Biochemical and Electrophysiological Characterization of Two Sea Anemone Type 1 Potassium Toxins from a Geographically Distant Population of Bunodosoma caissarum

 ,
, 
Abstract
:1. Introduction
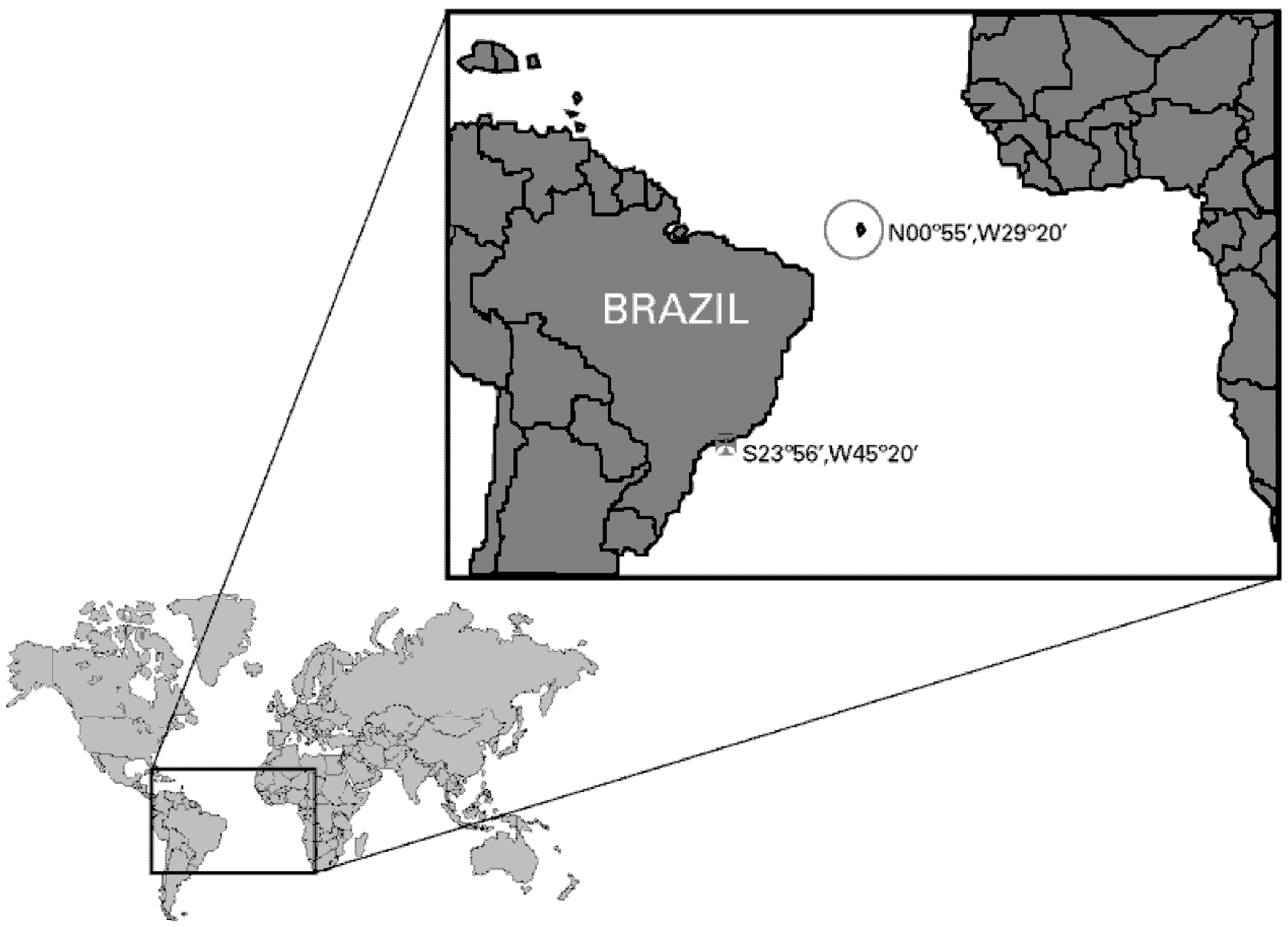
2. Results and Discussion
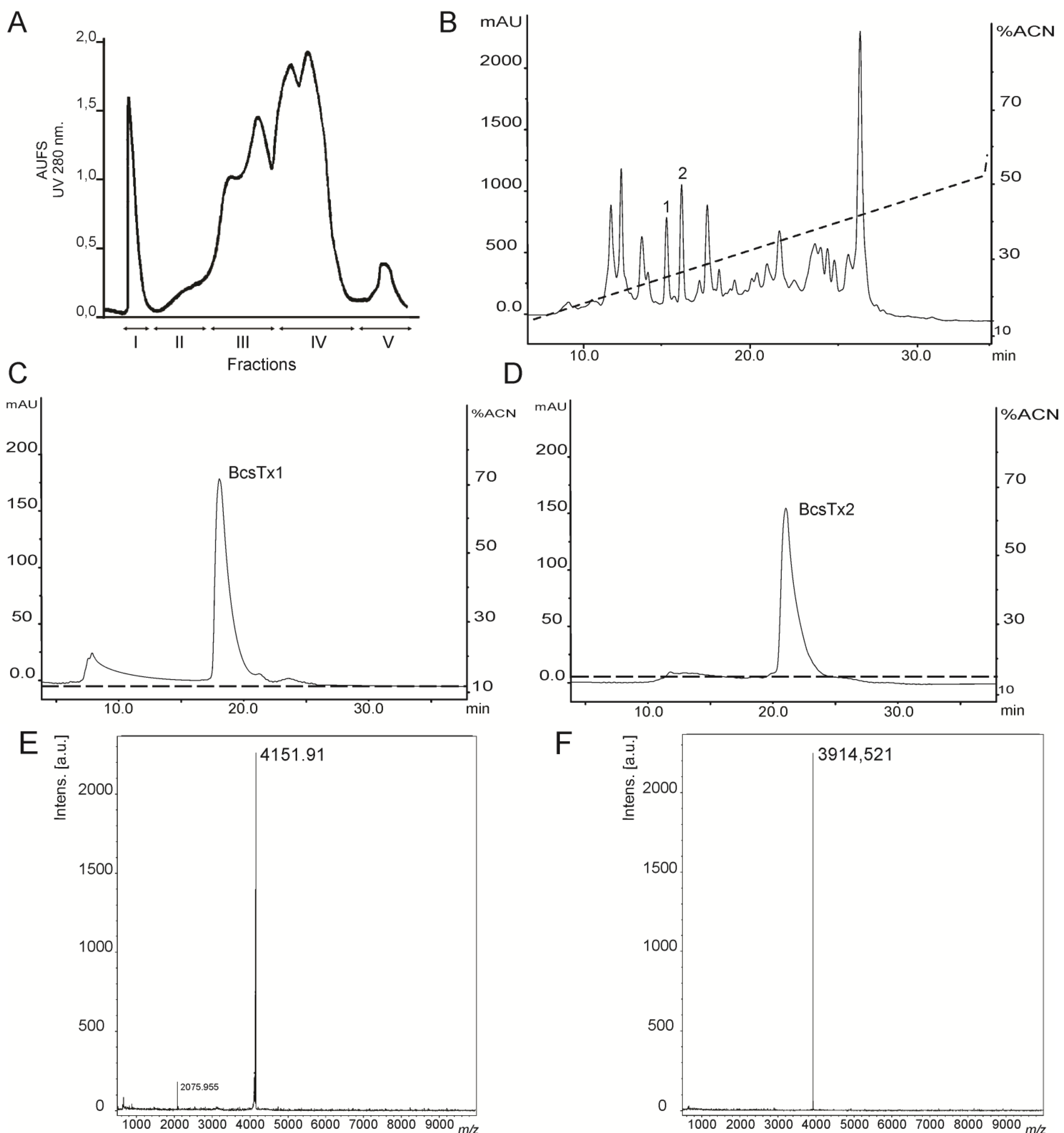
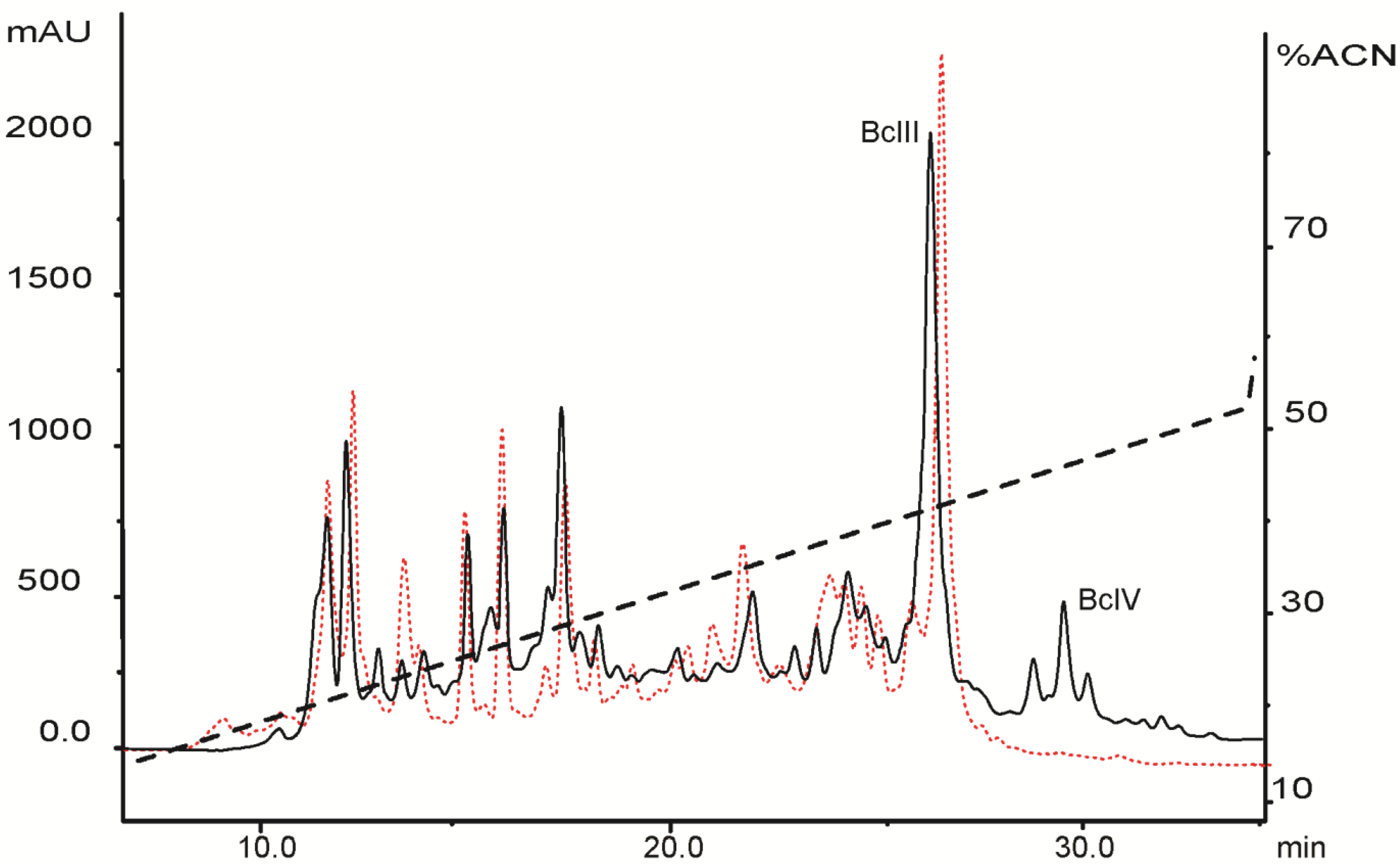
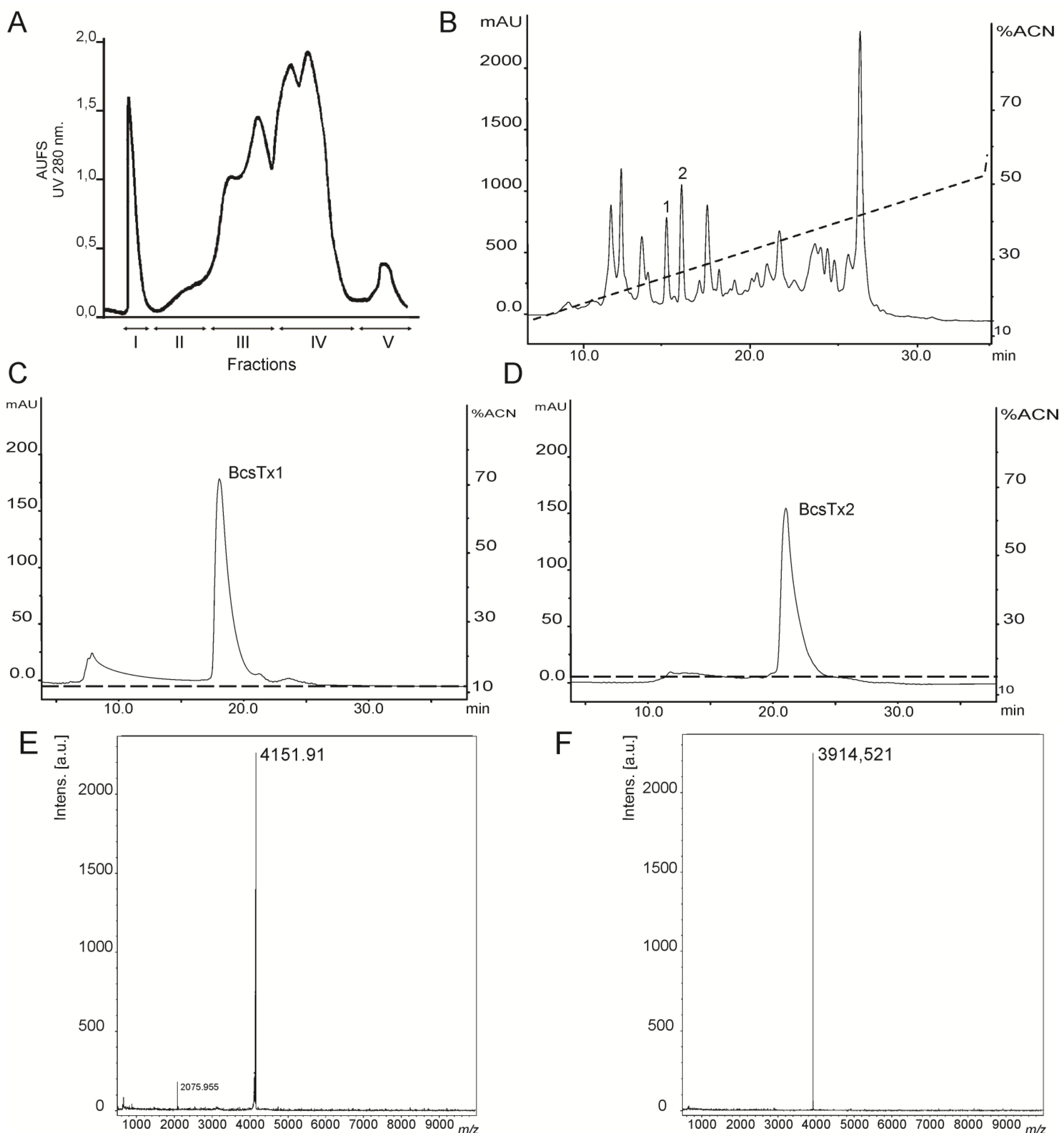
2.1. Venom Purification and Biochemical Characterization of BcsTx1 and BcsTx2
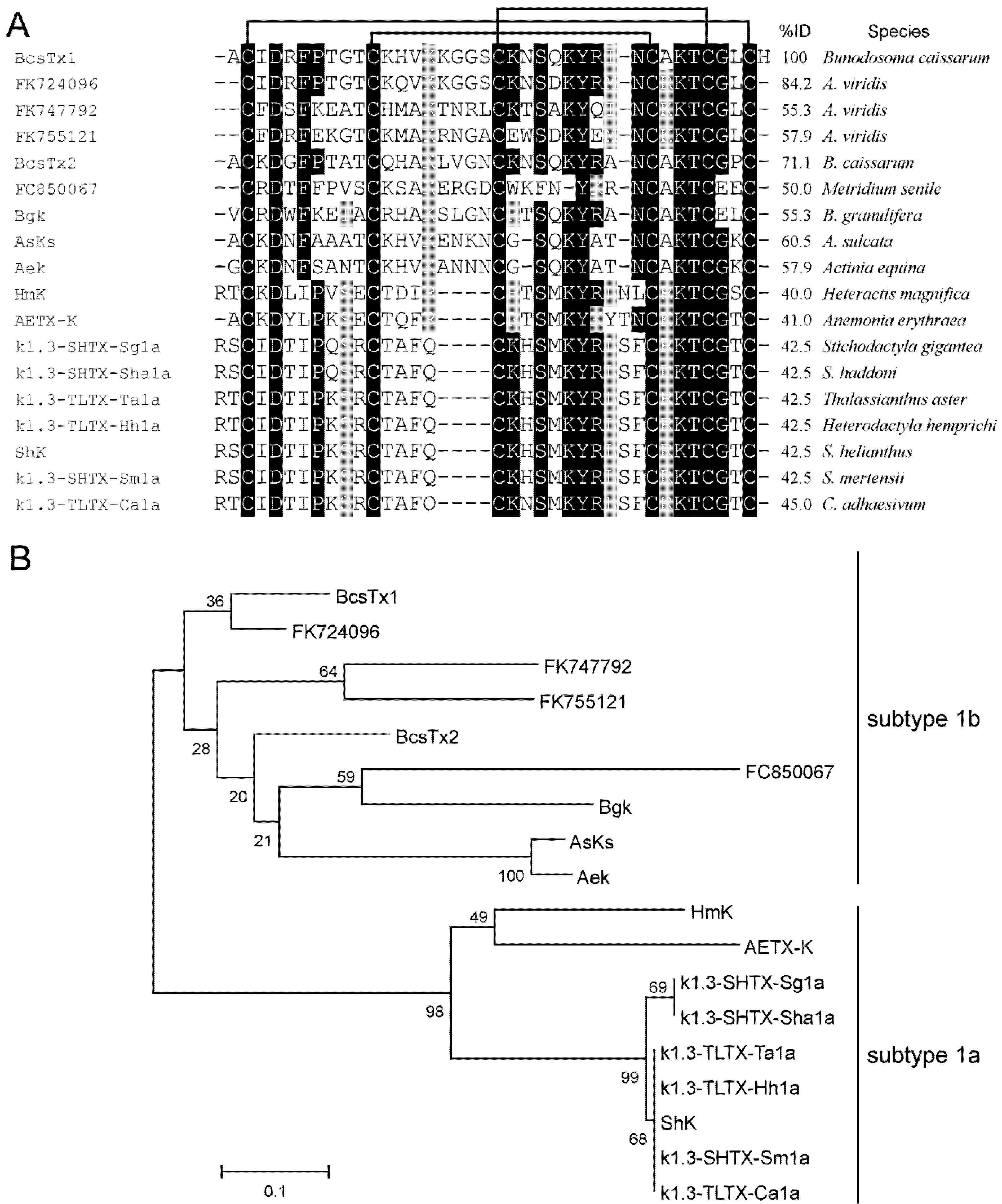
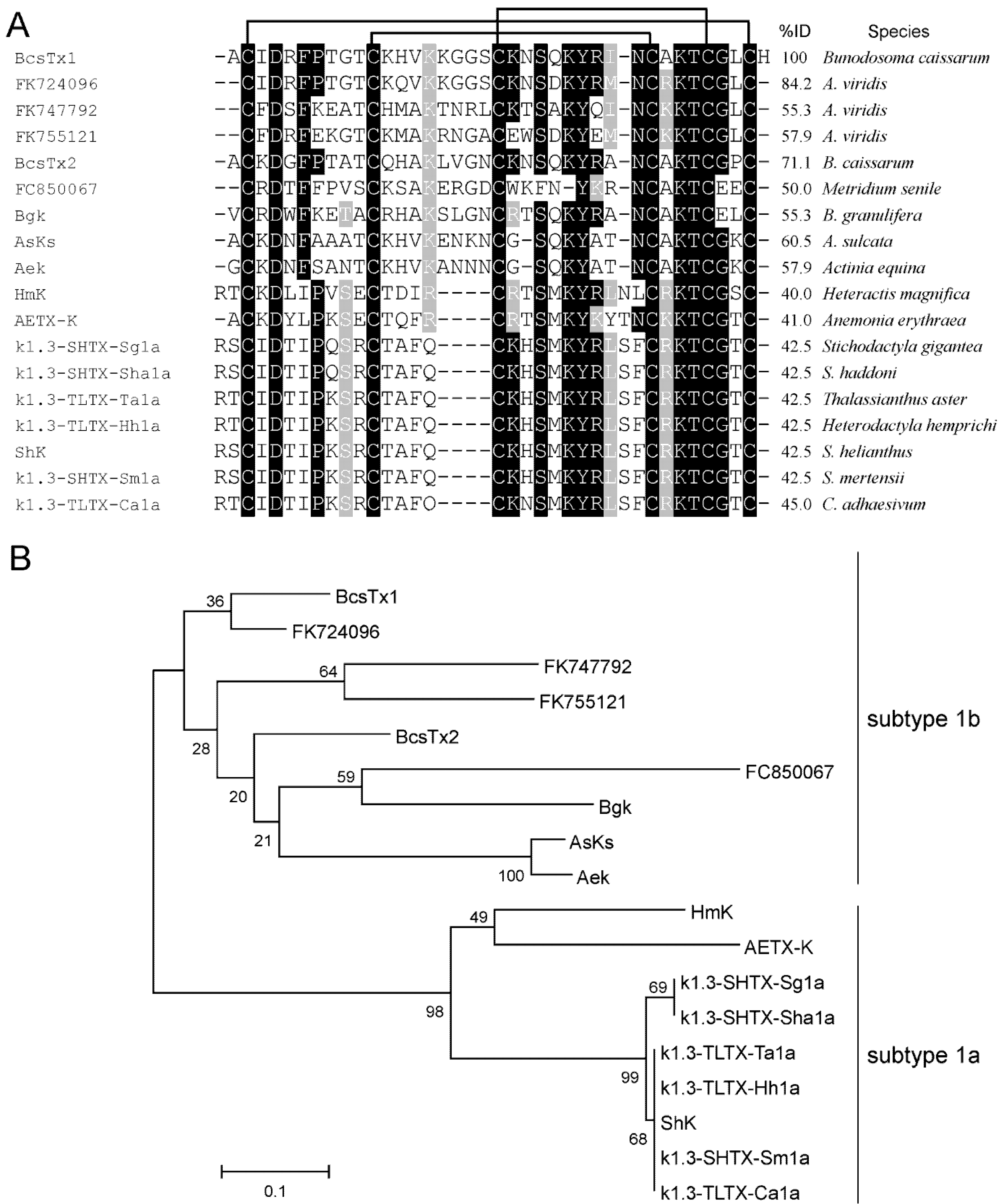
2.2. Amino Acid Sequences and Phylogenetic Analysis

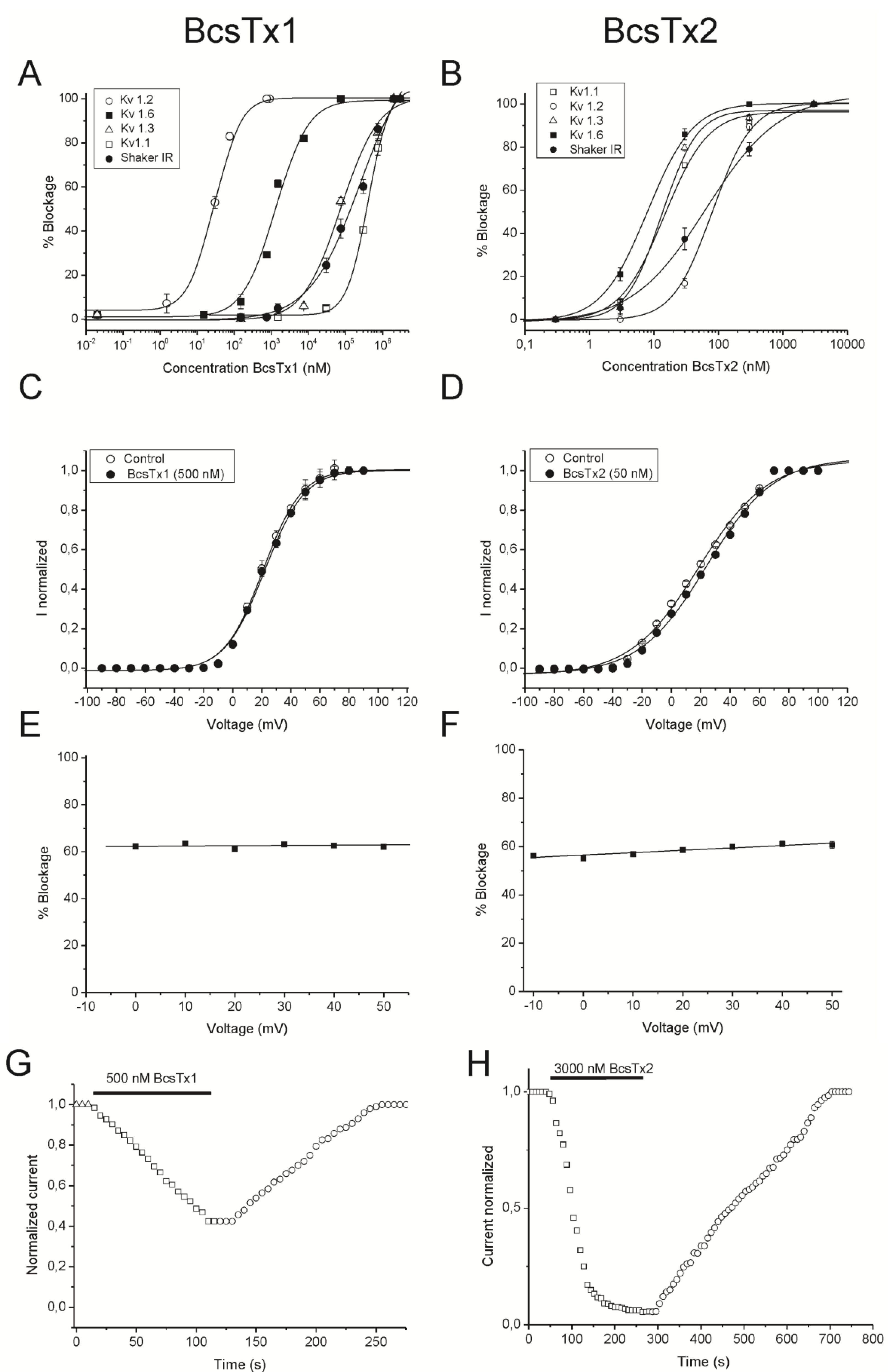
2.3. BcsTx1 and BcsTx2 Pharmacological Profiles
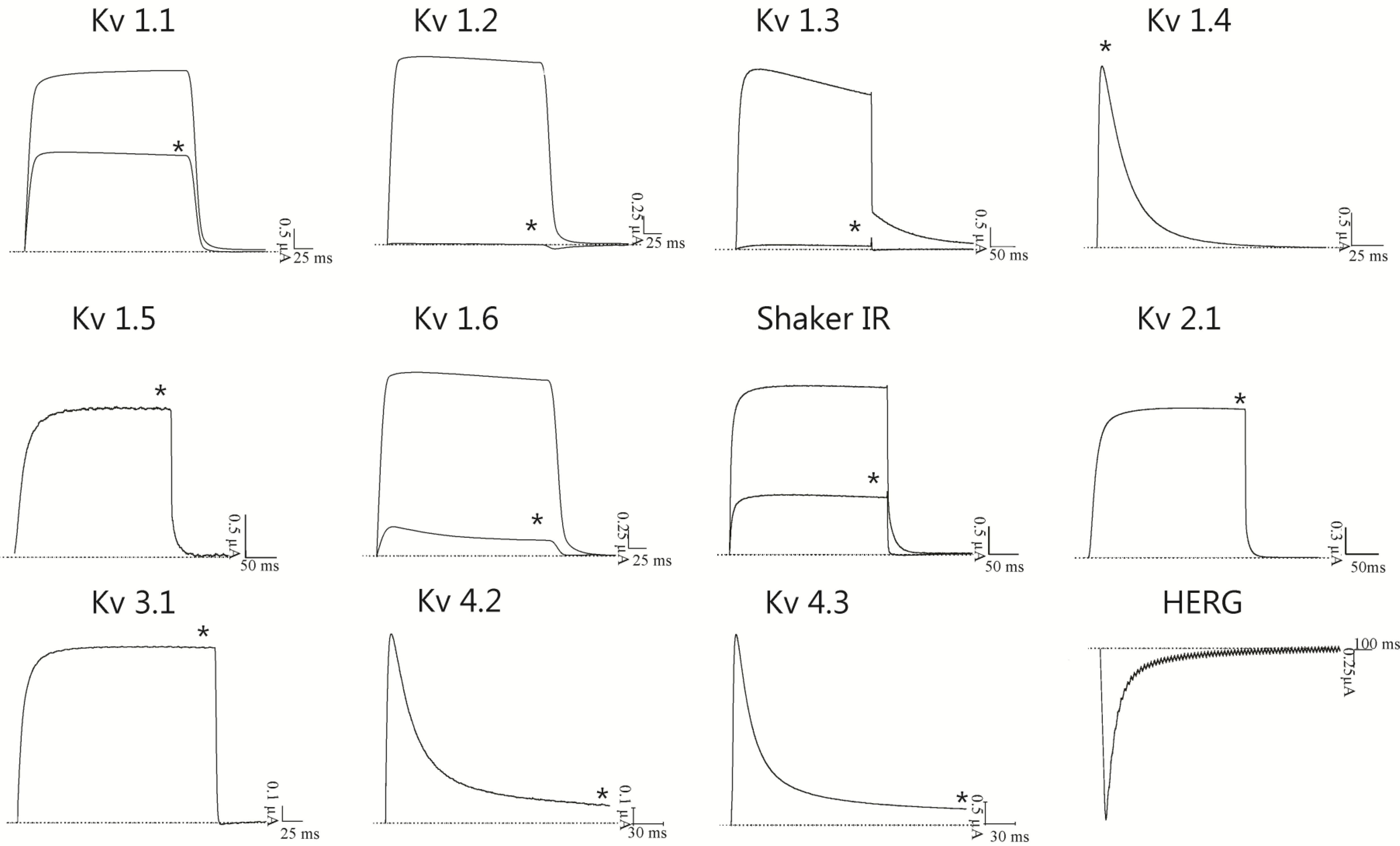

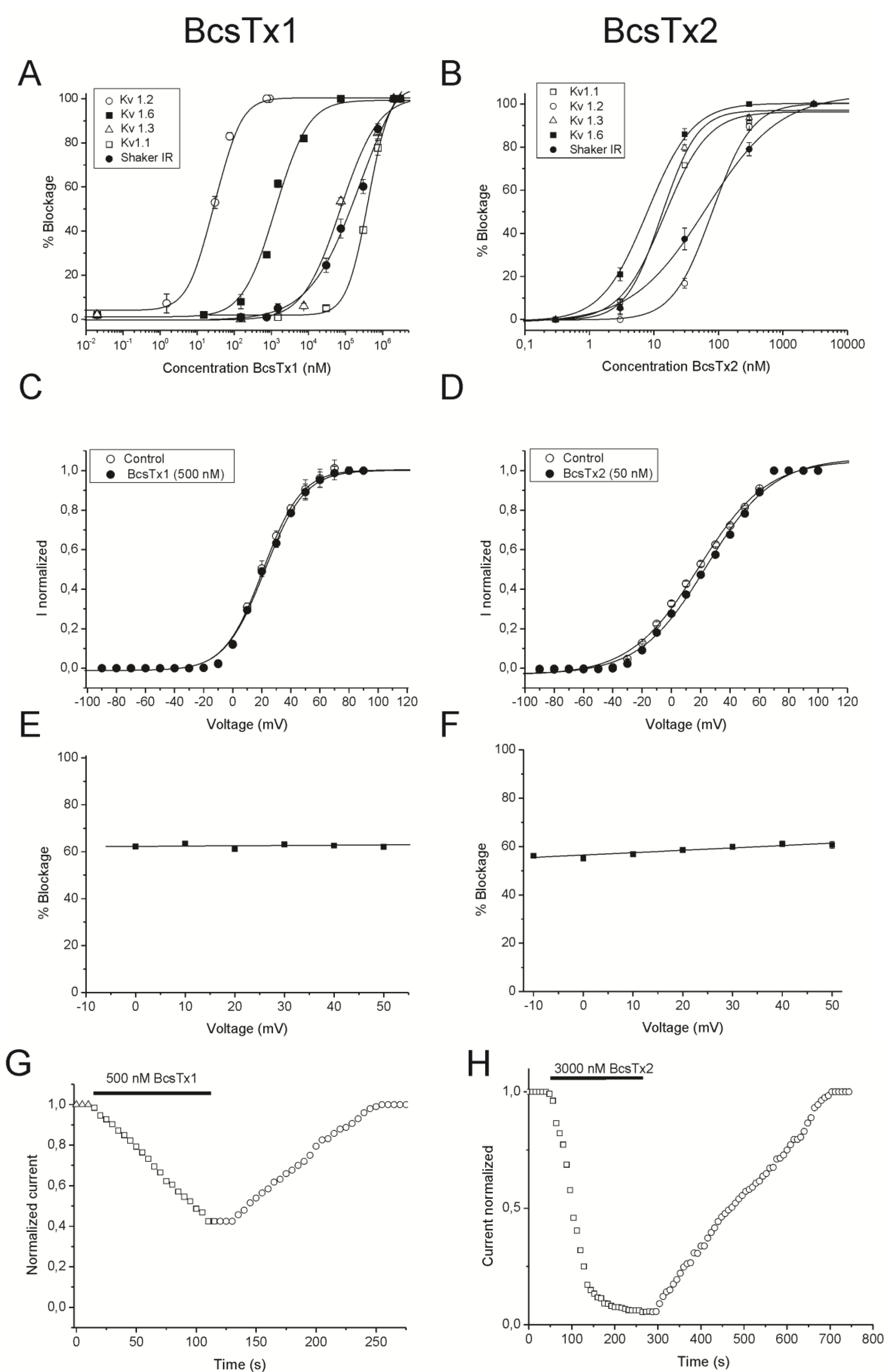

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoforms | BcsTx1 | BcsTx2 |
|---|---|---|
| KV1.1 | 405 ± 20.56 | 14.42 ± 2.61 |
| KV1.2 | 0.03 ± 0.006 | 80.40 ± 1.44 |
| KV1.3 | 74.11 ± 20.24 | 13.12 ± 3.29 |
| KV1.6 | 1.31 ± 0.20 | 7.76 ± 1.90 |
| Shaker IR | 247.69 ± 95.97 | 49.14 ± 3.44 |
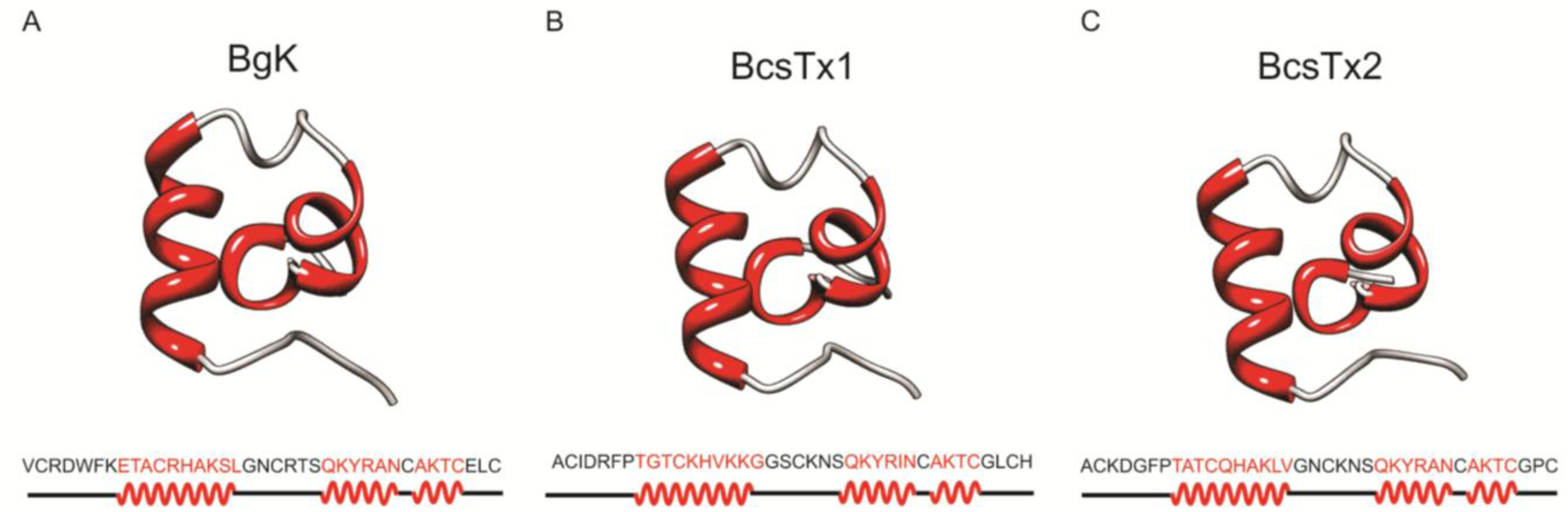
2.4. Bioinformatics Analysis
Molecular Models of BcsTx-1 and -2
3. Experimental Section
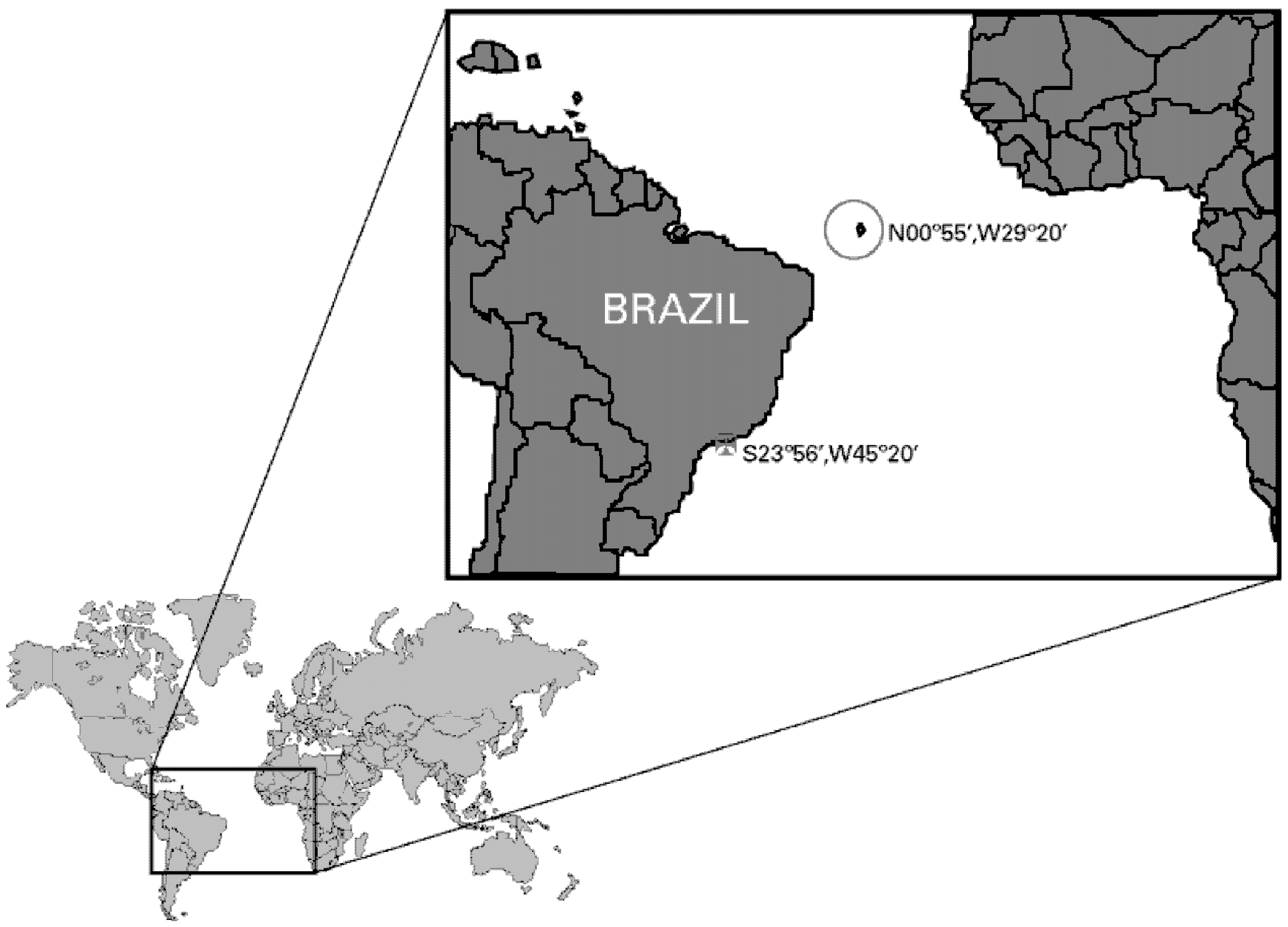
3.1. Sea Anemone Collection, Venom Isolation and Neurotoxins Purification
3.2. Mass Spectrometry Analysis
3.3. Amino Acid Sequence Determination
3.4. Expression of Voltage-Gated Ion Channels in Xenopus laevis Oocytes
3.5. Electrophysiological Recordings
3.6. Phylogenetic Analysis and Sequence Alignment
3.7. Structure Computational Modeling
3.8. Statistical Assessment
4. Conclusions
Acknowledgments
References
- Castaneda, O.; Harvey, A.L. Discovery and characterization of cnidarian peptide toxins that affect neuronal potassium ion channels. Toxicon 2009, 54, 1119–1124. [Google Scholar] [CrossRef]
- Nevalainen, T.J.; Peuravuori, H.J.; Quinn, R.J.; Llewellyn, L.E.; Benzie, J.A.; Fenner, P.J.; Winkel, K.D. Phospholipase A2 in cnidaria. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 731–735. [Google Scholar] [CrossRef]
- Kristan, K.C.; Viero, G.; Dalla Serra, M.; Macek, P.; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef]
- Fenical, W. Marine Soft Corals of the Genus Pseudopterogorgia—a Resource for Novel Anti-Inflammatory Diterpenoids. J. Nat. Prod. 1987, 50, 1001–1008. [Google Scholar] [CrossRef]
- Tibballs, J. Australian venomous jellyfish, envenomation syndromes, toxins and therapy. Toxicon 2006, 48, 830–859. [Google Scholar] [CrossRef]
- Lassen, S.; Helmholz, H.; Ruhnau, C.; Prange, A. Characterisation of neurotoxic polypeptides from Cyanea capillata medusae (Scyphozoa). Hydrobiologia 2010, 645, 213–221. [Google Scholar] [CrossRef]
- Bakus, G.J.; Targett, N.M.; Schulte, B. Chemical Ecology of Marine Organisms—an Overview. J. Chem. Ecol. 1986, 12, 951–987. [Google Scholar] [CrossRef]
- Honma, T.; Minagawa, S.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Novel peptide toxins from acrorhagi, aggressive organs of the sea anemone Actinia equina. Toxicon 2005, 46, 768–774. [Google Scholar] [CrossRef]
