Effects of Quercetin-Loaded Nanoparticles on MCF-7 Human Breast Cancer Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of QT-SLNs
2.2. Particle Size and Zeta Potential of QT-SLNs
2.3. Drug Entrapment Efficiency
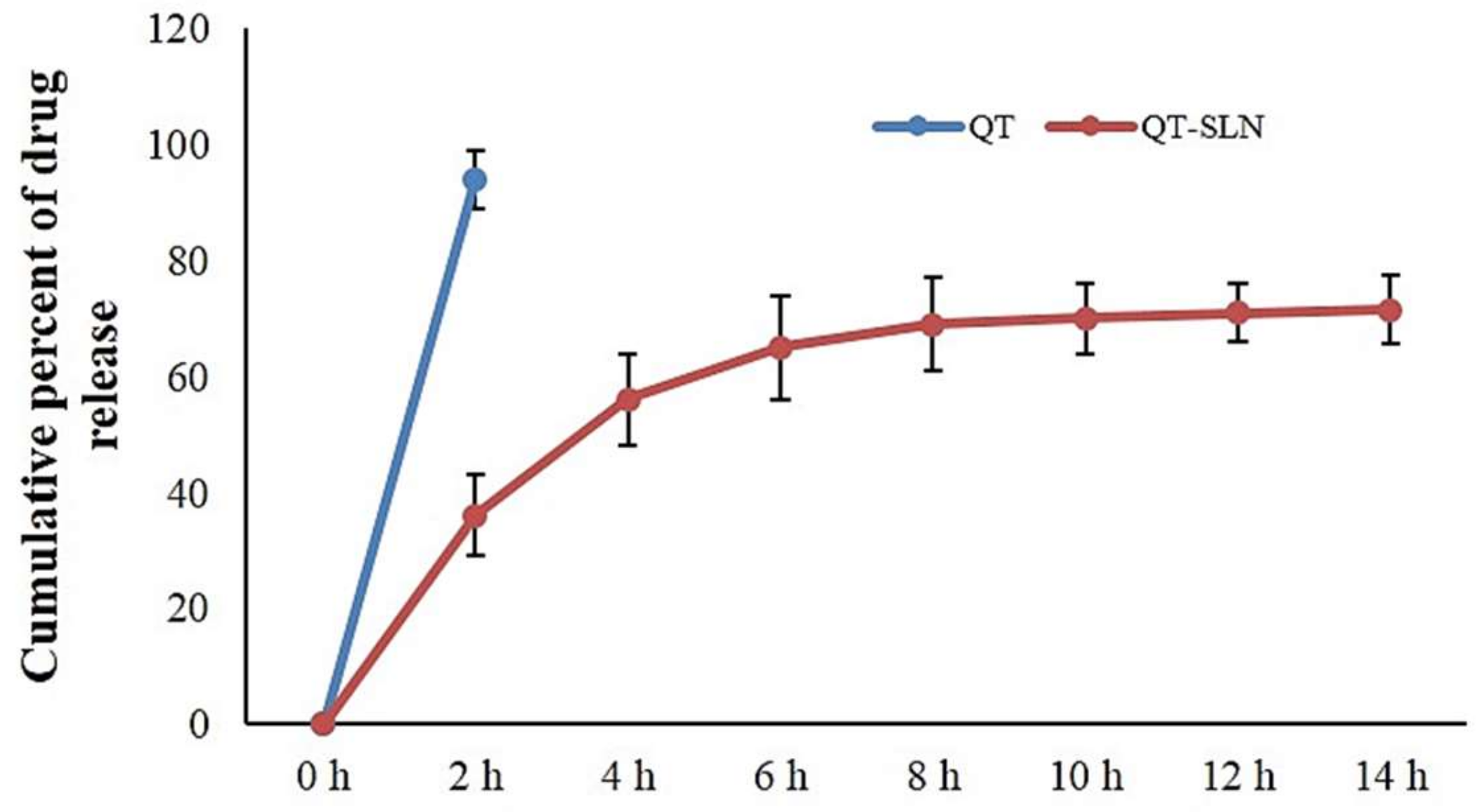
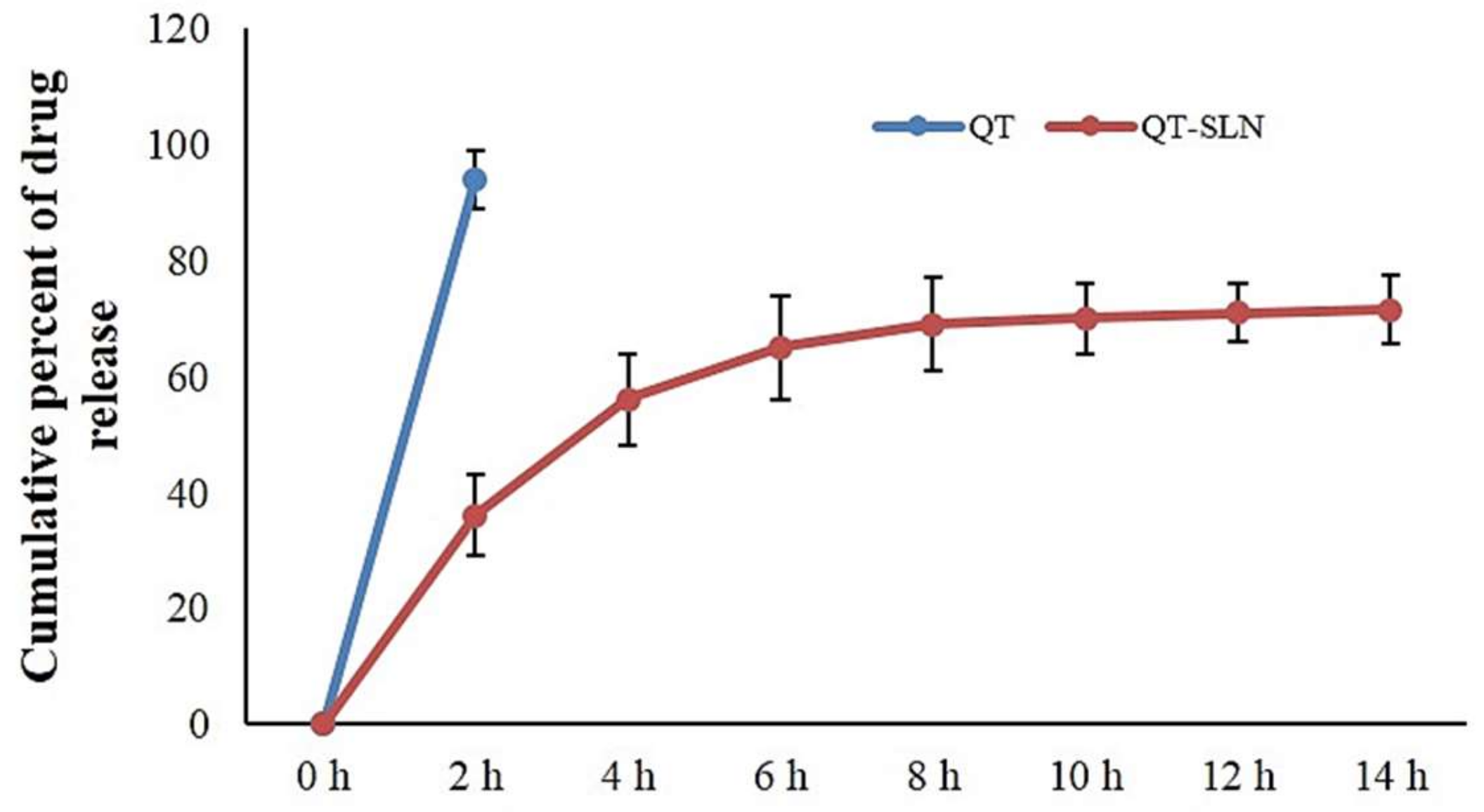
2.4. In Vitro Drug Release
2.5. Experimental Design
- Control: received only media
- Blank SLN: exposed to 25 µmol/mL of SLN without QT
- QT: treated by 25 µmol/mL of QT
- QT-SLN: treated with 25 µmol/mL of QT-SLNs
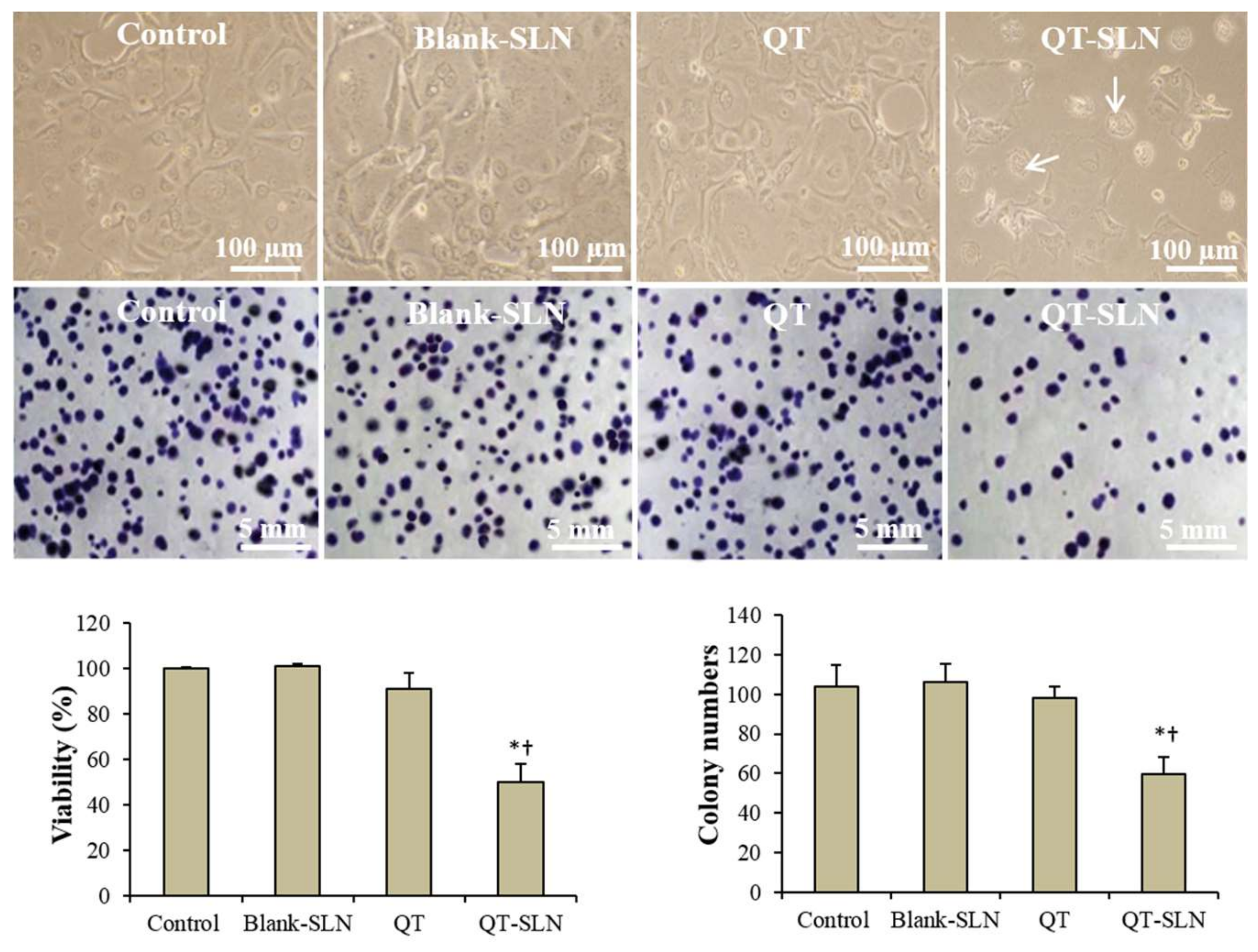
2.6. Cell Viability
2.7. Clonogenicity Assay
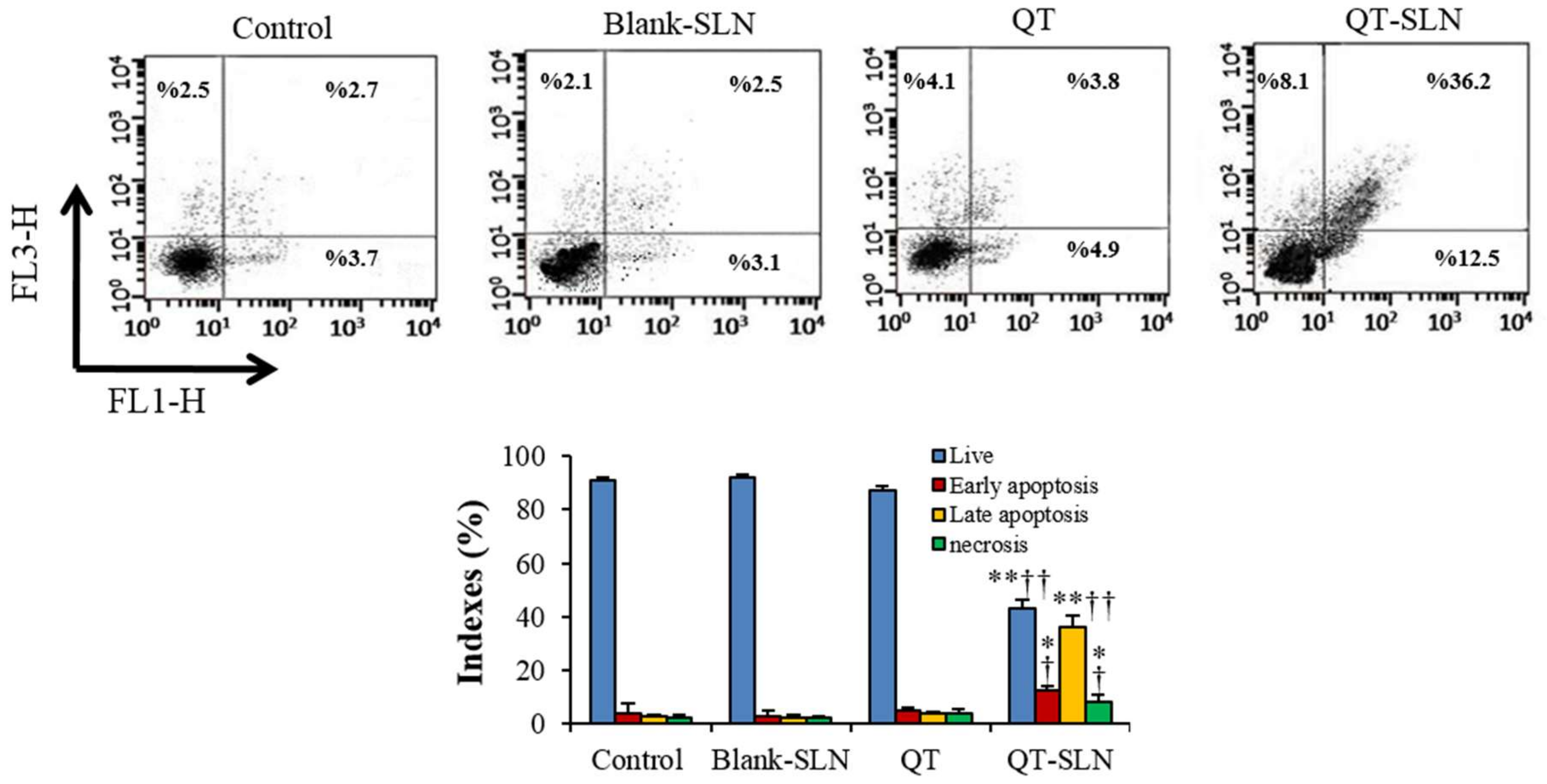
2.8. Annexin V-FITC/Propidium Iodide Apoptosis Assay
2.9. Real-Time Polymerase Chain Reaction
2.10. Western Analysis
2.11. Determination MDA Contents and Antioxidant Enzyme Activities
2.12. Determination of Intracellular ROS Levels
2.13. Statistical Analysis
3. Results
3.1. Characterization of QT-SLNs
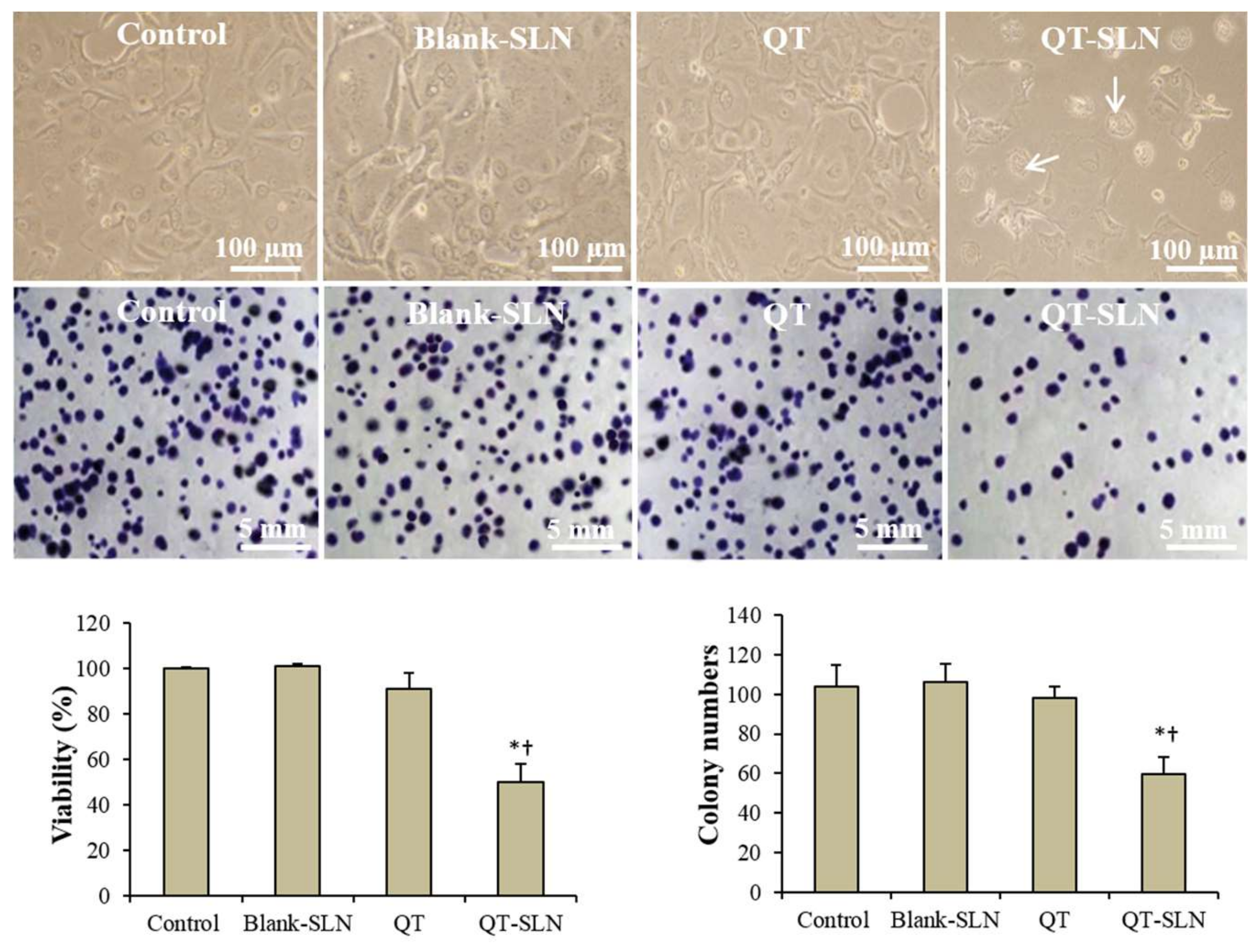
3.2. Cell Viability and Proliferation
3.3. Morphology Evaluation
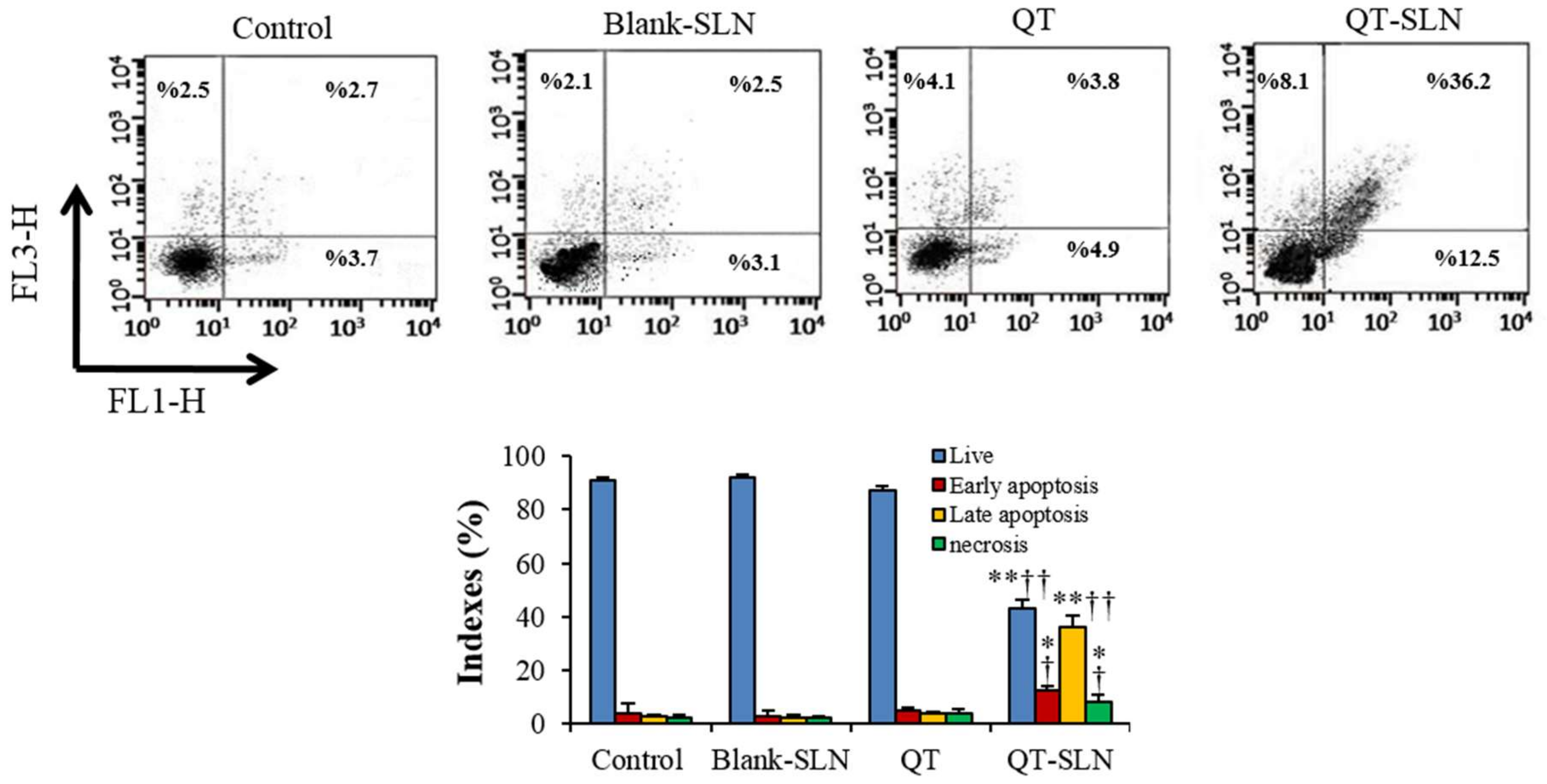
3.4. Annexin V-FITC/Propidium Iodide Apoptosis Assay
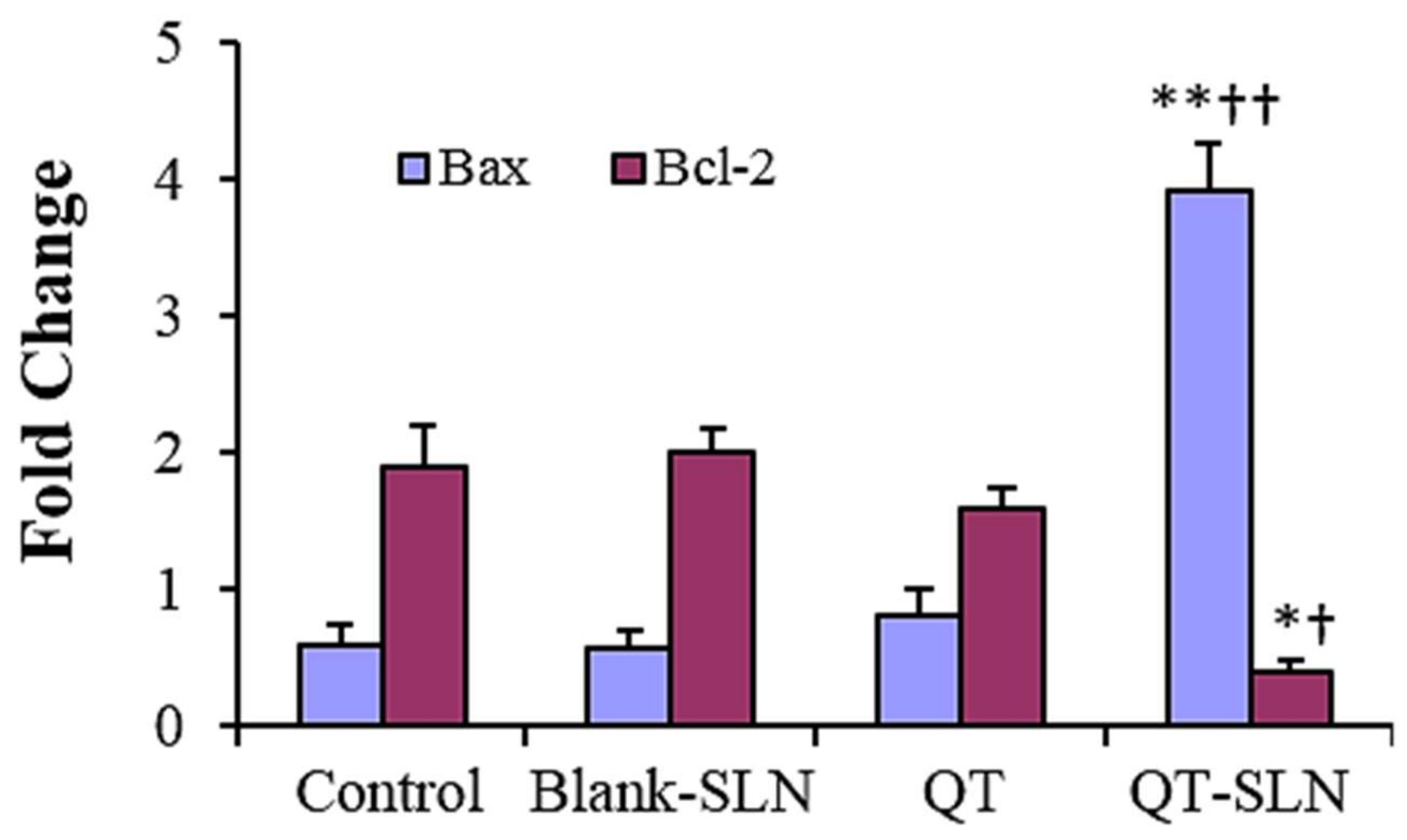
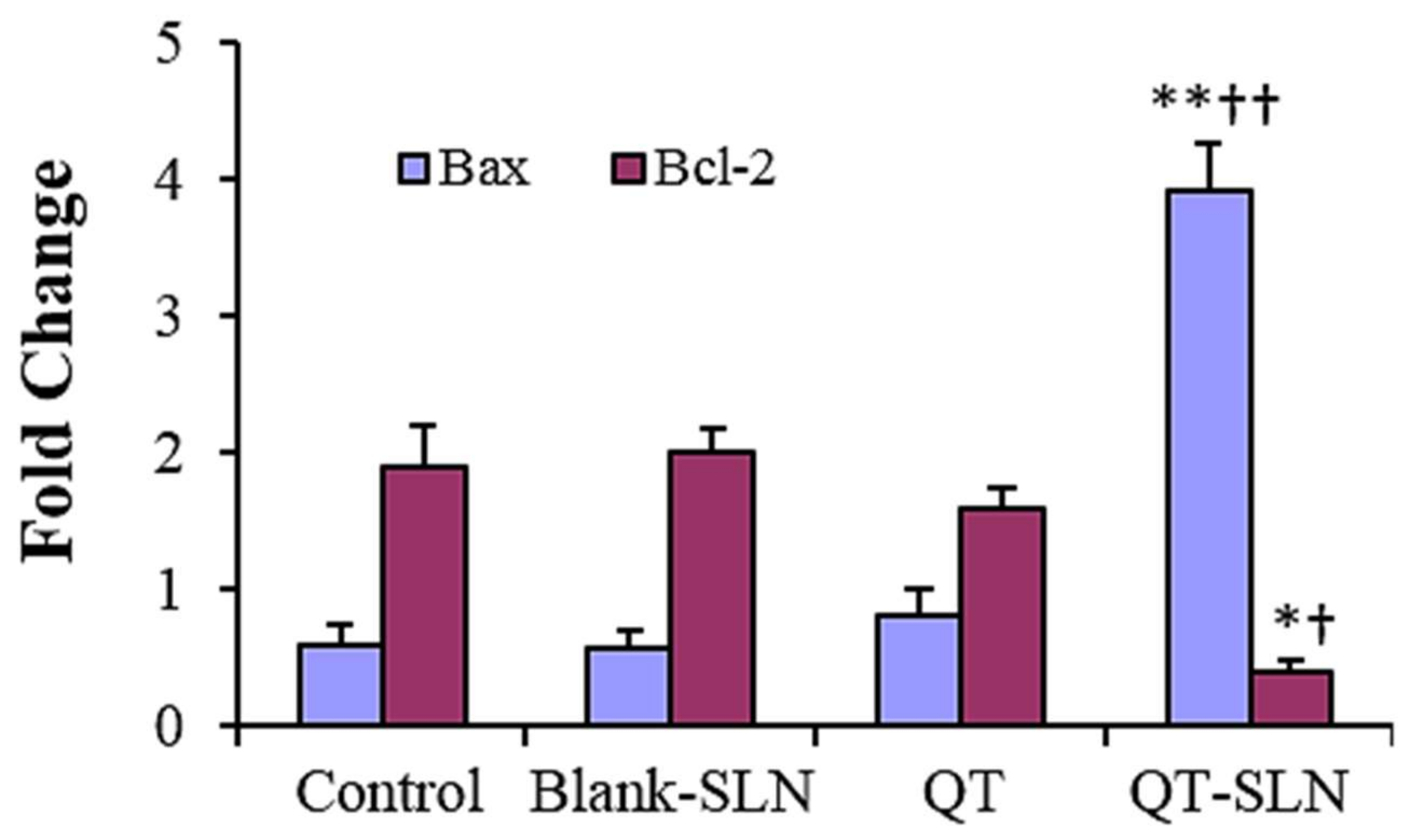
3.5. Quantitative Real-Time RT-PCR
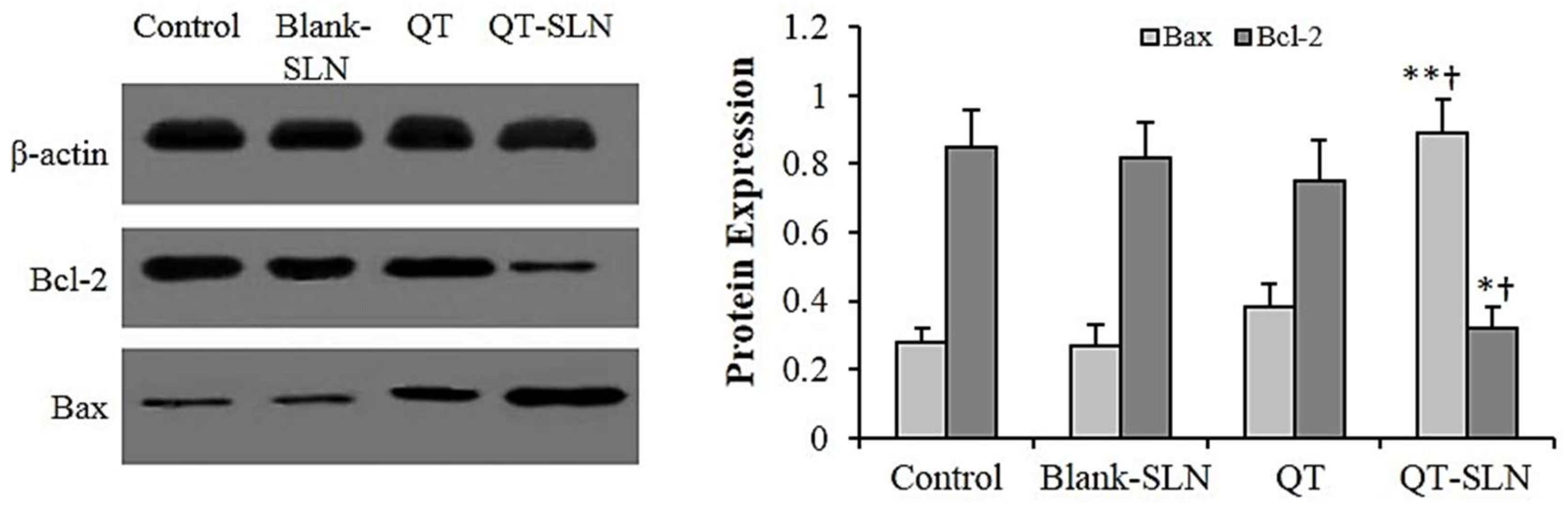
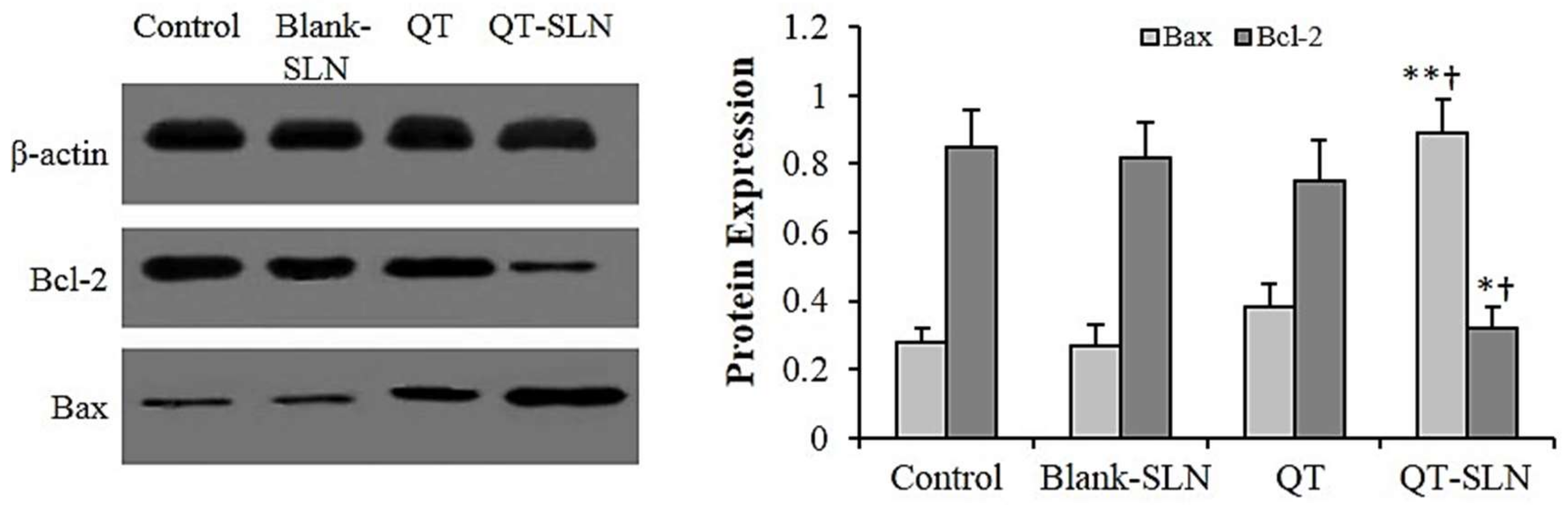
3.6. Western Analysis
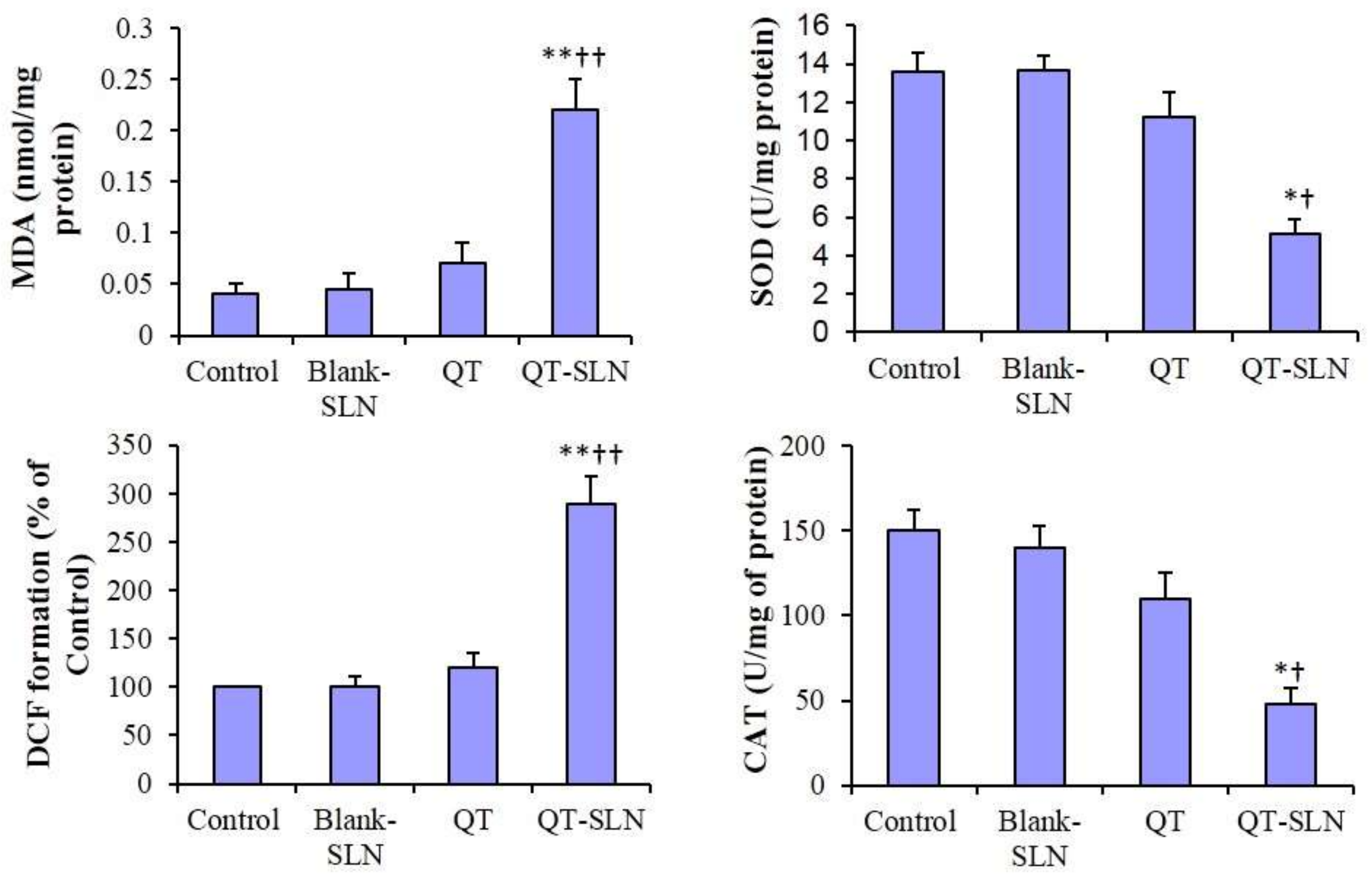
3.7. ROS Levels, MDA Content and Antioxidant Enzyme Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tinoco, G.; Warsch, S.; Glück, S.; Avancha, K.; Montero, A.J. Treating breast cancer in the 21st century: Emerging biological therapies. J. Cancer 2013, 4, 117–132. [Google Scholar] [CrossRef]
- Gadhwal, M.K.; Patil, S.; D’Mello, P.; Joshi, U.; Sinha, R.; Govil, G. Characterisation and antitumour activity of some Quercetin analogues. Ind. J. Pharm. Sci. 2013, 75, 233–237. [Google Scholar]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and cancer chemoprevention. Evid. Based Complement. Alternat. Med. 2011, 2011, 591356. [Google Scholar] [CrossRef] [PubMed]
- Dandawate, P.R.; Subramaniam, D.; Jensen, R.A.; Anant, S. Targeting cancer stem cells and signaling pathways by phytochemicals: Novel approach for breast cancer therapy. Semin. Cancer Biol. 2016, 40, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Sinha, D.; Biswas, J.; Nabavi, SM.; Bishayee, A. Tea phytochemicals for breast cancer prevention and intervention: From bench to bedside and beyond. Semin. Cancer Biol. 2017, 46, 33–54. [Google Scholar] [CrossRef]
