that the transcribed mRNA was polycistronic and the close relationship to genes exclusively from bacteria. Nevertheless, the low similarity of these genetic signatures to genes from Pseudomonas sp. or other prokaryotes hampered the correct identification of the bacterial producer. Finally, a strong correlation among bacterial abundance, the presence of Poribacteria and Sup-PKS was observed in the Caribbean I. felix, and it was proposed that Poribacteria was most likely the producer of mid-chain-branched fatty acids (MBFAs) from this sponge [127].
9. Cultivation of Irciniidae Species
Even though marine sponges produce the largest variety of secondary metabolites in marine ecosystems, few compounds have reached commercial production. This is because of the normally minute amounts readily found in the sponge body and of the impossibility to collect sufficiently large quantities of sponge biomass from their natural habitats, which is intolerable given the foreseen impacts on the marine environment and natural sponge populations. To circumvent this, many techniques have been developed to cultivate sponge species [6,128,129,130]. Below we highlight attempts to sponge cultivation using Irciniidae. We divide these attempts into three modalities: sponge mariculture, cell culture and larviculture.
9.1. Sponge Mariculture
In a preliminary study, Wilkinson and Vacelet [131] failed to transplant I. variabilis collected from Endoume (near Marseille) at 3–4 m from well-shaded cave walls and rock cliffs to a 7 m deep rock shelf around 30 m from the sampling place. All specimens died a few days after sampling, most likely due to severe damage caused by sampling, cutting and sewing of I. variabilis into plastic plaques.
Duckworth and colleagues [132] examined the feasibility of cultivating Psammocinia hawere in situ. Fifty specimens of P. hawere were collected from 10–20 m at “Ti Point Reef” (New Zealand). They were cut into cubes of four different sizes and transplanted to two locations, along with small whole sponges as undamaged controls. Three depths (5, 10 and 17 m) and two culture systems were used, and the experiment was conducted in summer (82 days) and winter (88 days). The survivorship was considered high (276 out of 360 explants), even though 2/3 of the explants (226) lost weight. P. hawere explants transported to deeper (10 and 17 m) waters in winter showed the highest growth and survivorship rates. This was related to the lower UV radiation as well as the cooler water temperatures, which helped to accelerate pinacoderm healing. P. hawere is known to incorporate detritus from the sediment into its body or fibres, which may have enhanced the surface consolidation during the healing process. Growth and survivorship improved with the size of the explant and the proportion of the intact pinacoderm. Independently of the culture systems used, P. hawere displayed better growth and survivorship at 10 and 17 m than at 5 m.
Van Treeck and colleagues [133] sampled 52 and 100 specimens of I. variabilis at the north-western coast of Corsica in summer and spring, respectively. They were transplanted to naturally formed sand patches in-between seagrass meadows at 15 m. Prior to transplanting, the sponge specimens were cut into different sizes underwater. The explants were placed between two frames, closed, mounted and transported to the experimental sites. The survival rates of I. variabilis were high within the first six months, followed by a reduction of 25% in population size, most likely due to a strong storm, after which the remaining sponge continued to grow. The specimens stocked in spring survived better. In the first 12 months, I. variabilis biomass increased by 90%; however, there was great variation between maximum and minimum rates (220 and −43%, respectively) [133].
In another study [134], I. ramosa was collected from Bone Lola reef (Indonesia) and cut in situ into explant sizes of around 30 mm where at least one side contained exopinacoderm. A polyethylene rope was passed through the sponge body, each rope carrying around nine explants. Ropes were attached to PVC frames that were placed horizontally ~20 cm above the reef bottom between 12–15 m deep, and secured with iron pegs at the exposed side of the submerged reef. A high survival rate of 92% was observed for I. ramosa during the four months. A slight but significant increase in length was detected, and a change in shape was observed which could not be precisely measured. However, only few explants grew during the trials [134].
9.2. Sponge Cell Culture
In a series of studies, de Rosa and colleagues [135,136,137] meticulously monitored the chemical composition of I. muscarum cells grown on a simple culture medium. Cell suspensions from I. muscarum (Gulf of Naples, Italy) were resuspended in sterile seawater complemented with antibiotics during the first two weeks of cultivation. The cultures were incubated at 18 °C in the dark and at 22 °C in the light and every third day the medium was renewed. From the third week onward, cultures were kept in Dulbecco’s modified Eagle’s medium supplemented with glucose. This medium was again renewed every third day. One week after inoculation, cells were observed attached to the bottom of culture chambers along with some aggregates. Cells were actively dividing in both culture conditions and stationary phase was reached after four days. Overall, 17 sterols were recovered. Around 90% of the total free sterols in cultured I. muscarum were represented by Δ5−7 sterols; from which 7-dehydrocholesterol, ergosterol and 7-dehydrositosterol prevailed. The medium was then supplemented with water-soluble cholesterol, which increased the number of observed cells by 70%. Oleic acid was one of the main acids in the lipids of the intact sponge and was found in low concentration in cultured cells. When the media were supplemented with oleic and linoleic water-soluble fatty acids, no growth promotion was observed. However, they were completely metabolized by the cells. The concentration of total lipids was higher in the intact animal, and cells developed in the dark had higher total lipids than those grown under light. Several structurally distinct groups of volatile compounds were observed, and their concentrations indicated that bacteria (from diet or symbionts) could be involved in their formation. A number of free amino acids were detected in the intact sponge, whereas practically none was found in the cell cultures. The major difference in carbohydrate composition between intact and cultured sponge cells was the presence of γ-lactones of 3-desoxy-arabino-hexonic acid and 3-desoxy-ribohexonic acids in higher concentrations in the cell cultures, suggesting shifts in their carbohydrate metabolism. The cultured cells did not produce any secondary metabolites as the major cell energy supplies had likely been allocated to the biosynthesis of primary compounds [135,136,137].