- Coll, J.C.; Labarre, S.; Sammarco, P.W.; Williams, W.T.; Bakus, G.J. Chemical Defenses in Soft Corals (Coelenterata, Octocorallia) of the Great Barrier-Reef—a Study of Comparative Toxicities. Mar. Ecol. Prog. Ser. 1982, 8, 271–278. [Google Scholar] [CrossRef]
- Bak, R.P.M.; Borsboom, J.L.A. Allelopathic Interaction between a Reef Coelenterate and Benthic Algae. Oecologia 1984, 63, 194–198. [Google Scholar] [CrossRef]
- Sheppard, C.R.C. Interspecific Aggression between Reef Corals with Reference to Their Distribution. Mar. Ecol. Prog. Ser. 1979, 1, 237–247. [Google Scholar] [CrossRef]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels—molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Hasegawa, Y.; Honma, T.; Nagashima, Y.; Shiomi, K. Screening and cDNA cloning of Kv1 potassium channel toxins in sea anemones. Mar. Drugs 2010, 8, 2893–2905. [Google Scholar] [CrossRef]
- Hille, B. The superfamily of voltage-gated channels. In Ion Channel of Excitable Membranes; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001; Volume Chapter 3, pp. 61–93. [Google Scholar]
- Armstrong, C.M.; Hille, B. Voltage-gated ion channels and electrical excitability. Neuron 1998, 20, 371–380. [Google Scholar] [CrossRef]
- Coetzee, W.A.; Amarillo, Y.; Chiu, J.; Chow, A.; Lau, D.; McCormack, T.; Moreno, H.; Nadal, M.S.; Ozaita, A.; Pountney, D.; et al. Molecular diversity of K+ channels. Ann. N. Y. Acad. Sci. 1999, 868, 233–285. [Google Scholar] [CrossRef]
- Gutman, G.A.; Chandy, K.G.; Grissmer, S.; Lazdunski, M.; Mckinnon, D.; Pardo, L.A.; Robertson, G.A.; Rudy, B.; Sanguinetti, M.C.; Stuhmer, W.; Wang, X.L. International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol. Rev. 2005, 57, 473–508. [Google Scholar] [CrossRef]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar. Biotechnol. (NY) 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Diochot, S.; Lazdunzki, M. Sea anemone toxins affecting potassium channels. In Marine Toxins as Research Tools; Fusetani, N., Kem, W., Eds.; Progress in Molecular and Subcellular Biology Volume 46; Springer: Berlin, Germany, 2009; pp. 99–122. [Google Scholar]
- Belem, M.J.C. Anatomy and biology of Bunodosoma caissarum Correa, 1964 (Cnidaria, Anthozoa, Actiniidae). I—Systematic position and morphological and microanatomical revision. An. Acad. Bras. Cienc. 1988, 61, 342–353. [Google Scholar]
- Zamponi, M.O.; Belem, M.J.C.; Schlenz, E.; Acuna, H. Distribution and some ecological aspects of Corallimorpharia and Actiniaria from shallow waters of the South American Atlantic Coast. Physis A 1998, 55, 31–45. [Google Scholar]
- Amaral, F.D.; Hudson, M.M.; da Silveira, F.L.; Migotto, A.E.; Pinto, S.M.; Longo, L. Cnidarians of Saint Peter and St. Paul Archipelago, Northeast Brazil. In Proceedings of 9th International Coral Reef Symposium, Bali, Indonesia, 23–27 October 2000; International Coral Reef Society: Bali, Indonesia, 2002; pp. 567–572. [Google Scholar]