- Lv, W.; Budke, B.; Pawlowski, M.; Connell, P.P.; Kozikowski, A.P. Development of small molecules that specifically inhibit the D-loop activity of RAD51. J. Med. Chem. 2016, 59, 4511–4525. [Google Scholar] [CrossRef] [PubMed]
- Aiello, F.; Carullo, G.; Giordano, F.; Spina, E.; Nigro, A.; Garofalo, A.; Tassini, S.; Costantino, G.; Vincetti, P.; Bruno, A.; et al. Identification of Breast Cancer Inhibitors Specific for G Protein-Coupled Estrogen Receptor (GPER)-Expressing Cells. Chem. Med. Chem. 2017, 12, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Singh, N. Morinda citrifolia (Noni) alter oxidative stress marker and antioxidant activity in cervical cancer cell Lines. Asian Pac. J. Cancer Prev. 2013, 14, 4603–4606. [Google Scholar] [CrossRef]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.M.; Dayem, A.A.; Cho, S.G. Correlation between oxidative stress, nutrition, and cancer initiation. Int. J. Mol. Sci. 2017, 18, E1544. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef]
- Davis, J.M.; Murphy, E.A. Effects of the dietary flavonoid quercetin upon performance and health. Curr. Sports Med. Rep. 2009, 8, 206–213. [Google Scholar] [CrossRef]
- Srivastava, S.; Somasagara, R.R.; Hegde, M.; Nishana, M.; Tadi, S.K.; Srivastava, M.; Choudhary, B.; Raghavan, S.C. Quercetin, anatural flavonoid interacts with DNA, arrests cell cycle and causes tumor regression by activating mitochondrial pathway of apoptosis. Sci. Rep. 2016, 6, 24049. [Google Scholar] [CrossRef] [PubMed]
- Tummala, T.; Lou, W.; Gao, A.C.; Nadiminty, N. Quercetin targets hnRNPA1 to overcome enzalutamide resistance in prostate cancer cells. Mol. Cancer Ther. 2017, 16, 2770–2779. [Google Scholar] [CrossRef] [PubMed]
- Lamson, D.W.; Brignall, M.S. Antioxidants and cancer, Part 3: Quercetin. Altern. Med. Rev. 2000, 5, 196–208. [Google Scholar]
- Dhumale, S.S.; Waghela, B.N.; Pathak, C. Quercetin protects necrotic insult and promotes apoptosis by attenuating the expression of RAGE and its ligand HMGB1 in human breast adenocarcinoma cells. IUBMB Life 2015, 67, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Abd-Rabou, A.A.; Ahmed, H.H. PEG decorated PLGA nano-prototype for delivery of bioactive compounds: A novel approach for induction of apoptosis in HepG2 cellline. Adv. Med. Sci. 2017, 62, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Garg, N.K.; Jain, A.; Kesharwani, P.; Jain, A.K.; Nirbhavane, P.; Tyagi, R.K. A synergistic approach ofadapalene-loaded nanostructured lipid carriers, and vitaminco-administration for treating acne. Drug Dev. Ind. Harm. 2016, 42, 897–905. [Google Scholar] [CrossRef]