9.3. Sponge Larviculture
With the aim of culturing sponges from larvae, de Caralt and colleagues [138] transported mature individuals of I. oros sampled from I’Escala (Mediterranean Sea) to an open aquaculture system, where they released larvae after about 45 days. Swimming larvae were then collected and transferred to six-multi-well dishes. These were placed in aquaria with filtered seawater (0.7 µm pore diameter) at the same temperature of the field (20 °C). Settlement of the larvae on the six-multi-well dish bottom started after 24 h and larvae metamorphosed into juveniles after five to seven days. An exhalant tube should be formed after this period; however, no visible inhalant/exhalant orifices were observed among the settlers of I. oros and, at this point, most juveniles died before the skeletal fibres could be formed. Although settlement success was considered high (94.5%), no survival of I. oros specimens was observed after 50 days of experiment. Success in settlement could be related to favourable conditions such as still water and no substrate competition found by the larvae in the laboratory [138]. The first two weeks of culture showed the maximum increase in area, but this was primarily due to rearrangement of the biomass during metamorphosis, rather than true growth [138]. Therefore, unknown and specific environmental cues are likely needed to enable full development of sponge larvae into mature individuals in the laboratory.
From the three modalities approached above, for Irciniidae species mariculture is apparently the most feasible (host) culturing methodology, rather than cell or larvae cultivation. However, whether irciniids will produce secondary metabolites under these conditions remains to be tested. Osinga and colleagues [128] reported on five sponge species that, grown in mariculture, were able to keep the biosynthesis of target metabolites, demonstrating that this approach can be rewarding. Optimization of explant size, depth of transplantation, temperature ranges, and exposure to light and water currents might improve sponge productivity and bioactive compound availability. Besides mariculture, ex-situ culture (i.e., aquaculture) of adult specimens and semi-synthesis of metabolites are promising alternatives. In the former, the sponge grows under controlled conditions in flow-through or recirculation systems, and the experimental costs and yields of bioactive compounds would be comparable to mariculture. In the latter, a biosynthetic precursor produced by a genetically modified bacterium triggers a limited number of further chemical reactions to the synthesis of the final product. For instance, using the antibiotic cyanosafracin B, which was obtained from Pseudomonas fluorescens via bacterial fermentation, an analogue of the antitumor compound ecteinascidin 743 isolated from the tunicate Ecteinascidia turbinata was effectively synthesized [130]. Combining biological and chemical approaches might be an effective way to cultivate sponges and recover their secondary metabolites.
10. Concluding Remarks and Outlook
Irciniidae-microbe interactions are pivotal to host health and have been evolving since the hosts’ origins. In recent years, a few fundamental issues regarding these interactions have been successfully addressed. For instance, we now know that distinct bacterial symbionts are selectively enriched within the sponge body and, eventually, vertically transmitted, and that such symbiont communities are stable along time and across geographical regions. Nevertheless, many other questions remain unanswered. How full symbiont genomes evolve across space and time, both within individuals of the same species and between different host species? Do single gene-based analyses (e.g., 16S rRNA gene sequencing) suffice to completely determine microbial diversification and evolution in marine sponges? What are the functions played by the symbionts in host health and functioning, and which symbiont does what? How do the vertically transmitted microorganisms affect larval settlement and juvenile establishment? What are the benefits of quorum sensing for the host? Are secondary metabolites produced by symbionts effectively involved in host’s chemical defence? Is there a true etiological agent (or many) underpinning disease outbreaks in Irciniidae species? Is the existent technology for their laboratory maintenance robust and widely applicable? Furthermore, the majority of microbiology studies of Irciniidae species have so far mainly determined bacterial community structures in these hosts. Although useful in shedding new light on our understanding of the diversity and stability of bacterial assemblages in marine sponges, these inventories usually do not unravel the biotechnological potential of the bacterial partners. Other microbial associates, such as archaea, fungi and other microeukaryotes—and their biotechnological value—have been left almost unaddressed. Thus, only by integrating the full holobiome in future surveys will a more comprehensive picture emerge, allowing better understanding of host function. Currently, several advanced molecular approaches such as metatranscriptomics, metaproteogenomics and single-cell genomics, may help answer these questions (Figure 6). Indeed, their recent application has already been useful in disentangling the phylogeny-function dilemma, and in unveiling the life-style and secondary metabolite biosynthesis capacity of several, so-far uncultured, marine sponge symbionts [15,125,139,140,141]. Finally, few alternative culturing protocols have been attempted so far for marine sponge microbial associates. The spectrum of microorganisms cultured from these hosts is likely to grow in the next few years as researchers develop dedicated protocols to the capture of “hard-to-culture” symbionts. In this regard, ever-increasing genetic datasets retrieved via cultivation-independent methods can help in the identification of symbiont metabolic pathways and physiological demands that could be explored in future cultivation trials. Given the aforementioned challenges, a joint interdisciplinary effort is clearly needed, from fields ranging from microbiology to chemistry, and from genetics to zoology, to advance the knowledge of the oldest extant metazoan-microbe interaction system on Earth. Ideally, this would couple laboratory experimentation with integrative systems biology strategies; therefore, we reinforce the need to select marine sponge models, and provide a multitude of reasons in favour of the Irciniidae family as one such model.
Supplementary Files
Supplementary File 1Supplementary File 2Acknowledgments
This research was financed by the Portuguese Foundation for Science and Technology (FCT) through the research projects PTDC/MAR/101431/2008 and PTDC/BIA-MIC/3865/2012, granted to RC. It was further partially supported by the European Regional Development Fund (ERDF) through the Operational Competitiveness Programme (COMPETE) and by national funds through FCT (Foundation for Science and Technology), under the project “PEst-C/MAR/LA0015/2011”. CCPH was the recipient of a PhD fellowship granted by FCT (Grant No. SFRH/BD/60873/2009). Massimiliano Cardinale is thanked for his guidance in FISH-CLSM analyses. We acknowledge Gobius Ciência e Comunicação (Portugal) for the artwork designs in Figure 1 and Figure 3.
