
Physical Activity and Post-Transcriptional Regulation of Aging Decay: Modulation of Pathways in Postmenopausal Osteoporosis



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Fate of MSCs: Age-Related Alterations
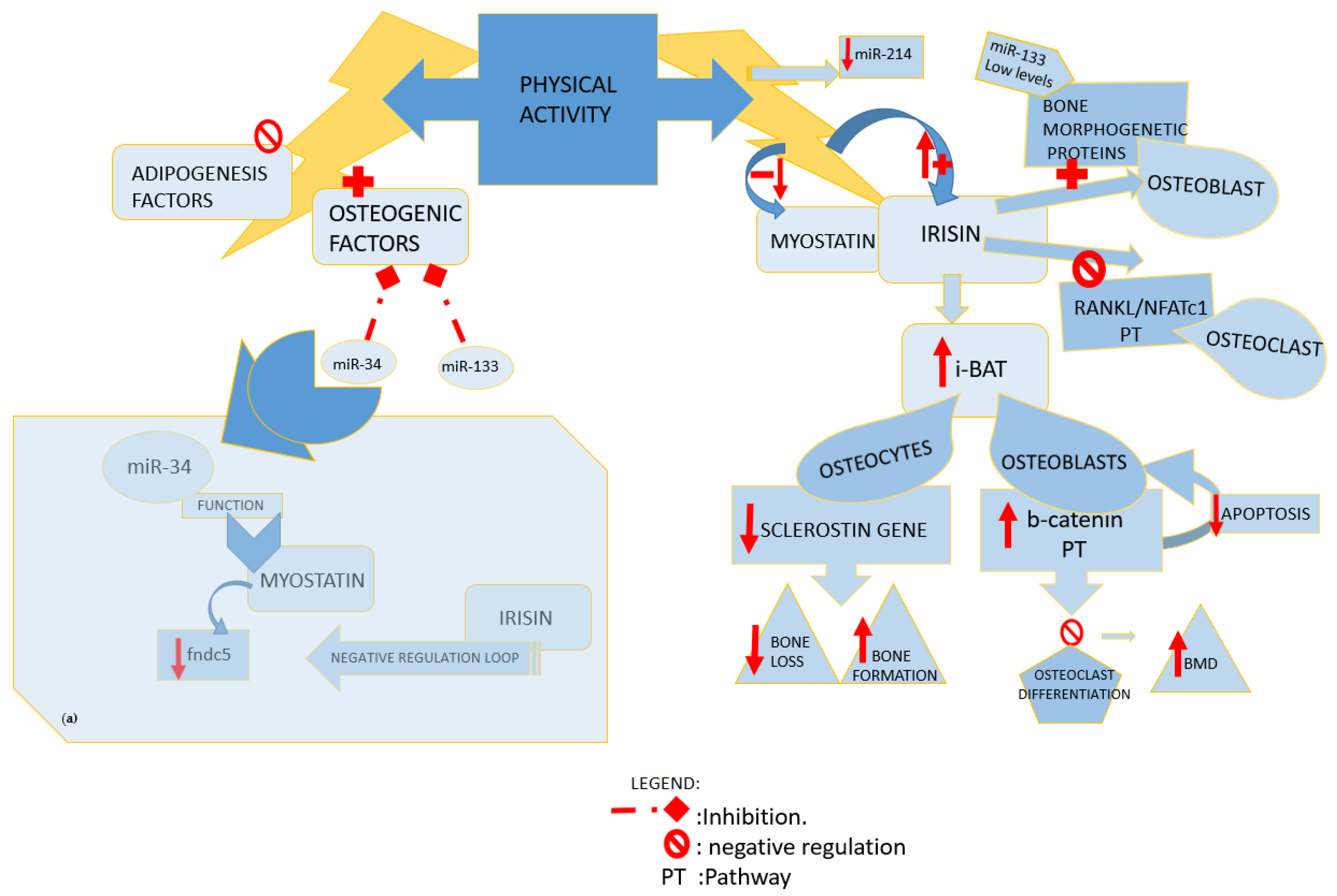
Fat Tissue and Bone-Muscle Unit: Physical Activity and Irisin, Myostatin, Sclerostin Crosstalk
4. Influence of Physical Activity on Post-Transcriptional Regulation of Fat, Muscle, and Bone Tissue Pathways
4.1. Impact of Different Types of Exercise on Bone Metabolism

{kind=link}
{kind=link}
{kind=link}
| Reference Year Author | Article Type | Type of Sample and Analysis | Physical Activity Type and Duration | Pathways Involvement | Effects |
|---|---|---|---|---|---|
| [31] 2019 Dalle Carbonare L. et al. | Article | -H.C. -n.22 M healthy and regularly active -Vitro analyses before and post run | -Half marathon performance | Upregulation of osteogenesis related genes Downregulation of Adipogenic commitment | Post activity increases sera levels of: RUNX2, MSX1, SPP1, BMP2, BMP6, and PPARγ gene Exercise counteracts chronic degenerative conditions |
| [30] 2020 Tobeiha, M. et al. | Review 21 studies | In 4 studies -O, F; -H/A | In most of the studies: high-intensity exercise | RANKL/RANK/ OPG pathway | Increases level of: -OPG Decreases level of: -RANKL Exercise promotes bone health |
| [35] 2019 Kang, Y.S. et al. | Article | -n.20 A. | -Swimming exercise. -16 weeks study period | Bone metabolism marker pathways beta-catenin | Increases level of: -Serum irisin - Bone tissue PGC-1α and FNDC5 |
| [36] 2015 Kim, H.J. et al. | Control study | -n.28 obese H, M/F -Y -3 h following acute HIIT | -HIIT vs. MICE. -8 weeks of exercise program (60 min/day, 5 times a week) | HIIT increases: -Fat oxidation during and following exercise -PGC-1α mRNA -Serum irisin level | |
| [37] 2021 Liu, Y. et al. | Control study | -n.40 O, A. | -High-intensity interval static training -8 weeks | Regulation of PGC1α/FNDC5/UCP1 signaling pathway | Increased levels of: -Serum irisin improvement in motor function of aged rats |
| [38] 2017 Eaton, M. et al. | Article | -n.10 H, Y, M. -Valuation before and after activity | -HIIT -20-day period of twice-daily | acute regulation of the mRNA myokine, interleukin-6, and FNDC5 | Increase in: -FNDC5 mRNA expression |
| [41] 2018 H. Shirvani and Arabzadeh, E. | Article | -n.32 A, M. | -HIIT vs. MICE -8 weeks | Crosstalk between skeletal muscle and adipose tissue | Increase in: -PGC-1α gene transcription; -Serum irisin |
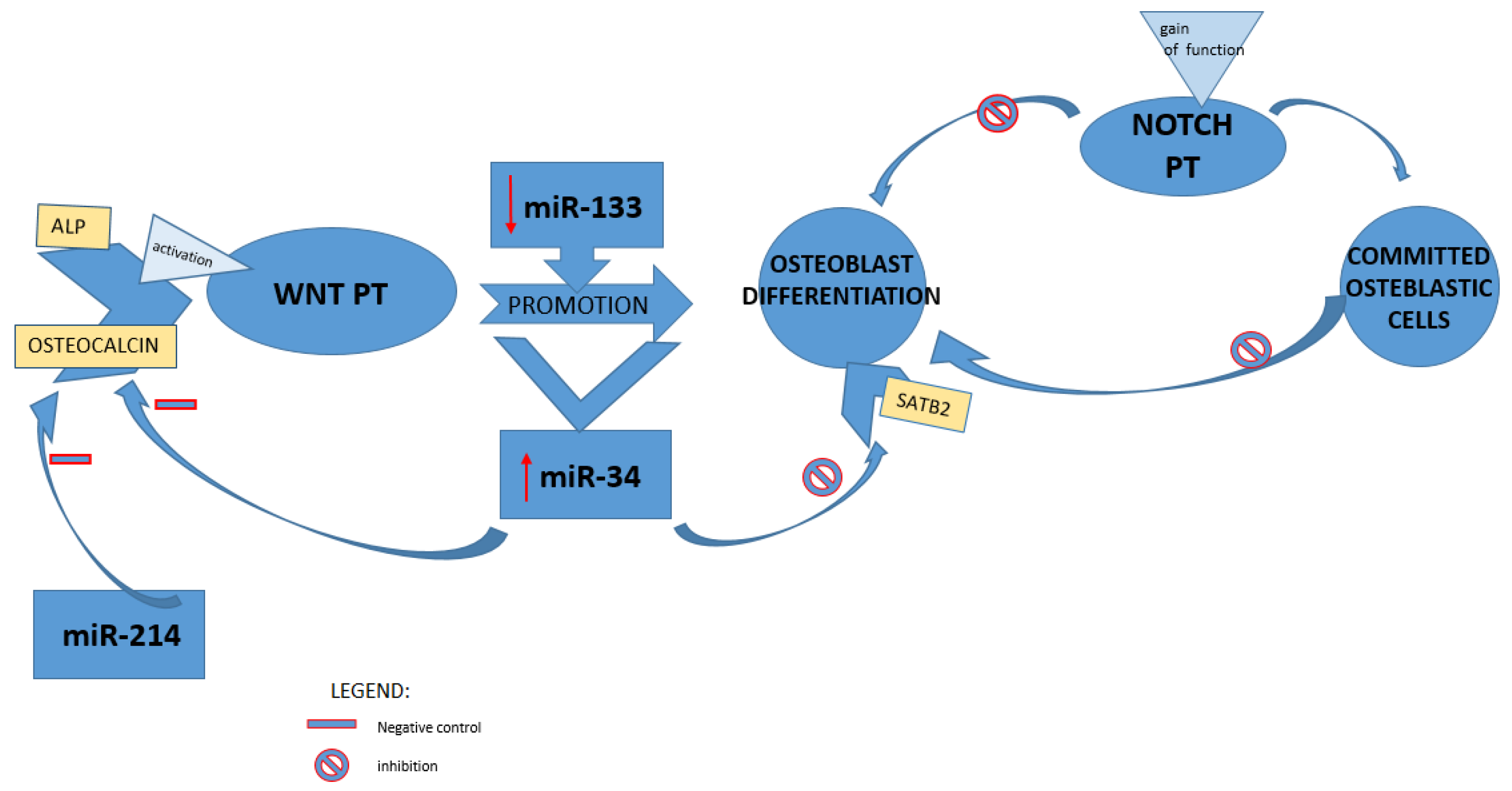
4.2. Involvement of Different Signaling Pathways and Related miRNAs in Bone Loss
4.3. Physical Activity and miRNAs Modulation in Prevent of Bone Loss: A Prospective Therapeutic Target
4.4. Possible Current Therapeutic Management of Postmenopausal Osteoporosis
5. Discussion
5.1. Pleiotropic Effect of Physical Activity on Altered Metabolism Age-Related
5.2. Influence of Different Types of Physical Activity
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Qi, Z.; Liu, W.; Lu, J. The mechanisms underlying the beneficial effects of exercise on bone remodeling: Roles of bone-derived cytokines and microRNAs. Prog. Biophys. Mol. Biol. 2016, 122, 131–139. [Google Scholar] [CrossRef]
- Colaianni, G.; Mongelli, T.; Colucci, S.; Cinti, S.; Grano, M. Crosstalk Between Muscle and Bone Via the Muscle-Myokine Irisin. Curr. Osteoporos. Rep. 2016, 14, 132–137. [Google Scholar] [CrossRef]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P. A PGC1α-dependent myokine that drives browning of white fat. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Dufrane, D. Impact of Age on Human Adipose Stem Cells for Bone Tissue Engineering. Cell Transplant. 2017, 26, 1496–1504. [Google Scholar] [CrossRef]
- Yen, K.L.; Capilla, E.; Rosen, C.J.; Gilsanz, V.; Pessin, J.E.; Judex, S.; Rubin, C.T. Mechanical stimulation of mesenchymal stem cell proliferation and differentiation promotes osteogenesis while preventing dietary-induced obesity. J. Bone Miner. Res. 2009, 24, 50–61. [Google Scholar] [CrossRef]