- Mombeini, M.; Saki, G.; Khorsandi, L.; Bavarsad, N. Effects of silymarin-loaded nanoparticles on HT-29 human colon cancer cells. Medicina 2018, 54, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, L.; Chen, T.; Guo, W.; Bao, X.; Wang, D.; Ren, B.; Wang, H.; Li, Y.; Wang, Y. Anticancer effects of resveratrol-loaded solid lipid nanoparticles on human breast cancer cells. Molecules 2017, 22, 1814. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, S.; Mukherjee, S.; Das, S.; Bhat, F.A.; Raja Singh, P.; Patra, C.R.; Arunakaran, J. Gold nanoparticles-conjugated quercetin induces apoptosis via inhibition of EGFR/PI3K/Akt-mediated pathway in breast cancer cell lines (MCF-7 and MDA-MB-231). Cell Biochem. Funct. 2017, 35, 217–231. [Google Scholar] [CrossRef]
- Govindaraju, S.; Rengaraj, A.; Arivazhagan, R.; Huh, Y.S.; Yun, K. Curcumin-conjugated gold clusters for bioimaging and anticancer applications. Bioconjug. Chem. 2018, 29, 363–370. [Google Scholar] [CrossRef]
- Nazar, M.; Talebi, A.R.; Hosseini Sharifabad, M.; Abbasi, A.; Khoradmehr, A.; Danafar, A.H. Acute and chronic effects of gold nanoparticles on sperm parameters and chromatin structure in mice. Int. J. Reprod. Biomed. 2016, 14, 637–642. [Google Scholar]
- Hwang, J.H.; Kim, S.J.; Kim, Y.H.; Noh, J.R.; Gang, G.T.; Chung, B.H.; Song, N.W.; Lee, C.H. Susceptibility to gold nanoparticle-induced hepatotoxicity is enhanced in a mouse model of nonalcoholic steatohepatitis. Toxicology 2012, 294, 27–35. [Google Scholar] [CrossRef]
- Isoda, K.; Daibo, T.; Yushina, K.; Yoshioka, Y.; Tsutsumi, Y.; Akimoto, Y.; Kawakami, H.; Taira, Y.; Taira, I.; Yanoshita, R.; et al. Hepatotoxicity, nephrotoxicity, and drug/chemical interaction toxicity of platinum nanoparticles in mice. Pharmazie 2017, 72, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Abbasalipourkabir, R.; Salehzadeh, A.; Abdullah, R. Antitumor activity of tamoxifen loaded solid lipid nanoparticles on induced mammary tumor gland in sprague-dawley rats. Afr. J. Biotechnol. 2010, 9, 7337–7345. [Google Scholar]