Author Contributions
Cristiane C. P. Hardoim and Rodrigo Costa conceived and designed the study. Cristiane C. P. Hardoim collected the literature and analyzed the data. Cristiane C. P. Hardoim and Rodrigo Costa wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, C.W.; Chen, J.Y.; Hua, T.E. Precambrian sponges with cellular structures. Science 1998, 279, 879–882. [Google Scholar] [PubMed]
- Van Soest, R.W.M.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; de Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.A. Global diversity of sponges (Porifera). PLoS One 2012, 7, e35105. [Google Scholar] [PubMed]
- Love, G.D.; Grosjean, E.; Stalvies, C.; Fike, D.A.; Grotzinger, J.P.; Bradley, A.S.; Kelly, A.E.; Bhatia, M.; Meredith, W.; Snape, C.E.; et al. Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature 2009, 457, 718–722. [Google Scholar]
- Vacelet, J.; Donadey, C. Electron-microscope study of association between some sponges and Bacteria. J. Exp. Mar. Biol. Ecol. 1977, 30, 301–314. [Google Scholar]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol. 2012, 10, 641–675. [Google Scholar] [PubMed]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [PubMed]
- Spalding, M.D.; Fox, H.E.; Halpern, B.S.; McManus, M.A.; Molnar, J.; Allen, G.R.; Davidson, N.; Jorge, Z.A.; Lombana, A.L.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 2007, 57, 573–583. [Google Scholar]
- Webster, N.S.; Taylor, M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [PubMed]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association: A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [PubMed]
- Taylor, M.W.; Hill, R.T.; Hentschel, U. Meeting report: 1st International Symposium on Sponge Microbiology. Mar. Biotechnol. 2011, 13, 1057–1061. [Google Scholar] [PubMed]
- Bayer, K.; Scheuermayer, M.; Fieseler, L.; Hentschel, U. Genomic mining for novel FADH2-dependent halogenases in marine sponge-associated microbial consortia. Mar. Biotechnol. 2013, 15, 63–72. [Google Scholar] [PubMed]
- Fieseler, L.; Horn, M.; Wagner, M.; Hentschel, U. Discovery of the novel candidate phylum “Poribacteria” in marine sponges. Appl. Environ. Microbiol. 2004, 70, 3724–3732. [Google Scholar] [PubMed]
- Hentschel, U.; Hopke, J.; Horn, M.; Friedrich, A.B.; Wagner, M.; Hacker, J.; Moore, B.S. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 2002, 68, 4431–4440. [Google Scholar] [PubMed]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N.; Perez, T.; Rodrigo, A.; Schupp, P.J.; Vacelet, J.; et al. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012, 6, 564–576. [Google Scholar]
- Siegl, A.; Hentschel, U. PKS and NRPS gene clusters from microbial symbiont cells of marine sponges by whole genome amplification. Environ. Microbiol. Rep. 2010, 2, 507–513. [Google Scholar] [PubMed]
- Fan, L.; Liu, M.; Simister, R.; Webster, N.S.; Thomas, T. Marine microbial symbiosis heats up: The phylogenetic and functional response of a sponge holobiont to thermal stress. ISME J. 2013, 7, 991–1002. [Google Scholar] [PubMed]
- Pantile, R.; Webster, N. Strict thermal threshold identified by quantitative PCR in the sponge Rhopaloeides odorabile. Mar. Ecol. Prog. Ser. 2011, 431, 97–105. [Google Scholar]
- Simister, R.; Taylor, M.W.; Tsai, P.; Webster, N. Sponge-microbe associations survive high nutrients and temperatures. PLoS One 2012, 7, e52220. [Google Scholar]
- Webster, N.S.; Cobb, R.E.; Negri, A.P. Temperature thresholds for bacterial symbiosis with a sponge. ISME J. 2008, 2, 830–842. [Google Scholar] [PubMed]
- Webster, N.S.; Taylor, M.W.; Behnam, F.; Lucker, S.; Rattei, T.; Whalan, S.; Horn, M.; Wagner, M. Deep sequencing reveals exceptional diversity and modes of transmission for bacterial sponge symbionts. Environ. Microbiol. 2010, 12, 2070–2082. [Google Scholar] [PubMed]
- Mohamed, N.M.; Colman, A.S.; Tal, Y.; Hill, R.T. Diversity and expression of nitrogen fixation genes in bacterial symbionts of marine sponges. Environ. Microbiol. 2008, 10, 2910–2921. [Google Scholar] [PubMed]
- Mohamed, N.; Cicirelli, E.; Kan, J.; Chen, F.; Fuqua, C.; Hill, R. Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ. Microbiol. 2008, 10, 75–86. [Google Scholar] [PubMed]
- Mohamed, N.; Rao, V.; Hamann, M.; Kelly, M.; Hill, R. Monitoring bacterial diversity of the marine sponge Ircinia strobilina upon transfer into aquaculture. Appl. Environ. Microbiol. 2008, 74, 4133–4143. [Google Scholar] [PubMed]
- Mohamed, N.M.; Saito, K.; Tal, Y.; Hill, R.T. Diversity of aerobic and anaerobic ammonia-oxidizing bacteria in marine sponges. ISME J. 2010, 4, 38–48. [Google Scholar] [PubMed]
- Zan, J.D.; Fuqua, C.; Hill, R.T. Diversity and functional analysis of luxS genes in vibrios from marine sponges Mycale laxissima and Ircinia strobilina. ISME J. 2011, 5, 1505–1516. [Google Scholar] [PubMed]
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez de Glasby, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schoenberg, C.; et al. World Porifera Database. Available online: http://www.marinespecies.org/porifera (accessed on 20 December 2013).