- David, V.; Martin, A.; Lafage-Proust, M.H.; Malaval, L.; Peyroche, S.; Jones, D.B.; Vico, L.; Guignandon, A. Mechanical loading down-regulates peroxisome proliferator-activated receptor γ in bone marrow stromal cells and favors osteoblastogenesis at the expense of adipogenesis. Endocrinology 2007, 148, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Valenti, M.T.; Deiana, M.; Cheri, S.; Dotta, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Dalle Carbonare, L.; Mottes, M. Physical Exercise Modulates miR-21-5p, miR-129-5p, miR-378-5p, and miR-188-5p Expression in Progenitor Cells Promoting Osteogenesis. Cells 2019, 8, 742. [Google Scholar] [CrossRef] [Green Version]
- An, J.H.; Ohn, J.H.; Song, J.A.; Yang, J.Y.; Park, H.; Choi, H.J.; Kim, S.W.; Kim, S.Y.; Park, W.Y.; Shin, C.S. Changes of microRNA profile and microRNA-mRNA regulatory network in bones of ovariectomized mice. J. Bone Miner. Res. 2014, 29, 644–656. [Google Scholar] [CrossRef]
- Colaianni, G.; Cinti, S.; Colucci, S.; Grano, M. Irisin and musculoskeletal health. Ann. N. Y. Acad. Sci. 2017, 1402, 5–9. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, Y.; Qiao, X.; Zeng, R.; Cheng, R.; Nie, Y.; Li, S.; A, R.; Shen, X.; Yang, M.; et al. Irisin ameliorates bone loss in ovariectomized mice. Climacteric 2020, 23, 496–504. [Google Scholar] [CrossRef]
- Yan, J.; Liu, H.J.; Guo, W.C.; Yang, J. Low serum concentrations of Irisin are associated with increased risk of hip fracture in Chinese older women. Jt. Bone Spine 2018, 85, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Farsani, Z.H.; Banitalebi, E.; Faramarzi, M.; Bigham-Sadegh, A. Effects of different intensities of strength and endurance training on some osteometabolic miRNAs, Runx2 and PPARγ in bone marrow of old male wistar rats. Mol. Biol. Rep. 2019, 46, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Moore, B.T.; Peng, X.H.; Fang, X.; Lappe, J.M.; Recker, R.R.; Xiao, P. Mir-133a in human circulating monocytes: A potential biomarker associated with postmenopausal osteoporosis. PLoS ONE 2012, 7, e34641. [Google Scholar] [CrossRef] [PubMed]
- Bottani, M.; Banfi, G.; Lombardi, G. Perspectives on mirnas as epigenetic markers in osteoporosis and bone fracture risk: A step forward in personalized diagnosis. Front. Genet. 2019, 10, 1044. [Google Scholar] [CrossRef]
- Rosen, C.J.; Ackert-Bicknell, C.; Rodriguez, J.P.; Pino, A.M. Marrow fat and the bone microenvironment: Developmental, functional, and pathological implications. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 109–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, G.M.; Griffith, J.F.; Yeung, D.K.W.; Leung, P.C.; Fogelman, I. Effect of increasing vertebral marrow fat content on BMD measurement, T-Score status and fracture risk prediction by DXA. Bone 2009, 44, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, A.; Rivas, D.; Duque, G. Effect of estrogens on bone marrow adipogenesis and Sirt1 in aging C57BL/6J mice. Biogerontology 2009, 10, 747–755. [Google Scholar] [CrossRef]
- Shockley, K.R.; Lazarenko, O.P.; Czernik, P.J.; Rosen, C.J.; Churchill, G.A.; Lecka-Czernik, B. PPARγ2 nuclear receptor controls multiple regulatory pathways of osteoblast differentiation from marrow mesenchymal stem cells. J. Cell. Biochem. 2009, 106, 232–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, S.; Bu, S.; Wang, Y.; Duan, Y.; Yang, S. Treadmill training prevents bone loss by inhibition of PPARγ expression but not promoting of Runx2 expression in ovariectomized rats. Eur. J. Appl. Physiol. 2011, 111, 1759–1767. [Google Scholar] [CrossRef]
- Cho, S.W.; Yang, J.Y.; Her, S.J.; Choi, H.J.; Jung, J.Y.; Sun, H.J.; An, J.H.; Cho, H.Y.; Kim, S.W.; Park, K.S.; et al. Osteoblast-targeted overexpression of PPARγ inhibited bone mass gain in male mice and accelerated ovariectomy-induced bone loss in female mice. J. Bone Miner. Res. 2011, 26, 1939–1952. [Google Scholar] [CrossRef]
- Restuccia, R.; Perani, F.; Ficarra, G.; Trimarchi, F.; Bitto, A.; di Mauro, D. Irisin and Vascular Inflammation: Beneficial Effects of a Healthy Lifestyle Beyond Physical Activity. Curr. Pharm. Des. 2021, 27, 2151–2155. [Google Scholar] [CrossRef] [PubMed]