- Badawi, N.M.; Teaima, M.H.; El-Say, K.M.; Attia, D.A.; El-Nabarawi, M.A.; Elmazar, MM. Pomegranate extract-loaded solid lipid nanoparticles: Design, optimization, and in vitro cytotoxicity study. Int. J. Nanomed. 2018, 13, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Stella, B.; Peira, E.; Dianzani, C.; Gallarate, M.; Battaglia, L.; Gigliotti, C.L.; Boggio, E.; Dianzani, U.; Dosio, F. Development and characterization of solid lipid nanoparticles loaded with a highly active doxorubicin derivative. Nanomaterials 2018, 8, 2. [Google Scholar] [CrossRef]
- Baek, J.S.; Na, Y.G.; Cho, CW. Sustained cytotoxicity of Wogonin on breast cancer cells by encapsulation in Solid Lipid Nanoparticles. Nanomaterials 2018, 8, 159. [Google Scholar] [CrossRef]
- Wang, W.; Chen, T.; Xu, H.; Ren, B.; Cheng, X.; Qi, R.; Liu, H.; Wang, Y.; Yan, L.; Chen, S.; et al. Curcumin-loaded solid lipid nanoparticles enhanced anticancer efficiency in breast cancer. Molecules 2018, 23, E1578. [Google Scholar] [CrossRef]
- Sun, J.; Bi, C.; Chan, H.M.; Sun, S.; Zhang, Q.; Zheng, Y. Curcumin-loaded solid lipid nanoparticles have prolonged in vitro antitumour activity, cellular uptake and improved in vivo bioavailability. Colloids Surf. B Biointerfaces 2013, 111, 367–375. [Google Scholar] [CrossRef]
- Wang, L.; Li, H.; Wang, S.; Liu, R.; Wu, Z.; Wang, C.; Wang, Y.; Chen, M. Enhancing the antitumor activity of berberine hydrochloride by solid lipid nanoparticle encapsulation. AAPS PharmSciTech 2014, 15, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, S.; Chen, R.; Wang, Y.; Li, H.; Wang, Y.; Chen, M. Oridonin loaded solid lipid nanoparticles enhanced antitumor activity in MCF-7 cells. J. Nanomater. 2014, 2014, 903646. [Google Scholar] [CrossRef]
- Masiiwa, W.L.; Gadaga, L.L. Intestinal permeability of artesunate-loaded solid lipid nanoparticles using the everted gut method. J. Drug Deliv. 2018, 2018, 3021738. [Google Scholar] [CrossRef]
- Vijayakumar, A.; Baskaran, R.; Jang, Y.S.; Oh, S.H.; Yoo, B.K. Quercetin-loaded solid lipid nanoparticle dispersion with improved physicochemical properties and cellular uptake. AAPS PharmSciTech 2017, 18, 875–883. [Google Scholar] [CrossRef]
- Abbasalipourkabir, R.; Salehzadeh, A.; Abdullah, R. Delivering tamoxifen within solid lipid nanoparticles. Pharm. Technol. 2011, 35, 74–79. [Google Scholar]
- Park, U.H.; Jeong, J.C.; Jang, J.S.; Sung, M.R.; Youn, H.; Lee, S.J.; Kim, E.J.; Um, S.J. Negative regulation of adipogenesis by kaempferol, a component of Rhizoma Polygonati falcatum in 3T3-L1 cells. Biol. Pharm. Bull. 2012, 35, 1525–1533. [Google Scholar] [CrossRef]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Prot. 2016, 1, 2315–2319. [Google Scholar] [CrossRef]