- Cook, S.C.; Bergquist, P.R. Family Irciniidae Gray, 1867. In System Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., van Soest, R.W.M., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 1022–1027. [Google Scholar]
- Cook, S.; Bergquist, P. New species of dictyoceratid sponges from New Zealand: Genus Ircinia (Porifera: Demospongiae: Dictyoceratida). N. Z. J. Mar. Freshw. Res. 1999, 33, 545–563. [Google Scholar]
- Wörheide, G.; Dohrmann, M.; Erpenbeck, D.; Larroux, C.; Maldonado, M.; Voigt, O.; Borchiellini, C.; Lavrov, D.V. Deep phylogeny and evolution of sponges (Phylum Porifera). In Advances in Marine Biology; Becerro, M.A., Uriz, M.J., Maldonado, M., Turon, X., Eds.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 1–18. [Google Scholar]
- Schmitt, S.; Weisz, J.; Lindquist, N.; Hentschel, U. Vertical transmission of a phylogenetically complex microbial consortium in the viviparous sponge Ircinia felix. Appl. Environ. Microbiol. 2007, 73, 2067–2078. [Google Scholar] [PubMed]
- Weisz, J.; Hentschel, U.; Lindquist, N.; Martens, C. Linking abundance and diversity of sponge-associated microbial communities to metabolic differences in host sponges. Mar. Biol. 2007, 152, 475–483. [Google Scholar]
- Yang, J.K.; Sun, J.; Lee, O.O.; Wong, Y.H.; Qian, P.Y. Phylogenetic diversity and community structure of sponge-associated bacteria from mangroves of the Caribbean Sea. Aquat. Microb. Ecol. 2011, 62, 231–240. [Google Scholar]
- Pita, L.; López-Legentil, S.; Erwin, P.M. Biogeography and host fidelity of bacterial communities in Ircinia spp. from the Bahamas. Microb. Ecol. 2013, 66, 437–447. [Google Scholar] [PubMed]
- Erwin, P.M.; López-Legentil, S.; González-Pech, R.; Turon, X. A specific mix of generalists: Bacterial symbionts in Mediterranean Ircinia spp. FEMS Microbiol. Ecol. 2012, 79, 619–637. [Google Scholar] [PubMed]
- Erwin, P.M.; Pita, L.; López-Legentil, S.; Turon, X. Stability of sponge-associated bacteria over large seasonal shifts in temperature and irradiance. Appl. Environ. Microbiol. 2012, 78, 7358–7368. [Google Scholar] [PubMed]
- Erwin, P.M.; López-Legentil, S.; Turon, X. Ultrastructure, molecular phylogenetics, and chlorophyll a content of novel cyanobacterial symbionts in temperate sponges. Microb. Ecol. 2012, 64, 771–783. [Google Scholar] [PubMed]
- Esteves, A.I.S.; Hardoim, C.C.P.; Xavier, J.R.; Gonçalves, J.M.S.; Costa, R. Molecular richness and biotechnological potential of bacteria cultured from Irciniidae sponges in the north-east Atlantic. FEMS Microbiol. Ecol. 2013, 85, 519–536. [Google Scholar] [PubMed]
- Hardoim, C.C.P.; Esteves, A.I.S.; Pires, F.R.; Gonçalves, J.M.S.; Cox, C.J.; Xavier, J.R.; Costa, R. Phylogenetically and spatially close marine sponges harbour divergent bacterial communities. PLoS One 2012, 7, e53029. [Google Scholar] [PubMed]
- Pita, L.; Turon, X.; López-Legentil, S.; Erwin, P.M. Host rules: Spatial stability of bacterial communities associated with marine sponges (Ircinia spp.) in the western Mediterranean Sea. FEMS Microbiol. Ecol. 2013, 86, 268–276. [Google Scholar] [PubMed]
- Usher, K.; Kuo, J.; Fromont, J.; Toze, S.; Sutton, D. Comparative morphology of five species of symbiotic and non-symbiotic coccoid Cyanobacteria. Eur. J. Phycol. 2006, 41, 179–188. [Google Scholar]
- Wilkinson, C.R. Microbial associations in sponges. III. Ultrastructure of in situ associations in coral-reef sponges. Mar. Biol. 1978, 49, 177–185. [Google Scholar]
- Webster, N.S.; Hill, R.T. The culturable microbial community of the Great Barrier Reef sponge Rhopaloeides odorabile is dominated by an alpha-proteobacterium. Mar. Biol. 2001, 138, 843–851. [Google Scholar]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [PubMed]
- Hardoim, C.C.P.; Costa, R.; Araújo, F.V.; Hajdu, E.; Peixoto, R.; Lins, U.; Rosado, A.S.; van Elsas, J.D. Diversity of bacteria in the marine sponge Aplysina fulva in Brazilian coastal waters. Appl. Environ. Microbiol. 2009, 75, 3331–3343. [Google Scholar] [PubMed]
- Kamke, J.; Taylor, M.W.; Schmitt, S. Activity profiles for marine sponge-associated bacteria obtained by 16S rRNA vs. 16S rRNA gene comparisons. ISME J. 2010, 4, 498–508. [Google Scholar] [PubMed]
- Sara, M. Ultrastructural aspects of symbiosis between two species of genus Aphanocapsa (Cyanophyceae) and Ircinia variabilis (Demospongiae). Mar. Biol. 1971, 11, 214–221. [Google Scholar]
- Wilkinson, C.R. Microbial associations in sponges. I. Ecology, physiology and microbial-populations of coral-reef sponges. Mar. Biol. 1978, 49, 161–167. [Google Scholar]
- Wilkinson, C.R. Microbial associations in sponges. II. Numerical-analysis of sponge and water bacterial populations. Mar. Biol. 1978, 49, 169–176. [Google Scholar]
- Hentschel, U.; Schmid, M.; Wagner, M.; Fieseler, L.; Gernert, C.; Hacker, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001, 35, 305–312. [Google Scholar] [PubMed]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [PubMed]