- Anastasilakis, A.D.; Polyzos, S.A.; Makras, P.; Gkiomisi, A.; Bisbinas, I.; Katsarou, A.; Filippaios, A.; Mantzoros, C.S. Circulating irisin is associated with osteoporotic fractures in postmenopausal women with low bone mass but is not affected by either teriparatide or denosumab treatment for 3 months. Osteoporos. Int. 2014, 25, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, H.C.; Zhang, D.; Yeom, H.; Lim, S.K. The novel myokine irisin: Clinical implications and potential role as a biomarker for sarcopenia in postmenopausal women. Endocrine 2019, 64, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Palermo, A.; Strollo, R.; Maddaloni, E.; Tuccinardi, D.; D’Onofrio, L.; Briganti, S.I.; Defeudis, G.; De Pascalis, M.; Lazzaro, M.C.; Colleluori, G.; et al. Irisin is associated with osteoporotic fractures independently of bone mineral density, body composition or daily physical activity. Clin. Endocrinol. 2015, 82, 615–619. [Google Scholar] [CrossRef]
- Zhou, K.; Qiao, X.; Cai, Y.; Li, A.; Shan, D. Lower circulating irisin in middle-aged and older adults with osteoporosis: A systematic review and meta-analysis. Menopause 2019, 26, 1302–1310. [Google Scholar] [CrossRef]
- Yuki, A.; Yotani, K.; Tamaki, H.; Kasuga, N.; Takekura, H. Upregulation of osteogenic factors induced by high-impact jumping suppresses adipogenesis in marrow but not adipogenic transcription factors in rat tibiae. Eur. J. Appl. Physiol. 2010, 109, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Shan, T.; Liang, X.; Bi, P.; Kuang, S. Myostatin knockout drives browning of white adipose tissue through activating the AMPK-PGC1-Fndc5 pathway in muscle. FASEB J. 2013, 27, 1981–1989. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, M.G.; Hamilton, D.L.; Pepin, M.; Patton, A.; Baar, K. Inhibition of Myostatin Signaling through Notch Activation following Acute Resistance Exercise. PLoS ONE 2013, 8, e68743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.; Sathiakumar, D.; Lua, B.J.G.; Kukreti, H.; Lee, M.; Mcfarlane, C. Myostatin signals through miR-34a to regulate Fndc5 expression and browning of white adipocytes. Int. J. Obes. 2017, 41, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobeiha, M.; Moghadasian, M.H.; Amin, N.; Jafarnejad, S. RANKL/RANK/OPG Pathway: A Mechanism Involved in Exercise-Induced Bone Remodeling. BioMed Res. Int. 2020, 2020, 6910312. [Google Scholar] [CrossRef] [Green Version]
- Dalle Carbonare, L.; Mottes, M.; Cheri, S.; Deiana, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Salvagno, G.L.; Lippi, G.; Valenti, M.T. Increased Gene Expression of RUNX2 and SOX9 in Mesenchymal Circulating Progenitors Is Associated with Autophagy during Physical Activity. Oxid. Med. Cell. Longev. 2019, 2019, 8426259. [Google Scholar] [CrossRef] [PubMed]
- Singulani, M.P.; Stringhetta-Garcia, C.T.; Santos, L.F.; Morais, S.R.L.; Louzada, M.J.Q.; Oliveira, S.H.P.; Chaves Neto, A.H.; Dornelles, R.C.M. Effects of strength training on osteogenic differentiation and bone strength in aging female Wistar rats. Sci. Rep. 2017, 7, 42878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Valverde, P.; Zhu, X.; Murray, D.; Wu, Y.; Yu, L.; Jiang, H.; Dard, M.M.; Huang, J.; Xu, Z.; et al. Exercise-induced irisin in bone and systemic irisin administration reveal new regulatory mechanisms of bone metabolism. Bone Res. 2017, 5, 16056. [Google Scholar] [CrossRef] [Green Version]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Pignataro, P.; Buccoliero, C.; Liu, P.; Lu, P.; Sartini, L.; Comite, M.D.; Mori, G.; et al. The myokine irisin increases cortical bone mass. Proc. Natl. Acad. Sci. USA 2015, 112, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.S.; Kim, J.C.; Kim, J.S.; Kim, S.H. Effects of swimming exercise on serum irisin and bone FNDC5 in rat models of high-fat diet-induced osteoporosis. J. Sports Sci. Med. 2019, 18, 596–603. [Google Scholar]
- Kim, H.J.; Lee, H.J.; So, B.; Son, J.S.; Yoon, D.; Song, W. Effect of aerobic training and resistance training on circulating irisin level and their association with change of body composition in overweight/obese adults: A pilot study. Physiol. Res. 2016, 65, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, C.; Liu, S.; Zhang, S.; Mao, Y.; Fang, L. Eight weeks of high-intensity interval static strength training improves skeletal muscle atrophy and motor function in aged rats via the PGC-1α/FNDC5/UCP1 pathway. Clin. Interv. Aging 2021, 16, 811–821. [Google Scholar] [CrossRef]