- Malpezzi, E.L.A.; Defreitas, J.C.; Muramoto, K.; Kamiya, H. Characterization of Peptides in Sea-Anemone Venom Collected by a Novel Procedure. Toxicon 1993, 31, 853–864. [Google Scholar] [CrossRef]
- Zaharenko, A.J.; Ferreira, W.A., Jr.; Oliveira, J.S.; Richardson, M.; Pimenta, D.C.; Konno, K.; Portaro, F.C.; de Freitas, J.C. Proteomics of the neurotoxic fraction from the sea anemone Bunodosoma cangicum venom: Novel peptides belonging to new classes of toxins. Comp. Biochem. Physiol. Part D Genomics Proteomics 2008, 3, 219–225. [Google Scholar] [CrossRef]
- Zaharenko, A.J.; Picolo, G.; Ferreira, W.A., Jr.; Murakami, T.; Kazuma, K.; Hashimoto, M.; Cury, Y.; de Freitas, J.C.; Satake, M.; Konno, K. Bunodosine 391: An analgesic acylamino acid from the venom of the sea anemone Bunodosoma cangicum. J. Nat. Prod. 2011, 74, 378–382. [Google Scholar] [CrossRef]
- Lagos, P.; Duran, R.; Cervenansky, C.; Freitas, J.C.; Silveira, R. Identification of hemolytic and neuroactive fractions in the venom of the sea anemone Bunodosoma cangicum. Braz. J. Med. Biol. Res. 2001, 34, 895–902. [Google Scholar] [CrossRef]
- Martins, R.D.; Alves, R.S.; Martins, A.M.; Barbosa, P.S.; Evangelista, J.S.; Evangelista, J.J.; Ximenes, R.M.; Toyama, M.H.; Toyama, D.O.; Souza, A.J.; et al. Purification and characterization of the biological effects of phospholipase A(2) from sea anemone Bunodosoma caissarum. Toxicon 2009, 54, 413–420. [Google Scholar] [CrossRef]
- Anderluh, G.; Macek, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Redaelli, E.; Zaharenko, A.J.; Cassulini, R.R.; Konno, K.; Pimenta, D.C.; Freitas, J.C.; Clare, J.J.; Wanke, E. Binding specificity of sea anemone toxins to Na(v)1.1–1.6 sodium channels—Unexpected contributions from differences in the IV/S3-S4 outer loop. J. Biol. Chem. 2004, 279, 33323–33335. [Google Scholar] [CrossRef]
- De Oliveira, J.S.; Zaharenko, A.J.; de Freitas, J.C.; Konno, K.; de Andrade, S.A.; Portaro, F.C.; Richardson, M.; Sant’anna, O.A.; Tambourgi, D.V. Caissarolysin I (Bcs I), a new hemolytic toxin from the Brazilian sea anemone Bunodosoma caissarum: purification and biological characterization. Biochim. Biophys. Acta 2006, 1760, 453–461. [Google Scholar] [CrossRef]
- Malpezzi, E.L.A.; Freitas, J.C. Hemolytic-Activity of the Nematocyst Venom from the Sea-Anemone Bunodosoma-Caissarum. Braz. J. Med. Biol. Res. 1991, 24, 1245–1249. [Google Scholar]
- Anderluh, G.; Sepcic, K.; Turk, T.; Macek, P. Cytolytic Proteins from Cnidarians—an Overview. Acta Chim. Slov. 2011, 58, 724–729. [Google Scholar]
- Anderluh, G.; Barlic, A.; Podlesek, Z.; Macek, P.; Pungercar, J.; Gubensek, F.; Zecchini, M.L.; Serra, M.D.; Menestrina, G. Cysteine-scanning mutagenesis of an eukaryotic pore-forming toxin from sea anemone: Topology in lipid membranes. Eur. J. Biochem. 1999, 263, 128–136. [Google Scholar] [CrossRef]
