Biomarker Potential of Plasma MicroRNA-150-5p in Prostate Cancer

 ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
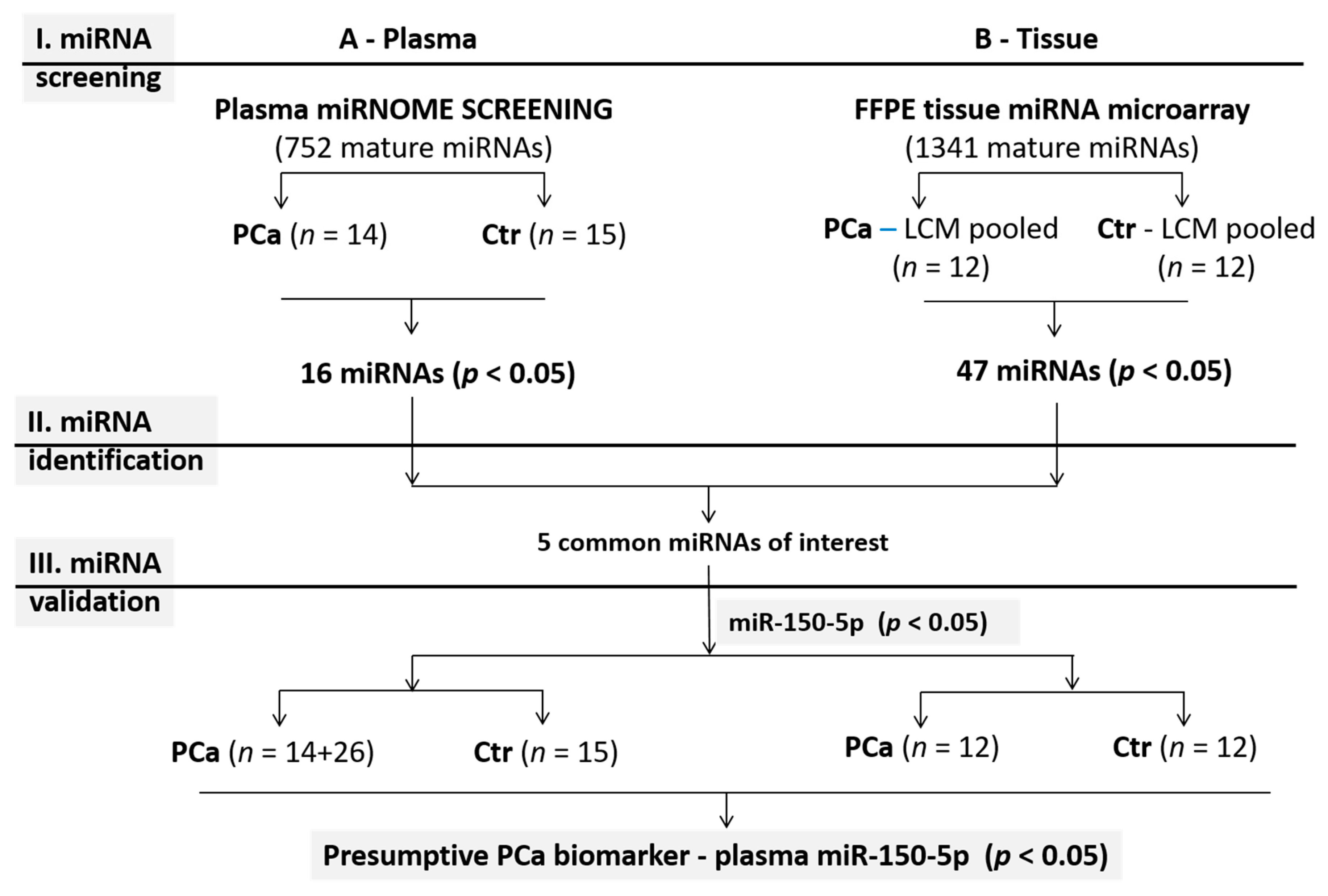
2.1. Study Design
2.2. Plasma Samples
2.3. Plasma miRNome Screening
2.4. Plasma miRNA Validation
2.5. MiRNA Analysis in FFPE Tissues
2.6. Statistical Analysis