- Thiel, V.; Imhoff, J.F. Phylogenetic identification of bacteria with antimicrobial activities isolated from Mediterranean sponges. Biomol. Eng. 2003, 20, 421–423. [Google Scholar] [PubMed]
- Enticknap, J.J.; Kelly, M.; Peraud, O.; Hill, R.T. Characterization of a culturable alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. Appl. Environ. Microbiol. 2006, 72, 3724–3732. [Google Scholar] [PubMed]
- O’Halloran, J.A.; Barbosa, T.M.; Morrissey, J.P.; Kennedy, J.; O’Gara, F.; Dobson, A.D.W. Diversity and antimicrobial activity of Pseudovibrio spp. from Irish marine sponges. J. Appl. Microbiol. 2011, 110, 1495–1508. [Google Scholar] [PubMed]
- Tabares, P.; Pimentel-Elardo, S.M.; Schirmeister, T.; Huenig, T.; Hentschel, U. Anti-protease and immunomodulatory activities of bacteria associated with Caribbean sponges. Mar. Biotechnol. 2011, 13, 883–892. [Google Scholar] [PubMed]
- Friedrich, A.B.; Fischer, I.; Proksch, P.; Hacker, J.; Hentschel, U. Temporal variation of the microbial community associated with the Mediterranean sponge Aplysina aerophoba. FEMS Microbiol. Ecol. 2001, 38, 105–113. [Google Scholar]
- Hugenholtz, P. Exploring prokaryotic diversity in the genomic era. Genome Biol. 2002, 3. [Google Scholar] [CrossRef]
- Usher, K.; Fromont, J.; Sutton, D.; Toze, S. The biogeography and phylogeny of unicellular cyanobacterial symbionts in sponges from Australia and the Mediterranean. Microb. Ecol. 2004, 48, 167–177. [Google Scholar] [PubMed]
- Schmitt, S.; Angermeier, H.; Schiller, R.; Lindquist, N.; Hentschel, U. Molecular microbial diversity survey of sponge reproductive stages and mechanistic insights into vertical transmission of microbial symbionts. Appl. Environ. Microbiol. 2008, 74, 7694–7708. [Google Scholar] [PubMed]
- Stabili, L.; Cardone, F.; Alifano, P.; Tredici, S.M.; Piraino, S.; Corriero, G.; Gaino, E. Epidemic mortality of the sponge Ircinia variabilis (Schmidt, 1862) associated to proliferation of a Vibrio bacterium. Microb. Ecol. 2012, 64, 802–813. [Google Scholar] [PubMed]
- Lee, O.O.; Wang, Y.; Yang, J.K.; Lafi, F.F.; Al-Suwailem, A.; Qian, P.Y. Pyrosequencing reveals highly diverse and species-specific microbial communities in sponges from the Red Sea. ISME J. 2011, 5, 650–664. [Google Scholar] [PubMed]
- Hardoim, C.C.P.; Cardinale, M.; Cúcio, A.; Esteves, A.I.S.; Berg, G.; Xavier, J.R.; Cox, C.; Costa, R. Bacterial cultivation bias examined for keratose marine sponges. Unpublised work. 2014. [Google Scholar]
- Hardoim, C.C.P.; Costa, R. Temporal dynamics of prokaryotic communities in the marine sponge Sarcotragus spinosulus. Mol. Ecol. 2014, 23, 3097–3112. [Google Scholar] [PubMed]
- Fiore, C.L.; Jarett, J.K.; Olson, N.D.; Lesser, M.P. Nitrogen fixation and nitrogen transformations in marine symbioses. Trends Microbiol. 2010, 18, 455–463. [Google Scholar] [PubMed]
- Zehr, J.P.; Mellon, M.T.; Zani, S. New nitrogen-fixing microorganisms detected in oligotrophic oceans by amplification of nitrogenase (nifH) genes. Appl. Environ. Microbiol. 1998, 64, 3444–3450. [Google Scholar] [PubMed]
- Zehr, J.P.; Jenkins, B.D.; Short, S.M.; Steward, G.F. Nitrogenase gene diversity and microbial community structure: A cross-system comparison. Environ. Microbiol. 2003, 5, 539–554. [Google Scholar] [PubMed]
- Man-Aharonovich, D.; Kress, N.; Bar Zeev, E.; Berman-Frank, I.; Beja, O. Molecular ecology of nifH genes and transcripts in the eastern Mediterranean Sea. Environ. Microbiol. 2007, 9, 2354–2363. [Google Scholar] [PubMed]
- Thakur, N.; Anil, A. Antibacterial activity of the sponge Ircinia ramosa: Importance of its surface-associated bacteria. J. Chem. Ecol. 2000, 26, 57–71. [Google Scholar]
- Thakur, N.; Anil, A.; Müller, W. Culturable epibacteria of the marine sponge Ircinia fusca: Temporal variations and their possible role in the epibacterial defense of the host. Aquat. Microb. Ecol. 2004, 37, 295–304. [Google Scholar]
- Pandey, S.; Sree, A.; Dash, S.S.; Sethi, D.P.; Chowdhury, L. Diversity of marine bacteria producing beta-glucosidase inhibitors. Microb. Cell. Fact. 2013, 12. [Google Scholar] [CrossRef]
- Asano, N. Glycosidase inhibitors: Update and perspectives on practical use. Glycobiology 2003, 13, 93R–104R. [Google Scholar] [PubMed]
- De Rosa, S.; Mitova, M.; Tommonaro, G. Marine bacteria associated with sponge as source of cyclic peptides. Biomol. Eng. 2003, 20, 311–316. [Google Scholar]
- Holden, M.T.G.; Chhabra, S.R.; de Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [PubMed]
- Degrassi, G.; Aguilar, C.; Bosco, M.; Zahariev, S.; Pongor, S.; Venturi, V. Plant growth-promoting Pseudomonas putida WCS358 produces and secretes four cyclic dipeptides: Cross-talk with quorum sensing bacterial sensors. Curr. Microbiol. 2002, 45, 250–254. [Google Scholar] [PubMed]
- Kalinovskaya, N.; Kuznetsova, T.; Rashkes, Y.; Milgrom, Y.; Milgrom, E.; Willis, R.; Wood, A.; Kurtz, H.; Carabedian, C.; Murphy, P.; et al. Surfactin-like structures of five cyclic depsipeptides from the marine isolate of Bacillus pumilus. Russ. Chem. Bull. 1995, 44, 951–955. [Google Scholar]