- Lin, C.W.; Hou, W.C.; Shen, S.C.; Juan, S.H.; Ko, C.H.; Wang, L.M.; Chen, Y.C. Quercetin inhibition of tumor invasion via suppressing PKCd/ERK/AP-1-dependent matrix metalloproteinase-9 activation in breast carcinoma cells. Carcinogenesis 2008, 29, 1807–1815. [Google Scholar] [CrossRef]
- Wu, Q.; Kroon, P.A.; Shao, H.; Needs, P.W.; Yang, X. Differential Effects of Quercetin and Two of Its Derivatives, Isorhamnetin and Isorhamnetin-3-glucuronide, in Inhibiting the Proliferation of Human Breast-Cancer MCF-7 Cells. J. Agric. Food. Chem. 2018, 66, 7181–7189. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, N.; Wang, J.; Liu, Z.; Wang, X.; Zhang, Q.; Liu, Q.; Gao, L.; Wang, R. Quercetin suppresses breast cancer stem cells (CD44+/CD24-) by inhibiting the PI3K/Akt/mTOR-signaling pathway. Life Sci. 2018, 196, 56–62. [Google Scholar] [CrossRef]
- Zhuang, Y.G.; Xu, B.; Huang, F.; Wu, J.J.; Chen, S. Solid lipid nanoparticles of anticancer drugs against MCF-7 cell line and a murine breast cancer model. Pharmazie 2012, 67, 925–929. [Google Scholar] [CrossRef]
- Abbasalipourkabir, R.; Salehzadeh, A.; Abdullah, R. Tamoxifen-loaded solid lipid nanoparticles-induced apoptosis in breast cancer cell lines. J. Exp. Nanosci. 2016, 11, 161–174. [Google Scholar] [CrossRef]
- Sun, M.; Nie, S.; Pan, X.; Zhang, R.; Fan, Z.; Wang, S. Quercetin-nanostructured lipid carriers: Characteristics and anti-breast cancer activities in vitro. Colloids Surf. B Biointerfaces 2014, 113, 15–24. [Google Scholar] [CrossRef]
- Fulda, S. Tumor resistance to apoptosis. Int. J. Cancer 2009, 124, 511–515. [Google Scholar] [CrossRef]
- Naumov, G.N.; Akslen, L.A.; Folkman, J. Role of angiogenesis in human tumor dormancy: Animal models of the angiogenic switch. Cell Cycle 2006, 5, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, K.; Velmurugan, B.; Gu, M.; Singh, R.P.; Agarwal, R. Inhibitory effect of silibinin against azoxymethane-induced colon tumorigenesis in A/J mice. Clin. Cancer Res. 2010, 16, 4595–4606. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.K.; Thanki, K.; Jain, S. Novel self-nanoemulsifying formulation of quercetin: Implications of pro-oxidant activity on the anticancer efficacy. Nanomedicine 2014, 10, 959–969. [Google Scholar] [CrossRef]