- Gunthorpe, L.; Cameron, A.M. Widespread but Variable Toxicity in Scleractinian Corals. Toxicon 1990, 28, 1199–1219. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Zaharenko, A.J.; Ferreira, W.A., Jr.; Konno, K.; Shida, C.S.; Richardson, M.; Lucio, A.D.; Beirao, P.S.; de Freitas, J.C. BcIV, a new paralyzing peptide obtained from the venom of the sea anemone Bunodosoma caissarum. A comparison with the Na+ channel toxin BcIII. Biochim. Biophys. Acta 2006, 1764, 1592–1600. [Google Scholar] [CrossRef]
- Moran, Y.; Weinberger, H.; Sullivan, J.C.; Reitzel, A.M.; Finnerty, J.R.; Gurevitz, M. Concerted evolution of sea anemone neurotoxin genes is revealed through analysis of the Nematostella vectensis genome. Mol. Biol. Evol. 2008, 25, 737–747. [Google Scholar] [CrossRef]
- Moran, Y.; Gurevitz, M. When positive selection of neurotoxin genes is missing—The riddle of the sea anemone Nematostella vectensis. FEBS J. 2006, 273, 3886–3892. [Google Scholar] [CrossRef]
- Darling, J.A.; Reitzel, A.M.; Finnerty, J.R. Regional population structure of a widely introduced estuarine invertebrate: Nematostella vectensis Stephenson in New England. Mol. Ecol. 2004, 13, 2969–2981. [Google Scholar] [CrossRef]
- Reitzel, A.M.; Darling, J.A.; Sullivan, J.C.; Finnerty, J.R. Global population genetic structure of the starlet anemone Nematostella vectensis: Multiple introductions and implications for conservation policy. Biol. Invasions 2008, 10, 1197–1213. [Google Scholar] [CrossRef]
- Nei, M.; Rooney, A.P. Concerted and birth-and-death evolution of multigene families. Annu. Rev. Genet. 2005, 39, 121–152. [Google Scholar] [CrossRef]
- Duda, T.F., Jr.; Chang, D.; Lewis, B.D.; Lee, T. Geographic variation in venom allelic composition and diets of the widespread predatory marine gastropod Conus ebraeus. PLoS One 2009, 4, e6245. [Google Scholar]
- Jakubowski, J.A.; Kelley, W.P.; Sweedler, J.V.; Gilly, W.F.; Schulz, J.R. Intraspecific variation of venom injected by fish-hunting Conus snails. J. Exp. Biol. 2005, 208, 2873–2883. [Google Scholar] [CrossRef]
- Ferreira Junior, R.S.; Sciani, J.M.; Marques-Porto, R.; Junior, A.L.; Orsi Rde, O.; Barraviera, B.; Pimenta, D.C. Africanized honey bee (Apis mellifera) venom profiling: Seasonal variation of melittin and phospholipase A(2) levels. Toxicon 2010, 56, 355–362. [Google Scholar] [CrossRef]
- Brand, J.M.; Blum, M.S.; Barlin, M.R. Fire ant venoms: Intraspecific and interspecific variation among castes and individuals. Toxicon 1973, 11, 325–331. [Google Scholar] [CrossRef]
- Deslippe, R.J.; Guo, Y.J. Venom alkaloids of fire ants in relation to worker size and age. Toxicon 2000, 38, 223–232. [Google Scholar] [CrossRef]
- Herzig, V.; John Ward, R.; Ferreira dos Santos, W. Intersexual variations in the venom of the Brazilian “armed” spider Phoneutria nigriventer (Keyserling, 1891). Toxicon 2002, 40, 1399–1406. [Google Scholar] [CrossRef]
- Binford, G.J. An analysis of geographic and intersexual chemical variation in venoms of the spider Tegenaria agrestis (Agelenidae). Toxicon 2001, 39, 955–968. [Google Scholar] [CrossRef]