- Prokof'eva, N.; Kalinovskaya, N.; Luk’yanov, P.; Shentsova, E.; Kuznetsova, T. The membranotropic activity of cyclic acyldepsipeptides from bacterium Bacillus pumilus, associated with the marine sponge Ircinia sp. Toxicon 1999, 37, 801–813. [Google Scholar] [PubMed]
- Simister, R.L.; Deines, P.; Botte, E.S.; Webster, N.S.; Taylor, M.W. Sponge-specific clusters revisited: A comprehensive phylogeny of sponge-associated microorganisms. Environ. Microbiol. 2012, 14, 517–524. [Google Scholar] [PubMed]
- Brochier-Armanet, C.; Boussau, B.; Gribaldo, S.; Forterre, P. Mesophilic crenarchaeota: Proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 2008, 6, 245–252. [Google Scholar] [PubMed]
- Spang, A.; Hatzenpichler, R.; Brochier-Armanet, C.; Rattei, T.; Tischler, P.; Spieck, E.; Streit, W.; Stahl, D.A.; Wagner, M.; Schleper, C. Distinct gene set in two different lineages of ammonia-oxidizing Archaea supports the phylum Thaumarchaeota. Trends Microbiol. 2010, 18, 331–340. [Google Scholar] [PubMed]
- Pester, M.; Schleper, C.; Wagner, M. The Thaumarchaeota: An emerging view of their phylogeny and ecophysiology. Curr. Opin. Microbiol. 2011, 14, 300–306. [Google Scholar] [PubMed]
- Lee, E.Y.; Lee, H.K.; Lee, Y.K.; Sim, C.J.; Lee, J.H. Diversity of symbiotic archaeal communities in marine sponges from Korea. Biomol. Eng. 2003, 20, 299–304. [Google Scholar] [PubMed]
- Webster, N.S.; Watts, J.E.M.; Hill, R.T. Detection and phylogenetic analysis of novel crenarchaeote and euryarchaeote 16S ribosomal RNA gene sequences from a Great Barrier Reef sponge. Mar. Biotechnol. 2001, 3, 600–608. [Google Scholar] [PubMed]
- Webster, N.S.; Negri, A.P.; Munro, M.; Battershill, C.N. Diverse microbial communities inhabit Antarctic sponges. Environ. Microbiol. 2004, 6, 288–300. [Google Scholar] [PubMed]
- Hoffmann, F.; Radax, R.; Woebken, D.; Holtappels, M.; Lavik, G.; Rapp, H.T.; Schlappy, M.L.; Schleper, C.; Kuypers, M.M.M. Complex nitrogen cycling in the sponge Geodia barretti. Environ. Microbiol. 2009, 11, 2228–2243. [Google Scholar] [PubMed]
- Han, M.Q.; Liu, F.; Zhang, F.L.; Li, Z.Y.; Lin, H.W. Bacterial and archaeal symbionts in the south China Sea sponge Phakellia fusca: Community structure, relative abundance, and ammonia-oxidizing populations. Mar. Biotechnol. 2012, 14, 701–713. [Google Scholar] [PubMed]
- Cardoso, J.; van Bleijswijk, J.D.L.; Witte, H.; van Duyl, F.C. Diversity and abundance of ammonia-oxidizing Archaea and Bacteria in tropical and cold-water coral reef sponges. Aquat. Microb. Ecol. 2013, 68, 215–230. [Google Scholar]
- Höller, U.; Wright, A.D.; Matthee, G.F.; Konig, G.M.; Draeger, S.; Aust, H.J.; Schulz, B. Fungi from marine sponges: Diversity, biological activity and secondary metabolites. Mycol. Res. 2000, 104, 1354–1365. [Google Scholar]
- Paz, Z.; Komon-Zelazowska, M.; Druzhinina, I.S.; Aveskamp, M.M.; Shnaiderman, A.; Aluma, Y.; Carmeli, S.; Ilan, M.; Yarden, O. Diversity and potential antifungal properties of fungi associated with a Mediterranean sponge. Fungal Divers. 2010, 42, 17–26. [Google Scholar]
- Bringmann, G.; Lang, G.; Gulder, T.; Tsuruta, H.; Muhlbacher, J.; Maksimenka, K.; Steffens, S.; Schaumann, K.; Stohr, R.; Wiese, J.; et al. The first sorbicillinoid alkaloids, the antileukemic sorbicillactones A and B, from a sponge-derived Penicillium chrysogenum strain. Tetrahedron 2005, 61, 7252–7265. [Google Scholar]
- Ereskovsky, A.V.; Gonobobleva, E.; Vishnyakov, A. Morphological evidence for vertical transmission of symbiotic bacteria in the viviparous sponge Halisarca dujardini Johnston (Porifera, Demospongiae, Halisarcida). Mar. Biol. 2005, 146, 869–875. [Google Scholar]
- Sharp, K.H.; Eam, B.; Faulkner, D.J.; Haygood, M.G. Vertical transmission of diverse microbes in the tropical sponge Corticium sp. Appl. Environ. Microbiol. 2007, 73, 622–629. [Google Scholar] [PubMed]
- Lee, O.O.; Chui, P.Y.; Wong, Y.H.; Pawlik, J.R.; Qian, P.Y. Evidence for vertical transmission of bacterial symbionts from adult to embryo in the Caribbean sponge Svenzea zeai. Appl. Environ. Microbiol. 2009, 75, 6147–6156. [Google Scholar] [PubMed]
- Lindquist, N.; Bolser, R.; Laing, K. Timing of larval release by two Caribbean demosponges. Mar. Ecol. 1997, 155, 309–313. [Google Scholar]
- Lopanik, N.; Lindquist, N.; Targett, N. Potent cytotoxins produced by a microbial symbiont protect host larvae from predation. Oecologia 2004, 139, 131–139. [Google Scholar] [PubMed]
- Lopanik, N.; Gustafson, K.R.; Lindquist, N. Structure of bryostatin 20: A symbiont-produced chemical defense for larvae of the host bryozoan, Bugula neritina. J. Nat. Prod. 2004, 67, 1412–1414. [Google Scholar] [PubMed]
- Lopanik, N.B.; Targett, N.M.; Lindquist, N. Isolation of two polyketide synthase gene fragments from the uncultured microbial symbiont of the marine bryozoan Bugula neritina. Appl. Environ. Microbiol. 2006, 72, 7941–7944. [Google Scholar] [PubMed]