2.7. Bioinformatics Analysis
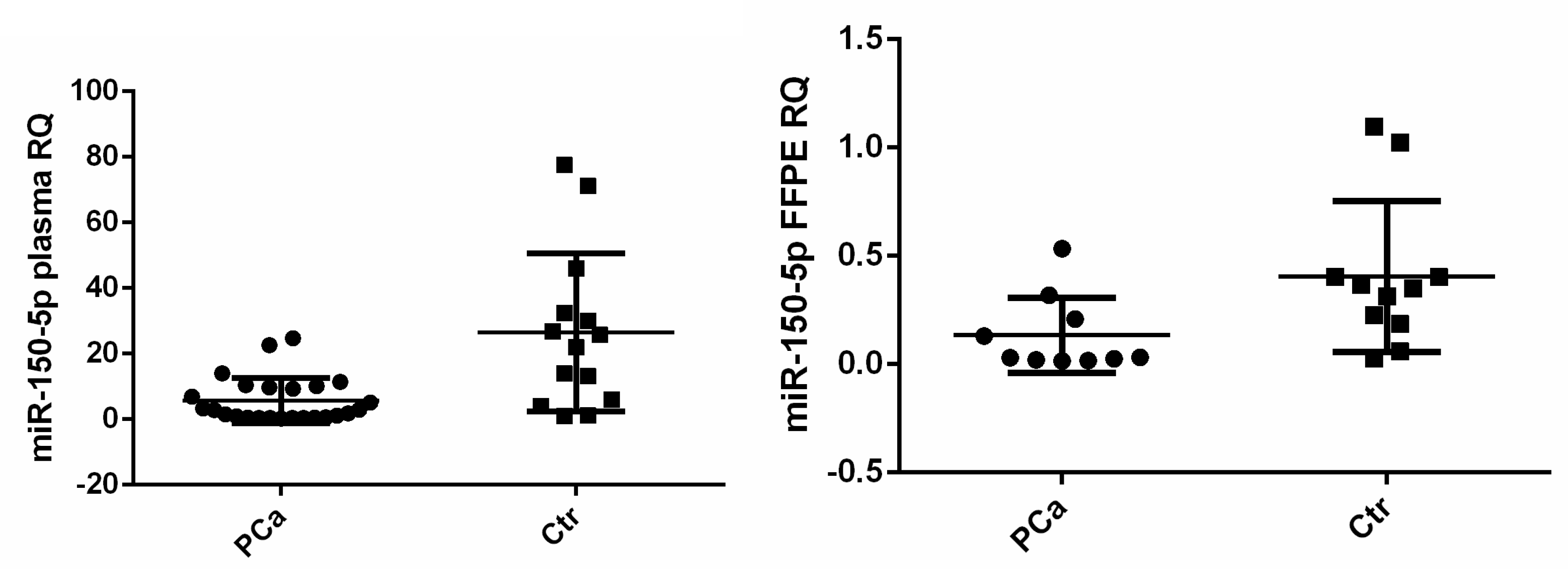
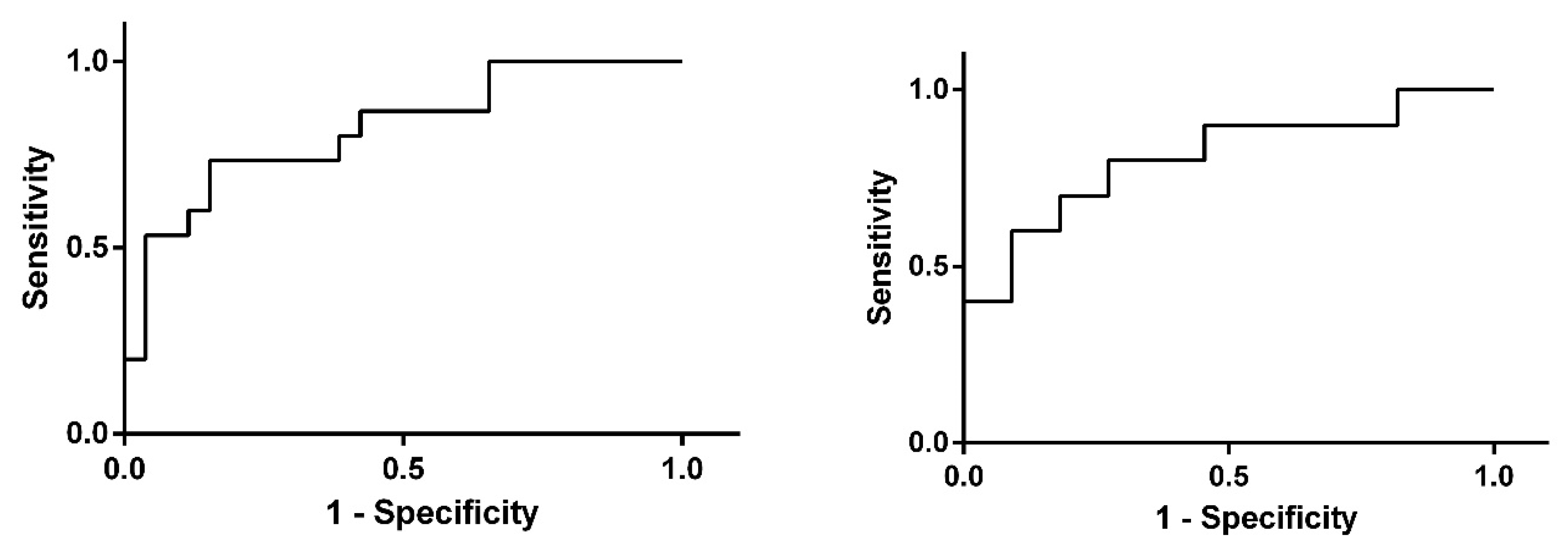
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- American Cancer Society. Cancer Facts & Figures 2018. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2018.html (accessed on 12 April 2018).