- Newton, K.A.; Clench, M.R.; Deshmukh, R.; Jeyaseelan, K.; Strong, P.N. Mass fingerprinting of toxic fractions from the venom of the Indian red scorpion, Mesobuthus tamulus: Biotope-specific variation in the expression of venom peptides. Rapid Commun. Mass Spectrom. 2007, 21, 3467–3476. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Omran, M.A.; Abdel-Nabi, I.M.; Ueda, H.; McVean, A. Intraspecific variation in the Egyptian scorpion Scorpio maurus palmatus venom collected from different biotopes. Toxicon 2009, 53, 349–359. [Google Scholar] [CrossRef]
- Gibbs, H.L.; Sanz, L.; Calvete, J.J. Snake population venomics: proteomics-based analyses of individual variation reveals significant gene regulation effects on venom protein expression in Sistrurus rattlesnakes. J. Mol. Evol. 2009, 68, 113–125. [Google Scholar] [CrossRef]
- Nunez, V.; Cid, P.; Sanz, L.; de La Torre, P.; Angulo, Y.; Lomonte, B.; Gutierrez, J.M.; Calvete, J.J. Snake venomics and antivenomics of Bothrops atrox venoms from Colombia and the Amazon regions of Brazil, Peru and Ecuador suggest the occurrence of geographic variation of venom phenotype by a trend towards paedomorphism. J. Proteomics 2009, 73, 57–78. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Abdel-Nabi, I.M.; El-Naggar, M.S.; Abbas, O.A.; Strong, P.N. Intraspecific variation in the venom of the vermivorous cone snail Conus vexillum. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 318–325. [Google Scholar]
- Dutertre, S.; Biass, D.; Stocklin, R.; Favreau, P. Dramatic intraspecimen variations within the injected venom of Conus consors: An unsuspected contribution to venom diversity. Toxicon 2010, 55, 1453–1462. [Google Scholar] [CrossRef]
- Pimenta, A.M.; de Marco Almeida, F.; de Lima, M.E.; Martin-Eauclaire, M.F.; Bougis, P.E. Individual variability in Tityus serrulatus (Scorpiones, Buthidae) venom elicited by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 413–418. [Google Scholar] [CrossRef]
- Kalapothakis, E.; Chavez-Olortegui, C. Venom variability among several Tityus serrulatus specimens. Toxicon 1997, 35, 1523–1529. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A. Intraspecific diversity of scorpions’ venom and its implication on the pathophysiological effects. J. Venom. Anim. Toxins Incl. Trop. Dis. Thesis 2008, 14, 191–192. [Google Scholar] [CrossRef]
- Andrade, D.V.; Abe, A.S. Relationship of venom ontogeny and diet in Bothrops. Herpetologica 1999, 55, 200–204. [Google Scholar]
- MacKessy, S.P.; Williams, K.; Ashon, K.G. Ontogenetic variation in venom composition and diet of Crotalus oreganus concolor: A case of venom paedomorphosis? Copeia 2003, 2003, 769–782. [Google Scholar] [CrossRef]
- Daltry, J.C.; Wuster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef]
- Cotton, J.; Crest, M.; Bouet, F.; Alessandri, N.; Gola, M.; Forest, E.; Karlsson, E.; Castaneda, O.; Harvey, A.L.; Vita, C.; Menez, A. A potassium-channel toxin from the sea anemone Bunodosoma granulifera, an inhibitor for Kv1 channels. Revision of the amino acid sequence, disulfide-bridge assignment, chemical synthesis, and biological activity. Eur. J. Biochem. 1997, 244, 192–202. [Google Scholar]