- Eaton, M.; Granata, C.; Barry, J.; Safdar, A.; Bishop, D.; Little, J.P. Impact of a single bout of high-intensity interval exercise and short-term interval training on interleukin-6, FNDC5, and METRNL mRNA expression in human skeletal muscle. J. Sport Health Sci. 2018, 7, 191–196. [Google Scholar] [CrossRef]
- Turner, C.H.; Takano, Y.; Owan, I. Aging changes mechanical loading thresholds for bone formation in rats. J. Bone Miner. Res. 1995, 10, 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Hettchen, M.; von Stengel, S.; Kohl, M.; Murphy, M.H.; Shojaa, M.; Ghasemikaram, M.; Bragonzoni, L.; Benvenuti, F.; Ripamonti, C.; Benedetti, M.G.; et al. Changes in menopausal risk factors in early postmenopausal osteopenic women after 13 months of high-intensity exercise: The randomized controlled ACTLIFE-RCT. Clin. Interv. Aging 2021, 16, 83–96. [Google Scholar] [CrossRef]
- Shirvani, H.; Arabzadeh, E. Metabolic cross-talk between skeletal muscle and adipose tissue in high-intensity interval training vs. moderate-intensity continuous training by regulation of PGC-1α. Eat. Weight Disord.-Stud. Anorex. Bulim. Obes. 2018, 25, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Encabo, P.; McNeil, J.; Boyne, D.J.; Courneya, K.S.; Friedenreich, C.M. Dose-response effects of exercise on bone mineral density and content in post-menopausal women. Scand. J. Med. Sci. Sports 2019, 29, 1121–1129. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, M.; Zhang, Q. The effectiveness of combined exercise interventions for preventing postmenopausal bone loss: A systematic review and meta-analysis. J. Orthop. Sports Phys. Ther. 2017, 47, 241–251. [Google Scholar] [CrossRef]
- Ju, Y.I.; Sone, T.; Ohnaru, K.; Tanaka, K.; Fukunaga, M. Effect of swimming exercise on three-dimensional trabecular bone microarchitecture in ovariectomized rats. J. Appl. Physiol. 2015, 119, 990–997. [Google Scholar] [CrossRef] [Green Version]
- Oh, T.; Tanaka, S.; Naka, T.; Igawa, S. Effects of high-intensity swimming training on the bones of ovariectomized rats. J. Exerc. Nutr. Biochem. 2016, 20, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Costa, A.M.; Palmeira-de-Oliveira, A.; Soares, J.; Monteiro, M.; Williams, J.H.H. The effects of combined training on bone metabolic markers in postmenopausal women. Sci. Sports 2016, 31, 152–157. [Google Scholar] [CrossRef]
- Mosti, M.P.; Kaehler, N.; Stunes, A.K.; Hoff, J.; Syversen, U. Maximal strength training in postmenopausal women with osteoporosis or osteopenia. J. Strength Cond. Res. 2013, 27, 2879–2886. [Google Scholar] [CrossRef]
- Gunendi, Z.; Ozyemisci-Taskiran, O.; Demirsoy, N. The effect of 4-week aerobic exercise program on postural balance in postmenopausal women with osteoporosis. Rheumatol. Int. 2008, 28, 1217–1222. [Google Scholar] [CrossRef]
- Mittlmeier, T. Interval Training Is Not Superior to Endurance Training With Respect to Bone Accrual of Ovariectomized Mice Mice and Experimental Design. Front. Physiol. 2020, 11, 1096. [Google Scholar] [CrossRef]
- Wei, J.; Shi, Y.; Zheng, L.; Zhou, B.; Inose, H.; Wang, J.; Guo, X.E.; Grosschedl, R.; Karsenty, G. miR-34s inhibit osteoblast proliferation and differentiation in the mouse by targeting SATB2. J. Cell Biol. 2012, 197, 509–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.M.; Nakashima, A.; Nashimoto, M.; Yawaka, Y.; Tamura, M. Bone morphogenetic protein-2 enhances Wnt/β-catenin signaling-induced osteoprotegerin expression. Genes Cells 2009, 14, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Hilton, M.J.; Tu, X.; Wu, X.; Bai, S.; Zhao, H.; Kobayashi, T.; Kronenberg, H.M.; Teitelbaum, S.L.; Ross, F.P.; Kopan, R.; et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat. Med. 2008, 14, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Bae, Y.; Yang, T.; Zeng, H.C.; Campeau, P.M.; Chen, Y.; Bertin, T.; Dawson, B.C.; Munivez, E.; Tao, J.; Lee, B.H. miRNA-34c regulates Notch signaling during bone development. Hum. Mol. Genet. 2012, 21, 2991–3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, M.; Uyama, M.; Sugiyama, Y.; Sato, M. Canonical Wnt signaling activates miR-34 expression during osteoblastic differentiation. Mol. Med. Rep. 2013, 8, 1807–1811. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Guo, B.; Li, Q.; Peng, J.; Yang, Z.; Wang, A.; Li, D.; Hou, Z.; Lv, K.; Kan, G.; et al. MiR-214 targets ATF4 to inhibit bone formation. Nat. Med. 2013, 19, 93–100. [Google Scholar] [CrossRef]