- Pernar, C.H.; Ebot, E.M. The Epidemiology of Prostate Cancer. Cold Spring Harb. Perspect Med. 2018. Available online: http://perspectivesinmedicine.cshlp.org/content/8/12/a030361.abstract (accessed on 23 May 2019). [CrossRef] [PubMed]
- Vanacore, D.; Boccellino, M. Micrornas in Prostate Cancer: An Overview. Oncotarget 2017, 8, 50240–50251. [Google Scholar] [CrossRef] [PubMed]
- Fabris, L.; Ceder, Y. The Potential of MicroRNAs as Prostate Cancer Biomarkers. Eur. Urol. 2016, 70, 312–322. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The Role of MicroRNAs in Human Cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Wang, K. The Ubiquitous Existence of MicroRNA in Body Fluids. Clin. Chem. 2017, 63, 784–785. [Google Scholar] [CrossRef]
- Balacescu, O.; Petrut, B. Urinary MicroRNAs for Prostate Cancer Diagnosis, Prognosis, and Treatment Response: Are We There Yet? Wiley Interdiscip. Rev. RNA 2017, 8, e1438. [Google Scholar] [CrossRef]
- Mihala, A.; Alexa, A.A. A Pilot Study on the Expression of MicroRNAs Resident on Chromosome 21 in Laser Microdissected FFPE Prostate Adenocarcinoma Samples. Rom. J. Morphol. Embryol. 2015, 56, 1063–1068. [Google Scholar]
- Seclaman, E.; Narita, D. MicroRNA Expression in Laser Micro-Dissected Breast Cancer Tissue Samples—A Pilot Study. Pathol. Oncol. Res. 2019, 25, 233–239. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Ding, J.; Li, X. TarPmiR: A New Approach for MicroRNA Target Site Prediction. Bioinformatics 2016, 32, 2768–2775. [Google Scholar] [CrossRef] [PubMed]
- Sticht, C.; De La Torre, C. Mirwalk: An Online Resource for Prediction of Microrna Binding Sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.S.; Schultz, N. Integrative Genomic Profiling of Human Prostate Cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44. [Google Scholar] [CrossRef] [PubMed]
- Dybos, S.A.; Flatberg, A. Increased Levels of Serum MiR-148a-3p Are Associated with Prostate Cancer. APMIS 2018, 126, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Ramalho-Carvalho, J.; Martins, J.B. Epigenetic Disruption of MiR-130a Promotes Prostate Cancer by Targeting SEC23B and DEPDC1. Cancer Lett. 2017, 385, 150–159. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, Y.; Song, J. MiR-150 Promotes the Cell Invasion of Prostate Cancer Cells by Directly Regulating the Expression of P53. Die Pharm. 2016, 71, 533–536. [Google Scholar]
- Liu, D.Z.; Zhang, H.Y. MIR-150 Promotes Prostate Cancer Stem Cell Development via Suppressing P27Kip1. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4344–4352. [Google Scholar]
- Hong, X.; Yu, J.J. MicroRNA-150 Suppresses Epithelial Mesenchymal Transition, Invasion and Metastasis in Prostate Cancer through the TRPM4-Mediated β-Catenin Signalling Pathway. Am. J. Physiol. Cell Physiol. 2018, 316, 463–480. [Google Scholar] [CrossRef] [PubMed]