- Castaneda, O.; Sotolongo, V.; Amor, A.M.; Stocklin, R.; Anderson, A.J.; Harvey, A.L.; Engstrom, A.; Wernstedt, C.; Karlsson, E. Characterization of a potassium channel toxin from the Caribbean Sea anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef]
- Gendeh, G.S.; Young, L.C.; de Medeiros, C.L.; Jeyaseelan, K.; Harvey, A.L.; Chung, M.C. A new potassium channel toxin from the sea anemone Heteractis magnifica: Isolation, cDNA cloning, and functional expression. Biochemistry 1997, 36, 11461–11471. [Google Scholar]
- Schweitz, H.; Bruhn, T.; Guillemare, E.; Moinier, D.; Lancelin, J.M.; Beress, L.; Lazdunski, M. Kalicludines and kaliseptine. Two different classes of sea anemone toxins for voltage sensitive K+ channels. J. Biol. Chem. 1995, 270, 25121–25126. [Google Scholar]
- Minagawa, S.; Ishida, M.; Nagashima, Y.; Shiomi, K. Primary structure of a potassium channel toxin from the sea anemone Actinia equina. FEBS Lett. 1998, 427, 149–151. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Honma, T.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Isolation and cDNA cloning of a potassium channel peptide toxin from the sea anemone Anemonia erythraea. Toxicon 2006, 48, 536–542. [Google Scholar] [CrossRef]
- Sabourault, C.; Ganot, P.; Deleury, E.; Allemand, D.; Furla, P. Comprehensive EST analysis of the symbiotic sea anemone, Anemonia viridis. BMC Genomics 2009, 10, 333. [Google Scholar]
- Dauplais, M.; Lecoq, A.; Song, J.X.; Cotton, J.; Jamin, N.; Gilquin, B.; Roumestand, C.; Vita, C.; deMedeiros, C.L.C.; Rowan, E.G.; Harvey, A.L.; Menez, A. On the convergent evolution of animal toxins—Conservation of a diad of functional residues in potassium channel-blocking toxins with unrelated structures. J. Biol. Chem. 1997, 272, 4302–4309. [Google Scholar] [CrossRef]
- Sabatier, J.M.; Jouirou, B.; Mouhat, S.; Andreotti, N.; de Waard, M. Toxin determinants required for interaction with voltage-gated K+ channels. Toxicon 2004, 43, 909–914. [Google Scholar] [CrossRef]
- Pennington, M.W.; Kem, W.R.; Mahnir, V.M.; Byrnes, M.E.; Zaydenberg, I.; Khaytin, I.; Krafte, D.S.; Hill, R. Identification of essential residues in the potassium channel inhibitor ShK toxin: Analysis of monosubstituted analogs. In Peptides: Chemistry, Structure and Biology; Kaumaya, P.T.P., Hodges, R.S., Eds.; Escom: Leiden, Netherlands, 1995; pp. 14–16. [Google Scholar]
- Gems, D.; Ferguson, C.J.; Robertson, B.D.; Nieves, R.; Page, A.P.; Blaxter, M.L.; Maizels, R.M. An abundant, trans-spliced mRNA from Toxocara canis infective larvae encodes a 26-kDa protein with homology to phosphatidylethanolamine-binding proteins. J. Biol. Chem. 1995, 270, 18517–18522. [Google Scholar]
- Gems, D.; Maizels, R.M. An abundantly expressed mucin-like protein from Toxocara canis infective larvae: The precursor of the larval surface coat glycoproteins. Proc. Natl. Acad. Sci. USA 1996, 93, 1665–1670. [Google Scholar] [CrossRef]
- Blaxter, M. Caenorhabditis elegans is a nematode. Science 1998, 282, 2041–2046. [Google Scholar] [CrossRef]