- Eisenberg-Lerner, A.; Bialik, S.; Simon, H.U.; Kimchi, A. Life and death partners: Apoptosis, autophagy and the cross-talk between them. Cell Death Diff. 2009, 16, 966–975. [Google Scholar] [CrossRef]
- Lee, D.H.; Szczepanski, M.; Lee, Y.J. Role of Bax in quercetin-induced apoptosis in human prostate cancer cells. Biochem. Pharmacol. 2008, 75, 2345–2355. [Google Scholar] [CrossRef]
- Duo, J.; Ying, G.G.; Wang, G.W.; Zhang, L. Quercetin inhibits human breast cancer cell proliferation and induces apoptosis via Bcl-2 and Bax regulation. Mol. Med. Rep. 2012, 5, 1453–1456. [Google Scholar] [CrossRef]
- Tserga, A.; Chatziandreou, I.; Michalopoulos, N.V.; Patsouris, E.; Saetta, A.A. Mutation of genes of the PI3K/AKT pathway in breast cancer supports their potential importance as biomarker for breast cancer aggressiveness. Virchows Arch. 2016, 469, 35–43. [Google Scholar] [CrossRef]
- Ren, X.; Zhao, B.; Chang, H.; Xiao, M.; Wu, Y.; Liu, Y. Paclitaxel suppresses proliferation and induces apoptosis through regulation of ROS and the AKT/MAPK signaling pathway in canine mammary gland tumor cells. Mol. Med. Rep. 2018, 17, 8289–8299. [Google Scholar] [CrossRef]
- Hasanzadeh, D.; Mahdavi, M.; Dehghan, G.; Nozad Charoudeh, H. Farnesiferol C induces cell cycle arrest and apoptosis mediated by oxidative stress in MCF-7 cell line. Toxicol. Rep. 2017, 4, 420–426. [Google Scholar] [CrossRef]
- Yao, Q.; Lin, M.; Wang, Y.; Lai, Y.; Hu, J.; Fu, T.; Wang, L.; Lin, S.; Chen, L.; Guo, Y. Curcumin induces the apoptosis of A549 cells via oxidative stress and MAPK signaling pathways. Int. J. Mol. Med. 2015, 36, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Haghiac, M.; Walle, T. Quercetin induces necrosis and apoptosis in SCC-9 oral cancer cells. Nutr. Cancer 2005, 53, 220–231. [Google Scholar] [CrossRef]
- Whelan, R.S.; Konstantinidis, K.; Wei, A.C.; Chen, Y.; Reyna, D.E.; Jha, S.; Yang, Y.; Calvert, J.W.; Lindsten, T.; Thompson, C.B.; et al. Bax regulates primary necrosis through mitochondrial dynamics. Proc. Natl. Acad. Sci. USA 2012, 109, 6566–6571. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wu, A.; Zhu, H.; Fang, H.; Xu, L.; Ye, J.; Shen, J. Melatonin is involved in the apoptosis and necrosis of pancreatic cancer cell line SW-1990 via modulating of Bcl-2/Bax balance. Biomed. Pharmacother. 2013, 67, 133–139. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Delarami Far, A.; Yari, A.; Heravi, R.E.; Tabrizian, K.; Taghdisi, S.M.; Sadegh, S.E.; Tsarouhas, K.; Kouretas, D.; Tzanakakis, G.; et al. Anticancer and apoptosis-inducing effects of quercetin in vitro and in vivo. Oncol. Rep. 2017, 38, 819–828. [Google Scholar] [CrossRef]