- Okato, A.; Arai, T. Dual Strands of Pre-MiR-150 (MiR-150-5p and MiR-150-3p) Act as Antitumor MiRNAs Targeting SPOCK1 in Naïve and Castration-Resistant Prostate Cancer. Int. J. Oncol. 2017, 51, 245–256. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Y. Aryl Hydrocarbon Receptor Enhances the Expression of MiR-150-5p to Suppress in Prostate Cancer Progression by Regulating MAP3K12. Arch. Biochem. Biophys. 2018, 654, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.R.; Khandrika, L. Focal Adhesion Kinase Controls Aggressive Phenotype of Androgen-Independent Prostate Cancer. Mol. Cancer Res. 2008, 6, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.-L.; Chen, L.-C. Emerging Roles of Focal Adhesion Kinase in Cancer. Biomed. Res. Int. 2015, 2015, 690690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, Y. MicroRNA Expression Profile in Chronic Nonbacterial Prostatitis Revealed by Next-Generation Small RNA Sequencing. Asian J. Androl. 2018, 21, 351. [Google Scholar]
- Valle, I.F.; Menichetti, G. Network integration of multi-tumour omics data suggests novel targeting strategies. Nat. Commun. 2018, 9, 4514. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, G.; Loizzo, D. Metabolomic profiling for the identification of novel diagnostic markers and therapeutic targets in prostate cancer: An update. Expert Rev. Mol. Diagn. 2019, 119, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Heinz, R.E.; Rudolph, M.C.; Ramanathan, P. Constitutive expression of microRNA-150 in mammary epithelium suppresses secretory activation and impairs de novo lipogenesis. Development 2016, 143, 4236–4248. [Google Scholar] [CrossRef]
- Kang, M.; Liu, X.; Fu, Y.; Garvey, W.T. Improved systemic metabolism and adipocyte biology in miR-150 knockout mice. Metabolism 2018, 83, 139–148. [Google Scholar] [CrossRef]
- Li, H.; Liu, J.; Cao, W. C-myc/miR-150/EPG5 axis mediated dysfunction of autophagy promotes development of non-small cell lung cancer. Theranostics 2019, 9, 5134–5148. [Google Scholar] [CrossRef]
- Cui, Y.J.; Liu, C.; Pang, F.W. MiR-150 inhibits hypoxia-induced autophagy in oral squamous carcinoma by negatively regulating BNIP3. Int. J. Clin. Exp. Pathol. 2017, 10, 2917–2927. [Google Scholar]
- Kim, T.H.; Jeong, J.Y.; Park, J.Y. miR-150 enhances apoptotic and anti-tumor effects of paclitaxel in paclitaxel-resistant ovarian cancer cells by targeting Notch3. Oncotarget 2017, 8, 72788–72800. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, X.; Long, X. miR-150 is a factor of survival in prostate cancer patients. JBUON 2015, 20, 173–179. [Google Scholar] [PubMed]
- Adams, B.D.; Guo, S.; Bai, H. An In Vivo Functional Screen Uncovers miR-150-Mediated Regulation of Hematopoietic Injury Response. Cell Rep. 2012, 2, 1048–1060. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Cases (n = 14) | Controls (n = 15) | Cases for Validation (n = 26) |
|---|---|---|---|
| Age (years ± SD) | 64.9 (±5.45) | 51.3 (±8.27) | 69.3 (±6.34) |
| PSA n (%) | |||
| <4 ng/mL | 0 (0.00) | 15 (100.00) | 0 (0.00) |
| 4–10 ng/mL | 6 (40.00) | 0 (0.00) | 13 (50.00) |
| ≥10 ng/mL | 9 (60.00) | 0 (0.00) | 12 (46.15) |
| Gleason Score n (%) | |||
| 5–6 | 2 (14.28) | 5 (19.23) | |
| 7 | 10 (71.43) | 14 (53.84) | |
| 8–10 | 2 (14.28) | 7 (26.92) |
| miRs Down | Fold Change | p-Value | miRs Up | Fold Change | p-Value |
|---|---|---|---|---|---|
| miR-130a-3p | −1.4640 | 0.0006 | miR-195-5p | 4.4277 | 0.0011 |
| miR-342-3p | −1.4793 | 0.0045 | miR-148a-3p | 1.6332 | 0.0084 |