- Daub, J.; Loukas, A.; Pritchard, D.I.; Blaxter, M. A survey of genes expressed in adults of the human hookworm, Necator americanus. Parasitology 2000, 120((Pt. 2)), 171–184. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; Renjifo, C.; de la Vega, R.C. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Pennington, M.W.; Byrnes, M.E.; Zaydenberg, I.; Khaytin, I.; de Chastonay, J.; Krafte, D.S.; Hill, R.; Mahnir, V.M.; Volberg, W.A.; Gorczyca, W.; et al. Chemical synthesis and characterization of ShK toxin: A potent potassium channel inhibitor from a sea anemone. Int. J. Pept. Protein Res. 1995, 46, 354–358. [Google Scholar]
- Kalman, K.; Pennington, M.W.; Lanigan, M.D.; Nguyen, A.; Rauer, H.; Mahnir, V.; Paschetto, K.; Kem, W.R.; Grissmer, S.; Gutman, G.A.; et al. ShK-Dap22, a potent Kv1.3-specific immunosuppressive polypeptide. J. Biol. Chem. 1998, 273, 32697–32707. [Google Scholar] [CrossRef]
- Yan, L.Z.; Herrington, J.; Goldberg, E.; Dulski, P.M.; Bugianesi, R.M.; Slaughter, R.S.; Banerjee, P.; Brochu, R.M.; Priest, B.T.; Kaczorowski, G.J.; et al. Stichodactyla helianthus peptide, a pharmacological tool for studying Kv3.2 channels. Mol. Pharmacol. 2005, 67, 1513–1521. [Google Scholar] [CrossRef]
- Rauer, H.; Pennington, M.; Cahalan, M.; Candı, K.G. Structural conservation of the pores of calcium-activated and voltage-gated potassium channels determined by a sea anemone toxin. J. Biol. Chem. 1999, 274, 21885–21892. [Google Scholar]
- Sabatier, J.M.; Mouhat, S.; Andreotti, N.; Jouirou, B. Animal toxins acting on voltage-gated potassium channels. Curr. Pharm. Des. 2008, 14, 2503–2518. [Google Scholar] [CrossRef]
- Peigneur, S.; Billen, B.; Derua, R.; Waelkens, E.; Debaveye, S.; Beress, L.; Tytgat, J. A bifunctional sea anemone peptide with Kunitz type protease and potassium channel inhibiting properties. Biochem. Pharmacol. 2011, 82, 81–90. [Google Scholar]
- Sabatier, J.M.; Mouhat, S.; Mosbah, A.; Visan, V.; Wulff, H.; Delepierre, M.; Darbon, H.; Grissmer, S.; de Waard, M. The “functional” dyad of scorpion toxin Pi1 is not itself a prerequisite for toxin binding to the voltage-gated Kv1.2 potassium channels. Biochem. J. 2004, 377, 25–36. [Google Scholar] [CrossRef]
- Batista, C.V.F.; Gomez-Lagunas, F.; de la Vega, R.C.R.; Hajdu, P.; Panyi, G.; Gaspar, R.; Possani, L.D. Two novel toxins from the Amazonian scorpion Tityus cambridgei that block Kv1.3 and Shaker BK+-channels with distinctly different affinities. Biochim. Biophys. Acta 2002, 1601, 123–131. [Google Scholar] [CrossRef]
- Rodriguez de la Vega, R.C.; Possani, L.D. Current views on scorpion toxins specific for K+-channels. Toxicon 2004, 43, 865–875. [Google Scholar] [CrossRef]
- Rogowski, R.S.; Collins, J.H.; ONeill, T.J.; Gustafson, T.A.; Werkman, T.R.; Rogawski, M.A.; Tenenholz, T.C.; Weber, D.J.; Blaustein, M.P. Three new toxins from the scorpion Pandinus imperator selectively block certain voltage-gated K+ channels. Mol. Pharmacol. 1996, 50, 1167–1177. [Google Scholar]