- Khoei, S.; Azarian, M.; Rafipour, M. The effect of quercetin and hyperthermia on spheroid model of DU145 prostate carcinoma cell line. J. Paramed. Sci. 2013, 4, 82–89. [Google Scholar]
- Hakimuddin, F.; Paliyath, G.; Meckling, K. Treatment of Mcf-7 breast cancer cells with a red grape wine polyphenol fraction results in disruption of calcium homeostasis and cell cycle arrest causing selective cytotoxicity. J. Agric. Food Chem. 2006, 54, 7912–7923. [Google Scholar] [CrossRef]
- Brusselmans, K.; De Schrijver, E.; Heyns, W.; Verhoeven, G.; Swinnen, J.V. Epigallocatechin-3-gallate is a potent natural inhibitor of fatty acid synthase in intact cells and selectively induces apoptosis in prostate cancer cells. Int. J. Cancer 2003, 106, 856–862. [Google Scholar] [CrossRef]
- Chen, Z.P.; Schell, J.B.; Ho, C.T.; Chen, K.Y. Green tea epigallocatechin gallate shows a pronounced growth inhibitory effect on cancerous cells but not on their normal counterparts. Cancer Lett. 1998, 129, 173–179. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Drug-Lipid Ratio | QT (mg) | Compritol (mg) | Tween 80 (mL) | EE (%) | LD (%) |
|---|---|---|---|---|---|---|
| QT-SLN1 | 1:1 | 50 | 50 | 6 | 67.6 ± 4.3 | 19.7 ± 2.6 |
| Blank-SLN1 | - | - | 50 | 6 | - | - |
| QT-SLN2 | 1:3 | 50 | 150 | 6 | 78.4 ± 4.8 | 21.5 ± 2.9 |
| Blank-SLN2 | - | - | 150 | 6 | - | - |
| QT-SLN3 | 1:5 | 50 | 250 | 6 | 89.3 ± 5.8 | 27.3 ± 3.9 |
| Blank-SLN3 | - | - | 250 | 6 | - | - |
| QT-SLN4 | 1:10 | 50 | 500 | 6 | 97.6 ± 2.3 | 28.3 ± 3.8 |
| Blank-SLN4 | - | - | 500 | 6 | - | - |
| QT-SLN5 | 1:15 | 50 | 750 | 6 | 98.9 ± 1.1 | 28.6 ± 4.1 |
| Blank-SLN5 | - | - | 750 | 6 | - | - |
| Treatments | Cells | 12 h | 24 h | 48 h | 72 h |
|---|---|---|---|---|---|
| QT | MCF-7 MCF-10A | 86.7 ± 7.5 218.3 ± 16.9 | 73.8 ± 7.1 201.2 ± 16.3 | 41.5 ± 6.1 **† 178.4 ± 14.6 * | 40.2 ± 5.8 **† 174.4 ± 15.1 * |
| QT-SLN | MCF-7 MCF-10A | 48.8 ± 4.3 198.5 ± 13.2 | 36.7 ± 3.5 * 182 ± 11.1 | 25.01 ± 2.4 **† 176.8 ± 12.5 | 24.7 ± 2.7 **† 171.3 ± 10.8 |
| Blank-SLN | MCF-7 MCF-10A | 293.2 ± 18.4 288.3 ± 17.8 | 289.7 ± 18.1 283.4 ± 19.1 | 284.5 ± 16.4 282.7 ± 16.9 | 281.4 ± 16.8 278.6 ± 18.2 |
| Formulation | Drug-Lipid Ratio | Particle Size | PDI | Zeta Potential (mV) |
|---|---|---|---|---|
| QT-SLN1 | 1:1 | 45.5 ± 3.5 | 0.112 ± 0.01 | −1.8 ± 0.26 |