- Yuan, Y.; Guo, J.; Zhang, L.; Tong, X.; Zhang, S.; Zhou, X.; Zhang, M.; Chen, X.; Lei, L.; Li, H.; et al. MiR-214 Attenuates the Osteogenic Effects of Mechanical Loading on Osteoblasts. Int. J. Sports Med. 2019, 40, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Kawao, N.; Iemura, S.; Kawaguchi, M.; Mizukami, Y.; Takafuji, Y.; Kaji, H. Role of irisin in effects of chronic exercise on muscle and bone in ovariectomized mice. J. Bone Miner. Metab. 2021, 39, 547–557. [Google Scholar] [CrossRef]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Oranger, A.; Mori, G.; Brunetti, G.; Colucci, S.; Cinti, S.; Grano, M. Irisin enhances osteoblast differentiation in vitro. Int. J. Endocrinol. 2014, 2014, 902186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Sun, K.; Zhao, S.; Geng, T.; Fan, X.; Sun, S.; Zheng, M.; Jin, Q. Irisin promotes osteogenic differentiation of bone marrow mesenchymal stem cells by activating autophagy via the Wnt//β-catenin signal pathway. Cytokine 2020, 136, 155292. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, W.; Huang, Y. MiRNA-133a is involved in the regulation of postmenopausal osteoporosis through promoting osteoclast differentiation. Acta Biochim. Biophys. Sin. (Shanghai) 2018, 50, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, X.X.; Takasaki, M.; Ohta, A.; Higuchi, M.; Ishimi, Y. Cooperative effects of exercise training and genistein administration on bone mass in ovariectomized mice. J. Bone Miner. Res. 2001, 16, 1829–1836. [Google Scholar] [CrossRef]
- Sun, X.; Li, F.; Ma, X.; Ma, J.; Zhao, B.; Zhang, Y.; Li, Y.; Lv, J.; Meng, X. The Effects of Combined Treatment with Naringin and Treadmill Exercise on Osteoporosis in Ovariectomized Rats. Sci. Rep. 2015, 5, 13009. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.A.; Cumming, R.G. Systematic review of randomized trials of the effect of exercise on bone mass in pre- and postmenopausal women. Calcif. Tissue Int. 2000, 67, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Roca-Rivada, A.; Castelao, C.; Senin, L.L.; Landrove, M.O.; Baltar, J.; Crujeiras, A.B.; Seoane, L.M.; Casanueva, F.F.; Pardo, M. FNDC5/Irisin Is Not Only a Myokine but Also an Adipokine. PLoS ONE 2013, 8, e60563. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.; Lu, Y.; Czernik, P.J.; Rosen, C.J.; Enerback, S.; Lecka-Czernik, B. Inducible brown adipose tissue, or beige fat, is anabolic for the skeleton. Endocrinology 2013, 154, 2687–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denham, J.; Gray, A.; Scott-Hamilton, J.; Hagstrom, A.D. Sprint Interval Training Decreases Circulating MicroRNAs Important for Muscle Development. Int. J. Sports Med. 2018, 39, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Jeon, J.; An, J.J.; Yi, H.K. Interval running training improves age-related skeletal muscle wasting and bone loss: Experiments with ovariectomized rats. Exp. Physiol. 2019, 104, 691–703. [Google Scholar] [CrossRef]
- Alipour, M.R.; Naderi, R.; Alihemmati, A.; Sheervalilou, R.; Ghiasi, R. Swimming training attenuates pancreatic apoptosis through miR-34a/Sirtu in1/P53 Axis in high-fat diet and Streptozotocin-induced Type-2 diabetic rats. J. Diabetes Metab. Disord. 2020, 19, 1439–1446. [Google Scholar] [CrossRef]
- Huang, C.C.; Wang, T.; Tung, Y.T.; Lin, W.T. Effect of exercise training on skeletal muscle SIRT1 and PGC-1α expression levels in rats of different age. Int. J. Med. Sci. 2016, 13, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; You, X.; Zhou, H.; Kang, A.; He, W.; Li, Z.; Li, B.; Zhu, H.; Zhao, Y.; Yu, G.; et al. p53 plays a central role in the development of osteoporosis. Aging (Albany NY) 2020, 12, 10473–10487. [Google Scholar] [CrossRef]