- Lopanik, N.B.; Shields, J.A.; Buchholz, T.J.; Rath, C.M.; Hothersall, J.; Haygood, M.G.; Hakansson, K.; Thomas, C.M.; Sherman, D.H. In vivo and in vitro trans-acylation by BryP, the putative bryostatin pathway acyltransferase derived from an uncultured marine symbiont. Chem. Biol. 2008, 15, 1175–1186. [Google Scholar] [PubMed]
- Sudek, S.; Lopanik, N.B.; Waggoner, L.E.; Hildebrand, M.; Anderson, C.; Liu, H.B.; Patel, A.; Sherman, D.H.; Haygood, M.G. Identification of the putative bryostatin polyketide synthase gene cluster from “Candidatus Endobugula sertula”, the uncultivated microbial symbiont of the marine bryozoan Bugula neritina. J. Nat. Prod. 2007, 70, 67–74. [Google Scholar] [PubMed]
- Jaeckle, W.B. Transport and metabolism of alanine and palmitic acid by field-collected larvae of Tedania ignis (Porifera, Demospongiae)—Estimated consequences of limited label translocation. Biol. Bull. 1995, 189, 159–167. [Google Scholar]
- Bandara, H.; Lam, O.L.T.; Jin, L.J.; Samaranayake, L. Microbial chemical signaling: A current perspective. Crit. Rev. Microbiol. 2012, 38, 217–249. [Google Scholar] [PubMed]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [PubMed]
- Skindersoe, M.E.; Ettinger-Epstein, P.; Rasmussen, T.B.; Bjarnsholt, T.; de Nys, R.; Givskov, M. Quorum sensing antagonism from marine organisms. Mar. Biotechnol. 2008, 10, 56–63. [Google Scholar] [PubMed]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V.J. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling 2011, 27, 893–905. [Google Scholar] [PubMed]
- Dieckmann, R.; Graeber, I.; Kaesler, I.; Szewzyk, U.; von Dohren, H. Rapid screening and dereplication of bacterial isolates from marine sponges of the Sula Ridge by Intact-Cell-MALDI-TOF mass spectrometry (ICM-MS). Appl. Microbiol. Biotechnol. 2005, 67, 539–548. [Google Scholar] [PubMed]
- Anand, T.P.; Bhat, A.W.; Shouche, Y.S.; Roy, U.; Siddharth, J.; Sarma, S.P. Antimicrobial activity of marine bacteria associated with sponges from the waters off the coast of south east India. Microbiol. Res. 2006, 161, 252–262. [Google Scholar] [PubMed]
- Hoffmann, M.; Fischer, M.; Ottesen, A.; McCarthy, P.J.; Lopez, J.V.; Brown, E.W.; Monday, S.R. Population dynamics of Vibrio spp. associated with marine sponge microcosms. ISME J. 2010, 4, 1608–1612. [Google Scholar] [PubMed]
- Flemer, B.; Kennedy, J.; Margassery, L.M.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D.W. Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. J. Appl. Microbiol. 2012, 112, 289–301. [Google Scholar] [PubMed]
- Webster, N.S. Sponge disease: A global threat? Environ. Microbiol. 2007, 9, 1363–1375. [Google Scholar]
- Gaino, E.; Pronzato, R.; Corriero, G.; Buffa, P. Mortality of commercial sponges—Incidence in two Mediterranean areas. Boll. Zool. 1992, 59, 79–85. [Google Scholar]
- Wulff, J.L. Rapid diversity and abundance decline in a Caribbean coral reef sponge community. Biol. Conserv. 2006, 127, 167–176. [Google Scholar]
- Maldonado, M.; Sanchez-Tocino, L.; Navarro, C. Recurrent disease outbreaks in corneous demosponges of the genus Ircinia: Epidemic incidence and defense mechanisms. Mar. Biol. 2010, 157, 1577–1590. [Google Scholar]
- Cebrian, E.; Uriz, M.J.; Garrabou, J.; Ballesteros, E. Sponge mass mortalities in a warming Mediterranean sea: Are Cyanobacteria-harboring species worse off? PLoS One 2011, 6, e20211. [Google Scholar] [CrossRef]
- di Camillo, C.G.; Bartolucci, I.; Cerrano, C.; Bavestrello, G. Sponge disease in the Adriatic Sea. Mar. Ecol. 2013, 34, 62–71. [Google Scholar]
- Pita, L.; Erwin, P.M.; Turon, X.; López-Legentil, S. Till death do us part: Stable sponge-bacteria associations under thermal and food shortage stresses. PLoS One 2013, 8, e80307. [Google Scholar] [PubMed]
- Luter, H.M.; Whalan, S.; Webster, N.S. Prevalence of tissue necrosis and brown spot lesions in a common marine sponge. Mar. Freshw. Res. 2010, 61, 484–489. [Google Scholar]
- Mohamed, N.M.; Enticknap, J.J.; Lohr, J.E.; McIntosh, S.M.; Hill, R.T. Changes in bacterial communities of the marine sponge Mycale laxissima on transfer into aquaculture. Appl. Environ. Microbiol. 2008, 74, 1209–1222. [Google Scholar] [PubMed]
- Isaacs, L.T.; Kan, J.J.; Nguyen, L.; Videau, P.; Anderson, M.A.; Wright, T.L.; Hill, R.T. Comparison of the bacterial communities of wild and captive sponge Clathria prolifera from the Chesapeake Bay. Mar. Biotechnol. 2009, 11, 758–770. [Google Scholar] [PubMed]
- Thomas, T.; Rusch, D.; DeMaere, M.Z.; Yung, P.Y.; Lewis, M.; Halpern, A.; Heidelberg, K.B.; Egan, S.; Steinberg, P.D.; Kjelleberg, S. Functional genomic signatures of sponge bacteria reveal unique and shared features of symbiosis. ISME J. 2010, 4, 1557–1567. [Google Scholar] [PubMed]
- Fan, L.; Reynolds, D.; Liu, M.; Stark, M.; Kjelleberg, S.; Webster, N.S.; Thomas, T. Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts. Proc. Natl. Acad. Sci. USA 2012, 109, E1878–E1887. [Google Scholar] [PubMed]