| Blank-SLN1 | - | 45.1 ± 3.2 | 0.114 ± 0.02 | −1.1 ± 0.35 |
| QT-SLN2 | 1:3 | 48.4 ± 3.9 | 0.118 ± 0.07 | −5.5 ± 1.12 |
| Blank-SLN2 | - | 47.9 ± 4.1 | 0.123 ± 0.06 | −8.9 ± 1.33 |
| QT-SLN3 | 1:5 | 58.3 ± 4.8 | 0.135 ± 0.07 | −12.6 ± 2.32 |
| Blank-SLN3 | - | 56.1 ± 4.6 | 0.127 ± 0.06 | −13.9 ± 1.87 |
| QT-SLN4 | 1:10 | 85.5 ± 8.5 | 0.152 ± 0.04 | −22.5 ± 0.6 |
| Blank-SLN4 | - | 84.7 ± 8.1 | 0.161 ± 0.05 | −23.6 ± 0.5 |
| QT-SLN5 | 1:15 | 99.6 ± 9.1 | 0.342 ± 0.04 | −18.9 ± 3.13 |
| Blank-SLN5 | - | 98.8 ± 8.7 | 0.316 ± 0.11 | −20.3 ± 2.58 |
| Groups | Control | Blank SLN | QT | QT-SLNs |
|---|---|---|---|---|
| Viability (%) | 100.0 ± 0.02 | 100.1 ± 0.09 | 101.2 ± 1.8 | 104.3 ± 2.1 |
| Colony numbers (%) | 1.42 ± 0.14 | 1.29 ± 0.22 | 1.38 ± 0.18 | 1.43 ± 0.12 |
| Early apoptosis (%) | 3.12 ± 0.56 | 2.96 ± 0.32 | 3.14 ± 0.62 | 2.87 ± 0.35 |
| Late apoptosis (%) | 2.57 ± 0.25 | 2.61 ± 0.25 | 2.26 ± 0.23 | 2.09 ± 0.15 |
| Necrosis (%) | 1.27 ± 0.08 | 1.01 ± 0.13 | 1.23 ± 0.05 | 1.26 ± 0.04 |
| Groups | Control | Blank SLN | QT | QT-SLNs |
|---|---|---|---|---|
| MDA (nmol/mg protein) | 0.042 ± 0.00 | 0.043 ± 0.00 | 0.041 ± 0.00 | 0.036 ± 0.00 |
| CAT (U/mg protein) | 130.3 ± 0.00 | 129.4 ± 6.51 | 131.7 ± 8.34 | 135.5 ± 9.13 |
| SOD (U/mg protein) | 13.71 ± 3.21 | 13.63 ± 3.08 | 13.86 ± 3.68 | 13.95 ± 4.05 |
| ROS (% of control) | 100 ± 0.00 | 99.72 ± 0.21 | 99.84 ± 0.31 | 99.47 ± 0.29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niazvand, F.; Orazizadeh, M.; Khorsandi, L.; Abbaspour, M.; Mansouri, E.; Khodadadi, A. Effects of Quercetin-Loaded Nanoparticles on MCF-7 Human Breast Cancer Cells. Medicina 2019, 55, 114. https://doi.org/10.3390/medicina55040114
Niazvand F, Orazizadeh M, Khorsandi L, Abbaspour M, Mansouri E, Khodadadi A. Effects of Quercetin-Loaded Nanoparticles on MCF-7 Human Breast Cancer Cells. Medicina. 2019; 55(4):114. https://doi.org/10.3390/medicina55040114
Chicago/Turabian StyleNiazvand, Firoozeh, Mahmoud Orazizadeh, Layasadat Khorsandi, Mohammadreza Abbaspour, Esrafil Mansouri, and Ali Khodadadi. 2019. "Effects of Quercetin-Loaded Nanoparticles on MCF-7 Human Breast Cancer Cells" Medicina 55, no. 4: 114. https://doi.org/10.3390/medicina55040114
APA StyleNiazvand, F., Orazizadeh, M., Khorsandi, L., Abbaspour, M., Mansouri, E., & Khodadadi, A. (2019). Effects of Quercetin-Loaded Nanoparticles on MCF-7 Human Breast Cancer Cells. Medicina, 55(4), 114. https://doi.org/10.3390/medicina55040114