- Leboff, M.S.; Greenspan, S.L.; Insogna, K.L.; Lewiecki, E.M.; Saag, K.G.; Singer, A.J.; Siris, E.S. The clinician’ s guide to prevention and treatment of osteoporosis. Osteoporos. Int. 2022, 1–54. [Google Scholar] [CrossRef]
- Rossi, L.M.M.; Copes, R.M.; Dal Osto, L.C.; Flores, C.; Comim, F.V.; Premaor, M.O. Factors related with osteoporosis treatment in postmenopausal women. Medicine (U.S.) 2018, 97, e11524. [Google Scholar] [CrossRef] [PubMed]
- Marcu, F.M.; Lazăr, L.; Cioară, F.; Nemeth, S.; Bungău, S.; Bănică, F. Clinical study regarding the rehabilitation treatment of osteoporotic patients. Farmacia 2021, 69, 123–128. [Google Scholar] [CrossRef]
- Camacho, P.M.; Petak, S.M.; Binkley, N.; Diab, D.L.; Eldeiry, L.S.; Farooki, A.; Harris, S.T.; Hurley, D.L.; Kelly, J.; Michael Lewiecki, E.; et al. American association of clinical endocrinologists/American college of endocrinology clinical practice guidelines for the diagnosis and treatment of postmenopausal osteoporosis-2020 update. Endocr. Pract. 2020, 26, 1–46. [Google Scholar] [CrossRef]
- De Castro Gomes, D.A.; Valadares, A.L.R.; Pinto-Neto, A.M.; Morais, S.S.; Costa-Paiva, L. Ability to follow drug treatment with calcium and vitamin D in postmenopausal women with reduced bone mass. Menopause 2012, 19, 989–994. [Google Scholar] [CrossRef]
- Somekawa, Y.; Chiguchi, M.; Ishibashi, T.; Aso, T. Soy intake related to menopausal symptoms, serum lipids, and bone mineral density in postmenopausal Japanese women. Obstet. Gynecol. 2001, 97, 109–115. [Google Scholar] [CrossRef]
- Zhang, X.; Shu, X.O.; Li, H.; Yang, G.; Li, Q.; Gao, Y.T.; Zheng, W. Prospective cohort study of soy food consumption and risk of bone fracture among postmenopausal women. Arch. Intern. Med. 2005, 165, 1890–1895. [Google Scholar] [CrossRef] [Green Version]
- Ţiţ, D.M.; Pallag, A.; Iovan, C.; Furău, G.; Furău, C.; Bungău, S. Somatic-vegetative symptoms evolution in postmenopausal women treated with phytoestrogens and hormone replacement therapy. Iran. J. Public Health 2017, 46, 1528–1534. [Google Scholar] [PubMed]
- Tit, D.M.; Bungau, S.; Iovan, C.; Cseppento, D.C.N.; Endres, L.; Sava, C.; Sabau, A.M.; Furau, G.; Furau, C. Effects of the hormone replacement therapy and of soy isoflavones on bone resorption in postmenopause. J. Clin. Med. 2018, 7, 297. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, G.; Sanchis-Gomar, F.; Perego, S.; Sansoni, V.; Banfi, G. Implications of exercise-induced adipo-myokines in bone metabolism. Endocrine 2016, 54, 284–305. [Google Scholar] [CrossRef]
- Liu, J. Irisin as an exercise-stimulated hormone binding crosstalk between organs. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 316–321. [Google Scholar]
- Tveit, M.; Rosengren, B.E.; Nilsson, J.A.; Karlsson, M.K. Exercise in youth: High bone mass, large bone size, and low fracture risk in old age. Scand. J. Med. Sci. Sports 2015, 25, 453–461. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tian, Y.; Valenzuela, P.L.; Huang, C.; Zhao, J.; Hong, P.; He, Z.; Yin, S.; Lucia, A. Myokine Response to High-Intensity Interval vs. Resistance Exercise: An Individual Approach. Front. Physiol. 2018, 9, 1735. [Google Scholar] [CrossRef]
- Hong, A.R.; Kim, S.W. Effects of resistance exercise on bone health. Endocrinol. Metab. 2018, 33, 435–444. [Google Scholar] [CrossRef]
- Huh, J.Y.; Siopi, A.; Mougios, V.; Park, K.H.; Mantzoros, C.S. Irisin in response to exercise in humans with and without metabolic syndrome. J. Clin. Endocrinol. Metab. 2015, 100, E453–E457. [Google Scholar] [CrossRef]
- Rashti, B.A.; Mehrabani, J.; Damirchi, A.; Babaei, P. The influence of concurrent training intensity on serum irisin and abdominal fat in postmenopausal women. Prz. Menopauzalny 2019, 18, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Rioux, B.V.; Goulet, E.D.B.; Johanssen, N.M.; Swift, D.L.; Bouchard, D.R.; Loewen, H.; Sénéchal, M. Effect of an acute exercise bout on immediate post-exercise irisin concentration in adults: A meta-analysis. Scand. J. Med. Sci. Sports 2018, 28, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Aboarrage Junior, A.M.; Teixeira, C.V.L.S.; Dos Santos, R.N.; Machado, A.F.; Evangelista, A.L.; Rica, R.L.; Alonso, A.C.; Barroso, J.A.; Serra, A.J.; Baker, J.S.; et al. A high-intensity jump-based aquatic exercise program improves bone mineral density and functional fitness in postmenopausal women. Rejuvenation Res. 2018, 21, 535–540. [Google Scholar] [CrossRef]