- Simister, R.; Taylor, M.W.; Tsai, P.; Fan, L.; Bruxner, T.J.; Crowe, M.L.; Webster, N. Thermal stress responses in the bacterial biosphere of the Great Barrier Reef sponge Rhopaloeides odorabile. Environ. Microbiol. 2012, 14, 3232–3246. [Google Scholar] [PubMed]
- Webster, N.; Pantile, R.; Botte, E.; Abdo, D.; Andreakis, N.; Whalan, S. A complex life cycle in a warming planet: Gene expression in thermally stressed sponges. Mol. Ecol. 2013, 22, 1854–1868. [Google Scholar] [PubMed]
- Duckworth, A.R.; Peterson, B.J. Effects of seawater temperature and pH on the boring rates of the sponge Cliona celata in scallop shells. Mar. Biol. 2013, 160, 27–35. [Google Scholar]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [PubMed]
- Piel, J.; Butzke, D.; Fusetani, N.; Hui, D.Q.; Platzer, M.; Wen, G.P.; Matsunaga, S. Exploring the chemistry of uncultivated bacterial symbionts: Antitumor polyketides of the pederin family. J. Nat. Prod. 2005, 68, 472–479. [Google Scholar] [PubMed]
- Schirmer, A.; Gadkari, R.; Reeves, C.D.; Ibrahim, F.; DeLong, E.F.; Hutchinson, C.R. Metagenomic analysis reveals diverse polyketide synthase gene clusters in microorganisms associated with the marine sponge Discodermia dissoluta. Appl. Environ. Microbiol. 2005, 71, 4840–4849. [Google Scholar] [PubMed]
- Siegl, A.; Kamke, J.; Hochmuth, T.; Piel, J.; Richter, M.; Liang, C.G.; Dandekar, T.; Hentschel, U. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J. 2011, 5, 61–70. [Google Scholar] [PubMed]
- Fisch, K.; Gurgui, C.; Heycke, N.; van der Sar, S.; Anderson, S.; Webb, V.; Taudien, S.; Platzer, M.; Rubio, B.; Robinson, S.; et al. Polyketide assembly lines of uncultivated sponge symbionts from structure-based gene targeting. Nat. Chem. Biol. 2009, 5, 494–501. [Google Scholar] [PubMed]
- Hochmuth, T.; Niederkruger, H.; Gernert, C.; Siegl, A.; Taudien, S.; Platzer, M.; Crews, P.; Hentschel, U.; Piel, J. Linking chemical and microbial diversity in marine sponges: Possible role for Poribacteria as producers of methyl-branched fatty acids. Chembiochem 2010, 11, 2572–2578. [Google Scholar] [PubMed]
- Osinga, R.; Tramper, J.; Wijffels, R.H. Cultivation of marine sponges. Mar. Biotechnol. 1999, 1, 509–532. [Google Scholar] [PubMed]
- Rinkevich, B. Marine invertebrate cell cultures: New millennium trends. Mar. Biotechnol. 2005, 7, 429–439. [Google Scholar] [PubMed]
- Sipkema, D.; Osinga, R.; Schatton, W.; Mendola, D.; Tramper, J.; Wijffels, R.H. Large-scale production of pharmaceuticals by marine sponges: Sea, cell, or synthesis? Biotechnol. Bioeng. 2005, 90, 201–222. [Google Scholar] [CrossRef]
- Wilkinson, C.R.; Vacelet, J. Transplantation of marine sponges to different conditions of light and current. J. Exp. Mar. Biol. Ecol. 1979, 37, 91–104. [Google Scholar]
- Duckworth, A.; Battershill, C.; Bergquist, P. Influence of explant procedures and environmental factors on culture success of three sponges. Aquaculture 1997, 156, 251–267. [Google Scholar]
- Van Treeck, P.; Eisinger, M.; Müller, J.; Paster, M.; Schuhmacher, H. Mariculture trials with Mediterranean sponge species—The exploitation of an old natural resource with sustainable and novel methods. Aquaculture 2003, 218, 439–455. [Google Scholar]
- De Voogd, N. An assessment of sponge mariculture potential in the Spermonde Archipelago, Indonesia. J. Mar. Biol. Assoc. UK 2007, 396, 1777–1784. [Google Scholar]
- De Rosa, S.; de Caro, S.; Tommonaro, G.; Slantchev, K.; Stefanov, K.; Popov, S. Development in a primary cell culture of the marine sponge Ircinia muscarum and analysis of the polar compounds. Mar. Biotechnol. 2001, 3, 281–286. [Google Scholar] [PubMed]
- De Rosa, S.; Tommonaro, G.; Slantchev, K.; Stefanov, K.; Popov, S. Lipophylic metabolites from the marine sponge Ircinia muscarum and its cell cultures. Mar. Biol. 2002, 140, 465–470. [Google Scholar]
- De Rosa, S.; de Caro, S.; Iodice, C.; Tommonaro, G.; Stefanov, K.; Popov, S. Development in primary cell culture of demosponges. J. Biotechnol. 2003, 100, 119–125. [Google Scholar] [PubMed]
- De Caralt, S.; Uriz, M.J.; Wijffels, R.H. Cell culture from sponges: Pluripotency and immortality. Trends Biotechnol. 2007, 25, 467–471. [Google Scholar] [PubMed]
- Piel, J.; Hui, D.Q.; Wen, G.P.; Butzke, D.; Platzer, M.; Fusetani, N.; Matsunaga, S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl. Acad. Sci. USA 2004, 101, 16222–16227. [Google Scholar] [PubMed]
- Liu, M.; Fan, L.; Zhong, L.; Kjelleberg, S.; Thomas, T. Metaproteogenomic analysis of a community of sponge symbionts. ISME J. 2012, 6, 1515–1525. [Google Scholar] [PubMed]
- Kamke, J.; Sczyrba, A.; Ivanova, N.; Schwientek, P.; Rinke, C.; Mavromatis, K.; Woyke, T.; Hentschel, U. Single-cell genomics reveals complex carbohydrate degradation patterns in poribacterial symbionts of marine sponges. ISME J. 2013, 7, 2287–2300. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).