| miR-20b-5p | −2.6733 | 0.0048 | miR-145-5p | 1.6182 | 0.0092 |
| miR-150-5p | −1.6961 | 0.0091 | let-7c | 1.5653 | 0.0213 |
| miR-324-3p | −1.5342 | 0.0108 | miR-365a-3p | 1.6709 | 0.0350 |
| miR-376a-3p | −2.6422 | 0.0160 | miR-199b-5p | 2.1282 | 0.0463 |
| miR-93-5p | −1.3391 | 0.0273 | |||
| miR-7-5p | −1.7245 | 0.0392 | |||
| miR-29b-3p | −1.3472 | 0.0418 | |||
| miR-215 | −1.4886 | 0.0433 |
| miR | FFPE Microarray | Plasma Screening | ||
|---|---|---|---|---|
| FC | p-Value | FC | p-Value | |
| miR-148a-3p | 1.647 | 0.012 | 1.633 | 0.008 |
| miR-130a-3p | −1.618 | <0.001 | −1.468 | 0.0005 |
| miR-150-5p | −1.409 | 0.002 | −1.696 | 0.009 |
| miR-145-5p | −1.748 | 0.020 | 1.618 | 0.009 |
| miR-365a-3p | −1.412 | 0.004 | 1.670 | 0.035 |
| Term (KEGG Pathway) | p-Value | Fold Enrichment | FDR | |
|---|---|---|---|---|
| miR-130a-3p, miR-150-5p, miR-145-5p, miR-365a-5p | hsa04510: Focal adhesion | 1.68 × 10−12 | 2.133 | 2.24 × 10−9 |
| hsa05200: Pathways in cancer | 6.35 × 10−9 | 1.636 | 8.44 × 10−6 | |
| hsa04512: ECM–receptor interaction | 5.77 × 10−8 | 2.402 | 7.67 × 10−5 | |
| hsa04722: Neurotrophin signaling pathway | 5.58 × 10−6 | 1.964 | 7.42 × 10−3 | |
| hsa04070: Phosphatidylinositol signaling | 6.28 × 10−6 | 2.077 | 8.35 × 10−3 | |
| hsa04151: PI3K-Akt signaling pathway | 2.18 × 10−5 | 1.491 | 2.89 × 10−2 | |
| hsa05222: Small-cell lung cancer | 2.81 × 10−5 | 2.079 | 3.73 × 10−2 | |
| miR-130a-3p, miR-150-5p | hsa05200: Pathways in cancer | 1.30 × 10−6 | 1.795 | 1.72 × 10−3 |
| hsa04510: Focal adhesion | 2.43 × 10−6 | 2.134 | 3.21 × 10−3 | |
| hsa04070: Phosphatidylinositol signaling | 1.60 × 10−5 | 2.608 | 2.12 × 10−2 | |
| miR-150-5p | hsa04510: Focal adhesion | 2.27 × 10−9 | 2.322 | 3.00 × 10−6 |
| hsa04512: ECM–receptor interaction | 2.58 × 10−6 | 2.699 | 3.42 × 10−3 | |
| miR-148a-3p | hsa04512: ECM–receptor interaction | 5.08 × 10−3 | 1.379 | 6.40 × 10−1 |
| hsa04510: Focal adhesion | 2.26 × 10−2 | 2.069 | 2.57 × 101 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paunescu, I.A.; Bardan, R.; Marcu, A.; Nitusca, D.; Dema, A.; Negru, S.; Balacescu, O.; Balacescu, L.; Cumpanas, A.; Sirbu, I.O.; et al. Biomarker Potential of Plasma MicroRNA-150-5p in Prostate Cancer. Medicina 2019, 55, 564. https://doi.org/10.3390/medicina55090564
Paunescu IA, Bardan R, Marcu A, Nitusca D, Dema A, Negru S, Balacescu O, Balacescu L, Cumpanas A, Sirbu IO, et al. Biomarker Potential of Plasma MicroRNA-150-5p in Prostate Cancer. Medicina. 2019; 55(9):564. https://doi.org/10.3390/medicina55090564
Chicago/Turabian StylePaunescu, Ionut Andrei, Razvan Bardan, Anca Marcu, Diana Nitusca, Alis Dema, Serban Negru, Ovidiu Balacescu, Loredana Balacescu, Alin Cumpanas, Ioan Ovidiu Sirbu, and et al. 2019. "Biomarker Potential of Plasma MicroRNA-150-5p in Prostate Cancer" Medicina 55, no. 9: 564. https://doi.org/10.3390/medicina55090564
APA StylePaunescu, I. A., Bardan, R., Marcu, A., Nitusca, D., Dema, A., Negru, S., Balacescu, O., Balacescu, L., Cumpanas, A., Sirbu, I. O., Petrut, B., Seclaman, E., & Marian, C. (2019). Biomarker Potential of Plasma MicroRNA-150-5p in Prostate Cancer. Medicina, 55(9), 564. https://doi.org/10.3390/medicina55090564