- Vainionpää, A.; Korpelainen, R.; Leppäluoto, J.; Jämsä, T. Effects of high-impact exercise on bone mineral density: A randomized controlled trial in premenopausal women. Osteoporos. Int. 2005, 16, 191–197. [Google Scholar] [CrossRef]
- Yu, P.; Hsu, W.; Hsu, W.; Kuo, L.; Lin, Z.; Shen, W.; Hsu, R.W. The effects of high impact exercise intervention on bone mineral density, physical fi tness, and quality of life in postmenopausal women with osteopenia. Medicine 2019, 98, e14898. [Google Scholar] [CrossRef]
- Singhal, V.; Lawson, E.A.; Ackerman, K.E.; Fazeli, P.K.; Clarke, H.; Lee, H.; Eddy, K.; Marengi, D.A.; Derrico, N.P.; Bouxsein, M.L.; et al. Irisin levels are lower in young amenorrheic athletes compared with eumenorrheic athletes and non-athletes and are associated with bone density and strength estimates. PLoS ONE 2014, 9, e100218. [Google Scholar] [CrossRef]
- Gaudio, A.; Rapisarda, R.; Xourafa, A.; Zanoli, L.; Manfrè, V.; Catalano, A.; Signorelli, S.S.; Castellino, P. Effects of competitive physical activity on serum irisin levels and bone turnover markers. J. Endocrinol. Investig. 2021, 44, 2235–2241. [Google Scholar] [CrossRef] [PubMed]
- Kemmler, W.; von Stengel, S.; Kohl, M. Exercise frequency and bone mineral density development in exercising postmenopausal osteopenic women. Is there a critical dose of exercise for affecting bone? Results of the Erlangen Fitness and Osteoporosis Prevention Study. Bone 2016, 89, 1–6. [Google Scholar] [CrossRef]
- Shanazari, Z.; Hemati Farsani, Z.; Faramarzi, M.; Banitalebi, E. MyomiR-OsteomiR crosstalk induced by different modes and intensities of exercise training and its role in controlling osteogenic differentiation in old male Wistar rats. Exp. Gerontol. 2021, 149, 111305. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Z.; Fu, Q.; Zhang, J. Plasma miRNA levels correlate with sensitivity to bone mineral density in postmenopausal osteoporosis patients. Biomarkers 2014, 19, 553–556. [Google Scholar] [CrossRef]
- Sansoni, V.; Perego, S.; Vernillo, G.; Barbuti, A.; Merati, G.; La Torre, A.; Banfi, G.; Lombardi, G. Effects of repeated sprints training on fracture risk-associated miRNA. Oncotarget 2018, 9, 18029–18040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemmler, W.; Shojaa, M.; Kohl, M.; von Stengel, S. Effects of Different Types of Exercise on Bone Mineral Density in Postmenopausal Women: A Systematic Review and Meta-Analysis; Springer: New York, NY, USA, 2020; Volume 107, ISBN 0123456789. [Google Scholar]

| Stratification of Risk | Low Risk (No Previous Spine or Hip Fracture; T-Score at Hip and Spine above −1.0; Normal Bmd) | Moderate Risk (No Previous Spine or Hip Fracture; T-Score between −1.0 and −2.5; Low Bone Mass) | High Risk (Prior Spine or Hip Fracture; or a Lumbar Spine or Hip T-Score of −2.5 or Below; Osteoporosis) | Very High Risk (Multiple Spine Fractures/Hip Fracture and T-Score of −2.5 or Lower at Lumbar Spine or Hip; Severe or Established Osteoporosis) |
|---|---|---|---|---|
| Non Pharmacological Treatments | Optimize Calcium and Vitamin D Status | |||
| Risk Appropriate Exercise | Risk Appropriate Exercise and Falls Preventions | |||
| Lifestyle recommendation | Weight-bearing activities; Muscle-strengthening activities; Balance, posture, and functional activities. | |||
| Pharmacological Treatments | Not recommended | Consider FDA-approved medical therapies in adults with low bone mass (osteopenia) and with a 10-year probability of a hip fracture ≥ 3% or 10-year probability of any major osteoporosis-related fracture ≥ 20%. | Initial treatment with inhibitors of bone resorption
| Initial treatment with stimulators of bone formation. Teriparatide or Abaloparatide treatment for up to 2 years or Romosozumab for 1 year. Following a course of anabolic treatment with antiresorptive osteoporosis therapies should be used to maintain bone density gains. |
| Follow-up | Regularly assess compliance and persistence with the therapeutic regimen (at least annually) | |||