- Garcia-Calvo, M.; Leonard, R.J.; Novick, J.; Stevens, S.P.; Schmalhofer, W.; Kaczorowski, G.J.; Garcia, M.L. Purification, characterization, and biosynthesis of margatoxin, a component of Centruroides margaritatus venom that selectively inhibits voltage-dependent potassium channels. J. Biol. Chem. 1993, 268, 18866–18874. [Google Scholar]
- Tudor, J.E.; Pallaghy, P.K.; Pennington, M.W.; Norton, R.S. Solution structure of ShK toxin, a novel potassium channel inhibitor from a sea anemone. Nat. Struct. Biol. 1996, 3, 317–320. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Macarthur, M.W.; Moss, D.S.; Thornton, J.M. Procheck—a Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Ramachandran, G.N.; Ramakrishnan, C.; Sasisekharan, V. Stereochemistry of polypeptide chain configurations. J. Mol. Biol. 1963, 7, 95–99. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method—a New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Honma, T.; Kawahata, S.; Ishida, M.; Nagai, H.; Nagashima, Y.; Shiomi, K. Novel peptide toxins from the sea anemone Stichodactyla haddoni. Peptides 2008, 29, 536–544. [Google Scholar] [CrossRef]
- Sali, A.; Blundell, T.L. Comparative Protein Modeling by Satisfaction of Spatial Restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Cheng, J.; Randall, A.Z.; Sweredoski, M.J.; Baldi, P. SCRATCH: A protein structure and structural feature prediction server. Nucleic Acids Res. 2005, 33, W72–W76. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Orts, D.J.B.; Peigneur, S.; Madio, B.; Cassoli, J.S.; Montandon, G.G.; Pimenta, A.M.C.; Bicudo, J.E.P.W.; Freitas, J.C.; Zaharenko, A.J.; Tytgat, J. Biochemical and Electrophysiological Characterization of Two Sea Anemone Type 1 Potassium Toxins from a Geographically Distant Population of Bunodosoma caissarum. Mar. Drugs 2013, 11, 655-679. https://doi.org/10.3390/md11030655
Orts DJB, Peigneur S, Madio B, Cassoli JS, Montandon GG, Pimenta AMC, Bicudo JEPW, Freitas JC, Zaharenko AJ, Tytgat J. Biochemical and Electrophysiological Characterization of Two Sea Anemone Type 1 Potassium Toxins from a Geographically Distant Population of Bunodosoma caissarum. Marine Drugs. 2013; 11(3):655-679. https://doi.org/10.3390/md11030655
Chicago/Turabian StyleOrts, Diego J. B., Steve Peigneur, Bruno Madio, Juliana S. Cassoli, Gabriela G. Montandon, Adriano M. C. Pimenta, José E. P. W. Bicudo, José C. Freitas, André J. Zaharenko, and Jan Tytgat. 2013. "Biochemical and Electrophysiological Characterization of Two Sea Anemone Type 1 Potassium Toxins from a Geographically Distant Population of Bunodosoma caissarum" Marine Drugs 11, no. 3: 655-679. https://doi.org/10.3390/md11030655
APA StyleOrts, D. J. B., Peigneur, S., Madio, B., Cassoli, J. S., Montandon, G. G., Pimenta, A. M. C., Bicudo, J. E. P. W., Freitas, J. C., Zaharenko, A. J., & Tytgat, J. (2013). Biochemical and Electrophysiological Characterization of Two Sea Anemone Type 1 Potassium Toxins from a Geographically Distant Population of Bunodosoma caissarum. Marine Drugs, 11(3), 655-679. https://doi.org/10.3390/md11030655