| Reassess Fracture Risk in 2 to 4 Years | Reassess Fracture Risk in 2 to 4 Years | |||
| Reference Year Author | Article Type | Population | Duration and Type of Exercise | Effects |
|---|---|---|---|---|
| [32] 2017 Singulani, M.P et al. | Article | Adult aged vs. exercised aged rats | -ST -16 weeks | ST: -Increase in: biomechanical parameters, runx2, osx gene, and bone matrix protein expression -Reduction in: pparγ expression and risk of fractures during senescent |
| [42] 2019 Gonzalo-Encabo, P. et al. | Clinical trial study | 400 idle postmenopausal woman | High rates vs. moderate rates of aerobic exercise -12 month | HIGH RATES OF AEROBIC EXERCISE: -Improves BMD -Attenuates age-related declines Benefits persist at intervention concludes |
| [43] 2017 Zhao, R. et al. | Review | 1061 postmenopausal women from 11 RCTs | -Combined exercise -Data at 12 month | Increase in BMD Prevents postmenopausal bone loss and helps in reducing risk of fracture |
| [44] 2015 Ju, Y. I. et al. [45] 2016 Oh, T. et al. | Articles | OVX rats | -Swimming -12 weeks -High-strength intermittent swimming training -6 weeks | -Trabecular architectural changes in cancellous bone -Beneficial effect on bone quantity and intensity |
| [46] 2016 Pereira, A. et al. | Article | 11 postmenopausal women | -Combined training -16 week | Improves dynamic muscular strength in lower and upper limbs No change in bone resorption |
| [47] 2013 Mosti, M.P. et al. | Article | 21 postmenopausal women with OP or osteopenia | -Squat exercise -12 weeks | MAXIMAL STRENGTH TRAINING: Improves BMD and bone mineral content |
| [48] 2008 Gunendi, Z. et al. | Article | 25 postmenopausal women without OP vs. 28 postmenopausal women with OP | -Submaximal aerobic exercise program on treadmill -Twice a week for 4 weeks | Improves static and dynamic postural balance in postmenopausal women with OP No changes in BMD |
| [49] 2020 Mittlmeier, T. | Article | OVX rats | Low-intensity endurance training on the treadmill vs. medium and HIIT -5 week | TREADMILL TRAINING: Influences differentially on the musculoskeletal unit |
| [57] 2021 Kawao, N. | Article | OVX mice | -Treadmill exercise with moderate intensity -8 week | Irisin high level might be related to increases in trabecular BMD but not with cortical BMD improvement in decreases in trabecular and cortical BMD |
| [67] 2019 Kim Jeong-Seok | Article | OVX rats | -Interval running -6 week | Decreases bone resorption prevents skeletal muscle wasting and bone loss in the elderly |
| [88] 2018 Aboarrage Junior, A.M. et al. | Article | 25 postmenopausal women | -High-intensity jump-based aquatic exercise program (HIIAE) | Improves BMD |
| [89] 2005 Vainionpää, A. et al. | Article | 120 premenopausal women | -Progressive high-impact exercises -3 times per week for 12 months | Improves BMD prevents OP |
| [90] 2019 Yu, P. et al. | Article | 100 postmenopausal women | -Aerobic dancing -24 weeks | Improves: BMD, muscle strength, agility. High-impact exercise better influence the trabecular bone than cancellous bone |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vita, F.; Gangemi, S.; Pioggia, G.; Trimarchi, F.; Di Mauro, D. Physical Activity and Post-Transcriptional Regulation of Aging Decay: Modulation of Pathways in Postmenopausal Osteoporosis. Medicina 2022, 58, 767. https://doi.org/10.3390/medicina58060767
Vita F, Gangemi S, Pioggia G, Trimarchi F, Di Mauro D. Physical Activity and Post-Transcriptional Regulation of Aging Decay: Modulation of Pathways in Postmenopausal Osteoporosis. Medicina. 2022; 58(6):767. https://doi.org/10.3390/medicina58060767
Chicago/Turabian StyleVita, Federica, Sebastiano Gangemi, Giovanni Pioggia, Fabio Trimarchi, and Debora Di Mauro. 2022. "Physical Activity and Post-Transcriptional Regulation of Aging Decay: Modulation of Pathways in Postmenopausal Osteoporosis" Medicina 58, no. 6: 767. https://doi.org/10.3390/medicina58060767
APA StyleVita, F., Gangemi, S., Pioggia, G., Trimarchi, F., & Di Mauro, D. (2022). Physical Activity and Post-Transcriptional Regulation of Aging Decay: Modulation of Pathways in Postmenopausal Osteoporosis. Medicina, 58(6), 767. https://doi.org/10.3390/medicina58060767